RELACIÓN DE PARENTESCO ENTRE CUATRO ESPECIES DEL GÉNERO
ATELOPUS (ANURA: BUFONIDAE) DE ALTA MONTAÑA COLOMBIANA,
BASADA EN CARACTERES OSTEOLÓGICOS
PAOLA NORIEGA NORIEGA
TRABAJO DE GRADO Presentado como Requisito Parcial
para Optar al Título de: BIÓLOGO
DIRECTOR
JULIO MARIO HOYOS HOYOS
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS
CARRERA DE BIOLOGÍA
NOTA DE ADVERTENCIA
Bogotá 16 de Febrero de 2007
Señores
PONTIFICIA UNIVERSIDAD JAVERIANA Cuidad
Estimados Señores:
Yo _Paola Noriega_Noriega_identificada con C.C. No. _52385925, autora del trabajo de grado Titulado,_Relación de parentesco entre cuatro especies del género Atelopus (Anura: Bufonidae) de la alta montaña Colombiana, basada en caracteres osteologicos, presentado como requisito para optar al título de _Biólogo, en el año de 2007; autorizo a la Universidad Javeriana a:
a) Reproducir el trabajo en medio digital o electrónico con el fin de ofrecerlo para la consulta en la Biblioteca General.____Si_____
b) Poner a disposición para la consulta con fines académicos, en la página web de la Facultad, de la Biblioteca General y en redes de información con las cuales tenga convenio la Universidad
Javeriana. __SI____
c) Enviar el trabajo en formato impreso o digital, en caso de que sea seleccionado para participar en concursos de trabajos de grado. ____SI_______
d) Distribuir ejemplares de la obra, para la consulta entre las entidades educativas con las que la facultad tenga convenio de intercambio de información, para que este sea consultado en las bibliotecas y centros de documentación de las respectivas entidades. __Si_
e) Todos los usos, que tengan finalidad académica. ___SI____
Los derechos morales sobre el trabajo son de los autores de conformidad con lo establecido en el artículo 30 de la Ley 23 de 1982 y el artículo 11 de la Decisión Andina 351 de 1993, los cuales son irrenunciables, imprescriptibles, inembargables e inalienables. Atendiendo lo anterior, siempre que se consulte la obra, mediante cita bibliográfica se debe dar crédito al trabajo y a su(s) autor(es). Este documento se firma, sin perjuicio de los acuerdos que el autor(es) pacte con la Unidad Académica referentes al uso de la obra o a los derechos de propiedad industrial que puedan surgir de la actividad académica.
ctÉ
la Noriega N.
AGRADECIMIENTOS
Expreso mi más sincero agradecimiento a todas aquellas personas que han estado
presentes durante toda mi carrera y durante la realización de éste proyecto,
principalmente a toda MI FAMILIA abuelas, padres, tíos (especialmente Connie) y
hermanos, que ha sido de gran apoyo, ayuda e incentivo para seguir adelante frente a
las dificultades que se han presentado en mi vida.
A JULIO MARIO HOYOS por todo lo que me enseñó al dirigir este trabajo, por sus
consejos, comentarios, por su gran paciencia y apoyo que me brindó durante este
camino. A ANDRES ACOSTA que me encaminó en el mundo de las ranas y por ser
de gran ayuda con sus sugerencias y guía especialmente durante la ausencia temporal
de mi director. Y a Sofía, Diana y otros compañeros que con su alegría compartieron
el laboratorio conmigo durante el desarrollo de este proyecto.
A la UNIVERSIDAD JAVERIANA por la formación tanto académica, como ética
que me ha dado durante todos estos años y por darme la oportunidad de desarrollar
este trabajo. AL MUSEO DE LA UNIVERSIDAD DE LA SALLE por su
colaboración para la transparentación y revisión de ejemplares de la colección de
anfibios, en especial al Hermano Roque y a Don Arturito Rodriguez por su ayuda y
amabilidad.
A mis compañeros y amigos, que realmente son pocos los verdaderos y sinceros, por
eso agradezco especialmente a MAO, JAIME, FERCHO, CAROLINA, FRANCY,
RAFA, DIANA y SANDRA, por estar siempre conmigo durante estos años, de una u
otra forma, teniendo siempre una palabra amable o una sonrisa.
Y finalmente a todos los que estuvieron presentes de algún modo dando de sí mismos
algo positivo, mucho o poquito, pero que igualmente me ayudaron, me empajaron y
TABLA DE CONTENIDO
RESUMEN ... 15
ABSTRACT... 16
1 INTRODUCCIÓN ... 17
2 MARCO TEÓRICO Y REVISIÓN DE LITERATURA ... 18
2.1 ANTECEDENTES... 18
2.2 GÉNERO ATELOPUS... 21
2.2.1 Generalidades ... 21
2.2.2 Origen del género... 22
2.2.3 Descripción del género ... 24
Atelopus muisca... 24
Atelopus lozanoi... 25
Atelopus marinkellei... 26
Atelopus mittermeieri... 27
2.2.4 Osteología del género ... 28
2.2.5 Distribución del género... 29
2.2.6 Estado de conservación del género ... 30
2.2.7 Sistemática del género... 32
2.3 ANÁLISIS FILOGENÉTICO... 34
2.3.1 Generalidades del análisis filogenético ... 34
2.3.2 Cladogramas ... 36
2.3.3 Método de comparación por grupo ajeno ... 37
3 FORMULACIÓN DEL PROBLEMA Y JUSTIFICACIÓN ... 38
3.1 FORMULACIÓN DEL PROBLEMA... 38
3.2 PREGUNTA DE INVESTIGACIÓN... 38
3.3 JUSTIFICACIÓN DE LA INVESTIGACIÓN... 39
4 OBJETIVOS ... 40
4.1 OBJETIVO GENERAL... 40
4.2 OBJETIVOS ESPECÍFICOS... 40
5 MATERIALES Y MÉTODOS ... 40
5.1 DISEÑO DE LA INVESTIGACIÓN... 40
5.1.1 Población de estudio y muestra... 41
5.1.2 Variables del estudio ... 42
5.2 MÉTODOS... 43
5.2.1 Recolección de información ... 43
5.2.3 Revisión de caracteres... 46
5.2.4 Selección de caracteres ... 47
5.2.5 Codificación de los caracteres ... 50
5.2.6 Realización de matrices... 51
5.2.7 Generación de cladogramas... 52
5.3 ANÁLISIS DE LA INFORMACIÓN... 53
6 RESULTADOS Y DISCUSIÓN... 54
6.1 RESULTADOS... 54
6.1.1 Revisión de ejemplares... 54
6.1.1.1 Morfometría... 54
6.1.1.2 Osteología... 54
6.1.1.2.1 Cráneo y aparato hioideo ... 55
6.1.1.2.2 Esqueleto axial ... 60
6.1.1.2.3 Esqueleto apendicular... 62
6.1.2 Caracteres Seleccionados ... 66
6.1.3 Generacion de cladogramas... 74
Cladogramas con caracteres de tipo CM-CB-CI. ... 75
Cladogramas con caracteres de tipo CB-CI. ... 84
Cladograma con caracteres de tipo CB-ncofl... 91
Cladogramas con caracteres de tipo P-CI. ... 92
Cladograma con caracteres de tipo P-ncofl... 98
Análisis de cladogramas. ... 99
6.2 DISCUSIÓN... 100
6.2.1 Caracteres utilizados en el análisis filogenético ... 100
6.2.2 Sinapomorfias de las especies del género Atelopus ... 104
6.2.3 Relaciones de parentesco propuestas... 108
6.2.4 Longitud e índice de consistencia... 110
7 CONCLUSIONES ... 112
8 RECOMENDACIONES ... 113
9 REFERENCIAS ... 114
FIGURAS-B ... 121
INDICE DE FIGURAS
Figura-A. 1; A. muisca. (Rueda, et al, 2004) ... 25
Figura-A. 2; A. lozanoi (Rueda, et al, 2004) ... 25
Figura-A. 3; A. marinkellei (Rueda, et al, 2004). ... 26
Figura-A 4; A. mittermeieri (Acosta, et al, 2006)... 27
Figura-A 5; A. mittermeieri (Rueda, et al, 2005)... 27
Figura-A 6; Cladograma con CM–CI, realizado con tres grupos ajenos, iniciando con D. labialis... 81
Figura-A 7; Cladograma con CM–CB–CI, realizado con tres grupos ajenos, iniciando con E. bogotensis. ... 82
Figura-A 8; Cladograma con CM–CB–CI, realizado con tres grupos ajenos, iniciando con C. granulosus. ... 83
Figura-A 9; Cladograma de consenso con CM–CB–CI, realizado con tres grupos ajenos. .. 84
Figura-A 10; Cladograma con CB–CI, realizado con tres grupos ajenos, iniciando con D. labialis... 89
Figura-A. 11; Cladograma con CB–CI, realizado con tres grupos ajenos, iniciando con E. bogotensis... 89
Figura-A. 12; Cladograma con CB–CI, realizado con tres grupos ajenos, iniciando con C. granulosus. ... 90
Figura-A. 13; Cladograma de Consenso con CB–CI, Realizado con tres Grupos Ajenos. ... 90
Figura-A. 14; Cladograma con CB–ncofl, realizado con tres grupos ajenos... 91
Figura-A. 15; Cladograma con P–CI, realizado con tres grupos ajenos, iniciando con D. Labialis... 96
Figura-A. 16; Cladograma con P–CI, realizado con tres grupos ajenos, iniciando con E. bogotensis... 96
Figura-A. 17; Cladograma con P–CI, realizado con tres grupos ajenos, iniciando con C. granulosus. ... 97
Figura-A. 18; Cladograma de Consenso con P–CI, realizado con tres grupos ajenos... 97
Figura-A. 19; Cladograma con P (CB)-ncofl., realizado con tres grupos ajenos. ... 98
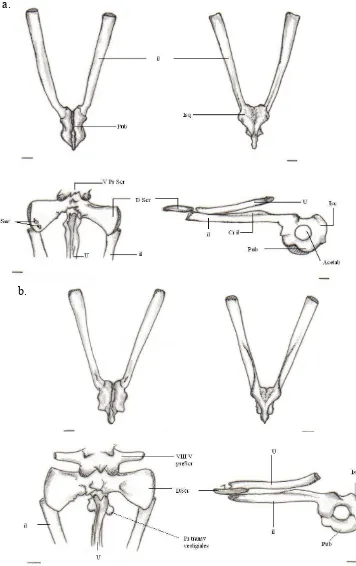
Figura-B. 1; Cráneo, vista dorsal, grupo propio ... 122
Figura-B. 2; Cráneo, vista dorsal, A. mittermeieri... 123
Figura-B. 3; Cráneo, vista dorsal, C. granulosus... 123
Figura-B. 4; Cráneo, vista dorsal, grupo ajeno ... 124
Figura-B. 5; Cráneo, vista ventral, grupo propio... 125
Figura-B. 6; Cráneo, vista ventral, grupo propio, A. mittermeieri... 126
Figura-B. 7; Cráneo, vista ventral, C. granulosus... 126
Figura-B. 8; Cráneo, vista ventral, grupo ajeno... 127
Figura-B. 9; Cráneo, vista lateral, grupo propio ... 128
Figura-B. 10; Cráneo, vista lateral, A. mittermeieri... 129
Figura-B. 11; Cráneo, vista lateral, C. granulosus... 129
Figura-B. 12; Cráneo, vista lateral, grupo ajeno... 130
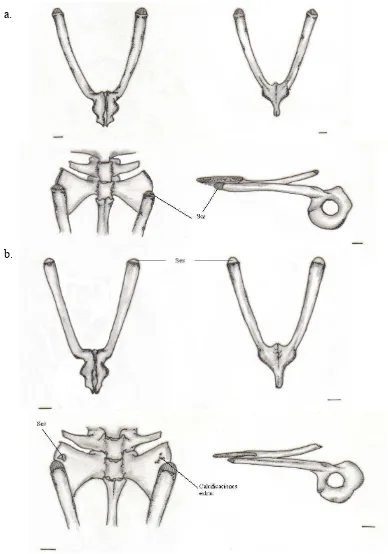
Figura-B. 13; Mandíbula, grupo propio... 131
Figura-B. 14; Mandíbula, grupo propio, A. mittermeieri... 132
Figura-B. 15; Mandíbula, grupo ajeno; C. granulosus... 132
Figura-B. 16; Mandíbula, grupo ajeno. ... 133
Figura-B. 17; Aparato hioideo, grupo propio ... 134
Figura-B. 18; Aparato hioideo, A. mittermeieri... 135
Figura-B. 19; Aparato hioideo, C. granulosus... 135
Figura-B. 20; Aparato hioideo, grupo ajeno ... 136
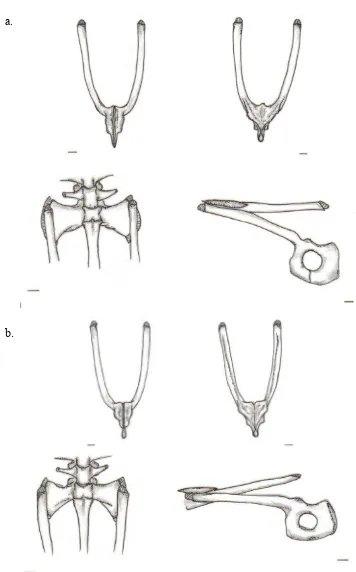
Figura-B. 21; Esqueleto apendicular, cintura pectoral, grupo propio... 137
Figura-B. 22; Esqueleto apendicular, cintura pectoral, A. mittermeieri.... 138
Figura-B. 23; Esqueleto apendicular, cintura pectoral, A. marinkellei... 139
Figura-B. 24; Esqueleto apendicular, cintura pectoral, C. granulosus.... 139
Figura-B. 25; Esqueleto apendicular, cintura pectoral, E. bogotensis... 140
Figura-B. 26; Esqueleto apendicular, cintura pectoral, D. labialis.... 141
Figura-B. 27; Esqueleto axial, columna vertebral, A. muisca... 142
Figura-B. 28; Esqueleto axial, columna vertebral, A. lozanoi... 143
Figura-B. 29; Esqueleto axial, columna vertebral, A. marinkellei.... 144
Figura-B. 30; Esqueleto axial, columna vertebral, A. mittermeieri... 145
Figura-B. 31; Esqueleto axial, columna vertebral, A. mittermeieri... 146
Figura-B. 32; Esqueleto axial, columna vertebral, C. granulosus... 147
Figura-B. 33; Esqueleto axial, columna vertebral, E. bogotensis... 148
Figura-B. 34; Esqueleto axial, columna vertebral, D. labialis... 149
Figura-B. 35; Esqueleto axial, columna vertebral, D. labialis... 150
Figura-B. 36; Esqueleto axial, cintura pélvica, A. muisca.... 151
Figura-B. 37; Esqueleto axial, cintura pélvica, A. lozanoi.... 152
Figura-B. 39; Esqueleto axial, cintura pélvica, A. mittermeieri.... 154
Figura-B. 40; Esqueleto axial, cintura pélvica, A. mittermeieri.... 155
Figura-B. 41; Esqueleto axial, cintura pélvica, C. granulosus.... 156
Figura-B. 42; Esqueleto axial, cintura pélvica, E. bogotensis.... 157
Figura-B. 43; Esqueleto axial, cintura pélvica, D. labialis... 158
Figura-B. 44; Esqueleto axial, cintura pélvica, D. labialis... 159
Figura-B. 45; Esqueleto apendicular, miembro anterior, grupo propio... 160
Figura-B. 46; Esqueleto apendicular, miembro anterior, A. marinkellei MLS 558... 161
Figura-B. 47; Esqueleto apendicular, miembro anterior, A. mittermeieri... 161
Figura-B. 48; Esqueleto apendicular, miembro anterior, A. mittermeieri... 162
Figura-B. 49; Esqueleto apendicular, miembro anterior, C. granulosus... 162
Figura-B. 50; Esqueleto apendicular, miembro anterior, E. bogotensis... 163
Figura-B. 51; Esqueleto apendicular, miembro anterior, D. labialis... 163
Figura-B. 52; Esqueleto apendicular, miembro anterior, D. labialis... 164
Figura-B. 53; Esqueleto apendicular, mano, A. muisca... 165
Figura-B. 54; Esqueleto apendicular, mano, A. lozanoi.... 166
Figura-B. 55; Esqueleto apendicular, mano, A. marinkellei... 167
Figura-B. 56; Esqueleto apendicular, mano, A. mittermeieri... 168
Figura-B. 57; Esqueleto apendicular, mano, A. mittermeieri... 169
Figura-B. 58; Esqueleto apendicular, mano, A. granulosus... 170
Figura-B. 59; Esqueleto apendicular, mano, E. bogotensis... 171
Figura-B. 60; Esqueleto apendicular, mano, D. labialis... 172
Figura-B. 61; Esqueleto apendicular, mano, D. labialis... 173
Figura-B. 62; Esqueleto apendicular, miembro posterior, A. muisca MUJ 179 ... 174
Figura-B. 63; Esqueleto apendicular, miembro posterior, A. muisca MUJ 180 ... 175
Figura-B. 64; Esqueleto apendicular, miembro posterior, A. lozanoi MUJ 519... 176
Figura-B. 65; Esqueleto apendicular, miembro posterior, A. lozanoi MUJ 952... 177
Figura-B. 66; Esqueleto apendicular, miembro posterior, A. marinkellei MLS 558, Hembra ... 178
Figura-B. 67; Esqueleto apendicular, miembro posterior, A. marinkellei MLS 558, Macho ... 179
Figura-B. 68; Esqueleto apendicular, miembro posterior, A. mittermeieri MUJ 3392... 180
Figura-B. 69; Esqueleto apendicular, miembro posterior, A. mittermeieri MUJ 4999... 181
Figura-B. 70; Esqueleto apendicular, miembro posterior, A. mittermeieri MUJ 5001... 182
Figura-B. 71; Esqueleto apendicular, miembro posterior, A. mittermeieri MUJ 5086... 183
Figura-B. 72; Esqueleto apendicular, miembro posterior, C. granulosus MUJ 4645 ... 184
Figura-B. 73; Esqueleto apendicular, miembro posterior, C. granulosus MUJ 4646 ... 185
Figura-B. 74; Esqueleto apendicular, miembro posterior, E. bogotensis MUJ 093 ... 186
Figura-B. 75; Esqueleto apendicular, miembro posterior, E. bogotensis MUJ 478 ... 187
Figura-B. 77; Esqueleto apendicular, miembro posterior, D. labialis MUJ 912 ... 189
Figura-B. 78; Esqueleto apendicular, miembro posterior, D. labialis LHA 022... 190
Figura-B. 79; Elementos sesamoideos, A. muisca MUJ 179 ... 191
Figura-B. 80; Elementos sesamoideos, A. muisca MUJ 180 ... 191
Figura-B. 81; Elementos sesamoideos, A. lozanoi MUJ 519... 192
Figura-B. 82; Elementos sesamoideos, A. lozanoi MUJ 952... 192
Figura-B. 83; Elementos sesamoideos, A. marinkellei MLS 558 Hembra ... 193
Figura-B. 84; Elementos sesamoideos, A. marinkellei MLS 558 Macho... 193
Figura-B. 85; Elementos sesamoideos, A. mittermeieri MUJ 3392... 194
Figura-B. 86; Elementos sesamoideos, A. mittermeieri MUJ 4999... 194
Figura-B. 87; Elementos sesamoideos, A. mittermeieri MUJ 5001... 195
Figura-B. 88; Elementos sesamoideos, A. mittermeieri MUJ 5086... 195
Figura-B. 89; Elementos sesamoideos, C. granulosus MUJ 4645 ... 196
Figura-B. 90; Elementos sesamoideos, C. granulosus MUJ 4646 ... 196
Figura-B. 91; Elementos sesamoideos, E. bogotensis MUJ 093 ... 197
INDICE DE TABLAS
Tabla 1; Ejemplares de la colección transparentados ... 44
Tabla 2; Ejemplares de la colección sometidos a proceso de transparentación ... 44
Tabla 3; Matrices realizadas ... 52
Tabla 4; Caracteres eliminados del análisis ... 67
Tabla 5; Caracteres usados en las matrices que incluyen los caracteres de tipo multiestado (CM)... 69
Tabla 6; Caracteres usados en las matrices que incluyen los caracteres de tipo binario (CB) ... 71
Tabla 7; Estados de carácter para la matriz de tipo CM–CB–CI ... 75
Tabla 8; Matriz de tipo CM–CB–CI. ... 80
Tabla 9; Estados de carácter para las matrices de tipo CB–CI ... 84
Tabla 10; Matriz de CB–CI. ... 88
Tabla 11; Matriz de CB–ncofl. ... 91
Tabla 12; Estados de caracteres para las matrices de tipo P- CI... 92
Tabla 13; Matriz de P-CI. ... 95
Tabla 14; Matriz de P (CB)-ncofl. ... 98
INDICE DE ANEXOS
ANEXO 1; Medidas Morfométricas ... 199
ANEXO 2; Morfometría de ejemplares transparentados ... 200
ANEXO 3; Abreviaturas ... 201
ANEXO 4; Descripción de Caracteres Osteológicos ... 203
ANEXO 5; Matriz general de datos - precaracteres y estados de carácter... 216
ANEXO 6; Fotos transparencias, Atelopus muisca MUJ 179 ... 224
ANEXO 7; Fotos transparencias, Atelopus muisca MUJ 180 ... 225
ANEXO 8; Fotos transparencias, Atelopus lozanoi MUJ 519 ... 226
ANEXO 9; Fotos transparencias, Atelopus lozanoi MUJ 952 ... 227
ANEXO 10; Fotos transparencias, Atelopus mittermeieri MUJ 3392 ... 228
ANEXO 11; Fotos transparencias, Atelopus mittermeieri MUJ 4999 ... 229
ANEXO 12; Fotos transparencias, Atelopus mittermeieri MUJ 5001 ... 230
ANEXO 13; Fotos transparencias, Atelopus mittermeieri MUJ 5086 ... 231
ANEXO 14; Fotos transparencias, Atelopus marinkellei MUS 558 Hembra... 232
ANEXO 15; Fotos transparencias, Atelopus marinkellei MUS 558 Macho ... 233
ANEXO 16; Fotos transparencias, Chaunus granulosus MUJ 4645 ... 234
ANEXO 17; Fotos transparencias, Chaunus granulosus MUJ 4646 ... 235
ANEXO 18; Fotos transparencias, Eleutherodactylus bogotensis MUJ 093 ... 236
ANEXO 19; Fotos transparencias, Eleutherodactylus bogotensis MUJ 478 ... 237
ANEXO 20; Fotos transparencias, Dendropsophus labialis LHA 022... 238
RESUMEN
Inicialmente es importante resaltar que hasta el momento no hay publicaciones
formales posteriores a Lynch (1993) acerca de la sistemática del grupo ignencens que incluyan las especies tratadas en este estudio, por consiguiente este trabajo buscó
presentar una hipótesis de parentesco de cuatro especies del género Atelopus
(Bufonidae; Anura) pertenecientes al grupo ignencens sensu Lynch (1993), fundamentada en caracteres osteológicos; ya que este grupo de especies no ha sido
posible verificarlo como una unidad monofilética basada en una posible sinapomorfia
como en el caso del grupo flavencens, sugerido como monofilético por Lynch (1993). Para alcanzar el objeto del trabajo se analizaron datos provenientes de 17 individuos
pertenecientes a siete especies en total, en lo posible hembra y macho para cada una,
construyendo matrices con caracteres multiestado y doble estado utilizando el método
cladístico de comparación por grupo ajeno. Para este grupo se usaron tres especies
(siete ejemplares) de las familias Hylidae, Brachycephalidae y Bufonidae, alternando
para el análisis el grupo ajeno inicial; así, mediante el programa Winclada-NONA
(Nixon, 2002) se generaron once cladogramas igualmente parsimoniosos y tres de
consenso estricto, los cuales fueron comparados determinando el índice de
consistencia y la longitud de cada uno; obteniendo como resultado hipótesis basadas
en posibles sinapomorfias que agrupan las especies del género Atelopus escogidas para este trabajo, proponiendo una aproximación en cuanto a las relaciones
filogenéticas del grupo ignencens. También se plantea la necesidad de incrementar el nivel del análisis a otras especies del género para poder verificar la posibilidad, que el
grupo ignencens sea una unidad monofilética.
Palabras Clave: Atelopus, cladograma, filogenia, grupo ajeno, grupo ignencens, grupo propio, índice de consistencia (IC), índice de retención (IR), longitud (L),
ABSTRACT
Initially it’s important to be clear that until the moment there aren’t formal
publications later to Lynch (1993) about the systematic of the group ignencens where they include the species tried in this study. For consequently, this work looked to
present a hypothesis of relationship of four species of the genus Atelopus (Bufonidae; Anura) belonging to the group ignencenssensu Lynch (1993), based in osteologycal characters; since this group of species has not been possible to verify it like an unit
monophyletic based on a possible synapomorphy, like the case of the group
flavencens, suggested as monophyletic by Lynch (1993). To reach the object of the work were analyzed in total dates that coming from 17 individuals belonging to seven
species, as much as possible female and male for each one, building matrix with
characters multistate and double state using the method comparison cladistic for other
outgroup. For this group were used three species (seven individuals) of the families
Hylidae, Brachycephalidae and Bufonidae, alternating for the analysis of the initial
outgroup; to this way, by means of the program Winclada-NONA (Nixon, 2002) were
generated eleven equally parsimonious cladograms and three of strict consent, which
were compared determining the index of consistency and the longitude of each one;
obtaining to hypothesis ace to result with possible synapomorphys that regroup the
species of genus Atelopus chosen for this work, proposed an approach to the phylogenetic relationships of the group ignencens. Also carries out about the necessity to increase the level from the analysis to other species of the gender to be
able to verify the possibility that the group ignencens will be a unit monophyletic.
1 INTRODUCCIÓN
Los anfibios poseen 5743 especies registradas alrededor del mundo (Rueda, et al,
2005); una gran parte de estas pertenecen al orden Anura, el cual para Colombia
consta de nueve familias, catalogándolo como el de mayor diversidad en este grupo
con cerca de 722 especies (Rueda, et al, 2004). Dentro de esta amplia diversidad, la familia Bufonidae distribuida entre 0 y 4500 msnm. Y caracterizada por individuos
con piel granular, pústulas y tubérculos, de hábitat terrestres y cavadoras, se
encuentran las conocidas “ranitas arlequín” pertenecientes al género Atelopus, las cuales son muy apetecidas por los traficantes debido a sus coloraciones vivas y
llamativas. Las especies que conforman este género se distinguen por sus rostros
puntiagudos, por pertenecer a ambientes húmedos y ser diurnas; para Colombia se
reconocen 38 especies descritas y 8 más por nominar (Rueda, et al, 2005) de las cuales varias ya están registradas en la lista de especies en peligro de extinción del
Libros Rojo para el país.
Las especies Atelopus muisca, A. lozanoi, A. mittermeieri y A. marinkellei escogidas para este trabajo, pertenecen al grupo ignescens sensu Lynch (1993) el cual no se ha podido comprobar como una unidad monofilética. Por esto, en busca de una
aproximación a la filogenia de este grupo de especies, se realizó el presente trabajo
donde se elaboraron hipótesis de parentesco para las cuatro especies de Atelopus
usando como grupos ajenos las especies Dendropsophus labialis, Eleutherodactylus bogotensis y Chaunus granulosus de las familias Hylidae, Brachycephalidae y Bufonidae respectivamente. Los ejemplares usados forman parte de las colecciones
biológicas de la Pontificia Universidad Javeriana y de la Universidad de la Salle,
individuos que fueron tratados con el método de aclaración enzimático y coloración
diferencial para la observación e identificación de caracteres osteológicos que facilitó
Por otro lado, la hipótesis de parentesco obtenida en este estudio en esencia, más que
ofrecer una solución al problema filogenético, buscó exponer un acercamiento a las
relaciones interespecíficas y además encontrar en el análisis de los huesos una
propuesta con respecto al manejo de los caracteres en los taxones trabajados. Por esto,
la importancia de éste trabajo radica en la contribución al conocimiento del género
Atelopus, específicamente del grupo ignescens y su filogenia; además, los datos obtenidos, pueden ser ampliados tanto en el número de caracteres como en el de
especies, en busca de aclarar la monofília del grupo, o pueden llegar a ser utilizados
en la reconstrucción filogenética de otros grupos de anuros.
2 MARCO TEÓRICO Y REVISIÓN DE LITERATURA
2.1 Antecedentes
Por más de tres siglos (XVI-XIX) el proceso del conocimiento científico sobre los
anfibios de Colombia estuvo dominado por investigadores extranjeros como Dumeril,
Bibron, Gray Günther y Boulenger, entre otros, pertenecientes a centros del
conocimiento científico de la época tal como los Museos de Historia Natural de París
y Londres; posteriormente para la década de 1910 los naturistas franceses, Hermano
Apolinar María (Nicolás Séller) y El Hermano Nicéforo María (Naline Rouhaire
Siauzade), sentaron definitivamente las bases para el estudio de los anfibios y reptiles
en Colombia, con la creación del Museo de la Salle en Bogotá y la edición del Boletín
de la Sociedad de Ciencias Naturales de la Salle (Rueda, et al, 2004). En la actualidad, existen varios grupos de trabajo en diferentes universidades del país, que
dirigen sus esfuerzos al conocimiento de la taxonomía de los anfibios de Colombia y
al establecimiento de sus relaciones filogenéticas, así como al avance del
conocimiento sobre su ecología y fisiología (Rueda, et al, 2004).
Sin embargo, aunque recientemente se encuentran numerosos estudios basados en
caracteres moleculares, aun estamos influenciados por las prácticas tradicionales
(Lynch, et al, 2001), así, como para la determinación de sus relaciones evolutivas. Tales influencias están marcadas por estudios monumentales como el de Hoffman
(1932) o Laurent (1940, 1941, 1942, 1943, entre otros), quienes realizaron unos
extensivos trabajos osteológicos para reorganizar grupos africanos y asiáticos de
ránidos; al igual que los realizados por autores, tales como Liem (1970) que utilizó
morfología interna y externa para establecer la sistemática y evolución de las ranas
arborícolas del viejo mundo; o Trueb (1973) quien trabajó significativamente en los
huesos de las ranas y su evolución, describiendo las generalidades de las
características óseas, especificando las particularidades de algunas familias y géneros,
llegando a determinar estados, patrones y tendencias evolutivas; otro igualmente
importante es el de Clarke (1981) que por su parte realizó un estudió de la osteología
comparada y las relaciones evolutivas en ránidos africanos .
También han sido determinantes numerosos estudios morfológicos mucho más
puntuales, como por ejemplo el de Ritland (1955) en el que se describen y analizan
aspectos de la anatomía post-craneal de Ascaphus. Pero acercándonos un poco más al grupo de estudio, como en el caso de algunos bufónidos, se identifican diversos
trabajos basados en la de osteología craneal, entre los que se incluyen los presentados
por Parker (1876), Ramaswami (1937), Sedra (1949), Sanders (1953), Baldauf (1959)
y Tihen (1962) (citados por McDiarmid 1971) los cuales fueron consultados e
interpretados por diferentes autores para hallar relaciones morfológicas en la familia y
los especimenes cercanos, entre ellos, Pramuk (2000, 2001, 2002 y 2006) quien ha
realizado diferentes estudios filogenéticos en bufónidos, basándose en caracteres
osteológicos y moleculares.
Por otro lado, ya concretando en el género Atelopus, es importante resaltar el hecho que se han estudiado la mayoría de las estructuras óseas, algunos músculos, la
biología, ecología, distribución, evolución y sistemática; por ejemplo, en el estudio de
McDiarmid (1971) sobre la morfología comparada y la evolución de este género
de los anuros de forma comparada ilustrando los cráneos de tres especies de este
género; también citó a Badenhorst (1945) que trabajó con la osteología craneal de
Melanophryniscus a Laurent (1942) quien discutió ciertos aspectos osteológicos de
Atelopus varius y Noble (1922) el cual ilustró un aspecto ventral de la columna vertebral de Atelopus varius, mostrando claramente la fusión de la primera vértebra truncada con el atlas, y con el cual estableció la importancia de las características
vertebrales para la clasificación de las ranas.Otro autor citado por McDiarmid (1971)
es Griffiths (1954), quien presentó un diagrama del cráneo de Atelopus ignescens.
Otro trabajo fundamental sobre este género, es el realizado por Lynch (1993),
respecto a la relación sistemática intraespecífica, en el que se generó un arreglo
notable en los grupos de especies; seguido por el estudio de Lötters (1996) quien
recopila la información de varios autores en diferentes especies del género; este
trabajo es una excelente guía que encierra un resumen de la biología, la distribución
del género y una lista de especies y subespecies reconocidas hasta el momento,
además presenta generalidades morfológicas en el aspecto osteológico, entre otras.
A nivel nacional se han realizado varios estudios taxonómicos en la colección de
anfibios de diferentes instituciones, con el género Atelopus, por ejemplo Ruiz, Ardila & Hernández (1994) describieron y nominaron tres nuevas especies endémicas de la
sierra Nevada de Santa Marta, A. laetissimus, A. nahumae y A. leoperezii. Se ha contribuido también con descripciones de especies para la ciencia (incluyendo sus
caracteres osteológicos), como es el caso Osorno & Ruiz (1994) que describieron dos
especies del grupo ignescens, A. simulatus y A. sernai y una del grupo longirostris, A. quimbaya.
Por otro lado, de las partes altas de la Cordillera Oriental de Colombia se destacan
estudios como el de Rueda & Hoyos (1991) en el que describen una nueva especie de
género como el trabajo de Grado de Medina (2000) el cual se basó en la miología y
osteología de A. muisca del Parque Nacional Natural Chingaza; por otro lado Osorno, Ardilla & Ruiz (2001) nominaron y describieron tres nuevas especies (A. guitarraensis, A. lozanoi y A. mandingues) pertenecientes al grupo ignescens (sensu
Lynch, 1993); y actualmente se esta investigando y trabajando en una nueva especie
para Colombia, A. mittermeieri y como primer resultado ya se obtuvo la reciente publicación (Acosta, et al, 2006) en la cual se encuentra la nominación y correspondiente descripción, con algunas implicaciones ecológicas.
En conclusión, es importante entender y tener en cuenta que al lado del conocimiento
de los rasgos estructurales, como el sistema esquelético, es importante entender la
filogenia, la etología y ecología del organismo. Por esto, trabajos de autores como
Parker (1881), Noble (1922), Griffiths (1954), entre otros, han sido recopilados por
Lötters (1996) y han contribuido a entender numerosos aspectos de la biología acerca
del grupo Atelopus.
2.2 Género Atelopus
2.2.1 Generalidades
Colombia es uno de los países con mayor diversidad en número de especies de
anfibios en el mundo. Estos nominados como clase anfibia esta compuesta por 3
ordenes vivientes (Lynch, et al, 2001), diferenciándose anatómicamente de otros grupos porque en el cráneo tienen dos cóndilos occipitales, un gran hueso escamoso
que usualmente esta fusionado con el parietal, una hendidura ótica cerrada; la fosa
postemporal y ectopterigoide ausente; no más de una vértebra sacra; piel glandular y
huevos anamnióticos (Avilán, 2000).
A esta clase anfibia pertenecen tres órdenes actuales: Anura (ranas y sapos), Urodela
conocidos en las regiones templadas y alcanzan una gran variedad en los trópicos. Su
alta especialización se manifiesta en las muchas modificaciones estructurales sobre
todo esqueléticas para poder saltar (Romer, et al, 1981), por ejemplo, presentan características anatómicas como una columna vertebral corta con 6-8 vértebras
presacras; presentan procesos transversos, presencia de urostilo, cintura pélvica
alargada; el sacro en la mitad del cuerpo; cola ausente en los adultos; extremidades
posteriores alargadas modificadas para saltar y una fusión entre la tibia y la fíbula
(Avilán, 2000).
En Colombia se han registrado cerca de 722 especies de ranas (Rueda, et al, 2004) llegando a ser uno de los países con mayor diversidad. Entre esta amplia gamma, se
encuentra la familia Bufonidae un grupo grande con una distribución cosmopolita, a
excepción de Australia donde está el único bufónido introducido Bufo marinus (=
Chaunus marinus), que se ha convertido en un parásito y peligro a la fauna local debido a su apetito voraz, gran tamaño, alta fecundidad y toxicidad.
El estereotipo de esta familia incluye características tales como: los miembros
delanteros acortados y los traseros usados para caminar o saltar, la piel seca y
verrugosa, las glándulas del parotoide detrás de los ojos. Los caracteres compartidos
dentro de la familia son: 1) presencia del órgano de Bidder; 2) dientes ausentes en la
mandíbula y la maxila; 3) músculo posterior del constrictor ausente; 4) músculo
depresor mandibulae se origina solamente del escamoso; 5) cráneo altamente osificado (la piel está osificada generalmente al cráneo) (Vélez, et al, 1997). Los bufónidos se ubican en el grupo Neobatrachia, pero las relaciones entre las familias
de estas ranas "avanzadas" son bastante polémicas (Rueda, et al, 2004).
2.2.2 Origen del género
El género Atelopus fue propuesto por Duméril y Bibron (1841) (McDiarmid, 1971); es el segundo más grande dentro de la familia Bufonidae y probablemente el más
menos 81 especies y subespecies nominales y 32 más por describir (La Marca et al,
2005; en : Acosta et al, 2006); endémico del neotrópico, agrupa especies de ranas
llamadas comúnmente “arlequín”, cuya distribución se extiende desde el noroccidente
de Costa Rica hasta el noroeste de Bolivia (Ruiz, et al,1994b); al parecer la mayor
diversidad de este grupo ha ocurrido en territorio colombiano (Rueda, et al, 1991) y Ecuatoriano, donde se han registrado el mayor numero de especies del género
(Lötters, 1996).
Presumiblemente, este género se ha adaptado al ámbito natural a orillas de arroyos y
se ha movido a las áreas montañosas y selváticas húmedas cálidas, que se hicieron
disponibles con el levantamiento de los Andes, durante el Terciario o Cretácico
superior (probablemente porque estuvo presente en Sudamérica antes del comienzo
del Cenozoico enhábitat del Sureste de Brasil (McDiarmid, 1971) y allí, adaptándose
a cursos de agua torrentosos, adquirió las adaptaciones que luego, como resultado de
movimientos tectónicos ocurridos durante el Mioceno, el Plioceno y Pleistoceno
Inferior, produjeron la ortogénesis andina y ofrecieron nuevos hábitat para la
ocupación por parte de elementos provenientes de tierras bajas (Ruiz, et al, 1994a); estos nuevos hábitat fueron el mayor factor que contribuyó a su radiación de especies
(McDiarmid, 1971).
Tal radiación hizo que se colonizaran diferentes ambientes como los presentes en la
compleja fisiografía del continente suramericano (extensas áreas tropicales,
subtropicales y bosques templados) y estos hábitat son en general un centro de
diversidad para infinidad de especies como los anuros, por lo tanto se pueden
encontrar especies de Atelopus al nivel del mar en climas cálidos, en bosques húmedos, otras especies están adaptadas al frío, en páramos y subpáramos hasta los
4500 m de elevación, aunque la distribución para varias de las especies ha sido
2.2.3 Descripción del género
Estas ranas son de tamaño pequeño o moderado (20-60 mm de longitud corporal); la
mayoría se caracterizan por los colores llamativos; considerablemente diferentes
porque su cabeza es usualmente larga y ancha; el hocico es fuertemente acuminado;
ojos relativamente pequeños, la pupila es horizontal elíptica; no tienen tímpano
externo, aunque pueden tener una elevación (supratímpanica) en esa misma área
(McDiarmid, 1971); extremidades posteriores proporcionalmente cortas; manos y
pies con el primer dedo bastante reducido, por lo general las manos carecen de
palmeaduras, dedos angostos y no finalizan en discos expandidos; los pies pueden
llegar a ser bastante palmeados; la textura de la piel puede variar desde lisa y delgada
hasta incluir tegumentos muy tuberculados y glandulares (Ruiz, et al, 2005).
En cuanto a la biología del género muchas especies son terrestres cuando adultas,
pero pueden ser semicavadoras (McDiarmid, 1971), se les encuentra cerca de áreas
muy húmedas y a muy baja altura del suelo; el apareamiento presenta amplexus axilar
y puede ser muy prolongado; las posturas son enrolladas alrededor de piedras u otro
objeto bajo el agua; tienen un numero bajo de huevos (200-300), dispuestos a manera
de una ristra o rosario; los renacuajos son reofílicos y se alimentan de microalgas; los
adultos se alimentan de hormigas, grillos y pequeños artrópodos en general; son
longevos (hasta 10 años) y de crecimiento lento (Ruiz, et al, 2005). A continuación se describen algunos aspectos referentes a las especies utilizadas en la realización de
este trabajo.
Atelopus muisca
A. muisca Rueda & Hoyos (1991), conocida como "sapito arlequín esmeralda"; endémica de Colombia; distribuida en los páramos y subpáramos entre 2900-3350
msnm (Rueda, et al, 1991); se encuentra en bosque húmedo montano, vegetación de páramo, preferiblemente al interior de los frailejonales Espeletia spp. y pajonales
laderas protegidas con bosques relictuales de encenillo Weinmannia spp., puede ser encontrada generalmente a la orilla de las quebradas o bajo rocas (Rueda, et al, 2004).
Es una especie perteneciente al grupo ignescens, tamaño corporal 30-35mm, reconocible de las demás integrantes del género por su patrón cromático verde
esmeralda, con algunas estructuras tegumentarias que van desde amarillo a verde
limón y en la región ventral verde esmeralda
pálido con diseño negro sobre el pecho
(Rueda, et al, 2004); palmas y plantas de color variado pardo oscuro con verde y amarillo
quemado hacia el extremo de los dedos; Pies
palmeados; no se conocen sus renacuajos;
posturas en largas hileras (Ruiz, et al, 2005).
Figura-A. 1; A. muisca. (Rueda, et al, 2004) El estudio de Medina (2000) en el cual se describe al detalle, la miología y osteología
de la especie, proporcionó numerosas contribuciones en la comprensión de los
diferentes aspectos de los caracteres descritos para A. muisca, los cuales fueron utilizados en este trabajo.
Atelopus lozanoi
A. lozanoi Osorno, Ardila & Ruiz (2001), llamada comúnmente como "sapito
arlequín de La Calera". Conocida
únicamente en el Páramo de Palacio y sus
alrededores en el Parque Nacional
Natural Chingaza (Lynch, et al, 2001), en
Figura-A. 2; A. lozanoi (Rueda, et al, 2004)
la vertiente oriental de la Cordillera Oriental, en el departamento de Cundinamarca,
difiere por el tamaño ya que A. lozanoi es ligeramente mayor (29-38mm) (Lynch, et al, 2001). Es una especie del grupo ignescens (sensu Lynch, 1993).
Igual que las otras especies del género son de hábitos diurnos, se encuentran
asociadas a quebradas del páramo, Depositan hileras de huevos (forma de rosarios) en
aguas torrentosas, los cuales se adhieren debajo de las rocas dentro de las quebradas
(Rueda, et al, 2004). Navas (1996) sugieren que sapos de esta especie están activos a
temperaturas ambientales muy bajas, porque de esta manera reducen el riesgo de
desecación sin reducir significativamente su desempeño en aquellas actividades que
realiza fuera de sus refugios (locomoción, forrajeo, vocalización) (Rueda, et al,
2004).
Atelopus marinkellei
A. marinkellei Cochran & Goin (1970). Se conoce sólo de la vertiente oriental del norte de la Cordillera Oriental de Colombia, departamento de Boyacá (entre los
2800-3650 metros) y típicamente en el Páramo de Vijagual (entre Tunja y Zetaquirá a
2660 m), Pisba y Toquilla; habita en los pajonales y
matorrales de páramo y subpáramo, muy cerca de
grandes ríos y quebradas de aguas limpias y
trasparentes. Presentan un tamaño entre 44-48 mm;
superficie ventral y dorsal de color negro o sepia con
pequeñas manchas semi-redondeadas de color
blanquecinas (Rueda, et al, 2004, 2005).
Figura-A. 3; A. marinkellei (Rueda, et al, 2004).
Algunas opiniones acerca de A. marinkellei y A. ebenoides, ambos conocidos comúnmente como “sapitos arlequín negro”, sostienen que deben ser reconocidas
como especies válida y no meramente como entidades subespecífica (A. ebenoides ebenoides y A. ebenoides marinkellei), como algunos autores consideran debido principalmente a sus distribuciones disyuntas, separadas entre sí por cientos de
A. ebenoides Rivero (1963) se caracteriza por habitar en los páramos de la vertiente occidental en el sur de Cordillera Central en los departamentos de Huila y Cauca a
3600 msnm, extendiéndose incluso hasta la Laguna de La Cocha en Nariño. Es una
rana robusta con extremidades relativamente cortas, presenta verrugas en el dorso y
lateralmente, la cabeza es relativamente lisa (Löthers, 1996), y presentan una longitud
entre 40-48mm; hembras adultas presentan pequeñas manchas semirredondeadas en
el dorso y el vientre de color amarillas (Rueda, et al, 2004). Lastimosamente debido a que no fue posible conseguir ejemplares pertenecientes a este grupo, no pudo ser
incluido en el análisis del proyecto como estaba presupuestado.
Atelopus mittermeieri
A. mittermeieri Acosta, Rueda, Velásquez, Sánchez & Peña (2006), conocida como Arlequín de Guanentá, está distribuida en la vertiente occidental de la Cordillera
Oriental de Colombia, específicamente en
el departamento de Santander, en tres
localidades aledañas, situadas dentro o en
la periferia del Santuario de Flora y Fauna
Guanentá-Alto Río Fonce, entre los
2250-2750 msnm en bosques de niebla (Rueda,
et al, 2005; Acosta, et al, 2006).
Figura-A 4; A. mittermeieri (Acosta, et al, 2006)
Exponente del grupo ignescens; presentan tamaño mediano y cuerpo robusto (Machos 32.9-33.3 y hembras 41.3-44.9); palmeadura manual vestigial,
membrana pedial extensa; miembros posteriores cortos; dorso
verrugoso de color café o negro con algunas manchas crema o
blancas redondeadas; el vientre varía de color café claro a rojo
encendido; hocico proyectado, tímpano y anillo timpánico
ausentes (Rueda, et al, 2005; Acosta, et al, 2006).
2.2.4 Osteología del género
Los caracteres osteológicos que identifican al género Atelopus según McDiarmid (1971) y Lötters (1996) son: hueso frontoparietal de extensión normal separado o
fusionado en la línea media; surco occipital serrado, la membrana de la plataforma
premaxilar es 2 veces el ancho de la membrana medial; la septomaxila tiene una
fuerte forma de U; Palatinos presentes; parasfenoides puntiagudo anteriormente,
superponiendo el complejo esfenetmoideo medial; el tronco del escamoso usualmente
presenta proceso anterior, el brazo dorsal se extiende sobre el proótico, manteniendo
la forma y tamaño en la mayoría de las especies del genero; cuadradojugal bien
desarrollado o reducido; pterigoide bien desarrollado; complejo esfenetmoideo
osificado; orbitosfenoides desosificado; cuadrado cartilaginoso, restringido a la parte
baja del suspensorium; la curva prearticular en la mandíbula posterior esta en punto
medio, el cartílago de Meckel es desosificado.
En cuanto al aparato hiolaringeo se caracteriza por: presentar la placa hioidea más
larga que ancha; el proceso anterior del hial muy pequeño y recto lateralmente; El
proceso anterolateral (hypobranchial I) presente con una base estrecha y el proceso
posterolateral (hypobranquial II) usualmente ausente.
El tronco del cuerpo esta conformado por siete vértebras presacras, presentando
fusión entre la primera vértebra y la segunda; procesos transversos presentan una
marcada variación intraespecífica en cuanto a forma, largo y dirección; sacro
usualmente separado de la vértebra; cóccix con articulación bicondilar o fusionada
con el sacro; diapófisis sacra grande, expandida y elípticamente aplanada; ilion y
diapófisis sacra articulada en una coyuntura plana; cintura pectoral firmisternal,
aunque para algunos autores hay controversia al respecto; esternón presente.
formula falangeal manual 2-2-3-3 o 1-2-3-3 (aunque hay variaciones) y formula
falangeal pedial 2-2-3-4-3.
2.2.5 Distribución del género
Los Andes que es el principal hábitat de este género, conforma un eje montañoso
topográficamente complejo, conformado por las Cordilleras Occidental, Central y
Oriental situadas hacia el occidente del país, separadas por los profundos y amplios
valles de los ríos Magdalena y Cauca. La Cordillera Occidental tiene una altura media
de 2000 metros, mientras que las Cordilleras Central y Oriental tienen alturas medias
superiores a los 3000 y hasta pueden sobrepasar los 4000; en algunas regiones
alcanzan elevaciones de más de 5000 metros sobre el nivel del mar en sus partes más
altas, como es la masa aislada de la Sierra Nevada de Santa Marta (Lynch, et al,
2001). Esta topografía es muy importante en la distribución latitudinal de varios
anuros.
La distribución del género es alopátrica (Lötters, 1996) y la mayoría de las especies
de Atelopus son encontradas en áreas tropicales del centro de Costa Rica hacia el Sur a través de Panamá, a lo largo de los Andes de la parte de Suramérica de Colombia,
Ecuador, Perú y Bolivia; y el área montañosa del norte de Venezuela. También se han
registrado poblaciones de algunas especies aisladas en la Guyana, Surinam, Guiana
francesa y noreste de Brasil (McDiarmid, 1971) y el estado de Recife, en la costa
oriental de Brasil (Rueda, et al, 1991); unas pocas especies habitan las pluviselvas tropicales de tierras bajas, en tanto que la gran mayoría (81%) viven en las zonas
montañosas en donde llegan incluso a penetrar en él limite de las nieves perpetuas
(Ruiz, et al, 2005).
Por ejemplo, hay registros de poblaciones distribuidas en el sistema montañoso de los
Andes colombianos a elevaciones comprendidas entre 300m y 4410m, como también
en Isla Gorgona y el litoral pacífico aledaño, mientras que otras viven en las tierras
algunas especies son encontradas incluso hasta los 4800 m, siendo por esto quizás,
dentro de los Anura, el género con mayor tolerancia térmica y altitudinal. Aunque no
existen especies conocidas representantes de este género en áreas de sabana naturales
con selvas riparias o freatófitas próximas a cursos de agua permanentes o transitorios,
como tampoco en áreas de bosque caducifolio (o hidrofíticos) isomegatérmicos como
los que caracterizan gran parte de la planicie litoral del norte de Colombia, se han
encontrado especies del género en el Macizo de la Sierra Nevada de Santa Marta, por
esto se considera el Macizo como un centro secundario de diversificación del género
(Ruiz, et al, 1994a).
Lötters (1996) recopiló cerca de 300 publicaciones sobre el género Atelopus donde se puede visualizar la distribución política del género en Colombia, destacándose los
departamentos de Amazonas, Antioquia, Boyacá, Cauca, Cesar, Córdoba,
Cundinamarca, Chocó, Guajira, Huila, Magdalena, Nariño, Putumayo, Quindío,
Risaralda y Tolíma.
2.2.6 Estado de conservación del género
Dentro de la diversidad biológica, los anfibios, conjuntamente con las aves, tienen
para nuestro país el mayor número de especies a nivel mundial, destacándose también
el alto grado de endemismo, que alcanza el 55% del total de especies registradas en el
país (Rueda, et al, 2004); por esto, es particularmente significativo que en Colombia uno de los grupos más vulnerables sean los anfibios. Es más, actualmente la crisis que
enfrenta este grupo es bastante dramática, ya que de las 5743 especies del mundo
cerca del 32% se encuentran amenazadas de extinción, incluidas las 337 especies
consideradas en Peligro Crítico (Rueda, et al, 2005).
A raíz de estas desapariciones rápidas, repentinas de un gran numero de poblaciones
de anfibios, se llevó a cabo el proceso de categorización de la fauna anfibia
amenazada de extinción en Colombia el cual dio como resultado 55 especies
arlequines endémicas que se encuentran extintas y muchas otras experimentando una
significativa reducción de los tamaños poblacionales durante los últimos 15 años
(Rueda, et al, 2005), incluyendo tres de las especies a trabajar como son: A. ebenoides (incluyendo sus entidades subespecíficas), A. muisca y A. lozanoi.
Las principales amenazas que afectan las poblaciones de anuros son la destrucción y
deterioro del hábitat natural; los efectos de fragmentación; los cambios climáticos y el
aumento de la radiación UV; la sobreexplotación de algunas especies; la introducción
de especies exóticas y la contaminación (Rueda, et al, 2004), aunque recientemente se ha descubierto que algunas desapariciones se encuentran asociadas a infecciones
provocada por hongos patógenos y como incidencia de este, otros patógenos como
virus y bacterias han aumentado, y se piensa que es probable que un conjunto de
problemas esté debilitando el sistema inmunológico de los anfibios, en general
(Rueda, et al, 2005).
Se considera que los criterios que hacen que se encuentren en Peligro critico (CR) las
especies A. muisca, A. lozanoi y la subespecie A. ebenoides ebenoides tanto en la Categoría Global UICN como en la Categoría Nacional de Amenaza de especies en
vía de extinción son: para el primer grupo, el rápido declive poblacional, la pequeña
área y la severa fragmentación, generando una disminución continua; en el segundo
grupo, se ha venido observando una disminución progresiva en sus poblaciones
naturales, mientras que para el ultimo se ha visto una rápida disminución poblacional
en un 90 por ciento de la distribución original (Rueda, et al, 2004). A. ebenoides merinkellei está en la Categoría UICN, en Peligro Critico ya que hasta mediados de 1995 eran muy comunes pero comenzaron a morirse en grandes cantidades a las
orillas de los caminos y acequias; actualmente son muy raras y tan solo se avistó un
individuo a comienzos del 2004 y en prospecciones de campo hachas en el 2005 no se
2.2.7 Sistemática del género
La sistemática de los anuros, en las categorías taxonómicas sobre niveles específicos
ha estado basada en relaciones evolutivas deducidas de datos conservados y
caracteres relativamente estables de la morfología interna principalmente (Brattstrom,
1957: en McDiarmid, 1971). Históricamente, para la identificación y clasificación de
las ranas hemos estado sujetos a disponer de caracteres anatómicos (Lynch, et al,
2001), aunque en muchos casos la morfología externa e interna se complementan con
la información que puede ser obtenida por la etología, ecología, compatibilidad
genética y bioquímica (McDiarmid, 1971). Así, bajo este esquema de obtención y
análisis de información, Lötters había registrado 66 especies de Atelopus hasta 1996, que han sido descritas con base sólo en su morfología externa, ya que las
características de hábitos se consideraron relativamente uniformes en la mayoría de
las especies del género así como la variación intraespecífica que muchas veces es más
grande que la variación interespecífica (Lötters, 1996).
Por esto, el énfasis en la morfología interna y los estudios detallados en diferentes
especies, principalmente de la osteología, son extremadamente importantes ya que
permiten comparar las formas ancestrales con las modernas o descendientes, para
poder entender la filogenia y contribuir ampliamente al conocimiento de las
relaciones evolutivas. Estos estudios se pueden realizar, ya sea, por el análisis de
grupos particulares de organismos relacionados, el análisis de un carácter
morfológico especifico o el de un sistema a través de un gran espectro de organismos
no relacionados. Un claro ejemplo de estos tipos de estudios es el realizado por
McDiarnid (1971) donde analiza el sistema músculo-esquelético de Atelopus.
Siguiendo el patrón de clasificación que se basa en la morfología, Peters (1973)
propuso para el género dos grupos fenéticos, Atelopus ignescens y A. longirostris,
basado en los índices de las relaciones morfométricas tales como: longitud cefálica
rostro-cefálica y de la misma con la longitud femoral, asociando las diferencias de color y la
distribución altitudinal, quedando el grupo longirostris que comprendía individuos de cuerpo delgado, con extremidades relativamente largas, y un grupo ignescens con cuerpos más robustos y extremidades relativamente cortas (Lötters, 1996).
Aunque McDiarmid (1971) anota, sin aplicarlo a la construcción de los grupos, que
en este género hay reducción en número y longitud de falanges en el dedo I, fue sólo
hasta la publicación de Lynch (1993), quien sugiere que en cualesquiera de los grupos
de Atelopus arriba anotados se presentan especies con miembros posteriores largos o cortos con relación a la longitud del cuerpo; así, buscando caracteres que permitan
construir grupos naturales, encontró que algunas de las especies del grupo ignescens
tienen un pulgar más largo (= dos falanges).
De lo anterior se concluye que el uso del número de falanges en el dedo I manual en
las especies de Atelopus, independiente del tamaño de los mismos, es de mayor utilidad para la constitución de grupos naturales que las relaciones morfométricas
entre segmentos corporales, diferencias altitudinales y deferencias en los colores
(Ruiz, et al, 1994b). Por esto, la presencia de las piezas tarsianas y el número de dedos en las manos y los pies, la forma terminal de los dedos, la presencia y
configuración de tubérculos en las palmas y la condición de la unión interdigital, son
características significativas para ser usadas en la sistemática de anuros y por ende en
este género (McDiarmid, 1971).
Lötters (1996) concluyó que “hay un notable arreglo en el grupo flavencens (sensu
Lynch 1993) que contiene 25 especies y ha sido sugerido como monofilético con base
en una sinapomorfìa hipotética, porque, los miembros de este grupo muestran una
reducción en el numero de falanges del pulgar (mostrando una formula 1-2-3-3). En
consecuencia todas las especies del reorganizado grupo presentan unidades filéticas
Por otro lado, también podemos considerar que en muchos casos, el aspecto
zoogeográfico es informativo para la taxonomía de Atelopus ya que los grupos de ranas de este género tienen un rango muy restrictivo, distribuciones alopátricas en
elevaciones altas y muchas formas sólo se conocen en áreas relativamente pequeñas,
como páramos, cimas de montañas o valles, áreas que actúan como barreras en el
flujo genético (Lötters, 1996).
Por ejemplo en la actualidad se presenta un conflicto taxonómico como se mencionó
anteriormente, en virtud del gran hiato geográfico existente entre las poblaciones de
A. e. ebenoides y A. e. marinkellei, del sur de la Cordillera Central y las de la porción septentrional de la Cordillera Oriental; sin embargo hasta el momento no se han
aportado las evidencias taxonómicas que justifiquen tal segregación (Rueda, et al,
2004). Lötters (1996), también identificó esta dualidad afirmando que, debido a su
distribución restringida, entre otras cosas, es posible que A. e. marinkellei sea una especie diferente a A. e. ebenoides, teniendo como base que la región Andina, al presentar condiciones ambientales que pueden cambiar radicalmente en distancias
verticales y horizontales cortas, como cursos de montañas discontinuas con rápidos
cambios de elevación, pueden causar diferentes meso y microclimas. Estos ambientes
heterogéneos pueden actuar no sólo como barreras ecológicas sino dificultar
físicamente el movimiento de las especies, por lo tanto estas barreras pueden resultar
en alopatría y endemismos.
2.3 Análisis filogenético
2.3.1 Generalidades del análisis filogenético
Para el estudio de los aspectos históricos de la evolución, la sistemática como ciencia
provee herramientas útiles para ello. Además de ser un buen método de clasificación
biológica que ayuda al estudio de la diversidad orgánica, este método de clasificación
agrupa organismos homogéneos entre sí (respecto a otro grupo de organismos
similares) o taxones, que comparten caracteres derivados (Schuch, 2000), en otras
palabras los taxones tienen caracteres o atributos que según como halla sido su origen
pueden definir niveles jerárquicos dentro de un grupo de organismos. Este método,
orientado por la escuela de sistemática cladística tiene como fundamento que la
escogencia de los caracteres derivados para el análisis tiene “valor”, es decir que es
con ellos que se logra establecer una relación de parentesco entre dos taxones, por lo
menos, con respecto a un tercero (Hoyos, 1994).
Es importante tener muy claro para este tipo de análisis, algunos términos básicos e
importantes correspondientes a los caracteres; por ejemplo, cuando un atributo
compartido por dos especies y es heredado a partir de un ancestro común, se
considera una homología, pero si los organismos no están emparentados pero
igualmente presentan características con función y apariencia similar (análogas) como
resultado de una evolución convergente se consideran homoplasias (Lipscomb, 1998),
estas ultimas generalmente definen grupos diferentes y conflictivos (Schuh, 2000),
que no pueden ser explicados.
Por otro lado, los caracteres se clasifican como ancestrales (plesiomórficos) o
derivados (apomórficos), y al aplicarlos en una serie de datos, si un carácter
apomórfico se presenta en más de un taxón, se denomina como sinapomórfico, pero
cuando la apomorfia se presenta en uno solo, es llamado autopomórfias. Por
consiguiente se puede decir que las sinapomorfias son atributos derivados,
compartidos y heredados de un ancestros común inmediato, además son las
importantes en el reconocimiento de los grupos monofiléticos (Schuh, 2000).
Finalmente, los caracteres pueden ser de dos tipos: binarios (doble estado) los cuales
presentan un solo estado derivado, y los multiestado que presentan más de un estado
La correcta identificación e interpretación de los diferentes conceptos y sus atributos
nombrados anteriormente, es de suma importancia en el momento de la construcción
de datos y el análisis de resultados, ya que la base para los estudios filogenéticos esta
dado por la selección de los caracteres y los taxones.
2.3.2 Cladogramas
A través del proceso de codificación de la información dada por los caracteres se
puede realizar un patrón de distribución de los mismos, a modo de representación
grafica denominado cladograma, este se caracteriza por presentar ramificaciones que
reflejan la jerarquía en las sinapomorfias, es decir las similitudes, más no las
diferencias, definiendo los grupos, la configuración y los niveles de jerarquía
(Galindo, 2004).
En el análisis cladístico, los cladogramas presentan las sinapomórfias ordenadas
dentro de una jerarquía anidada, a partir del criterio de parsimonia, escogiendo el
arreglo de los taxones que incluya el mayor número de caracteres explicados de la
manera más simple (Kitching, 1998). Este criterio se puede medir con la longitud
(número de pasos) que representa los cambios en los estados de carácter; así, cuando
los cladogramas tienen una menor longitud se asume un menor número de
homoplasias, es decir una hipótesis más parsimoniosa (Lipscomb, 1998), aunque
cuando el análisis genera múltiples cladogramas o si son pocos pero igualmente
parsimoniosos, la técnica de consenso constituye una herramienta más útil para la
escogencia de hipótesis permitiendo evaluar los resultados (Schuh, 2000).
Todo análisis necesita un fundamento matemático, por esto, los cladogramas cuentan
con una estadística descriptiva que se ha desarrollado con el fin de mostrar las
homoplasia requerida por un cladograma, esto se logra con el índice de consistencia
(IC) que permite medir la cantidad relativa de homoplasias en un cladograma, y el
partir de un grupo de datos, que es retenida como sinapomorfia en el cladograma
(Lipscomb, 1994; Schuh, 2000).
2.3.3 Método de comparación por grupo ajeno
Los estudios cladísticos utilizan las sinapomorfias para buscar grupos hermanos
dentro de un conjunto de taxones, lo cual es posible por medio de dos criterios: uno
llamado directo u ontogenético, y otro indirecto que implica el uso de grupos ajenos,
este ultimo tiene como utilidad el comparar un grupo de estudio (grupo propio) con
otro por fuera de él (grupo ajeno) y éste actúa como un “reemplazo” de los ancestros,
ante la imposibilidad de identificarlos de manera real, es decir ayuda a resolver y
determinar la polaridad de caracteres binarios, aplicando las siguientes reglas
especificas: si uno de los estados de carácter se encuentra tanto en grupo propio como
en grupo ajeno, se considera como primitivo; pero si el otro estado sólo esta en el
grupo propio, es considerado como derivado; y en los casos donde ambos estados se
encuentran tanto en el grupo propio como en el ajeno no es posible definir la
polaridad (Wiley, 1991; Hoyos, 1994).
Al usar métodos de comparación como este (que evalúa los datos bajo el criterio de
parsimonia) para la reconstrucción filogenética, buscando elaborar hipótesis de
parentesco y relaciones de grupos hermanos, el mejor camino es usar múltiples
grupos ajenos simultáneamente, debido a que excluye la posibilidad de hacer una
mala escogencia de un grupo ajeno, aumenta el nivel de generalidad y es una buena
herramienta para comprobar la monofilia del grupo propio, cuando por alguna razón
existen dudas al respecto (Barriel et al, 1998). Aunque, hay que tener en cuenta que el resultado depende de la escogencia del grupo ajeno inicial usado en el momento de la
realización del cladograma.
El análisis de comparación con grupo ajeno, puede hacerse por medio de diferentes
programas, para este trabajo se utilizó el programa de computador Winclada - NONA
fundamentales y formar un “único cladograma general” (aunque en algunas ocasiones
pueden ser más de uno), minimizando así la aceptación de presunciones (Hoyos,
1994).
3 FORMULACIÓN DEL PROBLEMA Y JUSTIFICACIÓN
3.1 Formulación del problema
En la proposición de hipótesis para aclarar relaciones filogenéticas, ha sido utilizada
como herramienta, la aplicación de caracteres anatómicos internos (osteológicos) con
buenos resultados, por esto, se consideró como un buen elemento, el cual proporciona
evidencias que pueden ser útiles para el acercamiento y aclaración en la filogenia,
como en el caso del grupo flavencens del género Atelopus, sugerido como monofilético por Lynch (1993), basándose en la reducción del número de falanges del
pulgar (formula 1-2-3-3).
Sin embargo, posteriormente no se han hecho publicaciones formales referentes a este
tema, manteniéndose hasta hoy, la reorganización planteada por Lynch (1993) para
los otros grupos de especies del género como simples unidades fenéticas, por
consiguiente el grupo ignensces no ha sido posible verificarlo como una unidad monofilética basada en una posible sinapomorfia, además que algunas de estas
presentan cierto conflicto sistemático, lo que generó para el desarrollo de este trabajo,
la escogencia de especies pertenecientes a este grupo.
3.2 Pregunta de investigación
Se formulo la siguiente pregunta, con base en lo anterior:
3.3 Justificación de la investigación
La existencia de vacíos informativos en cuanto a la herpetofauna en Colombia es muy
notoria y lamentable, considerado que es conocido como uno de los países de mayor
diversidad para este grupo, y no es sorpresivo que este reclamando estudios en
diferentes niveles (ecológicos, taxonómicos, filogenéticos) que generen conocimiento
de algunas especies y hasta que pueda ayudar a su mantenimiento y conservación.
Además, los estudios basados en la anatomía de las especies, siguen siendo
importantes y necesarios, ya que son usados por diversas y numerosas razones entre
las que se destacan el proveer evidencias para entender los procesos y productos de la
biología evolutiva, hacer predicciones, y ayudar a contribuir en resolver ciertas
relaciones de parentesco, como es el caso de este trabajo el cual tiene gran
importancia debido a que las especies estudiadas no poseen mucha información de su
filogenia en general y de la anatomía de algunas en particular.
Así, con este estudio se pretende progresar en el conocimiento que se posee acerca de
las especies de interés, resaltando el manejo de los caracteres osteológicos,
reconociendo que aunque en la morfología interna la variación es mínima, permite
hacer comparaciones que puede comprender las relaciones evolutivas tanto en un
género como en niveles superiores; por esto estas diferencias necesitan ser descritas y
evaluadas en el contexto filogenético para así lograr un mejor acercamiento y
entendimiento de la evolución y sistemática intragénerica del grupo (Lötters, 1996).
Por último, con los resultados obtenidos se pueden establecer futuras comparaciones
que puedan proponer hipótesis que a la luz de evidencias, continúen con la búsqueda