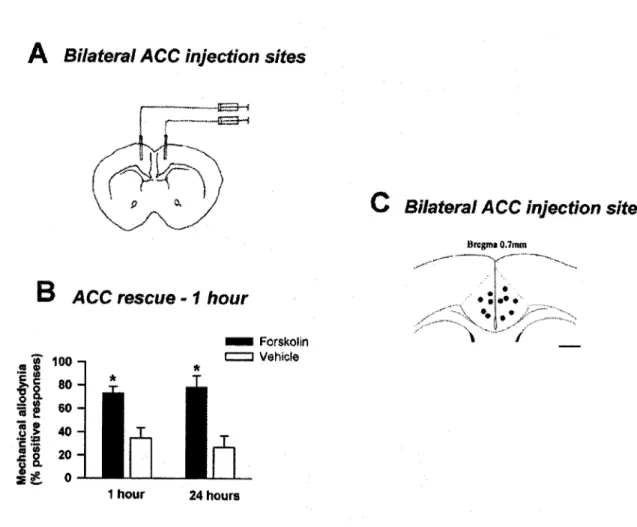
In a model of chronic muscle pain, ACI”; AC8” mice showed significantly reduced mechanical allodynia compared to wild-type mice. Excitatory synaptic transmission in layer I/II ACC neurons was enhanced on day 7 after CPN ligation. After CPN ligation in ACI mice, there was no increase in excitatory synaptic transmission in the anterior cingulate cortex (ACC), a brain region associated with the affective characteristics of pain.
These results suggest that AC1 in neuropathic pain is likely responsible for enhancing synaptic transmission in the ACC. I also thank other laboratory members, with whom I worked during my stay in the laboratory, for influencing me with their individual ways of approaching scientific problems. Fortnightly Zhuo lab journal clubs, intense neuroscience research atmosphere in the Physiology department and distinguished lectures under the program.
Chapter 4 Discussion
AC ACI“
ACI”
ACI”; ACI”
ACC AMPA
Chapter 1 Introduction
- Physiological versus pathological pain
- Animal models to study deep tissue and neuropathic pain
- Synaptic changes and pathological pain
Microinjection of opioid receptor agonists into the ACC produces potent anti-nociceptive effects (Lee et al. 1999a). However, tonic descending inhibition appears to be ineffective in the chronic phase of inflammation (reviewed in (Schaible et al., 2006)). Up-regulation of forebrain NUDA NR2B receptors was shown to contribute to behavioral sensitization following inflammation (Wu et al., 2005a).
The area of referred pain occurring in the same spinal segment may become tender or hyperalgesic (Procacci et al., 1986). Upregulation of the mRNAs for both AMPA and NMDA receptors was shown in diabetic neuropathy (Tomiyama et al., 2005). Nine trans-membrane adenylyl cyclases (tmACs) encoded by nine distinct genes called type 1 to type 9 were first discovered, followed by the discovery of soluble AC (SAC) (Buck et al., 1999).
An increase in phosphorylated CREB in the spinal cord occurs after repeated carrageenan-induced paw inflammation (Messersmith et al., 1998). Electrophysiological recordings from the ACC in humans have shown that some neurons respond to noxious stimuli (Hutchison et al., 1999).
Chapter 2 Materials and methods
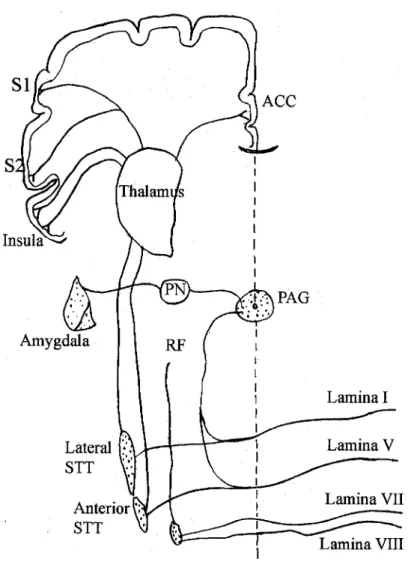
The whole-cell patch-clamp recording technique was used by stimulating projections from neurons in the mediodorsal thalamic nucleus that pass through the lower laminar layers to terminate in layers II/III of the ACC ( Wang and Shyu, 2004 ). A Rotarod apparatus (Med Associates, Georgina, Vermont) was used to assess the animals' ability to stand on a rotating rod that was initially accelerated from zero. The Rotarod test was performed by placing a mouse on a rotating drum and measuring the time each animal was able to maintain its balance while walking on top of the rod.
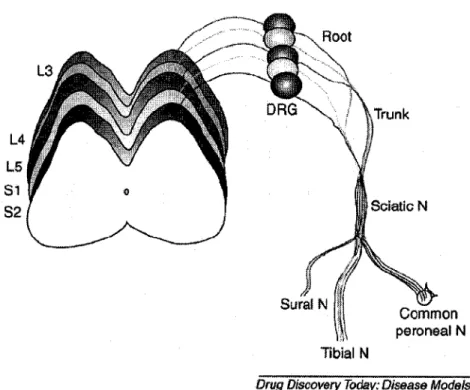
The needle was directed laterally to avoid any bone penetration and the tip was stopped in the middle of the muscle for injection. Nociceptors from the skin of the dorsal aspect of the foot, in addition to the medial and lateral sides, carry sensory stimuli via the common peroneal nerve. Behavioral responses were observed as a limb withdrawal reflex if the innocuous stimulus was perceived as painful by the animal.
Mechanical allodynia was assessed based on the response of the hind paw to the application of von Frey filaments (Steeling, Wood Dale and Illinois) to the point of flexion. The filament was applied over the dorsum of the foot while the animal was resting. Care was taken to apply the von Frey filament to the center of the dorsum of the foot while eliciting the paw withdrawal test as CPN does not supply the lateral aspects of the dorsum of the foot.
The heat source was positioned on the plantar skin of the hind limbs and a beam was fired simultaneously starting a built-in timer. The intensity of the heat source is adjusted with a constant voltage supply to obtain a baseline response time between 3-6 seconds and is kept constant throughout the experiments. After completion of the injection, the needle was held in place for 5 minutes before withdrawal.
Velocity of the microglial chemotaxis against 30 mM ATP puff application to spinal cord slices was performed as described in an early work (Wu et al., 2007).
Chapter 3 Results
- Establishing mouse models of chronic muscle and neuropathic pain
- Role of adenylyl cyclases in muscle pain
Mechanical allodynia in wild-type mice after injections of carrageenan on days 0 and 5 (marked as 5b) into left gastrocnemius muscle. Similar to single gene knockout mice, there was no significant difference between wild-type and double knockout mice, indicating that the absence of AC1, AC8, or a combination of both isoforms of neuronal adenylyl cyclases does not affect baseline motor coordination. There was no significant difference between wild-type and single- and double-knockout mice in the latency to fall, demonstrating that motor function is intact.
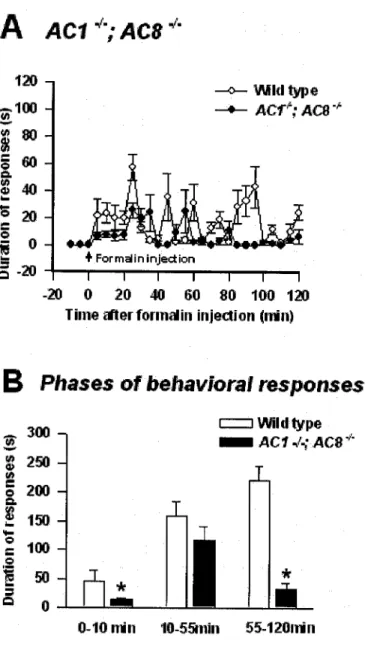
The first phase lasts 10 min after injection and was not significantly altered in AC]” and AC8” mice ( Fig. 7B, D ), indicating that AC1 and AC8 do not significantly contribute to the early phase of acute sensory responses to formalin. . A significant difference was observed between wild-type and ACI" mice at stage 2, (n= 8 mice, P < 0.05) indicating the specific role of this Ca"*/calmodulin-stimulated adenylyl cyclase during the intermediate phase of nociception. At the final stage, behavioral responses from AC]“ mice were significantly reduced compared to wild-type mice.
In contrast, AC8 mice did not show significantly different behavioral changes compared to the wild-type mice. Behavioral nociceptive responses to intramuscular formalin in AC8 mice plotted as early, intermediate, and late phases compared to wild-type mice. Effect of double gene deletion of ACI and AC8 on behavioral responses to intramuscular formalin compared to wild-type mice.
Nociceptive behavioral responses to intramuscular formalin in ACI”;AC8” mice plotted as early, intermediate, and late phases compared to wild-type mice. Although AC8” mice showed no significant reduction in behavioral responses (Fig. 7C,D), ACI”; AC8~ mice showed a significant difference in the late phase (n=8 mice, P<0.05), indicating that the AC1 isoform plays an important role in phase three of the acute persistent muscle pain. Using the chronic inflammatory pain model developed (details are given in Materials and Methods), the role of Ca'*/calmodulin-adenyly] cyclases was tested using ACI” ;AC8” mice.
At an early stage and up to day 8, the behavioral responses between wild type and .

- Developing a neuropathic pain model with least motor defects in
- Neuronal and non-neuronal changes in neuropathic pain
- Role of adenylyl cyclases in neuropathic pain
- Reduced mechanical allodynia in ACI” and ACI” ;AC8” mice after the CPN
- Changes in excitatory synaptic transmission in the ACC after CPN ligation
In addition to the ACC, the dorsal horn of the spinal cord is also involved. Second, most models involve muscle damage in part due to the deep location of the nerve. Mechanical allodynia was tested on postoperative day 1, while leg muscle strength was tested from day 3 onward.
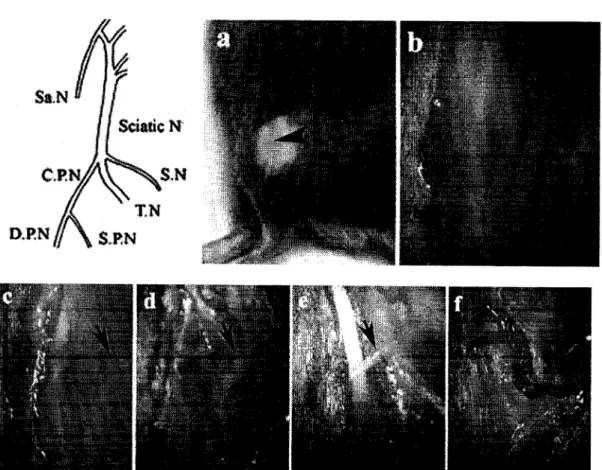
The SPN is traced from the bifurcation of the CPN to the superficial and deep branches (Lemont, 1975). The weakness observed is probably due to inflammatory swelling of the producing nerve. This indicates that nerve ligation did not cause any noticeable muscle weakness in the dorsiflexors (Figure 14).
After nerve injury and nerve ligation, morphological changes in microglia were reported at the level of the spinal cord (Eriksson et al., 1993). Representative sample of chemotaxis of microglial processes towards 30 mM ATP at day 14 after CPN ligation. upper panel) Side contralateral to the CPN connection. Binding of CPN resulted in decreased TMPase activity in the middle third of lamina Ili.
Not the lack of reduction in TMPase activity in the middle third of the inner lamina II on the right side (ligation side) at day 7 after CPN ligation. In addition, an additive effect of AC] and AC8 deficiency was observed on ACI”; AC8” mice during subcutaneous inflammation (Wei et al., 2002a) showing that there was no. Mechanical allodynia on the contralateral side may be due to central sensitization of supraspinal structures and descending modulatory systems.
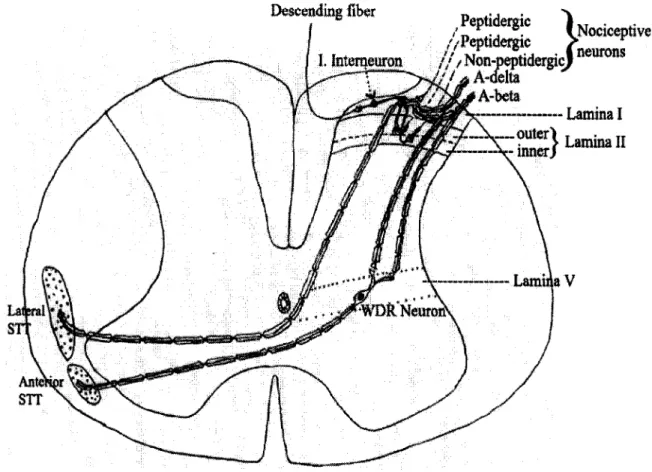
A later study of partial sciatic nerve ligation in mice showed only ipsilateral changes in mechanical allodynia and thermal hyperalgesia (Malmberg and Basbaum, 1998). In previous studies, it was shown that the fibers of the branches of the sciatic nerve have a topographic representation in the dorsal horn (Decosterd and Woolf, 2000). Electrophysiological experiments show that peripheral nerve injury alters excitatory synaptic transmission in lamina II of the dorsal horn (Kohno et al., 2003).
Chapter 5 Conclusion
The effect of Ca"*/calmodulin-stimulated adenylyl cyclases was investigated in the new neuropathic pain model. Microglia] activation in the spinal cord is implicated as one of the mechanisms leading to the chronic nature of neuropathic pain. NMDA EPSCs at glutamatergic synapses in the dorsal horn of the spinal cord of the postnatal rat.
Descending facilitatory modulation of a nociceptive behavioral response by stimulation in the anterior cingulate cortex of adult rats. Paired pulse and frequency facilitation in the CA1 region of the rat hippocampus in vitro. Effect of the {mu} opioid on excitatory and inhibitory synaptic inputs to periaqueductal gray-projecting neurons in the amygdala.
Effects of an acute muscle nerve section on the excitability of dorsal horn neurons in the rat. Thin fiber receptors responding to mechanical, chemical and thermal stimulation in canine skeletal muscle. Altered properties and laminar distribution of neuronal responses to peripheral stimulation in the SmI cortex of the arthritic rat.
Partial sciatic nerve injury in mice as a model of neuropathic pain: behavioral and neuroanatomical correlates. Long-term potentiation and long-term depression of primary afferent neurotransmission in the rat spinal cord. Transsynaptic degeneration in the superficial dorsal horn after sciatic nerve injury: effects of chronic constriction injury, transection, and strychnine.
Upregulation of mRNAs encoding AMPA and NMDA receptor subunits and metabotropic glutamate receptors in the dorsal horn of the spinal cord in a rat model of diabetes mellitus.