METODOLOGÍA SINTÉTICA
APLICADA A LA SÍNTESIS DE
FÁRMACOS
Tema 4
Inflamación: síntesis de
antiinflamatorios
Tema 4. Inflamación: síntesis de antiinflamatorios
4.1. El proceso inflamatorio 1
4.2. Mediadores de la inflamación 2
4.2.1. Metabolitos del ácido araquidónico 2
4.2.2 Aminas vasoactivas: histamina y serotonina 3
4.2.3. Citoquinas 4
4.2.4. Factor Activador de Plaquetas 4
4.2.5. Óxido nítrico 4
4.2.6. Especies de oxígeno reactivas 5
4.2.7. Constituyentes de los lisosomas de los leucocitos 6
4.2.8. Neuropéptidos 6
4.2.9. Mediadores derivados de proteínas plasmáticas 6
4.3. Efectos generales de la inflamación 7
4.3.1. Detención de la respuesta inflamatoria aguda 8
4.3.2. Inflamación crónica 8
4.4. Fármacos antiinflamatorios 9
4.4.1. Ciclooxigenasas 15
4.4.2. Modo de acción de los Antiinflamatorios No Esteroideos 23
4.4.2.1. Aspirina 23
4.4.2.2. Ibuprofeno y naproxeno 24
4.4.2.3. Indometacina y flurbiprofeno 25
4.4.2.4. Coxibes: inhibidores selectivos de COX-2 27
4.5. Síntesis de antiinflamatorios 29
4.5.1. Síntesis de ibuprofeno 29
4.5.1.1a. Análisis retrosintético 30
4.5.1.1b. Síntesis 30
4.5.1.1c. Cuestiones 31
4.5.1.2a. Análisis retrosintético de ibuprofeno mediante carbonilación 31
4.5.1.2b. Síntesis de ibuprofeno mediante carbonilación 31
4.5.1.2c. Cuestiones 32
4.5.1.3a. Análisis retrosintético de ibuprofeno mediante cianohidrina 33
4.5.1.3b. Síntesis de ibuprofeno mediante cianohidrina 33
4.5.2. Síntesis de flurbiprofeno 34
4.5.2.a. Análisis retrosintético 35
4.5.2.b. Síntesis 35
4.5.2.c. Cuestiones 36
4.5.3. Síntesis de naproxeno 37
4.5.3.1a. Análisis retrosintético 37
4.5.3.1b. Síntesis naproxeno 37
4.5.3.2a. Análisis retrosintético de naproxeno mediante
acoplamiento organometálico 39
4.5.3.2c. Cuestiones 42
4.5.4. Síntesis de indoprofeno 42
4.5.4.a. Análisis retrosintético 42
4.5.4.b. Síntesis 43
4.5.4.c. Cuestiones 43
4.5.5. Síntesis de indometacina 44
4.5.5.a. Análisis retrosintético 44
4.5.5.b. Síntesis 44
4.5.5.c. Cuestiones 45
4.5.6. Síntesis de sulindac 46
4.5.6.a. Análisis retrosintético 46
4.5.6.b. Síntesis 47
4.5.6.c. Cuestiones 47
4.5.7. Síntesis de etodolaco 48
4.5.7.a. Análisis retrosintético 48
4.5.7.b. Síntesis 48
4.5.7.c. Cuestiones 48
4.5.8. Síntesis de diclofenaco 49
4.5.8.a. Análisis retrosintético 49
4.5.8.b. Síntesis 50
4.5.8.c. Cuestiones 50
4.5.9. Síntesis de ketorolaco 51
4.5.9.1a. Análisis retrosintético 51
4.5.9.1b. Síntesis 52
4.5.9.1c. Cuestiones 53
4.5.9.2. Síntesis enantioselectiva de ketorolaco 53
4.5.10. Síntesis de zomepiraco 54
4.5.10.a. Análisis retrosintético 54
4.5.10.b. Síntesis 55
4.5.10.c. Cuestiones 55
4.5.11. Síntesis de piroxicam 55
4.5.11.a. Análisis retrosintético 55
4.5.6.b. Síntesis 56
4.5.11.c. Cuestiones 56
4.5.12. Síntesis de fenilbutazona 57
4.5.12.a. Análisis retrosintético 57
4.5.12.b. Síntesis 57
4.5.13. Síntesis de ácido flufenámico 58
4.5.13.a. Análisis retrosintético 58
4.5.13.b. Síntesis 58
4.5.13.c. Cuestiones 59
4.5.14.a. Análisis retrosintético 59
4.5.14.b. Síntesis 59
4.5.14.c. Cuestiones 60
4.5.15. Síntesis de oxaprocina 61
4.5.15.a. Análisis retrosintético 61
4.5.15.b. Síntesis 61
4.5.15.c. Cuestiones 61
4.5.16. Síntesis de nimesulida 63
4.5.16.a. Análisis retrosintético 63
4.5.16.b. Síntesis 63
4.5.16.c. Cuestiones 64
4.5.17. Síntesis de tenidap 64
4.5.17.a. Análisis retrosintético 64
4.5.17.b. Síntesis 65
4.5.17.c. Cuestiones 66
4.5.18. Síntesis de benzidamina 66
4.5.18.a. Análisis retrosintético 66
4.5.18.b. Síntesis 67
4.5.19. Síntesis de celecoxib (celebrex) 67
4.5.19.a. Análisis retrosintético 69
4.5.19.b. Síntesis 69
4.5.19.c. Cuestiones 70
4.5.20. Síntesis de etoricoxib 70
4.5.20.a. Análisis retrosintético 70
4.5.20.b. Síntesis 71
4.5.20.c. Cuestiones 72
4.5.21. Síntesis de refocoxib (vioxx) 73
4.5.21.1a. Análisis retrosintético 73
4.5.21.1b. Síntesis 73
4.5.21.1c. Cuestiones 74
4.5.21.a.2. Análisis retrosintético de rofexocib mediante
acoplamiento sp2-sp2 74
4.5.21.2b. Síntesis de rofexocib mediante acoplamiento sp2-sp2 75
4.5.21.2c. Cuestiones 76
4.5.22. Síntesis de lumiracoxib 77
4.5.22.1a. Análisis retrosintético 77
4.5.22.1b. Síntesis 78
4.5.22.1c. Cuestiones 78
4.5.22.2a. Análisis retrosintético de lumiracoxib mediante homologación 78
4.5.22.2b. Síntesis de lumiracoxib mediante homologación 79
4.5.22.2c. Cuestiones 79
4.6.1. Factores desencadenantes de la migraña 80
4.6.2. Factores de riesgo 80
4.6.3. Etapas del proceso migrañoso 81
4.7. Fármacos contra la migraña 81
4.8. Síntesis de triptanos 82
4.8.1. Síntesis de sumatriptan 82
4.8.1.a. Análisis retrosintético 82
4.8.1.b. Síntesis 83
4.8.1.c. Cuestiones 85
4.8.2. Síntesis de eletriptan 85
4.8.2.1a. Análisis retrosintético 85
4.8.2.1b. Síntesis 85
4.8.2.2a. Análisis retrosintético del eletriptan mediante síntesis del
indol de Fischer 87
4.8.2.2b. Síntesis 88
4.8.2.2c. Cuestiones 89
4.8.3. Síntesis de zolmitriptan 90
4.8.3.a. Análisis retrosintético 90
4.8.3.b. Síntesis 90
4.8.3.c. Cuestiones 91
4.8.4. Síntesis de naratriptan 91
4.8.4.a. Análisis retrosintético 91
4.8.4.b. Síntesis 92
4.8.4.c. Cuestiones 92
4.8.5. Síntesis de frovatriptan 92
4.8.5.a. Análisis retrosintético 92
4.1. El proceso inflamatorio
La inflamación es la respuesta del organismo frente a las agresiones del medio y está generada por los agentes inflamatorios. La respuesta inflamatoria ocurre sólo en tejidos conectivos vascularizados y surge con el fin defensivo de aislar y destruir al agente dañino, así como reparar el tejido u órgano dañado. Se considera a la inflamación un mecanismo de inmunidad innata, en contraste con la reacción inmune adaptativa, que es específica para cada tipo de agente infeccioso.
La inflamación se denomina en medicina con el sufijo -itis: faringitis, laringitis, colitis, conjuntivitis, etc.
Los agentes o condicionantes que pueden provocar la respuesta inflamatoria son los siguientes:
a) Las bacterias, virus, parásitos y hongos. Estos agentes infecciosos expresan compuestos patógenos que se unen a los RTT (receptores de tipo Toll, en inglés TLRs de Toll-like
receptors), proteínas transmembrana de tipo I que forman parte del sistema inmunitario
innato del organismo. Los TLRs detectan la presencia de agentes patógenos y desencadenan vías de señalización que estimulan la producción de diferentes mediadores, provocando en última instancia la respuesta inflamatoria (véase la figura 4.1).
Figura 4.1. Representación del modo de acción de los TLR
b) Los agentes que producen necrosis de los tejidos. Cuando estos agentes provocan la necrosis se produce la liberación de metabolitos, como ácido úrico, ADP o incluso ADN, que activan la respuesta inflamatoria. Los agentes capaces de necrosar tejidos son:
.- Agentes físicos, como radiaciones, frío, calor, rayos UV.
.- Agentes químicos, como venenos y toxinas.
.- Traumatismos y cuerpos extraños, que producen inflamación porque dañan los tejidos (necrosis) o aportan microbios.
.- Alteraciones vasculares, como por ejemplo las que producen isquemia.
Síntesis de antiinflamatorios 2
4.2. Mediadores de la inflamación
Los mediadores de la inflamación son pequeñas moléculas como prostaglandinas, leucotrienos y tromboxanos, aminoácidos modificados (histamina, serotonina) y pequeñas proteínas (citoquinas, factores de crecimiento, interleuquinas, etc) que provocan una respuesta en aquellas células que contienen receptores específicos en su membrana plasmática.
4.2.1. Metabolitos del ácido araquidónico
Los derivados del ácido araquidónico, también denominados eicosanoides, sirven como señales intra o extracelulares en la inflamación y en otros procesos biológicos, como en el caso de la hemostasis (conjunto de mecanismos que utiliza el organismo para detener los procesos hemorrágicos).
El ácido araquidónico (AA) es un derivado del ácido linoleico, que se encuentra normalmente esterificado en forma de fosfolípido en las membranas celulares. El AA se libera por acción de las fosfolipasas celulares, a partir de cualquier célula activada (plaquetas), estresada o a punto de morir por necrosis. Una vez liberado, el AA puede metabolizarse en leucotrienos por acción de las lipooxigenasas, y en tromboxanos, prostaciclinas o prostaglandinas por acción de las ciclooxigenasas (figura 4.2).
Fosfolípidos Fosfolípidos
Ácido araquidónico
Leucotrienos HPETE Lipoxinas
Tromboxanos Prostaciclinas Prostaglandinas
Factor activador de plaquetas
Citoquinas proinflamatorias
Fosfolipasa A2
Lipoxigenasa Ciclooxigenasa
Transcripción de fosfolipasa A2
Figura 4.2. Representación de la ruta metabólica de oxidación del ácido araquidónico
En la figura 4.3 se indican las estructuras de los metabolitos resultantes de las vías enzimáticas de oxidación del ácido araquidónico (en esta figura se ha dibujado arbitrariamente la estructura de un representante de cada familia de metabolitos).
COOH
Ácido araquidónico HO
HO OH
Prostaglandinas (prostaglandina F2 ) COOH
LOX
OH OH
COOH
Leucotrienos (leucotrieno LTB4)
HPETE (ácido 5-Hidroperoxieicosatetraenoico)
OOH
COOH
OH OH
OH COOH
Lipoxinas (lipoxina B4)
O
OH COOH
HP
OH
Tromboxanos (tromboxano B2) COX
O
HO OH
HOOC
Prostaciclina PGI2
Figura 4.3. Metabolitos resultantes de la oxidación del ácido araquidónico
4.2.2 Aminas vasoactivas: histamina y serotonina
Síntesis de antiinflamatorios 4
Figura 4.4. Estructuras de la histamina y de la serotonina
4.2.3. Citoquinas
Las citoquinas son pequeñas proteínas (entre 5 y 20 kD) que permiten el intercambio de información entre las células durante el proceso de inflamación, la hematopoyesis1 y las respuestas inmunes. Los factores de crecimiento que utilizan las células epiteliales para estimular su renovación son asimismo citoquinas.
4.2.4. Factor Activador de las Plaquetas
El Factor Activador de Plaquetas (en inglés Platelet Activating Factor, PAF) es un derivado de fosfolípidos mediador de la inflamación. Las principales acciones del PAF se enfocan a la agregación de las plaquetas, la vasoconstricción y broncoconstricción, la adhesión leucocitaria al endotelio, la quimiotaxis, la desgranulación, el estallido oxidativo y la activación de la síntesis de eicosanoides.
Figura 4.5. Estructura del Factor de Agregación de Plaquetas
4.2.5. Óxido nítrico
El óxido nítrico (NO) es un gas soluble producido en algunas neuronas del cerebro, macrófagos y células endoteliales. Actúa de forma paracrina (acción corta y local) sobre las células diana a través de la inducción de GMPc (guanosín monofosfato cíclico),el cual inicia una serie de sucesos intracelulares que acaban provocando la relajación del músculo liso (vasodilatación). La vida media in vivo del NO es muy corta, por lo que sólo actúa sobre las células muy próximas a su lugar de producción.
El NO se sintetiza a partir de L-arginina por la enzima NO-sintasa (NOS). Hay tres tipos de NOS: endotelial (eNOS), neuronal (nNOS) e inducible (iNOS). Las dos primeras son constitutivas, se expresan a niveles bajos y pueden activarse rápidamente aumentando los niveles de calcio intracelular. Sin embargo, la iNOS se activa solamente cuando los macrófagos y otras células son activados por citoquinas (como IFN-γ) o productos microbianos.
1 Proceso de formación, desarrollo y maduración de los elementos formes de la sangre (eritrocitos,
4.2.6. Especies de oxígeno reactivas
Las especies de oxígeno reactivas (en inglés ROS, de Reactive Oxigen Species) pueden liberarse al medio extracelular por los leucocitos después de que hayan sido activados por la presencia de microbios, quimioquinas, complejos inmunes, o después de la fagocitosis. Su producción depende de la activación del sistema NADPH oxidasa. Las principales especies producidas intracelularmente son el anión superóxido (O2-), el peróxido de hidrógeno H2O2 y el radical hidroxilo (·OH).
El anión superóxido puede combinarse con el óxido nítrico para formar especies reactivas del nitrógeno. Estas sustancias atacan todos los materiales biológicos (ADN, proteínas, lípidos, etc), ya sea arrancando electrones, arrancando átomos de hidrógeno o adicionándose sobre los enlaces dobles y reaccionando como potentes oxidantes. La consecuencia de estos procesos oxidantes es la alteración y la posterior pérdida de función de las moléculas afectadas.
La liberación extracelular de radicales libres de oxígeno (RLO) activa quimioquinas, citoquinas y moléculas de adhesión leucocitaria endotelial, amplificando la respuesta inflamatoria. En estas respuestas inflamatorias se provoca:
a) Daño de las células endoteliales, lo que consecuentemente produce un aumento de la permeabilidad vascular.
b) Daño a otras células, como glóbulos rojos o células del parénquima.
c) Inactivación de antiproteasas, como la α1-antitripsina, lo cual provoca un incremento de la destrucción tisular, como ocurre en el enfisema pulmonar.
El efecto negativo de los ERO se deja sentir cuando se produce un desequilibrio debido a una producción exagerada, o a una disminución de los sistemas de defensa, enzimáticos y no enzimáticos. El plasma, los fluidos tisulares y las células poseen enzimas y mecanismos antioxidantes que les permiten protegerse de los radicales libres de oxígeno. Entre estos se encuentran:
a) La enzima superóxido dismutasa, que convierte el anión superóxido en peróxido de hidrógeno.
b) La enzima catalasa, que destoxifica el peróxido de hidrógeno.
c) El glutatión peroxidasa, otro potente destoxificador del H2O2.
d) El ácido úrico, un potente antioxidante presente en el plasma en una concentración mucho mayor que el ascorbato (vitamina C).
e) La proteína ceruloplasmina, la principal transportadora de cobre en el suero.
f) La fracción plasmática libre de hierro de la proteína transferrina.
También existen compuestos de origen alimentario con capacidad antioxidante que intervienen en la neutralización de ERO como:
a) El α-tocoferol (vitamina E), compuesto liposoluble con capacidad de protección de las membranas celulares.
Síntesis de antiinflamatorios 6
c) El ascorbato (vitamina C), compuesto hidrosoluble capaz de regenerar los demás antioxidantes, como el glutatión o el α-tocoferol.
4.2.7. Constituyentes de los lisosomas de los leucocitos
Los neutrófilos y los monocitos contienen gránulos lisosomiales necesarios para la digestión de los materiales fagocitados. Si estos compuestos se vierten al exterior, pueden amplificar la respuesta inflamatoria, ya que tienen un efecto destructor sobre los tejidos (elastasas, colagenasas, proteasas, etc). Para contrarrestar su efecto, existen antiproteasas en el suero, fundamentalmente la α1-antitripsina, que es el principal inhibidor de la elastasa. Otra antiproteasa importante es la α2-macroglobulina.
4.2.8. Neuropéptidos
Los neuropéptidos son sustancias segregadas por los nervios sensoriales y por varios tipos de leucocitos, y juegan un papel en la propagación de la respuesta inflamatoria. Entre ellos se encuentran la sustancia P y la neurocinina A, pertenecientes a la familia de los taquininos producidos en el SNC y periférico. Los pulmones y el tracto gastrointestinal son ricos en fibras que contienen sustancia P. Este compuesto tiene, entre otras funciones, la de la transmisión de las señales dolorosas, la regulación de la presión sanguínea, la estimulación de la secreción de las células endocrinas y el aumento de la permeabilidad vascular.
4.2.9. Mediadores derivados de proteínas plasmáticas
Una gran variedad de fenómenos de la respuesta inflamatoria están mediados por proteínas plasmáticas que pertenecen a tres sistemas interrelacionados:
a) El sistema del complemento:2 las proteínas de este sistema están presentes en el plasma en forma inactiva, y cuando se activan se convierten en enzimas proteolíticas que degradan otras proteínas del complemento, formando una cascada. Los elementos que participan en el proceso inflamatorio se les conoce con el nombre de anafilotoxinas y son el C3a, C5a y en menor medida C4a. Estas enzimas estimulan la liberación de histamina por los mastocitos y, por tanto, producen vasodilatación. El C5a además tiene capacidad quimiotáctica y activa la lipooxigenasa, generando leucotrienos.
b) La coagulación: la inflamación aumenta la producción de algunos factores de la coagulación y convierte al endotelio en trombogénico. En contrapartida, la trombina promueve la inflamación mediante la activación de receptores denominados PAR
(protease-activated receptors), que activan diferentes respuestas como la movilización de selectina-P,
la producción de quimioquinas y citoquinas, la expresión de receptores para integrinas en el endotelio, la inducción de la COX-2 y la producción de prostaglandinas, la producción de NO y PAF, y cambios en la forma endotelial. Como la coagulación y la inflamación pueden iniciar un círculo vicioso de amplificación, la interferencia con la coagulación puede ser una estrategia terapéutica para reducir la inflamación en algunas patologías.
2 El sistema del complemento es uno de los componentes fundamentales de la respuesta inmunitaria
c) Las quininas son péptidos vasoactivos derivados de proteínas plasmáticas, denominadas quininógenos, por la acción de enzimas específicas denominadas calicreínas. El sistema de quininas está íntimamente ligado a la coagulación. Así, la forma activa del factor XII, FXIIa, convierte la precalicreína del plasma en calicreína, que corta una proteína del plasma de alto peso molecular para generar bradiquinina. La bradiquinina aumenta la permeabilidad vascular y causa contracción del músculo liso, dilatación de los vasos y dolor, efectos similares a los de la histamina. Por otro lado, la calicreína tiene efecto quimiotáctico, ya que convierte C5 del sistema del complemento en C5a (también quimiotáctico) y convierte el plasminógeno en plasmina para degradar el coágulo secundario. Los mediadores de la inflamación más importantes del conjunto de los tres sistemas son la bradiquinina el C3a, el C5a y la trombina. En la tabla 4.1 se resumen el papel de los mediadores en la respuesta inflamatoria.
Tabla 4.1
Mediadores Papel en la inflamación
Prostaglandinas Óxido nítrico Histamina
Vasodilatación
Histamina y Serotonina Bradiquinina
Leucotrienos
Factor activador de las plaquetas (PAF) Sustancia P
Aumento de la permeabilidad vascular
TNF, IL-1 Quimioquinas C3a, C5a Leucotrieno B4
Productos bacterianos, como péptidos N- formilmetil
Quimiotaxis, reclutamiento de leucocitos y activación
TNF, IL-1
Prostaglandinas Fiebre
Prostaglandinas
Bradiquinina Dolor
Enzimas lisosomiales de los leucocitos Especies reactivas del oxígeno
Óxido nítrico
Daño tisular
4.3. Efectos generales de la inflamación
Síntesis de antiinflamatorios 8
reactiva). Durante la fase reparadora juegan un papel clave en la activación y movilización de los leucocitos polimorfonucleares (leucocitos PMN) a partir de la médula ósea, así como en la activación de los fibroblastos
4.3.1. Detención de la respuesta inflamatoria aguda
Puesto que este potente proceso de defensa puede producir daños importantes en los tejidos del huésped, es importante mantenerlo bajo un estricto control. En parte, la inflamación desaparece simplemente porque los mediadores se producen en estallidos rápidos (sólo mientras persiste el estímulo), tienen vidas medias cortas, y son degradados tras su liberación. Los neutrófilos también tienen una vida media corta y mueren por apoptosis unas pocas horas después de dejar la sangre. Además, durante el desarrollo del proceso inflamatorio se disparan una serie de señales de STOP que sirven para terminar la reacción de forma activa. El proceso de parada se debe al cambio en el tipo de metabolitos producidos a partir del ácido araquidónico, deteniéndose la producción de leucotrienos proinflamatorios por lipoxinas antiinflamatorias.
Por otro lado, los macrófagos y otras células liberan citoquinas antiinflamatorias, como TGF-β e IL-10, produciendo mediadores lípidicos antiinflamatorios (como resolvinas y protectinas) derivados de ácidos grasos poliinsaturados, generando impulsos nerviosos (descargas colinérgicas) que inhiben la producción de TNF (Tumor Necrosis Factor) por los macrófagos.
4.3.2. Inflamación crónica
Cuando la inflamación se mantiene durante un tiempo prolongado (semanas o meses), se habla de inflamación crónica, en la que coexisten el daño tisular y los intentos de reparación, en diversas combinaciones. La inflamación crónica puede producirse por mantenimiento de la inflamación aguda (si no se resuelve la causa), o bien empezar de manera progresiva y poco evidente, sin las manifestaciones de la inflamación aguda. Este segundo caso es el responsable del daño tisular de algunas de las enfermedades humanas más invalidantes, como la artritis reumatoide, la aterosclerosis, la tuberculosis o la fibrosis pulmonar. Además, es importante en el desarrollo del cáncer y en enfermedades que anteriormente se consideraban exclusivamente degenerativas, como el Alzheimer. Entre las causas de la inflamación crónica se pueden distinguir:
a) Infecciones persistentes producidas por microbios difíciles de erradicar, como micobacterias, ciertos hongos, virus y parásitos.
b) Enfermedades mediadas por el sistema inmune debido a una sobredimensión de la respuesta inmunitaria.
4.4. Fármacos antiinflamatorios
Muchos medicamentos antiinflamatorios deben su modo de acción a la inhibición de la síntesis de prostaglandinas, sustancias de carácter lipídico derivadas del ácido araquidónico (véase la figura 4.6).
Figura 4.6. Estructuras de prostaglandinas de la series E y F (subserie 2)
Las series de las protaglandinas vienen determinadas por el tipo de sustitución que éstas exhiben en el anillo ciclopentánico. La subserie la determina el grado de insaturación de las cadenas laterales. En la figura 4.7 se representan algunas series y subseries de prostaglandinas.
Síntesis de antiinflamatorios 10
Los antiinflamatorios naturales, segregados por el propio organismo, son los derivados de los corticoides, sustancias de origen esteroideo de potente acción antiinflamatoria, pero que causan importantes efectos secundarios.
Los fármacos antiinflamatorios no esteroideos (AINEs) se denominan de esta forma en oposición a los corticoides. Los AINEs disminuyen la inflamación, el dolor y la fiebre inhibiendo la acción de las ciclooxigenasas, enzimas que participan en la biosíntesis de las prostaglandinas. Las funciones de las prostaglandinas se pueden resumir en cinco puntos:
a) Intervienen en la respuesta inflamatoria provocando la vasodilatación, el aumento de la permeabilidad de los tejidos permitiendo el paso de los leucocitos y actuando como antiagregante plaquetario estimulando las terminaciones nerviosas del dolor.
b) Aumentan la secreción de mucus gástrico y disminuyen la secreción de ácido gástrico.
c) Provocan la contracción de la musculatura lisa, lo que es especialmente importante en la zona uterina. De hecho, en el semen humano hay cantidades pequeñas de prostaglandinas que favorecen la contracción del útero y, como consecuencia, la ascensión de los espermatozoides a las trompas uterinas (trompas de falopio). Del mismo modo, durante la menstruación se produce la liberación de protaglandinas para favorecer el desprendimiento del endometrio. Los dolores menstruales son tratados muchas veces con inhibidores de la liberación de prostaglandinas.
d) Intervienen en la regulación de la temperatura corporal.
e) Controlan el descenso de la presión arterial al favorecer la eliminación de sustancias en el riñón.
Los AINEs se pueden clasificar en:
a) Salicilatos y derivados, como la aspirina (ácido acetilsalicílico), el benorilato, la salicilamida, el diflunisal, el clonixinato de lisina o el etersalato.
Figura 4.8. Estructuras de salicilatos AINEs
Figura 4.9. Estructuras de derivados indol-acéticos AINEs
El sulindac inhibe la producción de prostaglandinas, por lo que se indica para el alivio del dolor, fiebre y la inflamación. Aparte de la inhibición de la ciclooxigenasa, el sulindac inhibe el crecimiento de pólipos y lesiones precancerosas del colon, especialmente en pacientes con poliposis adenomatosa familiar.
c) Derivados aril-acéticos, como el etodolaco, que se utiliza para reducir la inflamación y para tratar dolores leves a moderados relacionados con la osteoartritis o la artritis reumatoide. En la figura 4.10 se indican las estructuras de otros derivados aril-acéticos con actividad antiinflamatoria.
Síntesis de antiinflamatorios 12
d) Ácidos enólicos.
d.1) Oxicames, como el piroxicam, que se emplea en tratamiento de los síntomas de la artritis reumatoide, osteoartritis, dolor menstrual primario y dolor posoperatorio.
Figura 4.11. Estructuras de oxicames AINEs
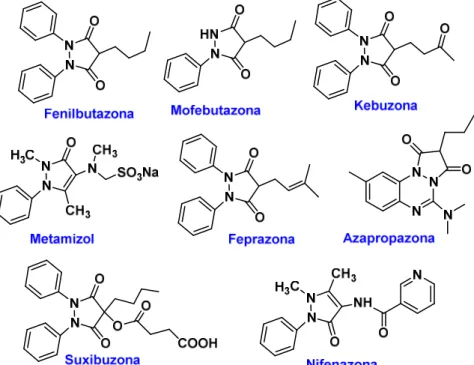
d.2) Pirazolonas, como la fenilbutazona, que se prescribe para el tratamiento del dolor crónico, incluyendo los síntomas de la artritis. Sin embargo, su uso es limitado en humanos por sus efectos adversos severos tales como la supresión de los glóbulos blancos y la anemia aplásica. En la figura 4.12 se indican las estructuras de otras pirazolonas con actividad antiinflamatoria.
Figura 4.12. Estructuras de pirazolonas AINEs
Figura 4.13. Estructuras de derivados arilpropiónicos AINEs
f) Fenematos, como el ácido meclofenámico, analgésico indicado para el tratamiento del dolor leve o moderado, y también indicado como antiinflamatorio y antipirético. En la figura 4.14 se indican las estructuras de otros fenematos con actividad antiinflamatoria.
Figura 4.14. Estructuras de fenematos AINEs
g) Coxibes, como el valdecoxib, que es un inhibidor selectivo de COX-2 y se prescribe para el tratamiento de los dolores mentruales, artritis y osteroartritis.
Síntesis de antiinflamatorios 14
h) Otros, como la nimesulida, el paracetamol, la namubetona, la diacereina, la tolmetina o la oxaprocina. La nimesulida es un antiinflamatorio relativamente COX-2 selectivo, con efectos analgésicos y antipiréticos. Está aprobado como indicación para el tratamiento del dolor agudo, la sintomatología de la osteoartritis y dismenorrea en adolescentes y adultos, por encima de los 12 años de edad. Sin embargo, este fármaco ha sido retirado del mercado debido a su potencial hepatotoxicidad.
N CH3
COOH O CH3 Tolmetina Nimesulida HO H N O CH3 Paracetamol NO2 O NH S H3C
O O MeO O CH3 Nabumetona O H3C
O
O CH3
O O O COOH Diacereina N O COOH Oxaprocina N N O Proquazona N
N O N
Benzidamina
N O N
NH2 O O
Morniflumato N O S O NH2 OH Cl Tenidap O OH HO HO
H2N
OH
Glucosamina
Figura 4.16. Estructuras de otros fármacos AINEs
El paracetamol también se incluye entre los AINEs, a pesar de su poca acción antiinflamatoria.
Tabla 4.2
Salicilatos Derivados
indolacéticos
Derivados
aril-acéticos Ácidos enólicos
Ácido acetilsalicílico Clonixinato de lisina Benorilato Diflunisal Salicilamida Etersalato Salsalato o ácido salicílico Acemetacina Glucametacina Indometacina Proglumetacina Oxametacina Sulindac Aceclofenaco Diclofenaco Etodolaco Fentiazaco Ketorolaco Bufexamaco Lonazolaco Alclofenaco Zomepiraco Difenpiramida Oxicames: Droxicam Meloxicam Piroxicam Tenoxicam Pirazolonas: Fenilbutazona Mofebutazona Oxifenbutazona Clofezona Kebuzona Metamizol (Dipirona) Feprazona Azapropazona Nifenazona Suxibuzona Aminofenazona
Derivados Arilpropiónicos Fenematos Otros
Butibufeno Fenoprofeno Fenbufeno Flurbiprofeno Benoxaprofeno Suprofeno Ibuprofeno Ibuproxam Ketoprofeno Dexketoprofeno Pirprofeno Indoprofeno Naproxeno Oxaprozina Tiaprofeno Dexibuprofeno Fenoprofeno Flunoxaprofeno Alminoprofeno Ácido meclofenámico Ácido mefenámico Ácido flufenámico Ácido tolfenámico Ácido niflúmico Etofenamato (tópico) Paracetamol Tolmetina Oxaprocina Nimesulida Nabumetona Diacereína Proquazona Benzidamina (tópico) Orgoteína Morniflumato Tenidap Glucosamina Glucosaminoglicano, polisulfato Coxibes: Celecoxib Rofecoxib Parecoxib Valdecoxib Etoricoxib 4.4.1. Ciclooxigenasas
Las ciclooxigenasas son los enzimas que catalizan algunas de las reacciones implicadas en la conversión del ácido araquidónico en prostaglandinas. El cuerpo produce dos tipos de ciclooxigenasas, la COX-1 y la COX-2 y ambas tienen funciones distintas.
La COX-1 participa en la señalización celular para mantener la homeostasis3 en el cuerpo, principalmente en el riñón, en las plaquetas y en la mucosa gástrica, donde cumple funciones de protección gastrointestinal.4
Por su parte, la COX-2 participa en la señalización que conlleva a la inflamación y al dolor. Los AINEs clásicos, como la aspirina, actúan inhibiendo principalmente ambos enzimas COX de modo no selectivo. Por ello, el uso de la aspirina puede traer
3 El concepto de homeostasis fue acuñado por el fisiólogo estadounidense Walter Bradford Cannon
(1871-1945) y se define como el conjunto de fenómenos de autorregulación que llevan al mantenimiento de la constancia en las propiedades y la composición del medio interno de un organismo.
4 Las alteraciones que aparecen a nivel gástrico o duodenal se deben a que los AINEs tienen
Síntesis de antiinflamatorios 16
complicaciones adversas, como sangramiento en el estómago debido a la destrucción de la mucosa gástrica.
Las ciclooxigenasas COX-1 y COX-2 se encuentran ancladas a la superficie de la membrana celular y contienen alrededor de 600 aminoácidos. Su centro activo se encuentra en el fondo de un estrecho túnel o canal hidrofóbico. Tres de las hélices alfa del dominio de unión a la membrana están en la entrada de este túnel. Las paredes del túnel están definidas por cuatro hélices alfa formadas por los residuos 106-123, 325-353, 379-384 y 520-535. Al fondo de este canal hidrofóbico se encuentra un importante residuo catalítico: la Tyr-385.
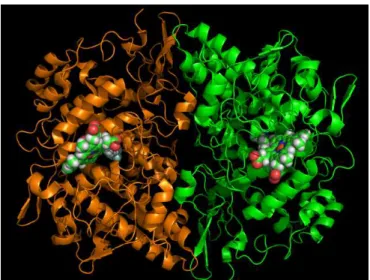
Figura 4.17. Representación de la enzima COX-2 y de los grupos hemo
Se piensa que la inhibición de la COX-2 es la responsable de la acción antiinflamatoria, analgésica y antipirética de los AINEs. Sin embargo, los inhibidores selectivos de COX-2 (Coxibes) no están exentos de riesgos secundarios porque la inhibición de COX-2 rompe el balance entre el efecto antitrombótico y el protrombótico (TxA2), incrementándose la posibilidad de una trombosis cardiovascular. Experimentalmente se ha demostrado que la inhibición selectiva de COX-2 en ratones produce trombogénesis acelerada y presión arterial elevada.
Las ciclooxigenasas tienen acción enzimática dual, puesto que tienen actividad peroxidasa y actividad ciclooxigenasa. El centro activo peroxidasa incluye una parte hemo, con el átomo de Fe(III) coordinado con la His-388 y la His-207. En la figura 4.17 se representa el dímero COX-2 y los grupos hemo que éste contiene.
Las ciclooxigenasas COX-1 y COX-2 son muy similares. Las principales diferencias, por lo que hace a sus centros activos, son el reemplazo de la isoleucina-434 y 523 de COX-1 por la menos voluminosa valina en COX-2, y la sustitución de la arginina-513 de COX-1 por histidina en COX-2. Este cambio genera un bolsillo lateral en el canal de acceso al centro activo de COX-2, en el cual interaccionan los AINEs que inhiben selectivamente a esta enzima.
el lado opuesto al del canal de entrada al centro activo ciclooxigenasa, que es la zona marcada con un asterisco en la figura 4.18. Se puede apreciar que la superposición de las dos enzimas es prácticamente total.
Figura 4.18. Superposición de las enzimas COX-1 y COX-2
En la figura 4.19 se representa la estructura del dímero de COX con indicación del dominio EGF (del inglés Epidermal Growth Factor) y el dominio MBD (del inglés Membrane
Binding Domains), que es el que se ancla en la superficie de la membrana celular. Existe
una sustancial diferencia en la zona MBD entre las dos isoformas de COX.
Figura 4.19. Dímero de COX mostrando los dominios de enlace a membrana (MBD)
Síntesis de antiinflamatorios 18
Figura 4.20. Anclaje de COX a la membrana celular
La unión de la proteína a la membrana se lleva a cabo mediante enlaces no covalentes, de tipo enlaces de hidrógeno y fuerzas electrostáticas. No intervienen fuerzas hidrófobas, ya que las proteínas periféricas sólo interactúan con la zona exterior de la membrana, nunca con su interior hidrófobo. Se unen a la cabeza polar de los lípidos de la bicapa o a determinadas regiones de las proteínas transmembrana. Las enzimas COX son globulares e hidrofílicas, ya que al estar en la periferia de la membrana están expuestas al medio acuoso extracitosólico.
La actividad peroxidasa hemo-dependiente de la COX está implicada en la formación de un radical Tyr-385 (radical tirosinilo), que es necesario para desempeñar la actividad ciclooxigenasa.
Esquema 4.1
En el esquema 4.2 se describen con más detalle los pasos de formación del radical 15-peroxi-PGG2 (compuesto IV del esquema 4.1). El proceso contiene las siguientes etapas:
1) Abstracción por parte del radical tirosinilo del átomo de hidrógeno en C-13 del ácido araquidónico y formación del radical 11-araquidonilo.
2) Oxidación del radical 11-araquidonilo y formación del radical 11-peroxi (intermedio II del esquema 4.1).
Síntesis de antiinflamatorios 20
Esquema 4.2
En la figura 4.21 se indica la estructura del enzima COX y la colocación del ácido araquidónico en el centro activo. El centro activo peroxidasa (centro POD) está colocado en la parte superior de la figura 4.21 y se encuentra expuesto al disolvente, de este forma el peróxido y los substratos exógenos tienen un fácil acceso a este centro enzimático.
El centro activo COX se sitúa en el interior de la enzima y sólo es accesible a lo largo de un surco hidrofóbico con una longitud de 12Å y una anchura de 6Å. El ácido araquidónico accede al centro activo del enzima y adquiere allí una conformación doblada que expone el hidrógeno pro-S del átomo de carbono C-13 a la abstracción por la tirosina-385 (véase el esquema 4.2).
En la figura 4.22 se muestra la colocación del araquidonato en el centro activo de COX-2. El grupo caboxilato forma un par iónico con la Arg-120 y está enlazado mediante puente de hidrógeno con la Tyr-355. El átomo de hidrógeno pro-S del C-13 del araquidonato se coloca en la proximidad del agente oxidante Tyr-385, colocándose el grupo metilo C-20 cerca de la Gly-533. Otros residuos claves son la Ser-530, que es el centro de acetilación de la aspirina, y los aminoácidos Val-523 y la Arg-513, cuyos restos permiten la unión de los grupos sulfonamida y sulfona de los fármacos de tipo diarilheterocíclico.
Figura 4.22. Colocación del araquidonato en el centro activo de COX-2
Las estructuras de los aminoácidos clave en los centros activos de las enzimas COX se indican en la figura 4.23:
Síntesis de antiinflamatorios 22
Es interesante señalar que a pesar de las grandes similitudes entre los enzimas COX-1 y COX-2, el ácido araquidónico se une de forma diferente en el centro activo de cada uno de estos dos enzimas.
En la parte izquierda de la figura 4.24 se representa la enzima COX-1 con el ácido araquidónico en su centro activo (parte inferior central de la estructura en modelo
space-filling en color gris). En la parte superior central de la enzima COX-1 se puede observar la
estructura, en modelo space-filling, del grupo hemo. En el recuadro izquierdo de la figura 4.24 se observa que el ácido araquidónico adopta una conformación alargada en el centro activo de COX-1.
En el recuadro derecho de la figura 4.24 se observa (parte inferior central en gris y en modelo space-filling) que el ácido araquidónico adopta una conformación más doblada en el centro activo de COX-2 (en la parte superior central se observa el grupo hemo).
Figura 4.24. Colocación de araquidonato en COX-1 y COX-2
Las diferentes propiedades y funciones de las enzimas COX-1 y COX-2 se pueden explicar por las pequeñas diferencias entre los centros activos de ambas enzimas. Así, la isoleucina-434 e isoleucina-523 de COX-1 son reemplazadas en COX-2 por el aminoácido valina, menos voluminoso, mientras que la arginina-513 de COX-1 es sustituida por histidina en COX-2. Estas diferencias estructurales entre los centros activos de ambas enzimas se han empleado en el desarrollo de inhibidores selectivos de éstos, en particular de COX-2.
Figura 4.25. Representación esquemática de los centros activos de COX-1 y COX-2
4.4.2. Modo de acción de los Antiinflamatorios No Esteroideos
4.4.2.1. Aspirina
La inhibición de COX-1 por aspirina es 170 veces mayor que la inhibición de COX-2. La aspirina bloquea la ciclooxigenasa mediante un mecanismo completamente distinto al del ibuprofeno. Así, después de la unión de la aspirina a la ciclooxigenasa se produce la transferencia del resto acilo de aquélla a un grupo hidroxilo de un residuo de serina (serina 530 de COX-1 o serina 516 de COX-2). Esta acetilación genera una forma catalíticamente inactiva de la COX-1, mientras que la COX-2 es incapaz de convertir el ácido araquidónico en PGH2. En su lugar se forma el ácido 15R-hidroxieicosatetraenoico (15R-HETE).
Figura 4.26. Interacción COX-1/aspirina mostrando la Ser-530 acetilada
Síntesis de antiinflamatorios 24
En la figura 4.27 se representa esquemáticamente la acción antiinflamatoria de la aspirina mediante acetilación del residuo de serina-530 del centro activo de COX-1.
Figura 4.27. Inactivación del centro activo de COX por acetilación con aspirina
4.4.2.2. Ibuprofeno y naproxeno
El ibuprofeno y el naproxeno ejercen su acción antiinflamatoria ocupando el centro activo de la ciclooxigenasa e impidiendo el acceso al mismo del ácido araquidónico.
Figura 4.20. Estructuras del ibuprofeno y naproxeno
En la figura 4.28 se indica la interacción de ibuprofeno con COX-1.
Estudios sobre COX-2 mutada han demostrado que los grupos naftilo del naproxeno (ácido (S)-6-metoxi-2-metil-2-naftalenacético) son esenciales para la inhibición de la enzima. La mutación de Trp-387 por Phe reduce significativamente la inhibición por el naproxeno. El cambio del átomo de oxígeno del naproxeno por un átomo de azufre incrementa la selectividad del fármaco hacia COX-2.
Figura 4.29. Estructuras del naproxeno y tionaproxeno
En la figura 4.30 se indica la interacción del naproxeno con COX-2:
Figura 4.30. Interacción COX-2/naproxeno
4.4.2.3. Indometacina y flurbiprofeno
La indometacina y el flurbiprofeno provocan la inhibición lenta de COX-1 y COX-2 mediante formación de un puente salino entre el grupo carboxilato del fármaco y el residuo Arg-120, que se encuentra en el túnel del acceso al centro activo.
Síntesis de antiinflamatorios 26
En la figura 4.32 se detalla la interacción de la indometacina (estructura en color verde y amarillo) con el centro activo de la COX.
Figura 4.32. Indometacina en el centro activo de COX
En la figura 4.33 se indica la interacción del flurbiprofeno (en amarillo) con el centro activo de COX-1, con indicación en modelo space-filling de los residuos Ile-434 (color cobre), His-513 (color verde), Phe-518 (color cobre) e Ile-523 (color cobre).
4.4.2.4. Coxibes: inhibidores selectivos de COX-2
En la figura 4.34 se muestran las estructuras de algunos inhibidores selectivos de COX-2. El rofecoxib, comercializado como Vioxx por la compañía Merck & Co, ha sido uno de los fármacos más recetados para combatir la osteoartritis, el dolor agudo y la dismenorrea.
Figura 4.34. Estructuras de inhibidores selectivos de COX-2
En el año 2004 la compañía Merck retiró del mercado el rofecoxib, ante la posible relación entre el aumento de infartos y derrames cerebrales y el consumo de este fármaco en pacientes que tomaban rofecoxib durante periodos prolongados de tiempo y en dosis relativamente elevadas.
A priori la inhibición selectiva de COX-2 es beneficiosa porque COX-2 participa en la biosíntesis de las prostaglandinas malas, responsables del dolor y la inflamación, mientras que COX-1 interviene en la biosíntesis de prostaglandinas buenas, responsables de la protección de la mucosa estomacal. De hecho los fármacos que inhiben la COX-1, como la aspirina, pueden provocar úlceras estomacales.
Las principales diferencias entre las dos isoformas de COX son el intercambio de isoleucina-434, isoleucina-523 y arginina-513 en COX-1 por valina-434, valina-523 e histidina-513 en COX-2.
En la figura 4.25 se han indicado de forma esquemática las diferencias en los centros activos de las enzimas COX debido, fundamentalmente, a la presencia de la isoleucina en la posición 523 de COX-1 y de valina en la posición 523 de COX-2. El aminoácido valina es más pequeño que la isoleucina y el vioxx puede entrar en el bolsillo de COX-2 ocupado por la valina, pero no puede entrar en el bolsillo enzimático de COX-1 porque esta enzima contiene isoleucina, cuyo mayor volumen impide la entrada del fármaco.
Síntesis de antiinflamatorios 28
Figura 4.35. Superposición de vioxx con los centros activos de COX-1 y COX-2
4.5. Síntesis de antiinflamatorios
4.5.1. Síntesis de ibuprofeno
El ibuprofeno es un antiinflamatorio no esteroideo (AINE), utilizado frecuentemente para el alivio sintomático del dolor de cabeza (cefalea), dolor dental (odontalgia), dolor muscular o mialgia, molestias de la menstruación (dismenorrea), dolor neurológico de carácter leve, síndrome febril y dolor tras cirugía (postquirúrgicos). También se usa para tratar cuadros inflamatorios, como los que se presentan en artritis, artritis reumatoide (AR) y artritis gotosa. El ibuprofeno es un inhibidor no selectivo de COX-1 y COX-2. El efecto antiinflamatorio y analgésico está relacionado con la inhibición de COX-2 mientras que la inhibición de COX-1 bloquea la formación de tromboxanos. La inhibición prolongada de COX-1 puede causar toxicidad gástrica ya que la actividad de COX-1 está relacionada con el mantenimiento de la mucosa gástrica.
El ibuprofeno fue desarrollado por la división de investigación de los laboratorios Boots y fue patentado en 1961. Este medicamento forma parte del listado de la Organización Mundial de la Salud de fármacos indispensables.
El ibuprofeno se administra como racemato. El diasteroisómero (-)-R es enzimáticamente isomerizado al (+)-S, pudiendo considerarse como un profármaco de este último. El mecanismo de isomerización implica una conversión inicial del enantiómero (-)-R en el tioéster de la coenzima A (compuesto 4.1 del esquema 4.3). Este intermedio, mediante tautomería ceto-enólica vía enol 4.2, probablemente mediada por un enzima, se epimeriza al intermedio 4.3, que por hidrólisis enzimática se transforma en el (+)-S-ibuprofeno.
Esquema 4.3
Síntesis de antiinflamatorios 30
4.5.1.1a. Análisis retrosintético
En el esquema 4.4 se indica un análisis retrosintético del ibuprofeno que se inicia con la interconversión de la función carboxilo en nitrilo. Esta operación genera el cianocompuesto 4.4 el cual, mediante una nueva operación IGF, se transforma en la oxima 4.5, que a su vez deriva del aldehído 4.6. En este punto del análisis retrosintético se lleva a cabo la escisión del grupo formilo. Esta operación genera el sintón aniónico no natural 4.7 y el sintón catiónico 4.8. El equivalente sintético del sintón aniónico 4.7 se explicará en el parte de síntesis, mientras que para el sintón catiónico 4.8 se empleará como equivalente sintético la aril metil cetona 4.9.
COOH
Ibuprofeno
CN
4.4 4.5
4.7
4.8 4.6
NOH
O H
O H + O
4.9
AGF IGF
IGF
Esquema 4.4
4.5.1.1b. Síntesis
En el esquema 4.5 se describe la síntesis del ibuprofeno según el análisis retrosintético indicado en el esquema anterior. Esta síntesis es la que patentó los laboratorios Boots en 1961.
Esquema 4.5
en medio ácido se provoca una reacción de hidrólisis, con descarboxilación concomitante, que conduce a la obtención del aldehído 4.6. Este compuesto se convierte en la oxima 4.5, la cual se deshidrata al nitrilo 4.6. La hidrólisis del nitrilo proporciona el ibuprofeno.
4.5.1.1c. Cuestiones
1) Proponga un mecanismo la formación del α,β-epoxiéster 4.11 mediante reacción de Darzens.
2) El cloroacetato de etilo actúa en la síntesis del ibuprofeno como equivalente sintético del anión de formilo. Después de la formación del α,β-epoxiéster 4.11, por reacción de la cetona con el cloroaceato de etilo, se obtiene el aldehído 4.6 mediante hidrólisis y descarboxilación del epoxiéster. Explique mecanísticamente la conversión del α,β-epoxiéster 4.11 en el aldehído 4.6.
3) Un método que permite la conversión de oximas en nitrilos se indica a continuación:5
Proponga un mecanismo para la reacción anterior.
4.5.1.2a. Análisis retrosintético mediante carbonilación
En el esquema 4.6 se describe un segundo análisis retrosintético para el ibuprofeno. La primera operación retrosintética desconecta el grupo carbonilo del fármaco y conduce al alcohol bencílico 4.12. El aumento del estado de oxidación de este compuesto proporciona la cetona 4.9, cuya síntesis ya se ha descrito en el esquema 4.4.
Esquema 4.6
4.5.1.2b. Síntesis de ibuprofeno mediante carbonilación
La síntesis del ibuprofeno, según el análisis retrosintético del esquema anterior, se describe en el esquema 4.7 y es la que desarrolló la empresa Hoechst para la producción del fármaco. La secuencia sintética comienza con la preparación de la aril metil cetona 4.9, que en este caso se lleva a cabo mediante reacción SEAr del isobutilbenceno con anhídrido acético en presencia de cantidades catalíticas de HF. La reducción del carbonilo cetónico, por hidrogenación molecular de la cetona 4.9 en presencia del catalizador Ni-Raney, conduce al alcohol bencílico 4.12 que se convierte en ibuprofeno mediante reacción de carbonilación catalizada por paladio en presencia de HI como promotor.
Síntesis de antiinflamatorios 32
Esquema 4.7
4.5.1.2c. Cuestiones
1) La síntesis del ibuprofeno de Hoechst es superior a la de Boots porque únicamente requiere de tres etapas, contra seis que necesita la de Boots, y porque todas las reacciones de la síntesis de Hoechst son catalíticas.
Explique por qué la reacción de acilación del isobutilbenceno 4.10 con anhídrido acético necesita cantidades estequiométricas de AlCl3 (síntesis de Boots, esquema 4.8), mientras que la acilación en presencia de HF (síntesis de Hoechst, esquema 4.8) es catalítica en el ácido protónico.
Esquema 4.8
2) La última etapa en la síntesis del ibuprofeno de Hoechst es la reacción de carbonilación catalizada por paladio del alcohol bencílico 4.12 (véase el esquema 4.6). No se disponen de datos más precisos sobre esta reacción puesto que la síntesis del ibuprofeno de Hoechst ha sido patentada y nunca ha sido publicada. No obstante, en la literatura científica se pueden encontrar métodos de carbonilación de alcoholes bencílicos, como el que han descrito Lin y Yamamoto,6 que consiguen la transformación directa de esta clase de alcoholes en los ácidos carboxílicos homólogos mediante reacción de carbonilación en medio acuoso en presencia de HI y de Pd(PPh3)4.
Proponga un mecanismo que explique la reacción anterior.
4.5.1.3a. Análisis retrosintético de ibuprofeno mediante cianohidrina
En el esquema 4.9 se indica un análisis retrosintético para el ibuprofeno que se inicia con el intercambio del grupo de ácido carboxílico por el grupo nitrilo. Este proceso genera el nitrilo 4.4 que por adición del grupo funcional hidroxilo se transforma en cianohidrina 4.13. La cianohidrina derivará de la aril metil cetona 4.9 que se desconecta, mediante una operación basada en una reacción SEAr, al cloruro de acetilo 4.14 y al isobutilbenceno 4.10.
COOH IGF
Ibuprofeno
CN
AGF CN
OH
IGF
O
O SEAr
Cl +
4.4 4.13
4.14
4.10 4.9
Esquema 4.9
4.5.1.3b. Síntesis de ibuprofeno mediante cianohidrina
La síntesis del ibuprofeno se inicia con la reacción de acilación Friedel-Crafts del isobutilbenceno 4.10 con el cloruro de acetilo 4.14 (esquema 4.10). Esta reacción proporciona la aril metil cetona 4.9 que se convierte en la cianohidrina 4.13 mediante reacción con NaCN. Finalmente, la hidrólisis del grupo nitrilo y la hidrogenolisis concomitante del grupo hidroxilo, por reacción de 4.13 con HI acuoso y fósforo, conduce al ibuprofeno.
Esquema 4.10
Síntesis de antiinflamatorios 34
Esquema 4.11
En el esquema 4.11 se indica la generación de hidrógeno molecular por reacción entre HI y el fósforo. En primer lugar (reacción 1), el ácido yodhídrico se disocia para formar yodo e hidrógeno molecular. Esta reacción se hace irreversible porque el yodo molecular reacciona con el fósforo en medio acuoso para generar el ácido hipofosforoso. La suma de las dos reacciones proporciona la reacción 3, en la cual la oxidación del fósforo por reacción con agua forma ácido hipofosforoso e hidrógeno molecular:
Esquema 4.12
La ecuación global ajustada para la transformación de la cianohidrina 4.13 en ibuprofeno, por reacción con HI y fósforo es la siguiente:
COOH
Ibuprofeno
CN OH
4.13
+ HI + 2 P + 5 H2O + 2 H3PO2 + NH4I
Esquema 4.13
4.5.2. Síntesis de flurbiprofeno
El flurbiprofeno se administra como racemato y se emplea en el tratamiento del dolor y de la artritis. Muy a menudo es uno de los componentes de las pastillas para la tos, como las tabletas Strepsils. La actividad antiinflamatoria del flurbiprofeno se debe al enantiómero
S. El enantiómero R carece de actividad antiinflamatoria y se ha demostrado que no inhibe
ninguna de las dos COX. El enantiómero R se bioconvierte en el S, aunque de manera muy poco eficiente ya que sólo el 1.5% del R se biotransforma en S.
4.5.2.a. Análisis retrosintético
El analisis retrosintético del flurbiprofeno se inicia con la desconexión del sistema bifenílico (esquema 4.14). Esta operación está basada en una reacción de acoplamiento arilo-arilo catalizada por paladio y genera el fragmento nucleofílico 4.17 (Y=metal o metaloide) y el fragmento electrofílico 4.18 (X=halógeno). Una operación de intercambio de grupo funcional convierte el dihaloácido 4.18 en el nitroácido 4.19. La última operación del análisis retrosintético desconecta la parte de acido propiónico del anillo aromático. Esta desconexión se basa en una reacción SNAr y genera el sintón nucleofílico 4.20 y el sintón electrofílico 4.21 (X=halógeno).
Esquema 4.14
4.5.2.b. Síntesis
Para la síntesis del flurbiprofeno se elige como material de partida el 2,4-difluoronitrobenceno 4.21 (esquema 4.15).7 La reacción SNAr entre este compuesto y el anión derivado del metilmalonato de dietilo 4.22, que se emplea como equivalente sintético del sintón aniónico 4.20, proporciona el diéster 4.23. La hidrólisis ácida del diéster, seguida de descarboxilación in situ, conduce al nitroácido 4.19, que se transforma en el anilinoácido 4.24 mediante hidrogenación. El dihaloácido 4.18, necesario para la proyectada reacción de acoplamiento bifenílico, se sintetiza a partir del anilinoácido 4.23 mediante reacción de Sandmeyer vía la correspondiente sal de arildiazonio. El flurbiprofeno se obtiene mediante reacción de acoplamiento de tipo Suzuki entre el dihaloácido 4.18 y el tetrafenoilborato sódico (NaBPh4) en presencia de paladio depositado sobre carbono.8
7 G. Lu, R. Franzen, X. J. Yu, Y. J. Xu. Chin. Chem. Lett. 2006, 17, 461-464.
Síntesis de antiinflamatorios 36
COOH
F O2N
4.18 COOEt
F O2N
F
+
4.21
EtOOC NaOH, DMF
23ºC
COOEt
F O2N
COOEt
HOAc, H2SO4,
H2O, reflujo,
(87% 2 pasos)
H2, Pd/C
23ºC (98%) COOH
F H2N
NaNO2, 40% HBr
CuBr, H2O (83%)
COOH
F Br
NaBPh4, Na2CO3, H2O
0.05 mol% de Pd/C al 5%, reflujo al aire durante 1 h (98%)
COOH F Flurbiprofeno 4.22 4.23 4.24 4.18 Esquema 4.15 4.5.2.c. Cuestiones
1) ¿Por qué la reacción SNAr del anión del metilmalonato de dietilo 4.22 sobre el 2,4-difluoronitrobenceno es regioselectiva? ¿Por qué no se sustituye el átomo de flúor en orto con respecto al grupo nitro?
2) Explique mecanísticamente la conversión de la arilamina 4.24 en el bromoarilo 4.25.
3) La reacción ajustada para el acoplamiento de Suzuki entre el compuesto 4.18 y el NaBPh4 es la siguiente:
Esquema 4.16
La reacción anterior se explica mediante la intervención de cuatro ciclos catalíticos. Con estos datos proponga un mecanismo que explique la formación del flurbiprofeno mediante la reacción de Suzuki.
4.5.3. Síntesis de naproxeno
primaria y los calambres menstruales. El naproxeno también está disponible como sal sódica, que se absorbe más rápidamente que el naproxeno en el tracto gastrointestinal.
4.5.3.1a. Análisis retrosintético
La retrosíntesis del naproxeno se inicia con la interconversión del grupo funcional carboxilo en éster (esquema 4.17). Esta operación genera el naproxenato de alquilo 4.25 que por escisión del grupo metilo, basada en una reacción SN2, conduce al compuesto 4.26. El siguiente paso retrosintético convierte el éster 4.26 en la naftilmetil cetona 4.27. En el esquema 4.16 a esta operación se ha indicado como migración 1,2 de grupo carbonilo. La última desconexión escinde el grupo acetilo y conduce al cloruro de acetilo 4.14 y al 2-metoxinaftaleno 4.28.
Esquema 4.17
4.5.3.1b. Síntesis
La primera síntesis a gran escala del naproxeno se llevó a cabo por la empresa Syntex en 1970 y permitía la producción de 500 kg de este fármaco.9 La síntesis del naproxeno se inicia con la obtención de metoxinaftil metil cetona 4.24 por reacción SEAr entre el 2-metoxinaftaleno 4.28 y el cloruro de acetilo (esquema 4.18). El paso clave de la síntesis es la migración 1,2 del grupo carbonilo cetónico, con aumento concomitante de su estado de oxidación. Esta conversión se consigue mediante reacción de la cetona 4.24 con morfolina y azufre a reflujo. El proceso proporciona la tiomorfolida 4.30 que por hidrólisis ácida se convierte en el ácido carboxílico 4.31. La esterificación de Fischer del ácido 4.31, seguida de enolización con NaH y alquilación con yoduro de metilo, proporciona el naproxenato de metilo racémico (+/-)-4.25. La saponificación de la función éster permite la obtención del naproxeno racémico (+/-)-4.32. La resolución óptica del racemato (+/-)-4.32 se lleva a cabo con el alcaloide (-)-cinconidina 4.33. Así, la reacción de (+/-)-4.32 con el alcaloide genera las correspondientes sales diastereoisoméricas, de las cuales la de estructura 4.34 cristaliza en el seno de la reacción. La separación de esta sal mediante filtración, seguida de tratamiento ácido, proporciona el (S)-naproxeno.
9 I. T. Harrison, B. Lewis, Peter, P. Nelson, W. Rokks, A. Roszkowski, A. Tomolonis, J. H. Fried. J.
Síntesis de antiinflamatorios 38
Esquema 4.18
Esquema 4.19
4.5.3.2a. Análisis retrosintético de naproxeno mediante acoplamiento organometálico
En el esquema 4.20 se indica un segundo análisis retrosintético para el naproxeno. Así, la desconexión de la parte de propionato sobre el éster 4.25 conduce al fragmento nucleofílico 4.41 (Y=metal) y al fragmento electrofílico 4.42 (X=halógeno). El compuesto organometálico 4.41 derivará del compuesto halogenado 4.43 (X=halógeno) que se obtendrá del 2-naftol 4.45 mediante halogenación SEAr seguida de metilación fenólica.
MeO O
OH
MeO O
OR
IGF C-C
MeO
O OR
MeO
X
HO
Naproxeno 4.25 4.41
4.43 4.45
Y X
+
HO
X IGF
C-X
IGF
SEAr
4.42
4.44
Esquema 4.20
4.5.3.2b. Síntesis de naproxeno mediante acoplamiento organometálico
Síntesis de antiinflamatorios 40
de cantidades estequiométricas de hidróxido de aluminio. El tercero es el empleo de disolventes poco apropiados a escala industrial como el nitrobenceno, que se utiliza como disolvente en la reacción de acilación. El cuarto es la utilización de reactivos peligrosos en grandes cantidades, como el yoduro de metilo y el hidruro sódico.
A fin de evitar los inconvenientes asociados a la síntesis del naproxeno, descrita en el esquema 4.18, durante los años 1972-1975, la empresa Syntex aplicó la secuencia sintética que se describe en el esquema 4.21.10 En esta síntesis el compuesto de partida es el -naftol 4.45, que por reacción con bromo molecular proporciona el dibromo-naftol 4.46. La reacción con hidrogenosulfito sódico provoca la eliminación reductiva regioselectiva del bromo en C-1 y proporciona el bromonaftol 4.44, que se convierte en el 2-bromo-6-metoxinaftaleno 4.34 por metilación con cloruro de metilo en medio básico. Este compuesto se transforma en el reactivo de Grignard 4.41, que por transmetalación con ZnCl2 y reacción con 2-bromopropanoato de etilo proporciona el éster racémico (+/-)-4.47. La hidrólisis del éster seguida de resolución con cinconidina permiten la obtención del (S)-naproxeno.
HO HO Br Br2 HO Br 4.46 NaHSO3 O OEt MeO Br 4.41 4.43 4.45 Br 4.42 4.44
CH3Cl, NaOH
(85-90% desde 4.45)
Mg MeO MgBr ZnCl2 MeO O OEt (+/-)-4.47
NaOH, H2O
MeO O
OH
(+/-)-4.32
1. Resolución con (-)-cinconidina 2. HCl ac.
MeO O
OH
Naproxeno
Br
Esquema 4.21
La aplicación industrial de la secuencia sintética del esquema 4.21 tampoco está exenta de inconvenientes. El primero de ellos es el empleo de cantidades estequiométricas de ZnCl2, que se requieren para generar el reactivo organometálico de tipo naftilzinc (no dibujado en el esquema 4.21). La utilización del ZnCl2 genera grandes cantidades de hidróxido de zinc como subproducto. El segundo inconveniente está relacionado con el acoplamiento del reactivo organometálico con el 2-bromopropanoato de etilo, que transcurre
con bajos rendimientos. Además, en esta reacción se forma también el 2-metoxinaftaleno, como subproducto de reducción, y el dímero binaftílico, como consecuencia de una vía secundaria que procede mediante acoplamiento radicalario del reactivo organometálico.
En los años 1976-1993 la producción de naproxeno por Syntex se llevó a cabo mediante la secuencia de reacciones que se indica en el esquema 4.22. La principal novedad de esta secuencia, en relación con la descrita en el esquema 4.21, estriba en el acoplamiento con el reactivo organometálico, que se lleva a cabo sobre la sal cloromagnésica 4.48 derivada del ácido 2-bromopropanoico.
Esquema 4.22
En la síntesis del esquema anterior no necesita ZnCl2 para la reacción de acoplamiento, y por tanto no generan residuos de zinc, y además el subproducto 2-metoxinaftaleno y el dímero binaftílico se forman en mucha menor proporción.
Otra mejora asociada al esquema anterior está relacionada con el paso de resolución óptica del racemato (+/-)-4.32. En esta secuencia la cinconidina se sustituye por una amina quiral preparada mediante aminación reductiva de la glucosa.