ERIKA LISETH VELANDIA BAUTISTA Código: 20082150056
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS FACULTAD DE CIENCIAS Y EDUCACIÓN
PROYECTO CURRICULAR DE LICENCIATURA EN QUÍMICA BOGOTÁ D.C.
ERIKA LISETH VELANDIA BAUTISTA
TRABAJO DE GRADO PARA OPTAR ELTÍTULO DE LICENCIADA EN QUÍMICA
DIRECTOR
CARLOS ARTURO GUERRERO FONSECA MD., MSc., PhD.
UNIVERSIDAD NACIONAL DE COLOMBIA
CODIRECTORA ADIS AYALA FAJARDO MSc
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS FACULTAD DE CIENCIAS Y EDUCACIÓN
PROYECTO CURRICULAR DE LICENCIATURA EN QUÍMICA BOGOTÁ D. C.
“La enseñanza que deja huella no es la que se hace de cabeza a cabeza, sino de corazón a corazón” Howard G. Hendricks
Este trabajo de investigación fue realizado gracias a la confianza que depositó una de las mejores docentes que he tenido, quien con su amor, entrega y dedicación originó en mí, no solo el gusto por la bioquímica sino que lo llevo más allá, lo llevo a una pasión y amor. Gracias profe por la motivación de emprender un camino por la bioquímica.
aquellas que con su aporte fue posible terminar esta investigación.
Agradezco a mis padres Miguel y Edilma quienes me dieron la vida, les doy gracias por su entrega, comprensión y apoyo en todas las decisiones que he tomado, por siempre estar conmigo e impulsarme hacia un futuro lleno de oportunidades y logros que alcanzar.
A mis hermanos Sandra y Jackson y a mi cuñado José por la colaboración en mi formación académica y personal y por forjar un carácter que me ha promovido a seguir adelante.
A mis sobrinos Miguel Ángel y Juliana quienes me levantan en mis decaídas con simplemente una sonrisa, un abrazo y un te quiero.
A mis profesores, compañeros y amigos de la Universidad Distrital por contribuir en mi formación disciplinar y humana. A Laura y Jennifer quienes nunca me dejaron desfallecer y siempre me dieron una palabra de aliento para seguir adelante y terminar esta investigación. A la profesora Adis Ayala por su formación en el área de Bioquímica y Biología Molecular y por su apoyo y colaboración en los arreglos de este escrito.
A la profesora Yurany Moreno quien creyó en mí y me abrió las puertas del mundo de la investigación ofreciéndome la posibilidad de realizar mi trabajo en la línea de investigación de Bioquímica y Biología Molecular.
Y en especial agradecimiento al Doctor Carlos Guerrero director del laboratorio de Biología Molecular de Virus de la Universidad Nacional de Colombia, quien con su entrega, apoyo, enseñanzas, dedicación, regaños y comprensión fue posible iniciar, desarrollar y culminar esta investigación. A mis compañeros del laboratorio, en especial a Natalia con quien inicie este bello camino, a Liliana, Rafael, José Luis y Wilder doy gracias por su ayuda y enseñanzas. De ellos aprendí grandes y valiosas cosas, no solo en la parte investigativa sino en la humana.
ABSTRACT
RESUMEN ... 11
ABSTRACT ... 7
INTRODUCCIÓN ... 1
1. ANTECEDENTES ... 3
2. JUSTIFICACIÓN ... 5
3. OBJETIVOS ... 7
3.1. GENERAL ... 7
3.2. ESPECÍFICOS ... 7
4. MARCO TEÓRICO ... 8
4.1. Virus oncolíticos ... 8
4.2. Reglamentación y consideraciones al realizar estudios para proponer a un virus como oncolítico. ... 12
4.2.1. Selectividad ... 12
4.2.2. Variantes moleculares ... 13
4.2.3. Selección de modelos animales y limitaciones asociadas a ellos ... 13
4.2.4. Farmacología-prueba de concepto (POC) ... 13
4.2.5. Biodistribución ... 13
4.2.6. Consideraciones para la excreción viral ... 13
4.2.7. Toxicología y estudios de seguridad terapéutica ... 14
4.2.8. Estudios de buenas prácticas de laboratorio (BPL) ... 14
4.2.9. Estudios clínicos ... 14
4.2.10. Farmacocinética, farmacodinamia y actividad biológica ... 14
4.2.11. Bioseguridad ... 14
4.2.12. Principios éticos generales ... 14
4.3. Rotavirus ... 15
4.3.1. Estructura ... 15
4.3.2. Genoma ... 15
4.5. Proteínas de choque térmico ... 22
4.5.1. Proteínas de choque térmico y virus. ... 23
4.5.2. Familias de Hsp ... 25
5. METODOLOGIA ... 32
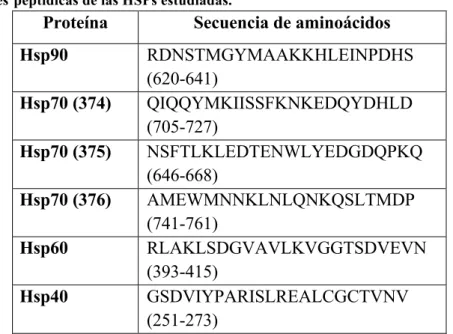
5.1. Secuencias de los peptidos sinteticos de cada Hsp. ... 32
5.2. Rotavirus WTEW ... 32
5.3. KATO III ... 34
5.4. Expresión de las HSPs en la membrana citoplasmática de las células KATO III. ... 34
5.5. Aislamiento de fracciones enriquecidas de membrana citoplasmática (FEMC). ... 34
5.6. Ensayos de evaluación de la interacción entre las HSPs con el rotavirus. ... 35
5.6.1. Ensayo de interacción. ... 35
5.6.2. Ensayos de Bloqueo ... 36
5.7. Papel de las HSPs durante el proceso infeccioso del rotavirus WTEW. ... 38
5.7.1. Ensayo de bloqueo. ... 38
5.7.2. Ensayos de competencia ... 39
5.8. Técnicas empleadas ... 40
5.8.1. ELISA de captura. ... 40
5.8.2. Inmunocitoquímica. ... 41
5.9. Análisis estadístico ... 41
6. RESULTADOS ... 42
6.1. Las proteínas de choque térmico (Hsp90, Hsp70, Hsp60 y Hsp40) se expresan en la membrana citoplasmática de las células KATO III. ... 42
6.2. Las proteínas de choque térmico (Hsp90, Hsp70, Hsp60 y Hsp40) de la membrana citoplasmática de las células KATO III interaccionan con el rotavirus WTEW. ... 43
6.3. La unión del rotavirus a las fracciones enriquecidas de membrana disminuye al ser bloqueadas con anticuerpos anti-HSPs. ... 44
8. CONCLUSIONES ... 64
9. PROYECCIONES DE LA INVESTIGACIÓN ... 64
BIBLIOGRAFIA ... 65
LISTA DE FIGURAS
Figura 1. Diagrama de la replicación de oncovirus en células sanas y células de cáncer……9 Figura 2. Estructura del rotavirus………...15 Figura 3. Ciclo replicativo de Rotavirus………20 Figura 4. Citometría de flujo e inmunofluorescencia de las células KATO III…………...41 Figura 5. Interacción entre el rotavirus WTEW y las proteínas de choque termico expresadas en la membrana de las KATO III………....42 Figura 6. Disminución de la interacción Disminución de la interacción entre HSPs-WTEW al preincubar membrana enriquecida con anticuerpos anti-HSPs. ELISA de captura………44 Figura 7. Disminución de la interacción rotavirus-HSPs cuando las fracciones de membrana son incubadas previamente con anti-HSPs de la porción F(ab’)2………...46 Figura 8. Los péptidos sintéticos de las regiones diferenciadas de las HSPs no alteran la interacción del rotavirus WTEW a las fracciones de membrana. 48
LISTA DE TABLAS
ABREVIATURAS Ac: Anticuerpo.
ADP: Adenosina difosfato.
AdV: Virus de la enfermedad aleutiana. AEC: Aminoetilcarbazol.
AKT: Proteína quinasa B. AS: Ácido sialico.
ATCC: American type culture collection. ATP: Adenosina trifosfato.
BLP: Buenas prácticas de laboratorio. BSA; Albumina de suero bovino. CD: Cytosine deaminase.
CEA: Carcinoembryonic antigen. CRC: Colorectal cancer.
DLP: Partícula de doble capa.
DMEM: Medio de cultivo celular Dulbecco’s modified Eagle’s médium. DNA: Ácido desoxirribonucleico.
dRNA: RNA de doble cadena.
ELISA: Enzyme linked immuno sorbent assay. ECwt: Cepa de rotavirus murino.
F(´ab)2: Porción del anticuerpo que se une al antígeno. Fc: Porción extrema del anticuerpo.
FEMC: Fracciones Enriquecida de Membrana Citoplasmática. FITC: Isotiocianato de fluoresceína.
GFP: green fluorescent protein.
GM-CSF: granulocyte-macrophage colony-stimulating factor. HCC: hepatocellular carcinoma.
h.p.i.: Horas post infección. HSFs: Factores de shock térmico.
Hsp40: Proteína de choque térmico 40. Hsp60: Proteína de choque térmico 60. Hsp70: Proteína de choque térmico 70. Hsp90: Proteína de choque térmico 90. HRP: Horseradish peroxidase.
HSPs: Proteínas de choque térmico. HTLV-1: Virus humano T-linfotrópico.
ICH: International conference on harmonisation. ICP: infected cell protein.
mtHsp70: Proteína de choque térmico 70 mitocondrial. MOI: Multiplicidad de infección.
MV: Measles virus.
NDV: Newcastle disease virus. NIS: Sodium iodide symporter. NSP: Non structural protein. NTP: Nucleótido trifosfato. NTPasa: Nucleótido trifosfatasa.
OMS: Organización Mundial de la Salud. OPD: O-fenilendiamina dihidrocloruro. PDI: Proteína disulfuro isomerasa. PMSF: Fluoruro de fenil metil sulfonilo. PS: Péptido sintético.
PVDF: Difluoruro de polivinilo. RE: Retículo esdoplasmático. RNA: Ácido ribonucleico.
RNAi: Ácido ribonucleico interferente.
SCCHN: squamous cell carcinoma of the head and neck. SDS: Dodecil sulfato de sodio.
SDS-PAGE: Electroforesis en gel de poliacrilamida dodecil sulfato de sodio. siRNA: Ácido ribonucleico silenciador.
TK: thymidine kinase. TLP: Partícula de triple capa.
TRUY: Aislamiento rotaviral (mezcla de cepas TRF, RRV, UK y YM). VGF: vaccinia growth factor.
VP: Viral protein.
Wt1-5: Aislamiento rotaviral (mezcla de cepas aisladas de heces humanas). WTEW: Aislamiento rotaviral (mezcla de cepas WWM, TRUY, ECwt y Wt1-5). WWM: Aislamiento rotaviral (mezclar de cepas Wa, Wi y M69).
αVβ3: Integrina alfa cinco beta tres.
RESUMEN
En el laboratorio de Biología Molecular de Virus de la Universidad Nacional de Colombia se está investigando si rotavirus WWM, TRUY, Wt1-5, O-ECwt y WTEW (generados en el laboratorio), pueden utilizarse como virus oncolítico. Trabajos previos sugieren que los aislamientos rotavirales infectan las líneas celulares tumorales Sp2/0- Ag14 (mieloma murino), U937 (linfoma histiocitico), REH (leucemia linfocítica aguda), PC3 (adenocarcinoma grado IV de próstata). MDA y MCF-7 (adenocarcinoma de mama)
Este trabajo tuvo por objetivo determinar si hay o no interacción del rotavirus WTEW con las proteínas de choque térmico Hsp90, Hsp70, Hsp60, Hsp40 durante la entrada a la línea tumoral KATO III (carcinoma de estómago). Para esto se aislaron fracciones enriquecidas de membranas citoplasmáticas, se incubaron con rotavirus o se bloquearon con anticuerpos específicos anti-Hsp (completos o la porción F(ab’)2) y se adicionó rotavirus MOI 8. La interacción se observó mediante técnicas de ELISA de captura, coinmunoprecipitación y Western Blot. Para determinar el papel funcional de las HSPs, las células se incubaron con anticuerpos F(ab’)2 y luego con rotavirus por 12h a 37°C. El rotavirus se incubó con proteínas HSPs completas solubles durante 1h a 37°C y luego el complejo HSPs-rotavirus se adicionó a las células por 12h a 37°C. La infección en ambos casos se evaluó mediante inmunocitoquímica. Finalmente se incubaron las células con el rotavirus WTEW por 1 h a 4°C y luego se le adicionaron los péptidos sintéticos de cada Hsp por 1h a 4°C dejando por 12 h a 37°C en cámara de CO2. La infección se evaluó por ELISA de captura. Se determinó que existe interacción entre rotavirus WTEW con las HSPs de manera directa y funcional ya que al bloquear o alterar dicha unión se disminuye la infección, lo que sugiere que el rotavirus WTEW interacciona con las HSPs de la línea KATO III facilitando el ingreso del rotavirus a la célula.
ABSTRACT
In the Molecular Biology of Virus located at Universidad Nacional de Colombia, the research to determine whether rotavirus WWM, Wt1-5, O-ECwt and WTEW (generated in the laboratory) can be used as oncolytic virus. Preliminary research suggests that rotaviral isolates infected tumor cell lines Sp2/0-Ag14 (murine myeloma), U937 (histiocytic lymphoma), REH (Acute Lymphocytic Leukemia), PC3 (prostate adenocarcinoma grade IV). MDA and MCF-7 (breast adenocarcinoma).
This study was aimed to determine whether there was or not WTEW rotavirus interaction with heat shock protein Hsp90, Hsp70, Hsp60, Hsp40 during the entry to the tumor cell line KATO III (gastric carcinoma). For this, fractions enriched with cytoplasmic membranes were isolated, incubated with rotavirus or blocked with specific anti-Hsp (complete or portion F(ab')2) and was added rotavirus MOI 8. The interaction was observed by capture ELISA, co-immunoprecipitation and western blotting. To determine the functional role of HSPs, the cells were incubated with antibody F(ab')2 and then with rotavirus for 12 h at 37 °C. Rotavirus was incubated with soluble proteins HSPs complete for 1 h at 37 °C and then the HSPs rotavirus complex was added to the cells for 12 h at 37 °C. In both cases the infection was assessed by immunocytochemistry. Finally cells rotavirus WTEW incubated for 1 h at 4 °C and then were added the synthetic peptides each Hsp for 1 h at 4 °C leaving 12 h at 37 °C in CO2 chamber. Infection was evaluated capture ELISA. It was determined that there is interaction between the rotavirus WTEW and HSPs was direct and functional manner as to block or alter binding said infection is decreased, suggesting that interacts rotavirus WTEW and HSPs of facilitating the entry of rotavirus to the cell KATO III line.
INTRODUCCIÓN
La palabra “Cáncer” designa un grupo de enfermedades que pueden afectar cualquier parte del organismo, se conoce también como «tumores malignos» o «neoplasias malignas». Una característica del cáncer es la multiplicación rápida de células anormales que se extienden más allá de sus límites habituales y puede invadir partes adyacentes del cuerpo o propagarse a otros órganos, este proceso es conocido como metástasis, principal causa de muerte por cáncer. La proliferación celular incontrolada es el sello distintivo del cáncer y las células tumorales generan daño a los genes que regulan directamente sus ciclos (1). Según la OMS en su informe de febrero de 2013, el cáncer es una de las principales causas de muerte en todo el mundo; en 2012 causó 8,2 millones de defunciones siendo los de mayor mortalidad por año el cáncer de pulmón, estómago, hígado, colon y mama (2).
Para combatir el cáncer, se han desarrollado tratamientos como la quimioterapia y radioterapia con el fin de controlarlo. Sin embargo, los efectos secundarios de estos procedimientos suelen ser desde moderados hasta graves, lo cual obedece al tipo de paciente y otros factores, resultando desde afecciones cutáneas, vomito, caída del cabello, debilitamiento, problemas de coagulación hasta alteraciones de los sentidos, del funcionamiento de ciertos órganos y del sistema inmune, producto de los efectos tóxicos en las células normales, además son tratamientos que no son totalmente efectivos y no son definitivos (OMS, 2013). Gracias al progreso de la biología molecular se ha avanzado en el desarrollo de nuevas técnicas de tratamiento contra el cáncer como la viroterapia que emplea virus para controlar y hasta eliminar el tumor presente (3). El propósito de este procedimiento con virus oncolíticos es destruir las células cancerígenas o utilizarlo como coadyuvante en otros tratamientos establecidos como la quimioterapia y radioterapia (4).
Los virus empleados en la viroterapia se pueden dividir en dos categorías: con efecto oncolítico selectivo, como el virus de la enfermedad de Newcastle (NDV), reovirus y herpesvirus y virus modificados genéticamente de manera que adquieren una mayor selectividad por las células tumorales con capacidad de replicación y lisis en su interior. Se conoce que los virus más destacados clínicamente son el virus de la enfermedad de Newcastle (NDV) y el reovirus, entre los virus de ARN, y el vaccinia, herpes simplex (HSV) y adenovirus, entre los virus de ADN, siendo su principal tarea la de adherirse a la célula blanco mediante receptores específicos para cada virus, penetrar, hospedarse y replicarse solamente en las células tumorales induciendo la apoptosis o la lisis celular y posiblemente liberando viriones que no infecten el tejido sano (5-6).
aislamientos rotavirales con las proteínas de choque térmico (Hsp) expresadas en células que sufren estrés, tal es el caso de las células de cáncer que se encuentran en constante estrés celular como las de linfoma U-937 (7-9) y leucemia REH (9) por lo que se consideran como las células apropiadas para ser tratadas con este virus lítico.
1. ANTECEDENTES
La actividad oncolítica de los virus se dió a conocer por Dock y DePance quienes reportaron casos de regresiones tumorales utilizando los virus de influenza y rabia pero por las limitaciones tecnológicas estos estudios no pudieron ser continuados. A finales del siglo pasado y gracias al desarrollo de la biología molecular, se retomó el estudio del tratamiento del cáncer empleando virus, consiguiendo así, el desarrollo de la viroterapia que emplea virus denominados oncolíticos que infectan células cancerígenas pero no células normales (10-11).
En el laboratorio de Biología Molecular del Virus de la facultad de Medicina de la Universidad Nacional de Colombia se han realizado varios estudios para determinar el papel oncolítico del rotavirus en diferentes líneas celulares de carcinoma como lo describe Guerrero 2011 en su trabajo investigativo, con el cual logró establecer el potencial del rotavirus como virus oncolítico en un modelo celular in vitro de mieloma murino utilizando una metodología dividida en tres etapas: 1) Generación de los aislados rotavirales a evaluar, 2) Determinación de la expresión de las proteínas Hsp90, 70, 60, 40, Hsc70, integrina αVβ3 y PDI en la línea celular de mieloma murino Sp2/0 Ag14 y, 3) Caracterización de la susceptibilidad de la línea celular ante la infección por rotavirus aislados y seleccionados en este laboratorio. Las técnicas que empleó para desarrollar estas metodologías fueron inmunocitoquímica e inmunfluorescencia. Con los resultados obtenidos, el investigador concluyó que los 5 rotavirus aislados (ECwt, WTEW, TRUY, WWM y Wt1-5) infectan y lisan la línea tumoral de mieloma murino Sp2/0-Ag14 y desencadenan cambios en la expresión de las proteínas celulares Hsc70, Hsp90 y Hsp70, además sugiere que los aislamientos de rotavirus estudiados son potenciales virus oncolíticos (12).
Alfonso y Rodríguez con su trabajo realizado en el 2012, lograron determinar la unión de los aislamientos rotavirales Wt 1-5, TRUY, WWM y ECwt y WTEW con las proteínas de choque térmico Hsp90 y Hsp70 durante el proceso infeccioso en la línea celular de leucemia mieloide humana U937 mediante ensayos de inmunocitoquímica, inmunofluorecencia, ELISA de captura, co-inmunoprecipitacion y western blot. Los investigadores concluyeron que el rotavirus usa como moduladores de entrada este tipo de proteínas superficiales que podrían estar acopladas o trabajando en conjunto (8).
En la investigación realizada por Hernández, 2013, se evaluó la interacción entre el rotavirus WTEW y las proteínas de choque térmico en las líneas celulares de linfoma U- 937 y leucemia REH mediante ensayos de unión como ELISA de captura y co- inmunoprecipitación. También estableció la entrada del virus por estas proteínas expresadas en la membrana celular empleando ensayos de competencia usando fracciones enriquecidas de HSPs solubles o péptidos sintéticos derivados de HSPs preincubados con WTEW. Además empleó anticuerpos específicos contra dominios de Hsp (Hsp90, Hsp70, Hsp60 y Hsp40) buscando bloquear la infectividad. Con los resultados obtenidos pudo concluir que las proteínas de choque térmico presentes en la membrana citoplasmática se unen a rotavirus WTEW durante el proceso de entrada a las líneas celulares U-937 y REH (9).
Patiño y Pedroza (2013) determinaron el potencial oncolítico de los cinco aislamientos rotavirales (Wt 1-5, TRUY, WWM y ECwt y WTEW) en la línea celular Sp2/0 Ag14 y la disminución de la infección al ser bloqueada con anticuerpos anti-HSPs. Mediante la técnica de inmunocitoquimica evaluaron las unidades formadoras de foco (partículas infecciosas), por la técnica de yoduro de propidio y TUNEL determinaron los efectos citotóxicos de los cinco aislamientos y por ultimo evaluaron mediante inmunocitoquimica la participación de las HSPs en el proceso de penetración del rotavirus a la célula. Los resultados encontrados sugirieron que los cinco aislamientos rotavirales infectan y lisan la línea celular tumoral Sp2/0 Ag14 y utilizan las proteínas de choque térmico expresadas en membrana cuando ingresan a la célula. Las investigadoras concluyeron que los cinco rotavirus estudiados pueden ser potenciales virus oncolíticos (13).
2. JUSTIFICACIÓN
El cáncer es una de las primeras causas de mortalidad en el mundo. Para el año 2012 se diagnosticaron cerca de 14,1 millones de nuevos casos y se presentó una mortalidad de 8,2 millones (2). Gracias a los desarrollos tecnológicos se han generado tratamientos y se han estudiado nuevas alternativas de combatir esta enfermedad. A parte de los tratamientos ya conocidos, existen desarrollos a nivel investigativo que emplean variados elementos biológicos que pueden dar una luz que promueva la curación del cáncer o la mejora de tratamientos que no generen tantos ni tan graves efectos secundarios. Tal es el caso de los virus, cuya propiedad de invasividad e infectividad, que es ávida de interacción con las proteínas celulares, los hace candidatos promisorios para el tratamiento de dicho padecimiento (4).
En la actualidad se han realizado varios estudios en viroterapia con el fin de postular diferentes virus como oncolíticos y así servir como terapia complementaria a tratamientos de cáncer como la radioterapia, quimioterapia y cirugía, entre otros ya existentes. Los virus oncolíticos deben sufrir modificaciones para que ataquen la célula blanco, es decir, para que se adhieran, penetren, repliquen, destruyan las células de cáncer y se propaguen a otras células o tejidos cercanos que presenten las mismas características de las células objetivo. Una de las principales ventajas de esta terapia vírica según Merchán et al, es su selectividad hacia las células tumorales y en combinación con otro tipo de tratamiento como la quimioterapia, es la destrucción de células tumorales en gran cantidad y por ende la reducción del tamaño del tumor. Los virus son modificados genéticamente, ya sea realizando una deleción de un gen como en el caso de ONYX 015 (Adenovirus) o de OncoVEXGM-CSF® (Virus de Herpes simple) o JX-594 (Virus de Vaccinia) o modificando los genes, en el caso de Reolysin® (Reovirus) alterando su genoma para que aproveche de la vía Ras (14). Pero la exclusividad de estos virus oncoliticos para atacar las células tumorales presenta una limitante para tratar diferentes tipos de cáncer ya que existe diversidad entre las células del mismo cáncer y por tanto en los diferentes tipos de cáncer, es decir, estos virus solo pueden tratar ciertos tumores malignos.
Las proteínas de choque térmico (Hsp) se encargan de reparar daños estructurales de las proteínas y funcionan como citoprotectoras cuando la célula está sometida a estrés. Las células tumorales, al encontrarse en ambientes hostiles, expresan las proteínas HSPs que favorecen su supervivencia y su posterior diseminación a todo el organismo. Este mecanismo es aprovechado en investigación con la finalidad de que el rotavirus continúe su curso natural como virus lítico, destruyendo las células de carcinoma (15).
3. OBJETIVOS
3.1. GENERAL
Determinar si se da interacción del rotavirus WTEW con las proteínas de choque térmico Hsp90, Hsp70, Hsp60, Hsp40 de la línea tumoral KATO III.
3.2. ESPECÍFICOS
3.2.1. Evaluar el tipo de interacción entre el rotavirus WTEW y las proteínas de choque térmico en la línea tumoral KATO III.
4. MARCO TEÓRICO
El cáncer ha sido una de las enfermedades más estudiadas en los últimos tiempos ya que los tratamientos convencionales como la radioterapia y quimioterapia no solo atacan las células objetivo “que en estos casos son las cancerosas” sino también las células sanas. En la quimioterapia se pueden emplear diferentes tipos de medicamentos como los que bloquean la capacidad de reproducción de las células en general, es decir, también afectan las células sanas de cabello, garganta y otras que constantemente se regeneran, además se sabe que este tratamiento reduce el número de glóbulos blancos volviendo susceptible al paciente a contraer fácilmente alguna enfermedad. En el caso de la radioterapia, se emplean diferentes tipos de radiación que atacan mayormente a las células de cáncer que son más sensibles que las células normales pero que igualmente son afectadas con la diferencia de que estas últimas tienen capacidad de recuperarse. Estos tratamientos generan efectos secundarios que incluyen náuseas, vómito, diarrea, caída del pelo, cansancio, probabilidad de contraer una infección, fatiga, cambios en la piel y pérdida del apetito (16). Estos tratamientos no han sido suficientes para controlar la enfermedad, por este motivo se vio necesario desarrollar procedimientos alternos que contribuyan a disminuir la incidencia del cáncer.
4.1. Virus oncolíticos
Figura 1. Diagrama de la replicación de oncovirus en células sanas y células de cáncer. El virus ataca y penetra las células tumorales y normales, pero la infección en la célula normal es evitada por la misma célula ya que tiene la habilidad de reconocer y destruir el material genético anormal. Como resultado del tropismo natural hacia el receptor especifico de la célula tumoral, se producen viriones que lisan la célula liberándose así, las nuevas partículas virales que infectan células de las mismas características de las que infectaron. Tomado y modificado de Alemany. R. (2013) (18).
Estos virus atacan específicamente a las células tumorales por los receptores que éstas sobreexpresan y/o mediante la explotación de vías moleculares de señalización, asociados con la transformación maligna para su replicación generando efectos citotóxicos y aumentando la lisis de las células cancerígenas. Este proceso es más eficiente si se incorporan transgenes terapéuticos o inmunomoduladores en los virus. Se ha comprobado que la infección de virus oncolíticos a tumores tiene menos efectos secundarios tóxicos que los otros tipos de tratamiento empleados para combatir el cáncer (21).
Se emplean virus de tipo salvaje con efecto oncolítico selectivo como los reovirus, parvovirus y virus herpes; y virus modificados genéticamente con el fin de aumentar la selectividad por las células tumorales, la capacidad de replicación y lisis celular sin afectar el tejido sano adyacente. Se puede aumentar la selectividad del virus por la célula tumoral mediante la modificación del genoma para que las proteínas de la cápside no reconozcan las células sanas o para que su replicación solo tenga lugar en el interior de la célula tumoral alterando la transcripción en la célula diana (22-23).
herpes simplex tipo 1 (G207 y C1716), entre otros, no son tóxicos y son oncolíticos (Tabla 1) (27-29).
Tabla 1. Virus estudiados para ser propuestos como virus oncoliticos.
VIRUS NOMBRE MODIFICACIÓN TUMOR
Adenovirus Onyx-015 E1B-55k-E3B Metástasis pulmón. Oncorine E1B-55k-E3 Cáncer de cabeza y cuello. Virus del herpes
simple
Oncovex GM-CSF Tumores sólidos.
ICP34.5 Melanoma. ICP47 (-) Melanoma
Sarampión MV-CEA CEA Cáncer de ovario y glioma.
MV-NIS NIS Mieloma, cáncer de ovario,
mesotelioma y SCCHN. Virus de la
enfermedad de Newcastle
NDV-HUJ - Glioma.
PV701 - Tumores sólidos.
MTH-68/H
- Tumores sólidos.
NV1020 - Tumores sólidos.
Reovirus Reolysin - Glioma.
Cáncer peritoneal, tumores sólidos y CRC.
Cáncer de páncreas, SCCHN, melanoma, cáncer de pulmón.
SCCHN. Retrovirus Toca 511 CD Glioma. Vaccinia
(Wyeth strain)
JX-594 GM-CSF CRC
TK (-) Tumores sólidos, HCC, Tumores sólidos pediátricos, melanoma. Melanoma y CRC.
HCC y CRC. Vaccinia
(Western Reserve)
wDD-CDSR
TK-, VGF- Tumores sólidos
Vaccinia (Lister)
GL-ONC1 Renilla luciferasa Tumores sólidos
GLV-h68 GFP, β-gal SCCHN
El virus del herpes fue el primer virus oncolítico aprobado en pacientes con glioma maligno ya que tiene una gran capacidad para replicarse en tejido neuronal. Entre 1997 y 1999, se analizó la dosis, la seguridad del tratamiento y la replicación selectiva del virus HSV-1716 Y G207 en el tratamiento de tumores malignos en el cerebro (30).
Por otra parte, el potencial oncolítico del reovirus se empezó a estudiar en 1977, cuando se determinó la susceptibilidad del virus por células cancerígenas más no por las células normales (31). Posteriormente, se reportó que al infectar células WI-38 (fibroblastos de pulmón humano embrionario) con reovirus (que normalmente no son susceptibles) junto con “antígeno T mayor de SV-40”, el virus se replicaba (32).
Tabla 2. Los oncovirus en fase más avanzada. VECTOR/
VIRUS
CANCER TRANSGÉ N FASE DE DESARROLLO Adenovirus p53 (Gendicine®) Tumores sólidos, sobre todo de cabeza y cuello
p53 Comercializado en
China desde 2003
Adenovirus Oncorine® (H-101) Tumores sólidos, sobre todo de cabeza y cuello Virus oncolítico delecionad o en E1B-55K
Comercializado en China desde 2005
Herpes simplex OncoVEXG M
-CSF
Melanoma Virus oncolítico que expresa GM-SCF
Fase clínica III
Vaccinia virus JX-594 (Pexa- Vec) Carcinoma hepatocelular Virus oncolítico que expresa GM-SCF
Fase clínica III
Tomado y modificado de Alemany. R. (2013) (18).
4.2. Reglamentación y consideraciones al realizar estudios para proponer a un virus como oncolítico.
La International Conference on Harmonisation (ICH) es una organización internacional que se ha encargado de reglamentar las consideraciones y pasos a tener en cuenta cuando se inicia un trabajo con un virus buscando utilizarlo como agente oncolítico. (35).
parte, aunque la selectividad también puede ser demostrada en estudios in vivo no clínicos, siempre hay que tener en cuenta que este aspecto no representa una medida directa del potencial antineoplásico de los virus oncolíticos. Por lo tanto, para posibilitar su uso como agente anticancerígeno, se deben realizar ensayos que arrojen cualquier evidencia de actividad biológica relacionada con la propagación viral en la masa tumoral.
4.2.2. Variantes moleculares: La caracterización de cada uno de los virus oncolíticos incluye la verificación de la existencia de sus correspondientes variantes moleculares, de las cuales se deben tener en cuenta las que resultan de la replicación selectiva o del perfil oncolítico. La naturaleza y la cantidad de variantes determinan la estrategia experimental a seguir para su selección, evaluando la estabilidad genética de los vectores antineoplásicos.
4.2.3. Selección de modelos animales y limitaciones asociadas a ellos: Se debe tener en cuenta, para la selección de un modelo animal, el propósito del estudio, el tropismo viral, la infectividad, la capacidad de replicación, el potencial citopático, el efecto antitumoral del virus oncolítico, susceptibilidad a la infección y al ciclo viral, además de la inhabilidad para controlar la respuesta inmune.
4.2.4. Farmacología-‐prueba de concepto (POC): Evalúa los aspectos relacionados con la bioactividad, el potencial mecanismo de acción y el perfil farmacológico de los virus oncolíticos, es decir, que su capacidad para inducir los efectos biológicos deseados in vivo sea verificable. La importancia de estos estudios radica en que ayudan a definir un intervalo de dosis farmacológicamente activas, estableciendo una que sea óptima y otra mínimamente efectiva, definir una ruta potencialmente adecuada para el suministro de un determinado virus oncolítico y fijar un esquema de dosificación para las fases iniciales de los respectivos ensayos clínicos.
4.2.5. Biodistribución: Se analiza la propagación de los virus oncolíticos en los órganos blanco y en los que no lo son, para lo cual se pueden aplicar ensayos que detecten proteínas virales (inmunocitoquímica), o sondas que identifiquen ácidos nucleicos virales, entre otros.
una potencial manifestación notable de efectos secundarios a largo plazo, tanto en estudios clínicos como en no clínicos.
4.2.7. Toxicología y estudios de seguridad terapéutica: El perfil de expresión de transgenes (si están presentes), y el de biodistribución y persistencia de los virus oncolíticos orientan con frecuencia la duración de los correspondientes estudios toxicológicos y la fijación de los intervalos de tiempo para el sacrificio de los animales. Ante esta situación, que puede ser bastante variable, la evaluación toxicológica de este tipo de vectores debe ser lo suficientemente exhaustiva para identificar, caracterizar y cuantificar el potencial de toxicidad local y sistémica posterior al suministro viral.
4.2.8. Estudios de buenas prácticas de laboratorio (BPL): Los criterios de valoración de seguridad son a menudo obtenidos a partir de ensayos realizados con animales portadores de tumores, de los cuales pueden resultar situaciones únicas del cuidado animal. Para algunos estudios que emplean métodos especializados de prueba puede resultar difícil cumplir plenamente las BPL, haciéndose más evidente esta situación si las leyes regionales exigen su cumplimiento.
4.2.9. Estudios clínicos: Se deben realizar estudios que indiquen la dosis a usar en un paciente para determinar un nivel de dosis inicial seguro. También se debe tener en cuenta el potencial de replicación del virus oncolítico en tejidos que no son su objetivo.
4.2.10. Farmacocinética, farmacodinamia y actividad biológica: Se deben emplear ensayos de infectividad para monitorear la proliferación de los virus oncolíticos. Otros enfoques pueden ser usados para vigilar el ciclo viral de este tipo de partículas, dentro de los que se incluyen los niveles de algún marcador viral o la actividad de un transgen en especial. Es de gran utilidad la información que se obtiene de la patología neoplásica en la medida en que sea posible practicar previamente una resección o una biopsia.
4.2.11. Bioseguridad: Es importante que al administrar tratamientos basados en virus oncolíticos se sigan las precauciones habituales para el manejo de material infeccioso y las pautas de bioseguridad o sus equivalentes. También se deben seguir las regulaciones emitidas por las entidades competentes, los países, los estados, y las localidades. Generalmente, como parte del protocolo clínico, todas las autoridades reguladoras establecen la necesidad de usar algunas formas de barrera protectora durante la duración de los correspondientes ensayos clínicos como una medida estándar para evitar la transmisión persona a persona.
humanos deben tener en cuenta los tres principios éticos básicos: respeto por las personas, beneficencia y justicia. En forma general, se concuerda en que estos principios guían la preparación responsable de protocolos de investigación (36).
4.3. Rotavirus
Se conoce que el rotavirus pertenece a la familia Reoviridae, tiene un genoma ARN bicatenario envuelto por tres cápsides proteícas y no posee envoltura de bicapa lipídica (37) (Figura 2). Este virus ataca los enterocitos maduros de niños menores de 5 años provocando diarrea, en algunas ocasiones vómito y deshidratación severa siendo una de las causas principales de muerte por gastroenteritis ya que el virus lisa las células provocando un desequilibrio iónico en el sistema gastrointestinal (38).
Figura 2. Estructura del rotavirus. Las partículas virales maduras de rotavirus poseen tres capas proteicas; la capa externa conformada por VP4 (color rojo) y VP7 (color amarillo), la capa intermedia conformada por VP6 (color azul) y la capa interna conformada por VP2 (color verde) y VP1 y VP3 (color rojo). Electroforesis del genoma viral y proteínas estructurales o no estructurales de células infectadas (+) o no (-). Tomado y modificado de Angel, J. et al. (2007) (39) y Ruiz et al. (2009) (40)
4.3.1. Estructura
La apariencia del rotavirus es similar a una rueda de 75 nm de diámetro, presenta una estructura icosaédrica conformada en su centro por el genoma del virión rodeado por subunidades polipeptídicas que constituyen las tres cápsides de proteína (41-43).
4.3.2. Genoma
para seis no estructurales (NSP: Non Structural Protein), cada una codificada por un gen excepto para la NPS5 y NPS6 codificadas por un mismo fragmento. (44).
4.3.3. Proteínas estructurales
4.3.3.1. Nucleocápside
Esta capa interna está compuesta en mayor proporción por la proteína estructural VP2 que contiene el material genético del virión y en menor cantidad por la VP1, caracterizada por ser una ARN polimerasa dependiente de ARN, VP3 que es una proteína guanilil transferasa. VP1 y VP3 están encargadas del proceso de transcripción viral (44).
VP1: Esta proteína de 1088 aminoácidos es codificada por el segmento 1 del genoma viral (45). Estudios realizados sugieren que esta proteína es la ARN polimerasa dependiente de ARN porque la secuencia de la proteína contiene regiones que tienen homología con motivos conservados entre la ARN polimerasas dependientes de ARN de otros virus. La VP1 está presente en partículas virales transcripcionalmente activas y se caracteriza por ser un componente común de estas con actividad de ARN polimerasa (transcriptasa o replicasa) (46).
VP2: Conformada por 881 aminoácidos, es codificada por el segmento 2 del genoma y es la más abundante de la cápside interna. Esta proteína no depende de otros productos virales como ARN u otras proteínas; como es el caso de la VP1, pero tiene afinidad por el ARN lo que sugiere que podría estar relacionada con el empaquetamiento y la conformación del genoma del rotavirus (46). Esta proteína es la única que tiene sitios de unión con el material genético del virus dsARN (47).
VP3: Esta proteína de 835 aminoácidos es codificada por el segmento 3 del ARN (48). Al igual que VP1 existen muy pocas copias de este polipéptido por virión. A esta proteina se le ha caracterizado como guanilil-transferasa del ARN viral porque su secuencia es similar a otras guanilil-transferasas estudiadas, se une a GTP de modo covalente en una reacción reversible es un componente esencial de los complejos de transcripción y no de los complejos replicativos (46).
4.3.3.2. Cápside intermedia
interna las partículas de doble capa (DLPs) (49).
VP6: Es la proteína más abundante del virión conformada por 397 aminoácidos y codificada por el gen 6 del dsARN. Esta proteína interacciona con la proteína VP2, hacia el interior de la partícula y con VP4 y VP7 de la capa externa. VP6 es una proteína ampliamente estudiada ya que se ha comprobado que contiene determinantes antigénicos conservados entre todas las cepas de rotavirus del grupo A. La función de la VP6 es estructural ya que mantiene la conformación apropiada y la organización del complejo transcripcional en la nucleocápside del virus (46). Esta proteína es usada para ensayos de diagnóstico para la identificación de partículas virales por su capacidad inmunogénica y antigénica (47).
4.3.3.3. Cápside externa
Está formada por las proteínas VP4 y VP7, que son las responsables de la adhesión y penetración al enterocito y por lo tanto determinan la infectividad del virus, siendo los serotipos de los grupos A al C los que infectan a los humanos (49).
VP4: Esta proteína conformada por 776 aminoácidos es codificada por el gen 4 y se encarga del reconocimiento de los receptores celulares para la adherencia y entrada del virus a la célula huésped.
La infectividad del virus se incrementa cuando es tratado con tripsina ya que produce un rompimiento de VP4 en dos proteínas de menor peso molecular, la VP8 y la VP5 lo que sugiere que la penetración del rotavirus puede ser iniciada por las proteínas generadas o por un cambio conformacional de VP4 (49-50).
VP8: Está conformada por 247 aminoácidos y está involucrada en la unión con el ácido sálico que se encuentra presente en las glicoproteinas (AS).
VP5: La conforman 529 aminoácidos que interaccionan con la integrina α2β1 y HSC70 (51).
4.3.4. Proteínas no estructurales
Las NSPs son sintetizadas en el citoplasma de la célula durante el ciclo replicativo del rotavirus para regular la síntesis de las proteínas virales (VP) (53-54). Se distinguen 6 proteínas no estructurales:
NSP1: Proteína codificada por el gen 5 que se une al ARN.
NSP2: Es codificada por el segmento 7 del genoma rotaviral, es una proteína de unión de ARN necesaria en la replicación del material genético.
NSP3: Codificada por el segmento 8 del genoma ARN, se encarga de finalizar la síntesis proteica de la célula huésped para dar inicio con la viral.
NSP4: Esta proteína es codificada por el gen 10 de dsARN. Se caracteriza por ser una enterotoxina que induce el principal síntoma de infección rotaviral, la diarrea (55-56).
NSP5: Se codifica por el gen 11 del genoma de rotavirus y en las células infectadas se forma en el viroplasma.
NSP6: Codificada por el segmento 11 al igual que la NSP5. Se une al ácido nucleico (56).
Tabla 3. Resumen de las proteínas estructurales y no estructurales de rotavirus y su función.
Proteina del rotavirus
Función
VP1 RNA polimerasa dependiente de RNA; se une a ssRNA y forma complejo con VP3; se localiza dentro de la cápside interna.
VP2 Proteína estructural de la cápside interna; coopera con proteína VP1 en actividad de replicasa.
VP3 Guanidil transferasa y metil transferasa; hace parte del complejo de transcripción del virión junto con VP1.
VP4 Proteína de unión membrana celular; forma de trímeros de VP4 y en su clivaje genera los fragmentos VP5 y VP8.
VP6 Principal componente del virión; tiene estructura homotrimérica; se requiere para la transcripción.
Proteina del rotavirus
Función
NSP1 Asociada al citoesqueleto; parece ser la proteína responsable de la supresión de la respuesta al interferón alfa.
NSP2 NTPasa y helicasa de unión no específica a cadena simple de RNA; está involucrada en la formación del viroplasma; se une a NSP5 y VP1; esencial para la síntesis de dsRNA.
NSP3 Homodímero; se une al región 3’ del RNA viral; Se une al factor de elongación eIF4G1 y está involucrada en la regulación transcripcional.
NSP4 Enterotoxina viral; receptor para la entrada de partículas de doble capa a través de la membrana del retículo endoplasmático; glicoproteína; regula los niveles de calcio y la replicación del RNA viral.
NSP5 Interactúa con NSP2 y NSP6; forma homomultímeros; sufre
O-glicosilación; se une a ssRNA; componente del viroplasma; esencial para a replicación viral.
NSP6 Producto del segundo marco de lectura abierto del segmento del gen II; interactúa con NSP5; hace parte del viroplasma.
Tomado de Angel, J., et al. (2007) (57)
4.4. Ciclo infeccioso del rotavirus
El rotavirus infecta enterocitos maduros de las vellosidades del intestino delgado, entrando a la célula por un proceso coordinado de interacciones con varios ligandos y cambios conformacionales en la proteína VP4 de la cápside del virión que sufre un rompimiento por la tripsina intestinal generando dos polipéptidos VP8 y VP5, los cuales permiten la adhesión a la célula por las interacción con ácido siálico (AS) y con la integrina α2β1 respectivamente, además de la interacción de VP7 con otras proteínas celulares, como Hsc70 y la integrina αvβ3 (58-59).
La replicación viral se da en el citoplasma en un tiempo aproximado de 14 horas pasando por varios procesos:
a) Se adhieren las proteínas VP4 y VP7 con los receptores celulares como las integrinas
α2β1, α4β1, αxβ2, αvβ3 y la proteína HSC 70.
b) Se genera la penetración directa o por endocitosis.
c) Se remueve la cápside externa y se activa la transcripción. d) Se produce el ARN viral.
e) El ARN es sintetizado por la ARN polimerasa viral.
virales estructurales y 5 no estructurales.
g) Se da el ensamblaje de la nucleocápside y del genoma generando el viroplasma. h) Se maduran los DLPs a TLPs: se produce una glicosilación de VP7 en el retículo endoplasmático rugoso y la NSP4 actúa como un receptor intracelular para DLPs, i) Se liberan las partículas infecciosas por lisis celular.
Figura 3. Ciclo replicativo de Rotavirus. El virus se une a los receptores celulares y penetra la celula. En el interior de la celula se activa la transcriptasa viral y los ARNm dirigen la sisntesis de las proteínas virales. Estas proteínas se acumulan en estructuras llamadas viroplasmas donde se replica el ARN viral y se ensamblan las partículas de dos capas (DLPs) para seguir con la morfogénesis en el retículo endoplasmatico (RE). Tomado y modificado de Shane, D., et al, (2012) (60)
4.4.1. Entrada de rotavirus a la célula
El rotavirus entra por de endocitosis, en un proceso que se da en presencia de tripsina; proteasa que cliva VP4 en VP5 y VP8. Por otra parte, la maduración de los endosomas genera cambios como el descenso en el pH y en el estado redox de la célula que permiten a rotavirus sensar su ubicación en la célula y usar esta información para establecer el momento de desensamble. El virus se internaliza en vesículas endocíticas recubiertas de clatrina que presentan un decrecimiento en la concentración de calcio promovido por la bomba endosomal protón-ATPasa. Como consecuencia se solubilizan las proteínas de superficie de la partícula viral y permeabiliza la membrana de la vesícula liberando la partícula de doble capa transcripcionalmente activa en el citosol. La dinamina y la clatrina parecen estar implicadas, no solo en la ruta de endocitosis sino que también en el último paso de entrada durante el movimiento del virus desde la membrana plasmática al citosol (62). Sin embargo, la internalización de todas las cepas rotavirales en vesículas endocíticas no parece depender única y exclusivamente de la presencia de clatrina, ya que cepas de rotavirus como la RRV son capaces de infectar células MA104 incluso haciendo uso de un iRNA contra la cadena pesada de clatrina, sugiriendo así un mecanismo independiente de esta ruta. Experimentos transfectando un mutante de dinamina II indicaron un efecto de la infectividad en todas las cepas de rotavirus empleadas (UK, TRF, Wa, RRV, REO) lo que indicó que existen varias alternativas en el proceso de entrada del virus vía endocitosis dependiendo de la cepa rotaviral (63). El modelo de endocitosis, el cual parece ser dependiente de calcio, se ha planteado con base en la observación de que VP4 y VP7 son capaces de permeabilizar membranas y en el hecho de que la bafilomicina A (agente que bloquea la bomba ATPasa endosomal) bloquea la infección por rotavirus. Es así como en la infección parece estar jugando un rol biológico clave el colesterol y la dinamina (64).
La cápside externa de los virus está conformada por la proteína icosahédrica VP4 y por la glicoproteína VP7, las cuales están implicadas en la interacción con la célula (65) (64). Durante el proceso infeccioso parecen interactuar en la superficie celular con moléculas que contienen ácido siálico, con integrinas (αvβ3, α2β1, α4β7, α4β1 y αXβ2) Hsc70 y PDIs. Estas interacciones inducen un cambio conformacional en la cápside, preparando el virus para desensamblarse y penetrar en la membrana (66-72).
microdominio, en células MA104 y en enterocitos (66, 71-74).
La interacción con ácido siálico parece depender del genotipo de VP4 del virus y no del origen de la especie, por otra parte, hay evidencia de que el gangliósido GM3 sirve como co- receptor del ácido siálico para rotavirus de especie porcina OSU. Se ha propuesto además que rotavirus de diversos orígenes reconocen galactosa, un componente de los receptores tipo glicoproteína de la línea celular MA104. Respecto a las integrinas, la proteína VP4 de rotavirus contiene un motivo secuencia de un tripéptido para las integrinas α2β1 y α4β1 mientras que la proteína VP7 contiene sitios de unión a integrinas para las denominadas
αxβ2 y α4β1 (67, 63).
Otra de las integrinas que parece estar involucrada en la entrada de varias cepas rotavirales post acoplamiento es la αvβ3 y con ella ciertas proteínas de membrana conocidas como proteínas de choque térmico específicamente Hsc70. En resumen, el contacto inicial es a través de un receptor celular que contenga ácido siálico, probablemente un gangliósido que use el dominio viral VP8 de VP4. Esto induce un cambio conformacional en VP4 que permite que el virus interactúe con la integrina α2β1 a través de VP5. Posteriormente estarían involucradas las proteínas virales VP5 y VP7 junto con αvβ3, αxβ2 y la proteína de choque térmico Hsc70. La interacción con Hsc70 está mediada por un dominio viral de VP5, que está localizado entre los aminoácidos 642 y 659, postulando una interacción de tipo ligando-receptor; no es claro si la actividad chaperona de la proteína de choque térmico juega algún rol durante la entrada del virus (63).
4.5. Proteínas de choque térmico
Las proteínas de choque térmico, más conocidas como Hsp (Heat Shock Protein), se expresan de forma constitutiva en las células procariotas y eucariotas. Estas proteínas se expresan cuando se da un cambio de temperatura significativo activando su función citoprotectora como mecanismo de defensa de la célula tras la reacción en situación de estrés celular (agresiones ambientales, exposición a tóxicos, metales pesados, hipoxia, infecciones virales, estados febriles o inflamatorios) disminuyendo los efectos de esta actividad (14).
proteínas (75-76).
4.5.1. Proteínas de choque térmico y virus.
Las infecciones causadas por virus en células de mamífero generan alteraciones en la síntesis de las proteínas de choque térmico (Hsp) que se encuentran en el retículo endoplasmático. Las HSPs chaperonas asisten en la organización de la nucleocapside viral al provocarse una acumulación rápida de las proteínas virales sintetizadas para dar origen a un nuevo virión (77-79).
Por la asociación encontrada entre las proteínas de choque térmico y los virus, se ha propuesto hipotéticamente que estos virus (aquellos con capacidad oncolítica) puedan eventualmente convertirse en herramientas de terapia contra el cáncer. La hipótesis se propone debido a que las proteínas de choque térmico se sobreexpresan en cáncer de seno, melanomas, hepatocarcinoma y tumores cerebrales entre algunos tipos de cáncer (80-83).
La respuesta al choque térmico es un mecanismo de defensa fundamental contra efectos deletéreos a nivel celular tal como el mal plegamiento de proteínas que trae consigo agregación proteica y la alteración del estado redox. Dicha señal de estrés resulta en la activación de factores de transcripción de choque térmico conocidos como HSFs que transcriben las HSPs involucradas en plegamiento y degradación. Ya que las HSPs se involucran directamente en el plegamiento y la apoptosis, se dice que los virus podrían eventualmente usar esas proteínas para el correcto ensamblaje y plegamiento de las proteínas virales manipulando el destino de la célula huésped. En condiciones de no estrés los HSFs están localizados en el citoplasma de células de mamífero y son regulados negativamente, permaneciendo como monómero asociado a chaperonas y co-chaperonas, incluidas la Hsp90 y Hsp70. En condición de estrés las chaperonas liberan el factor de transcripción y este se oligomeriza a un estado trimérico translocándose al núcleo, allí es fosforilado permitiendo la unión al DNA en promotores de secuencias conocidas como elementos de respuesta al choque térmico (84).
factores de choque térmico o inducir la síntesis de HSPs (84). Para evitar la agregación proteica los virus parecen usar las chaperonas celulares de retículo endoplasmático para el plegamiento óptimo de sus dominios, por ejemplo, la Hsp90 (Grp94) y la Hsp70 (Bip) (85).
El tropismo por las HSPs se ha documentado en el virus de la encefalitis, el virus del dengue y el retrovirus humano HTLV-1 (86-88). El primer paso en la infección de la célula blanco consiste en la unión del virión a la superficie de la célula huésped. Se ha asociado la Hsp90 y la Hsp70 con la entrada del virus a la célula independientemente que dichas proteínas no tienen señales de exporte a membrana externa. Es claro que su expresión es común en ciertos tipos de células normales así como en células tumorales facilitando, en teoría, la entrada del virus a la célula.
Luego de una serie de complejos pasos de fusión de membrana y transporte se ha vinculado a la Hsp70 con el desensamblaje de estructuras proteicas oligoméricas del virus, incluyendo las vesículas de clatrina que sirven para la entrada de algunos virus como el AdV y el virus del papiloma (89). Por otra parte, en la transcripción del genoma viral las HSPs parecen tener un rol importante, resaltando el caso de los virus de DNA los cuales transcriben sus genes en el núcleo y hacen uso de la maquinaria transcripcional de huésped y de factores como el caso de la Hsp70. Un ejemplo, es la co-chaperona de la familia Hsp70 Bag-1 la cual interactúa físicamente con factores de iniciación de la transcripción para estimular la actividad transcripcional especulándose que dicha interacción activa o estabiliza factores de transcripción (84). Hsc70 asiste en el ensamblaje del virión de los papilomavirus donde parece interactuar y colocalizar en el núcleo con proteínas como VP1, evitando el ensamblaje de la cápside viral en presencia de Ca2+. Sin embargo, la cápside parece ensamblarse cuando se adiciona un complejo proteico que además de Hsc70 incluye proteínas como Hsp40, con dominio J (90).
Las HSPs parecen asistir en el plegamiento de estructuras virales recién sintetizadas, favoreciendo la organización de la nucleocápside viral, tanto en plantas como en animales, con el propósito de mantener la calidad de las proteínas virales hasta los últimos estados de ciclo viral (84) y posiblemente protegiendo al virus hasta que infecte una nueva célula huésped.
4.5.2. Familias de Hsp
Las familias de las HSPs se clasifican según su peso molecular en varios grupos entre los que se encuentra la familia de pequeñas Hsp (Hsp20, Hsp25, Hsp27), familia de Hsp40, familia Hsp60, familia Hsp70, familia Hsp90 y familia de Hps110 (14):
4.5.2.1. Hsp90
Asiste en la señalización y manteniendo de la integridad funcional de proteínas, ya que ciertas proteínas tienen dificultad en permanecer en sus estados fisiológicamente competentes y requieren la asistencia de proteínas de la familia Hsp90 para su estabilización. Usualmente estas chaperonas trabajan en asociación con la familia de las Hsp70 y algunas otras proteínas auxiliares mediando su función primaria que evita la agregación y favorecer el plegamiento adecuado. Al formar complejos con moléculas de señalización ayudan en la translocación, al correcto compartimento subcelular y al ensamblaje de los complejos de señalización. La Hsp90 parece unirse además a receptores intracelulares y a proteínas transductoras de señales con el propósito de facilitar su plegamiento y estabilidad (94).
plegamiento se exponen dominios que son objeto de modificaciones postraduccionales (85).
La Hsp90 se ha asociado con la infección por rotavirus SA11 en células MA104, ya experimentos utilizan 17-N-alilamino-17-dimetoxigeldanamicina (17-AAG) o bloquear la traducción de esta proteína a través de un RNAi, se disminuyó la infección por rotavirus, además otra evidencia sugiere que Hsp90 trabaja junto con AKT en una vía de señalización clave para la replicación viral (96). De hecho, la Hsp90 parece permitir el correcto plegamiento y estabilización de la proteína NSP3 a través de una interacción directa favoreciendo su unión al RNA y al factor de elongación eIF4G (97).
4.5.2.2. Hsp70
La familia de las Hsp70 asiste en el plegamiento de cadenas polipeptídicas nacientes, los miembros eucariotas de esta familia se unen a zonas hidrofóbicas de las cadenas sintetizadas, ayudando a transportar proteínas a través de membranas de organelos tales como retículo endoplasmático y mitocondria. Estas familias trabajan de manera cooperativa con los miembros de la familia Hsp60 evitando la agregación de proteínas mal plegadas. En citosol de células eucariotas está la forma constitutiva así como la isoforma inducida por estrés. La Hsp70 se une al sustrato involucrando ciclos sucesivos de estados de unión a ATP de baja afinidad y estados de unión a ADP de alta afinidad. Este ciclo es regulado por un dominio denominado J de la Hsp40 y por factores intercambiadores de nucleótidos que controlan la duración de la unión al sustrato. En el proceso de plegamiento los péptidos se unen a DnaK (De la familia Hsp40) a través de interacciones hidrofóbicas y puentes de hidrógeno con el péptido naciente, de esta manera la Hsp70 reconoce características estructurales propias de péptidos nacientes como la exposición de aminoácidos hidrofóbicos. Las Hsp70 poseen dos dominios, un dominio N-terminal ATPasa y un dominio C-N-terminal de unión al sustrato. La estructura tridimensional de este tipo de proteínas muestra que su dominio ATPasa está organizado en dos subdominios globulares que pueden formar cuatro lóbulos distintos que permiten que la molécula de ATP esté rodeada por iones magnesio y potasio. El dominio C- terminal tiene cuatro hojas plegadas β con una serie de “loops” que permiten la unión al sustrato donde la conformación de estas hojas lleva a la exposición de residuos hidrofóbicos que reconocen aminoácidos de las mismas características en proteínas mal plegadas (98).
o denominadas GRP170 que se encuentran en el retículo endoplasmático las cuales se regulan por estrés y presentan un segmento extendido de unión a péptidos (95). Los miembros de las Hsp70 actúan en ambos lados de la membrana celular y bacteriana ensamblando y desensamblando complejos proteicos oligoméricos controlando así la actividad y la estabilidad de quinasas y factores de transcripción. El accionar de este tipo de proteínas está sujeto además a la unión de factores intercambiadores de nucleótidos los cuales se han clasificado en cinco familias: GrpE (procariotas), Bag (citosol de eucariotas), HspBP1 (citosol y retículo endoplasmático), Bap (citosol) y las Hs170 (retículo endoplasmático) (85).
La Hsp70 parece asociarse positivamente con la infectividad de rotavirus RF, ya que se ha visto su sobreexpresión entre la primera y la quinta hora post-infección, sugiriendo un rol en los primeros eventos de infección por rotavirus (99). Además estudios donde se evaluó el perfil de genes expresados en células HT29 tras la infección con las cepas de rotavirus SA11, A5-13 y Wa, identificaron una sobreexpresión de la Hsp70 utilizando los tres tipos de virus, sugiriendo algún rol biológico ya sea negativo o positivo sobre la infección con estas cepas de rotavirus (100).
4.5.2.3. Hsp60
La Hsp60 también parece estar involucrada en procesos de plegamiento proteico en eucariotas. Reside generalmente en mitocondria donde parece establecer interacciones con la Hsp10 en el mencionado proceso de plegamiento. Hay evidencia que indica que también esta proteína de choque térmico interactúa con la Hsp70 (isoforma mitocondrial). También se ha localizado en citosol y en membrana celular tanto de células normales como de células tumorales; asociada en células tumorales a la integrina αVβ3, involucrada en procesos de adhesión y metástasis en algunos tipos de cáncer. Su rol en cáncer está vinculado a mitigación de señales apoptóticas, pérdida de senescencia celular y proliferación descontrolada (101).
A la fecha no se ha documentado interacción o influencia de Hsp60 con rotavirus sin embargo, con el virus de la hepatitis B se ha dado evidencia de una interacción con la polimerasa del virus de la hepatitis B, siendo esta interacción un paso crucial para que la enzima exponga su sitio activo (102). Su influencia negativa sobre la replicación del virus de la hepatitis B se observó in vivo tras utilizar un siRNA contra Hsp60 (103). Existe evidencia de que la Hsp60 podría estar haciendo parte de un complejo proteico que involucra al factor transcripcional HBx del virus de la hepatitis B, donde ensayos de “pull-down” in vitro utilizando proteínas recombinantes y ensayos in vivo empleando la línea celular HepG2, indican que la Hsp60 estaría interactuando con HBx donde se especula que la Hsp60 podría tener función de chaperona molecular permitiendo el correcto plegamiento de la proteína viral (104).
Cuando se analizó su incidencia en la replicación del virus murino de la hepatitis se encontró que Hsp60 junto con otras chaperonas como Hsp70 y Hsp40 están formando un complejo proteico que se une a la región 3’ del RNA viral in vitro. Otros ensayos como inmunoprecipitación junto con ensayos de amplificación en tiempo real por reacción en cadena de la polimerasa confirmaron en células 17C1-1 la existencia de un complejo proteico de Hsp60, Hsp40 y mtHsp70, sugiriendo así algún rol en la replicación viral (105).
4.5.2.4. Hsp40
La proteína Hsp40 se ha encontrado asociada a la Hsp70 cumpliendo funciones individuales muy particulares. Así por ejemplo se ha encontrado una unidad proteica de Hsp70 unida a múltiples unidades de Hsp40 formando complejos de plegamiento de péptidos nacientes. Su principal función parece ser la regulación de la unión de un péptido naciente al bolsillo de ATP de la Hsp70 a través de un dominio conocido como (PPDs). Además de facilitar la unión parece asistir la actividad ATPasa de la Hsp70 a través de un dominio conocido como J, el cual presenta 4 hélices alfa y un dominio HPD (106). Sin embargo, parece tener actividad chaperona asistiendo en el plegamiento de proteínas nacientes de manera independiente al accionar de Hsp70, o también asistiendo como co- chaperona a proteínas como la Hsp90 (107).
presentar únicamente un dominio J y la región C-terminal (106).
Lejos de estas regiones conservadas se encuentran dominios de la Hsp40 que determinan diversa funcionalidad, así por ejemplo cierta región de aminoácidos asiste a la proteína disulfuro isomerasa en la formación de puentes disulfuro en retículo endoplasmático. De modo que su funcionalidad parece depender tanto de la secuencia como de su localización celular; se encuentra ampliamente distribuida en núcleo, endosoma, mitocondria, retículo endoplasmático y ribosoma. Parece presentarse en esta familia “splicing” alternativo cuando el organismo coordina su expresión en un único tejido (107).
La Hsp40 junto con Hsp70 parece tener un rol importante en la patogenicidad del virus del papiloma humano y el virus de la inmunodeficiencia tipo I. Al parecer Hsp40 regula el estrés celular producto de la infección viral además de estar involucrada en el ciclo viral (108). En la infección del virus de la influenza la proteina Hsp40 estaría eliminando la represión celular sobre eIF-2α asegurando la síntesis de las proteínas virales (109). En la infección por el virus aviar CELO, la expresión de la proteína viral Gam/1 se asocia de manera directa con el aumento en la expresión y la colocalización de la Hsp70 y de la Hsp40. Gam/1 parece activar la expresión de la Hsp40 y la respuesta de choque térmico de la célula huésped (110).
Tabla 4. Proteínas de choque térmico localización y función.
FAMILIA NOMBRE LOCALIZACIÓN FUNCIÓN CONFORMACIÓN
Pequeñas Hsp
p20 Citoplasma Vaso
relajación
De 8 a 24 unidades
Hsp22o α B-cristalina
Citoplasma/Núcleo Estabilización del
citoesqueleto Hsp25
(ratón)
Citoplasma/Núcleo Chaperona
Hsp27 (humana)
Citoplasma/Núcleo Dinámica de la actina
Hsp40
Hsp40 Citoplasma Chaperona Monomero
Hsp47 Ret. Endoplásmico Control de síntesis del colágeno
Hsp60 Hsp58 Mitocondrias Chaperona 14 unidades
FAMILIA NOMBRE LOCALIZACIÓN FUNCIÓN CONFORMACIÓN
Hsp70
Hsc70 (73) Citoplasma Chaperona Monómero
Hsc70 (72) Citoplasma/Núcleo Chaperona
Hsp75 Mitocondria Chaperona
Grp78 Ret. Endoplásmico Chaperona
Hsp90
Hsp90α (86) Citoplasma Unión a
receptores de esteroides
Dimero
Hsp90β (84) Citoplasma Unión a
receptores de esteroides Grp94 Ret. Endoplásmico Chaperona
Hsp110
Hsp105 Citoplasma Chaperona
citoprotectora
6 a 7 unidades
Hs0110 Nucléolo/citoplasma Chaperona citoprotectora Tomado de Silva, C., et al (1999) (111)