INMUNOLOGÍA
de Kuby
00 MAQ. PRELI-KINDT.indd i
Célula estromática de la médula ósea
Monocito Macrófago
Célula dendrítica
Eritrocito
Mastocito
Basófilo Eosinófilo
Plaquetas
Célula presentadora de antígeno
Célula B
MHC clase II
Célula TH CD4
Célula B
Neutrófilo
Célula propia alterada MHC clase I
Célula TC CD8 Péptido
antigénico MHC clase I
MHC clase II
Citocina
Receptor de citocina CD4
CD8 Anticuerpo
Receptor de célula T
CD3
Célula TH Célula TC Célula T citotóxica Célula asesina natural
Célula plasmática Timocito
inmaduro
00 MAQ. PRELI-KINDT.indd ii
INMUNOLOGÍA
de Kuby
SEXTA EDICIÓN
MÉXICO • BOGOTÁ • BUENOS AIRES • CARACAS • GUATEMALA • LISBOA MADRID • NUEVA YORK • SAN JUAN • SANTIAGO
SAO PAULO • AUCKLAND • LONDRES • MILÁN • MONTREAL • NUEVA DELHI SAN FRANCISCO • SINGAPUR • ST. LOUIS • SIDNEY • TORONTO
Thomas J. Kindt
National Institutes of Health
Richard A. Goldsby
Amherst College
Barbara A. Osborne
University of Massachusetts at Amherst
Traducción:
Roberto Palacios Martínez
Universidad Autónoma de Baja California
00 MAQ. PRELI-KINDT.indd iii
Supervisor de edición: Camilo Heras Martínez
Supervisora de producción: Ángela Salas Cañada
Composición y formación: Ediciones y Recursos Tecnológicos, S.A. de C.V.
Diseño de portada: Impulso Creativo Publicidad y Diseño, S.C.
INMUNOLOGÍA de Kuby
Prohibida la reproducción total o parcial de esta obra, por cualquier medio, sin autorización escrita del editor.
DERECHOS RESERVADOS © 2007, respecto a la segunda edición en español por McGRAW-HILL INTERAMERICANA EDITORES, S.A. de C.V.
A subsidiary of Th e McGraw-Hill Companies, Inc.
Prolongación Paseo de la Reforma 1015, Torre A, Piso 17, Col. Desarrollo Santa Fe, Delegación Álvaro Obregón
C. P. 01376, México, D. F.
Miembro de la Cámara Nacional de la Industria Editorial Mexicana, Reg. núm. 736
ISBN 13: 978-970-10-6454-2 ISBN 10: 970-10-6454-2
Translated from the sixth english edition of Kuby Immunology
Copyright © 2007, 2003, 2000, 1997, 1994, 1992 by W.H. Freeman and Company. All Rights Reserved
ISBN 13: 978-1-4292-0211-4 ISBN 10: 1-4292-0211-4
1234567890 09865432107 Impreso en México Printed in Mexico
NOTA
La medicina es una ciencia en constante desarrollo. Conforme surjan nuevos conocimientos, se requeri-rán cambios de la terapéutica. El (los) autor(es) y los editores se han esforzado para que los cuadros de dosifi cación medicamentosa sean precisos y acordes con lo establecido en la fecha de publicación. Sin embargo, ante los posibles errores humanos y cambios en la medicina, ni los editores ni cualquier otra per-sona que haya participado en la preparación de la obra garantizan que la información contenida en ella sea precisa o completa, tampoco son responsables de errores u omisiones, ni de los resultados que con dicha información se obtengan. Convendría recurrir a otras fuentes de datos, por ejemplo, y de manera particular, habrá que consultar la hoja informativa que se adjunta con cada medicamento, para tener certeza de que la información de esta obra es precisa y no se han introducido cambios en la dosis recomendada o en las contraindicaciones para su administración. Esto es de particular importancia con respecto a fármacos nue-vos o de uso no frecuente. También deberá consultarse a los laboratorios para recabar información sobre los valores normales.
00 MAQ. PRELI-KINDT.indd iv
Dra. Alicia del Toro Arreola
Doctora en Ciencias Biomédicas, Orientación en Inmunología, Centro Universitario de Ciencias de la Salud de la
Universidad de Guadalajara.
Dra. Susana del Toro Arreola
Doctora en Ciencias de la Salud, Orientación Biomédica, Centro Universitario de Ciencias de la Salud de la Universidad de Guadalajara.
Profesor Investigador Titular C
Centro Universitario de Ciencias de la Salud Universidad de Guadalajara.
Dra. Trinidad García Iglesias
Doctora en Inmunología
Centro Universitario en Ciencias de la Salud de la Universidad de Guadalajara.
Profesora de Bioquímica en el Centro Universitario de Ciencias de la Salud de la Universidad de Guadalajara.
Miembro de Biólogos Colegiados de Jalisco A.C.
Dra. Cecilia Magdalena Guillén Vargas
Doctora en Inmunología y Profesora Investigadora Titular A de la Universidad de Guadalajara, Centro Universitario de Ciencias de la Salud, Departamento de Fisiología.
Dra. Ana Molina Ocaña
Doctora en Ciencias Biológicas
Investigadora y Docente en la Sección de Inmunopatología Experimental e Inmunoquímica de Investigación, Hospital Ramón y Cajal (Madrid).
Investigadora y Docente en el Department of Nutritional Chemistry, School of Medicine, University of Tokushima (Japón).
Investigadora y Docente en el Área de Inmunología, Facultad de Biología, Universidad de Vigo (Pontevedra). Miembro de la Sociedad Española de Inmunología (SEI).
Dr. Saturnino Muñoz Martínez
Doctor en Ciencias Biológicas
Investigador y Docente en la Sección de Inmunopatología Experimental e Inmunoquímica de Investigación, Hospital Ramón y Cajal (Madrid).
Investigador y Docente en el Department of Nutritional Chemistry, School of Medicine, University of Tokushima (Japón).
Investigador y Docente en el Área de Inmunología, Facultad de Biología, Universidad de Vigo (Pontevedra). Miembro de la Sociedad Española de Inmunología (SEI).
Dr. Pedro Ernesto Sánchez Hernández
Doctor en Ciencias Biomédicas con orientación a inmunología Centro Universitario de Ciencias de la Salud – Centro Universitario de Ciencias Biológicas y Agropecuarias de la Universidad de Guadalajara.
Profesor de Inmunología del Centro Universitario de Ciencias de la Salud de la Universidad de Guadalajara.
00 MAQ. PRELI-KINDT.indd v
Thomas J. Kindt
es requerido con regularidad como consultor en temas de inmu-nología y enfermedades infecciosas por organizaciones gubernamentales y priva-das, y ha fungido por muchos años como director de investigación intramuros en el National Institute of Allergy and Infectious Diseases de los National Institutes of Health, un cargo que lo mantiene en contacto diario con la vanguardia de la inmu-nología clínica y experimental. Es profesor adjunto en el Departamento de Biología de la University of New Mexico y pertenece a la Regional Association of Medical and Biological Organizations con sede en New Mexico.Richard A. Goldsby
enseña inmunología a estudiantes de licenciatura y posgrado en el Amherst College. Sus intereses en la investigación incluyen tecnologías para generar anticuerpos humanos y sometidos a ingeniería genética en biorreactores animales. En muchas ocasiones ha sido director de curso en el Chautauqua Short Course Program de la National Science Foundation, donde presenta los avances vi-gentes en la inmunología a profesores universitarios.Barbara A. Osborne,
de la University of Massachusetts at Amherst, es una con-tribuyente reconocida en las áreas rápidamente cambiantes de muerte celular pro-gramada y desarrollo de reacciones de células T. Investigadora muy activa, Barbara también imparte cursos de inmunología a estudiantes de licenciatura y posgrado.Janis Kuby,
quien murió en 1997, enseñó en la San Francisco State University y en la University of California at Berkeley. La profesora Kuby fue quien inició este libro y es autora de las primeras tres ediciones. Su enseñanza experta y sus habilidades para la escritura hicieron de Inmunología el texto más vendido para el curso; su visión de la obra como una forma de combinar el contenido actualizado con un formato accesible y pedagógicamente rico persiste en la nueva edición.De izquierda a derecha:Richard A. Goldsby, Barbara A. Osborne y Th omas J. Kindt
00 MAQ. PRELI-KINDT.indd vi
emoción y alegría.
Esperamos que las generaciones futuras de inmunólogos
encuentren el tema tan satisfactorio como nosotros.
00 MAQ. PRELI-KINDT.indd vii
Prefacio
E
n la segunda edición de Inmunología, Janis Kuby escri-bió: “...el crecimiento continuo de la inmunología es in-evitable y desafía tanto a la comunidad médica como a la• Células, moléculas antimicrobianas solubles y receptores unidos a membrana colaboran para montar un ataque instantáneo contra los agentes infecciosos.
• El sistema inmunitario actúa no sólo como primera línea de defensa, sino también como un activador esencial para el sistema inmunitario adaptativo.
• Los defectos en los componentes del sistema inmunitario innato a menudo dan por resultado reacciones débiles o inadecuadas del sistema inmunitario adaptativo.
Hincapié en la pertinencia clínica
Una inmunorreacción defi ciente o excesiva puede tener conse-cuencias nefastas. Es fundamental que quienes estén interesa-dos en seguir carreras médicas comprendan el funcionamiento de este sistema. En todo el libro se cubre una amplia gama de enfermedades e infecciones nuevas, y se actualizan los ensayos Enfoque clínico y sus correspondientes Preguntas de estudio al fi nal del capítulo. En esta edición se incorporan:
• Explicación de la presentación cruzada en lo que se refi ere a inmunidad a virus y otros agentes infecciosos (cap. 8). académica para mantenerse actualizada.” Nuestro objetivo con
cada nueva edición de Inmunología es presentar el conocimiento a una nueva generación de científi cos y profesionales médicos. Debemos dar a quienes se aproximan por primera vez al tema un panorama amplio del campo de la inmunología. Tenemos que mantenernos actualizados. Y debemos además introducir los experimentos y modelar los sistemas sobre los cuales se ha construido nuestro conocimiento del sistema inmunitario.
Nuevo capítulo 3: Inmunidad innata
Los medios por los cuales se dedujeron los mecanismos del sis-tema inmunitario innato y el rápido avance en el conocimiento sobre esta rama de la inmunidad se encuentran entre los de-sarrollos más impactantes en inmunología desde la edición anterior de este libro. El nuevo capítulo 3, Inmunidad innata, explora el modo en que
• Las actividades de efectores inmunitarios como los receptores de reconocimiento de patrón se integran en la inmunorreacción innata.
Célula bacteriana (E. coli) Organización de la pared celular
Membrana externa
Membrana interna Peptidoglucano Lipopolisacárido (endotoxina)
FIGURA 3-9 Lipopolisacárido (LPS) en la pared celular de E. coli. El LPS es un potente estímulo de la inmunidad innata.[Micrografía de Gary Gaugler/Visuals Unlimited.]
00 MAQ. PRELI-KINDT.indd viii
• Mayor cobertura del receptor de célula T γδ, incluida una imagen tridimensional reciente que revela diferencias en el modo en que los receptores de célula T γδ y αβ se unen a antígeno (caps. 9 y 10).
Nuevas técnicas
Las siguientes exposiciones de técnicas modernas importantes han producido fascinantes conocimientos nuevos en el campo de la inmunología:
• Descripción de la técnica de resonancia de plasmones
superfi ciales y su aplicación a interrogantes básicas en
inmunología (cap. 6).
• Uso de tetrámeros antígeno-MHC marcados para etiquetar
receptores de célula T unidos a membrana (cap. 14). • Ilustración del poder de la microscopia bifotónica para
seguir el recorrido de células en un ganglio linfático (caps. 11 y 22).
Organización actualizada
Tras consultar con numerosos profesores de inmunología, en la sexta edición se realizaron los siguientes cambios de organiza-ción para mejorar la secuencia de exposiorganiza-ción y evitar redundan-cias:
• Se combinaron los capítulos sobre anticuerpo y antígeno (cap. 4).
• El capítulo acerca del complemento se adelantó, para situarlo inmediatamente después de los capítulos sobre anticuerpo (cap. 7).
• Se combinaron los capítulos de MHC y presentación de antígeno (cap. 8).
Herramientas pedagógicas
Figuras para visualización de conceptos
Varios conceptos son especialmente cruciales para que los es-tudiantes establezcan conocimientos fi rmes de inmunología. En este libro se emplean imágenes diseñadas de manera espe-cífi ca con este fi n, para ayudar a los estudiantes a dominar el material.
• Los conceptos principales se ilustran en fi guras para
visualización de conceptos. Estas fi guras resumen ideas
y procesos importantes de tal manera que un texto escrito no puede lograr por sí solo; a menudo se disponen como diagramas “de recorrido”, que incluyen leyendas más extensas y sistemáticas que ayudan a visualizar procesos clave.
• Se hace uso consistente de iconos, los cuales representan diversas células del sistema inmunitario y moléculas de membrana importantes, para ayudar a los estudiantes a visualizar relaciones complejas. Estos iconos aparecen al principio del libro en un cuadro, que constituye una guía accesible.
• Una exposición más amplia de las citocinas y su cometido en infl amación y enfermedad (cap. 12 en adelante). • Últimos descubrimientos sobre la diversidad de receptores
de célula NK y el modo en que su variabilidad genética infl uye en la susceptibilidad a enfermedades (cap. 14). • Nuevo ensayo Enfoque clínico sobre la infl uencia de
KIR/MHC en la enfermedad (cap. 14).
• Nueva cobertura de la tolerancia central y periférica y el modo en que se relacionan con enfermedad
autoinmunitaria y rechazo de aloinjertos (cap. 16). • Nuevas exposiciones sobre métodos para aliviar el
sufrimiento causado por diversos trastornos de
autoinmunidad como esclerosis múltiple, lupus eritematoso y enfermedad de Crohn (cap. 16).
• Cobertura del uso clínico creciente de los anticuerpos monoclonales como agentes terapéuticos (caps. 4, 5, 6, 16, 17 y otros).
• Mayor cobertura de enfermedades infecciosas, incluyendo descripciones de grupos de patógenos importantes y las inmunorreacciones características que provocan; material actualizado sobre gripe, incluidas las cepas aviares y las amenazas que representan para las poblaciones humanas; y por qué las enfermedades micóticas han aumentado en grado signifi cativo debido a la propagación del SIDA y al aumento de la cantidad de personas que toman
medicamentos contra enfermedades autoinmunitarias (cap. 18).
• Cobertura del SARS, incluidos el descubrimiento y la determinación del modo en que saltó de los animales al ser humano (caps. 18 y 19).
• Nuevos datos sobre consecuencias del SIDA (cap. 20). • Informe sobre la relación entre el virus del papiloma humano
(HPV) y el cáncer cervicouterino, los ensayos de vacunas para su prevención, y un nuevo Enfoque clínico donde se examinan más a fondo estos descubrimientos (cap. 21).
Mayor cobertura sobre señalización
En los últimos pocos años ha habido un gran avance en el co-nocimiento sobre los procesos que ocurren después de que los receptores se unen a sus ligandos. Ahora se dedica una sección a presentar el tema general de la transducción de señales, donde se resume el patrón general de señalización y se nombran algu-nos de los componentes clave más universales. Por ejemplo, se incluyen
• Una nueva sección donde se describe la transducción de señales que sigue a la unión de los receptores tipo Toll con sus ligandos (cap. 3).
• Mayor cobertura de las interacciones moleculares implicadas en la migración y la extravasación celulares (caps. 3 y 13).
• Nuevos detalles sobre las vías de señalización que llevan a maduración, diferenciación y activación de diversos tipos celulares (caps. 10 y 11).
00 MAQ. PRELI-KINDT.indd ix
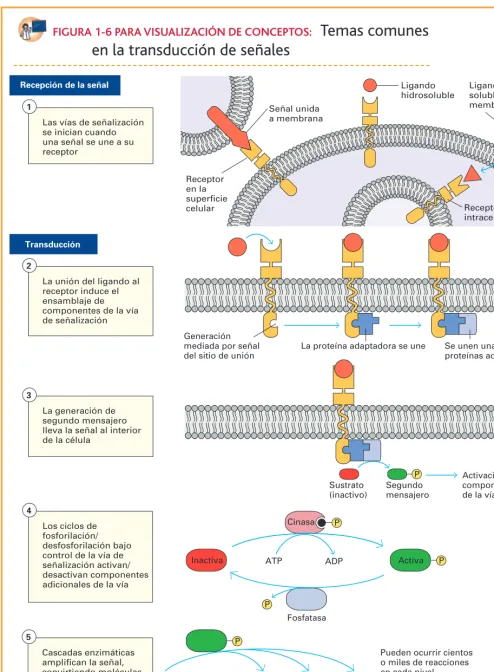
FIGURA 1-6 PARA VISUALIZACIÓN DE CONCEPTOS: Temas comunes
en la transducción de señales
P
P P
P
P Las vías de señalización
se inician cuando una señal se une a su receptor
La unión del ligando al receptor induce el ensamblaje de componentes de la vía de señalización
La generación de segundo mensajero lleva la señal al interior de la célula
Los ciclos de fosforilación/ desfosforilación bajo control de la vía de señalización activan/ desactivan componentes adicionales de la vía
Cascadas enzimáticas amplifican la señal, convirtiendo moléculas a sus formas activas
Recepción de la señal
Transducción
Señal unida a membrana
Ligando hidrosoluble
Ligando soluble en membrana
1
2
3
4
5
Inactiva ATP ADP Activa
Sustrato (inactivo)
Segundo mensajero
Activación de componentes de la vía Receptor
en la superficie celular
Generación mediada por señal del sitio de unión
La proteína adaptadora se une Se unen una o más proteínas adicionales
Cinasa
Fosfatasa
Pueden ocurrir cientos o miles de reacciones en cada nivel
Efectores metabólicos
Receptor intracelular
00 MAQ. PRELI-KINDT.indd x
Preguntas de estudio
Ideadas por Janis Kuby, las Preguntas de estudio de Inmunolo-gía han demostrado ser un valioso recurso para instructores y estudiantes por igual. En todos los capítulos de la sexta edición se presentan preguntas nuevas y revisadas, incluida una serie totalmente nueva titulada Analice los datos, donde se utilizan bibliografía moderna y datos cuantitativos, y se desafía a los es-tudiantes a extrapolar información con las herramientas adqui-ridas en el estudio del texto. Las preguntas se complementan con respuestas ampliadas y actualizadas como material de res-paldo al fi nal del libro.
Reconocimientos
Debemos un agradecimiento especial a varios colegas que ayu-daron a realizar complejas revisiones y que realizaron lecturas detalladas, todo lo cual desembocó en grandes mejoras al tex-to. Entre estos notables contribuyentes se incluyen los doctores J. Donald Capra y Kendra White de la Oklahoma Medical Re-search Foundation, la doctora JoAnn Meerschaert de la Saint Cloud State University, el doctor Jiri Mestecky de la Universi-ty of Alabama at Birmingham, el doctor Jonathan Yewdell de NIAID, NIH, el doctor James Kindt de la Emory University, la doctora Johnna Wesley de la Brown University, y el doctor Eric Long de NIAID, NIH. Esperamos que el producto fi nal refl eje la alta calidad de lo aportado por estos expertos y por todos aque-llos que se enumeran más adelante, que proporcionaron ideas críticas y orientación.
También deseamos expresar nuestra gratitud y aprecio a la doc-tora JoAnn Meerschaert de la Saint Cloud State University por haber escrito excelentes problemas nuevos, y al doctor Stephen K. Chapes, de la Kansas State University, por crear las preguntas de la sección Analice los datos.
Agradecemos a los siguientes revisores sus comentarios y su-gerencias acerca del manuscrito durante la elaboración de esta sexta edición. Su maestría y su agudeza contribuyeron enorme-mente a este libro.
Ruth D. Allen, Indiana University-Purdue University
Indianapolis
Avery August, Th e Pennsylvaia State University
Pamela J. Baker, Bates College
Kenneth J. Balazovich, University of Michigan
Cynthia L. Baldwin, University of Massachusetts Amherst Scott R. Barnum, University of Alabama, Birmingham Stephen H. Benedict, University of Kansas
Earl F. Bloch, College of Medicine Howard University Lisa Borghesi, University of Pittsburgh
Lauren Brossay, Brown University
Jane Bruner, California State University, Stanislaus James W. Campbell, Rice University
Stephen Keith Chapes, Kansas State University Koteswara R. Chintalacharuvu, UCLA
Jefrey R. Dawson, Duke University, School of Medicine Janet M. Decker, University of Arizona
Michael Edidin, Th e Johns Hopkins University
Sherry D. Fleming, Kansas State University
Scott C. Garman, University of Massachusetts, Amherst Elizabeth Godrick, Boston University
Sandra O. Gollnick, Roswell Park Cancer Institute Hans W. Heidner, Th e University of Texas at San Antonio
Vincent W. Hollis, Jr., Howard University W. Martin Kast, University of Southern California Dennis J. Kitz, Southern Illinois University, Edwardsville Katherine L. Knight, Loyola University
Paul M. Knopf, Brown University
Kay K. Lee-Fruman, California State University, Long Beach Alan D. Levine, Case Western Reserve University
Judith Manning, University of Wisconsin School of Medicine James A. Marsh, Cornell University College of Veterinary
Medicine
John Martinko, Southern Illinois University Carbondale Andrea M. Mastro, Th e Pennsylvania State University
Jennifer M. Mataraza, Boston College Dennis W. McGee, Binghamton University JoAnn Meerschaert, Saint Cloud State University Jiri Mestecky, University of Alabama, Birmingham Michael F. Minnick, University of Montana
Th omas W. Molitor, University of Minnesota, College of
Veterinary Medicine
David M. Mosser, University of Maryland Rita B. Moyes, Texas A&M University Philip C. Nelson, University of Pennsylvania Alma Moon Novotny, Rice University Kim O’Neill, Brigham Young University Luke O’Neill, Trinity College, Dublin, Ireland Leonard D. Pearson, Colorado State University Christopher A. Pennel, University of Minnesota Wendy R. Raymond, Williams College
Robert C. Rickert, University of California, San Diego Kenneth H. Roux, Florida State University
Abhineet Sheoran, Tuft s Cummings School of Veterinary
Medicine
Michail Sitkovsky, Northeastern University Robert C. Sizemore, Alcorn State University Gary Splitter, University of Wisconsin, Madison Douglas A. Steeber, University of Wisconsin, Milwaukee Lisa Steiner, Massachusetts Institute of Technology Jeff rey L. Stott, University of California, Davis School of
Veterinary Medicine
Denise G. Wingett, Boise State University Jon Yewdell, NIH-NIAID
Kirk Ziegler, Emory University School of Medicine
Asimismo, deseamos dar las gracias a nuestros experimenta-dos y talentosos colegas de W. H. Freeman and Company. Nues-tro agradecimiento en especial a Kate Ahr, Georgia Lee Hadler, Karen Taschek, Vicki Tomaselli, Paul Rohloff , Susan Timmins, Ted Szczepanski, Hannah Th onet y Nick Tymoczko. La ejecu-ción de este trabajo no habría sido posible sin la férrea determi-nación de nuestro editor de desarrollo, Morgan Ryan, quien nos ayudó a ilustrar la historia de la inmunología.
00 MAQ. PRELI-KINDT.indd xi
Prefacio
Contenido resumido
PARTE I
Introducción
1
Panorama general del sistema inmunitario
1
2
Células y órganos del sistema inmunitario
23
3
Inmunidad
innata
52
PARTE II
Respuestas de las células B y T
4
Antígenos y anticuerpos
76
5
Organización y expresión de los genes de inmunoglobulina
111
6
Interacciones antígeno-anticuerpo: principios y aplicaciones
145
7
Sistema
del
complemento
168
8
Complejo mayor de histocompatibilidad y presentación de antígeno
189
9
Receptor de célula T
223
10
Maduración, activación y diferenciación de la célula T
245
11
Generación, activación y diferenciación de la célula B
271
PARTE III
Mecanismos inmunoefectores
12
Citocinas
302
13
Activación y migración de leucocitos
327
14
Reacciones citotóxicas mediadas por células
351
15
Reacciones de hipersensibilidad
371
16
Tolerancia y autoinmunidad
401
PARTE IV
El sistema inmunológico en la salud y la enfermedad
17
Inmunología de los trasplantes
425
18
Inmunorreacción a las enfermedades infecciosas
447
19
Vacunas
475
20
SIDA y otras inmunodefi ciencias
493
21
Cáncer y sistema inmunitario
525
22
Sistemas
experimentales
546
Apéndice I: Antígenos CD
A-1
Apéndice II: Citocinas
A-27
Glosario
G-1
Respuestas a las preguntas de estudio
R-1
Índice
alfabético
I-1
00 MAQ. PRELI-KINDT.indd xii
Prefacio viii
PARTE I Introducción
1
Panorama general del sistema
inmunitario
1
Perspectiva histórica 2
Los estudios pioneros sobre la vacunación abrieron
el campo para la inmunología 2
La vacunación es una tarea continua a nivel mundial 3
Primeros estudios sobre inmunidad humoral y celular 4
Desafíos teóricos 5
Infección e inmunidad 7
Inmunidad innata y adaptativa 8
Las células fagocíticas constituyen una barrera contra
las infecciones 9
Algunas moléculas solubles contribuyen a la inmunidad
innata 9 La colaboración entre la inmunidad innata y la adaptativa
incrementa la inmunorreactividad 9
La inmunidad adaptativa es altamente específi ca 10
Linfocitos y células presentadoras de antígeno cooperan
en la inmunidad adaptativa 12
Las células presentadoras de antígeno interactúan con
células T 14
Las inmunorreacciones humoral y celular tienen distintas
funciones efectoras 14
Los receptores de antígeno de los linfocitos B y T son diversos 14 Las moléculas del complejo mayor de histocompatibilidad
unen péptidos antigénicos 16
La selección de antígeno por los linfocitos causa
expansión clonal 16
Disfunción inmunitaria y sus consecuencias 18
ENFOQUE CLÍNICO ALERGIAYASMACOMOPROBLEMASGRAVES
DESALUDPÚBLICA 20
2
Células y órganos
del sistema inmunitario
23
Hematopoyesis 23
La hematopoyesis se regula a nivel genético 24
En la hemostasia hematopoyética intervienen muchos
factores 26 La muerte celular programada es un mecanismo
homeostático esencial 26
Las células madre hematopoyéticas pueden enriquecerse 28
Células del sistema inmunitario 30
Células linfoides 30
ENFOQUE CLÍNICO CÉLULASMADRE: USOSCLÍNICOSYPOTENCIAL 32
Linfocitos B (células B) 34
Linfocitos T (células T) 34
Las poblaciones de células B y T comprenden subpoblaciones
de clonas 35
Células asesinas naturales 35
Fagocitos mononucleares 36
La fagocitosis es seguida de la digestión y presentación
de antígeno 36
Células granulocíticas 37
Células cebadas 38
Células dendríticas 38
Células dendríticas foliculares 40
Órganos del sistema inmunitario 40
Órganos linfoides primarios 40
Órganos linfoides secundarios 43
Células y órganos linfoides: comparaciones
evolutivas 49
3
Inmunidad innata
52
Barreras anatómicas 53
Conexiones entre la inmunidad innata
y la adaptativa 55
Infl amación 57
La extravasación leucocitaria es un proceso altamente
regulado de múltiples pasos 59
Moléculas solubles y receptores relacionados
con membrana 59
Los péptidos antimicrobianos contribuyen a la defensa innata
contra bacterias y hongos 59
Las proteínas de la reacción de fase aguda contribuyen a la
inmunidad innata 61
La inmunidad innata utiliza diversos receptores para detectar infección 61
xiii
00 MAQ. PRELI-KINDT.indd xiii
Receptores tipo Toll 62
Tipos celulares de inmunidad innata 65
Los neutrófi los se especializan en fagocitosis y matanza 65 ENFOQUE CLÍNICO LAPROTEÍNA C REACTIVAESUNMARCADOR
CLAVEDERIESGOCARDIOVASCULAR 66
Los macrófagos despliegan varios recursos contra los
patógenos 66 Las células NK son una importante primera línea de
defensa contra los virus y constituyen una señal
de activación clave para otras células 68
Las células dendríticas atacan patógenos e invocan
inmunorreacciones adaptativas al activar células T 68
Vías de transducción de señales 69
La señalización por TLR es típica de las vías de transducción
de señales 69
Ubicuidad de la inmunidad innata 71
PARTE II Respuestas
de las células B y T
4
Antígenos y anticuerpos
76
Inmunogenicidad y antigenicidad 77
Los haptenos son valiosos instrumentos de investigación
y diagnóstico 77
Las propiedades del inmunógeno contribuyen a la
inmunogenicidad 78 El sistema biológico contribuye a la inmunogenicidad 80
Epítopos 81
Los epítopos de células B tienen propiedades
características 81
Estructura básica y función de los anticuerpos 84
Los anticuerpos son heterodímeros 85
Métodos químicos y enzimáticos revelaron la estructura
básica del anticuerpo 85
La determinación de las secuencias de la cadena ligera
reveló regiones constantes y variables 87
Existen cinco clases principales de cadenas pesadas 87 Las inmunoglobulinas poseen múltiples dominios con base
en el plegamiento de la inmunoglobulina 87
Sitio de unión de anticuerpos 89
Las CDR unen antígeno 90
La unión de antígeno puede inducir cambios
conformacionales 92
Dominios de región constante 93
Funciones efectoras mediadas por anticuerpo 94
El anticuerpo promueve la opsonización 94
Los anticuerpos activan el complemento 95
La citotoxicidad mediada por células dependiente
de anticuerpo (ADCC) destruye otras células 95
Algunos anticuerpos pueden cruzar capas epiteliales por
transcitosis 95
Clases de anticuerpos y actividades biológicas 95
Inmunoglobulina G (IgG) 95
Inmunoglobulina M (IgM) 96
ENFOQUE CLÍNICO TERAPÉUTICAPASIVACONANTICUERPO 98
Inmunoglobulina A (IgA) 99
Inmunoglobulina E (IgE) 100
Inmunoglobulina D (IgD) 100
Determinantes antigénicos en inmunoglobulinas 100
Isotipo 101 Alotipo 101 Idiotipo 101
Receptor de célula B 102
Los receptores Fc se enlazan con regiones Fc de anticuerpos 102
Superfamilia de las inmunoglobulinas 103
Anticuerpos monoclonales 105
Los anticuerpos monoclonales tienen usos clínicos
importantes 106 Las abzimas son anticuerpos monoclonales que catalizan
reacciones 106
5
Organización y expresión
de los genes de inmunoglobulina
111
Diseño de un modelo genético compatible
con la estructura de la inmunoglobulina 112
Modelos de línea germinal y de variación somática
propuestos para explicar la diversidad de anticuerpos 113 Dreyer y Bennett propusieron un revolucionario modelo
de dos genes-un polipéptido 113
La bomba de Tonegawa: los genes de la inmunoglobulina
se reordenan 114
Organización multigénica de genes
de inmunoglobulina 115
Cada familia multigénica tiene características distintas 115
Familia multigénica de la cadena pesada 115
Reordenamientos génicos de región variable 116
El DNA de la cadena ligera experimenta
reordenamientos VJ 117
El DNA de cadena pesada experimenta
reordenamientos VDJ 118
Mecanismo de los reordenamientos de DNA
de región variable 119
Secuencias señal dirigen la recombinación 119
Los segmentos génicos se unen mediante recombinasas 119 Los reordenamientos del gen de inmunoglobulina (Ig)
pueden ser productivos o improductivos 121
00 MAQ. PRELI-KINDT.indd xiv
La exclusión alélica asegura la especifi cidad
antigénica única 121
Generación de diversidad de anticuerpos 123
Existen numerosos segmentos génicos V, D y J de la línea
germinal 123
La unión combinatoria VJ y VDJ genera diversidad 123 La fl exibilidad de unión contribuye a la diversidad 123 La adición P añade diversidad a secuencias palindrómicas 125 La adición N promueve una considerable diversidad por la
agregación de nucleótidos 125
La hipermutación somática agrega diversidad en segmentos
génicos ya reordenados 125
Un origen último de la diversidad es la asociación
combinatoria de cadenas pesada y ligera 127
La diversifi cación de los genes de inmunoglobulina difi ere
entre las especies 127
Cambio de clase entre genes de la región constante 128
La desaminasa de citidina inducida por activación (AID) media tanto la hipermutación somática como el
cambio de clase 128
Expresión de genes de inmunoglobulina 130
Los transcritos primarios de cadena pesada experimentan
procesamiento diferencial del RNA 130
Síntesis, ensamblaje y secreción
de inmunoglobulinas 133
Regulación de la transcripción de genes
de inmunoglobulina 133
El reordenamiento del DNA acelera en gran medida
la transcripción 135
En las células T está inhibida la expresión de los genes
de inmunoglobulina 135
Genes de anticuerpos e ingeniería de anticuerpos 136
Los anticuerpos monoclonales quiméricos y humanizados
poseen un gran potencial clínico 136
Se han creado ratones con loci de inmunoglobulina
humana 137 Las bibliotecas de exhibición en fago permiten
producir anticuerpos monoclonales sin necesidad
de inmunización 137
ENFOQUE CLÍNICO TERAPÉUTICADELLINFOMANO HODGKIN YOTRASENFERMEDADESMEDIANTEANTICUERPOSELABORADOSPOR
INGENIERÍAGENÉTICA 140
6
Interacciones
antígeno-anticuerpo: principios
y aplicaciones
145
Potencia de las interacciones antígeno-anticuerpo 145
La afi nidad de anticuerpo es una medida cuantitativa
de la fuerza de la unión 145
La avidez del anticuerpo incorpora la afi nidad de múltiples
sitios de unión 148
Reactividad cruzada 149
Resonancia de plasmones superfi ciales 149
La SPR puede usarse para caracterizar las especifi cidades
de epítopo de grupos de anticuerpos 150
Reacciones de precipitación 151
Las reacciones de precipitación en gel producen líneas de
precipitina visibles 151
La inmunoelectroforesis combina electroforesis e
inmunodifusión doble 152
Reacciones de aglutinación 153
La hemaglutinación se utiliza en la tipifi cación sanguínea 153 La aglutinación bacteriana se emplea para diagnosticar
infecciones 154 La aglutinación pasiva es útil con antígenos solubles 154 En la inhibición de la aglutinación, la ausencia de
aglutinación es diagnóstica de antígeno 154
Radioinmunoensayo 154
Ensayo de inmunosorbente ligado a enzima 155
Existen múltiples variantes de ELISA 155
Western blotting 158
Inmunoprecipitación 158
Inmunofl uorescencia 160
Citometría de fl ujo y fl uorescencia 161
Alternativas a las reacciones antígeno-anticuerpo 162
ENFOQUE CLÍNICO CITOMETRÍADEFLUJOYTIPIFICACIÓNDE
LEUCEMIAS 163
Microscopia inmunoelectrónica 164
7
Sistema del complemento
168
Funciones del complemento 168
Componentes del complemento 169
Activación del complemento 169
La vía clásica se inicia con la unión de antígeno y anticuerpo 170
La vía alterna es independiente de anticuerpo 173
La vía de lectina se inicia con la unión de proteínas del
hospedador a superfi cies microbianas 175
Las tres vías del complemento convergen en el complejo
de ataque a membrana 175
Regulación del sistema del complemento 177
Consecuencias biológicas de la activación
del complemento 180
El complejo de ataque a membrana puede lisar una amplia
gama de células 180
Los productos de escisión de componentes del complemento
median la infl amación 182
00 MAQ. PRELI-KINDT.indd xv
ENFOQUE CLÍNICO HEMOGLOBINURIAPAROXÍSTICANOCTURNA:
UNDEFECTODELAREGULACIÓNDELALISISPORCOMPLEMENTO 183
La unión de C3b y C4b facilita la opsonización 184
El sistema del complemento también neutraliza la
infectividad vírica 184
El sistema del complemento depura inmunocomplejos
de la circulación 185
Defi ciencias de complemento 185
8
Complejo mayor
de histocompatibilidad
y presentación de antígeno
189
Organización general y herencia del MHC 190
El MHC codifi ca tres clases de moléculas principales 190 Las formas alélicas de los genes MHC se heredan en
grupos unidos llamados haplotipos 191
Las cepas endogámicas de ratón han sido de utilidad
en el estudio del MHC 193
Moléculas y genes MHC 193
Las moléculas clase I tienen una cadena pesada de
glucoproteína y una cadena ligera proteínica pequeña 193 Las moléculas clase II tienen dos cadenas glucoproteínicas
distintas 195 La disposición de exones e intrones en los genes clase I
y clase II refl eja su estructura de dominio 196
Las moléculas clase I y clase II muestran polimorfi smo
en la región que se une a péptidos 197
Las moléculas clase I y clase II muestran diversidad dentro de una especie, y se presentan múltiples
formas de ellas en un individuo 199
Mapa genómico detallado de los genes MHC 201
La región de la clase I humana abarca alrededor de 2 000 kb en el extremo telomérico del complejo de antígenos de
histocompatibilidad leucocíticos (HLA) 202
Los genes del MHC clase II se localizan en el extremo
centromérico del HLA 203
Los genes del MHC clase III del ser humano están entre
las clases I y II 203
Expresión celular de moléculas MHC 203
Regulación de la expresión del MHC 204
MHC y susceptibilidad a enfermedades 205
MHC e inmunorreactividad 206
Restricción de células T a MHC propio 207
Función de las células presentadoras
de antígeno 207
Es necesario que el antígeno sea procesado para que
las células T lo reconozcan 208
La mayoría de las células puede presentar antígeno con MHC clase I; la presentación con MHC clase II se restringe a células presentadoras de
antígeno (APC) 209
Pruebas de la existencia de diferentes vías
de procesamiento y presentación de antígeno 209
Antígenos endógenos: vía citosólica 210
Complejos de proteasa llamados proteasomas generan
los péptidos para presentación 211
Los péptidos se transportan del citosol al retículo
endoplásmico rugoso 211
Los péptidos se ensamblan con MHC clase I auxiliados
por carabinas moleculares 212
ENFOQUE CLÍNICO LADEFICIENCIADETRANSPORTADORES RELACIONADOSCONLAPRESENTACIÓNDEANTÍGENO (TAP) CAUSA
UNADIVERSAGAMADEENFERMEDADES 213
Antígenos exógenos: vía endocítica 214
Los péptidos se generan a partir de moléculas
internalizadas en vesículas endocíticas 214
La cadena invariante guía el transporte de moléculas
MHC clase II a las vesículas endocíticas 214
Los péptidos se ensamblan con moléculas MHC clase II
por desplazamiento de CLIP 215
Presentación cruzada de antígenos exógenos 217
Presentación de antígenos no peptídicos 217
9
Receptor de célula T
223
Primeros estudios sobre el receptor de célula T 223
Experimentos clásicos demostraron la restricción
del receptor de célula T al MHC propio 224
El uso de anticuerpos clonotípicos permitió aislar
receptores de célula T 224
El gen de la cadena del TCR se clonó mediante
hibridación sustractiva 224
Receptores de célula T ␣ y ␥␦: estructuras
y funciones 226
Organización y reordenamiento de los genes
del TCR 228
Los genes de la región variable del TCR se reordenan
de manera similar a los genes de anticuerpo 229
Mecanismo de los reordenamientos del DNA del TCR 230
Exclusión alélica de los genes de TCR 231
Los genes del TCR reordenados se ensamblan a partir
de segmentos génicos V, D y J 232
ENFOQUE CLÍNICO REORDENAMIENTOSDECÉLULAS T COMO
MARCADORESDECÉLULASCANCEROSAS 231
La diversidad de TCR se genera en forma similar a la
diversidad de anticuerpos pero sin mutación somática 232
Complejo receptor de célula T: TCR-CD3 235
Moléculas de membrana accesorias
de la célula T 236
Correceptores CD4 y CD8 se unen a regiones conservadas
de las moléculas MHC clase II o I 236
00 MAQ. PRELI-KINDT.indd xvi
La afi nidad del TCR por complejos péptido-MHC es
intensifi cada por correceptores 238
Estructuras tridimensionales de complejos
TCR-péptido-MHC 240
Los TCR interactúan de manera diferente con moléculas
clase I y clase II 241
Alorreactividad de las células T 241
10
Maduración, activación
y diferenciación de la célula T
245
Timo y maduración de la célula T 245
Selección tímica del repertorio de células T 248
La selección positiva asegura la restricción en MHC 249 La selección negativa asegura la autotolerancia 250 Algunos experimentos revelaron los elementos esenciales
de las selecciones positiva y negativa 250
Algunos temas centrales de la selección tímica aún no
se resuelven 251
Activación de la célula T 254
La unión del TCR inicia múltiples vías de señalización 254 ¿Cuántos complejos de TCR deben ensamblarse para
inducir la activación de la célula T? 258
Para la activación completa de las células T se requieren
señales coestimuladoras 259
Cuando no existe una señal coestimuladora se presenta
anergia clonal 259
Los superantígenos inducen la activación de células T
al unir el TCR y el MHC II de modo simultáneo 260
Diferenciación de la célula T 261
Las células T activadas generan células T efectoras y
de memoria 262
Una subpoblación CD4CD25 de células T regula
de modo negativo las inmunorreacciones 263
Las células presentadoras de antígeno tienen propiedades
coestimuladoras características 263
Muerte celular y poblaciones de células T 264
ENFOQUE CLÍNICO LAFALTADEAPOPTOSISCAUSAHOMEOSTASIS
DEFECTUOSADELINFOCITOS 266
11
Generación, activación
y diferenciación de la célula B
271
Maduración de la célula B 271
Las células B progenitoras proliferan en la médula ósea 272 El reordenamiento del gen de inmunoglobulina (Ig)
produce células B inmaduras 273
Para el desarrollo de la célula B es esencial el receptor
de célula pre-B 274
En experimentos de desactivación génica se identifi caron
factores de transcripción esenciales 275
Los marcadores de superfi cie celular identifi can las etapas
del desarrollo 275
Las células B-1 son un subconjunto de células B que se
renuevan por sí mismas 275
En la médula ósea se seleccionan negativamente células B autorreactivas 276 Es posible rescatar células B autorreactivas al editar genes
de cadena ligera 278
Activación y proliferación de la célula B 278
Los antígenos dependientes e independientes del timo
tienen diferentes requerimientos para reaccionar 278 Dos tipos de señales impulsan a las células B hacia el ciclo
celular y a través de él 279
La transducción de señales activadoras incluye
heterodímeros Ig-/Ig- 279
La señalización de células B es iniciada por la unión de antígeno e induce muchas vías de transducción de
señales 281 El complejo correceptor de célula B puede intensifi car las
reacciones de la célula B, y el CD22 es capaz de inhibirlas 281 ENFOQUE CLÍNICO AGAMMAGLOBULINEMIALIGADAALSEXO:
UNAFALLAENLATRANSDUCCIÓNDESEÑALESYELDESARROLLO
DELACÉLULA B 284
Las células TH tienen acciones esenciales en la mayor parte
de las reacciones de la célula B 285
Es posible la selección negativa de células B autorreactivas
maduras en la periferia 287
Reacción humoral 289
Las respuestas primaria y secundaria difi eren en grado
signifi cativo 289
Las células T colaboradoras tienen un papel crítico en
la reacción humoral a conjugados de hapteno y portador 290
Sitios in vivo para la inducción de reacciones
humorales 292
Centros germinales y diferenciación de la célula B
inducida por antígeno 292
La maduración de la afi nidad es el resultado de mutaciones
y selecciones repetidas 293
Las células B de memoria y las células plasmáticas se
generan en centros germinales 296
Regulación de la inmunorreacción efectora 297
Diferentes antígenos pueden competir entre sí 297
La presencia de anticuerpo puede suprimir la reacción
al antígeno 297
PARTE III Mecanismos
inmunoefectores
12
Citocinas
302
Propiedades de las citocinas 302
Las citocinas pertenecen a cuatro familias estructurales 305
00 MAQ. PRELI-KINDT.indd xvii
Las citocinas tienen múltiples funciones biológicas 306
Receptores de citocinas 307
Los receptores de citocina pertenecen a cinco familias 308 Las subfamilias de los receptores de citocina clase I tienen
en común subunidades de señalización 309
IL-2R es uno de los receptores de citocinas estudiado de
modo más extenso 311
Los receptores de citocina inician la señalización 312
Antagonistas de citocinas 314
Secreción de citocinas por los subconjuntos
TH1 y TH2 314
El desarrollo de los subconjuntos TH1 y TH2 depende
del ambiente de las citocinas 316
Los perfi les de citocinas son regulados de manera
cruzada 317 El balance TH1/TH2 determina los resultados fi nales
de una enfermedad 318
Enfermedades relacionadas con citocinas 318
El choque séptico es común y potencialmente letal 318 El choque tóxico bacteriano se debe a superantígenos 319 La actividad de las citocinas se relaciona con los cánceres
linfoide y mieloide 319
La enfermedad de Chagas es causada por un parásito 320
Tratamientos basados en citocinas 320
Citocinas en la hematopoyesis 321
ENFOQUE CLÍNICO TERAPÉUTICACONINTERFERONES 322
13
Activación y migración
de leucocitos
327
Moléculas de adhesión celular 327
Quimiocinas 329
Los perfi les de receptores de quimiocina median
la actividad de los leucocitos 331
Extravasación de leucocitos: el paradigma
de los pasos múltiples 332
Recirculación de linfocitos 334
Extravasación de linfocitos 334
Las vénulas con endotelios altos (endotelios venulares
altos, HEV) son sitios de extravasación de linfocitos 334 El direccionamiento (tráfi co) de los linfocitos es guiado
por perfi les y señales de los receptores 336
Los linfocitos vírgenes recirculan hacia tejido linfoide
secundario 336 Los linfocitos efectores y de memoria adoptan patrones
de tráfi co distintos 337
Otros mediadores de la infl amación 338
La lesión tisular activa el sistema de las cininas 338
El sistema de la coagulación proporciona mediadores de la
infl amación generados por la fi brina 338
El sistema fi brinolítico proporciona mediadores de la
infl amación generados por la plasmina 338
El sistema del complemento produce anafi latoxinas 338 Algunos lípidos actúan como mediadores de la infl amación 339 Algunas citocinas son mediadores importantes de la
infl amación 339
Proceso infl amatorio 340
Los neutrófi los tienen un papel temprano e importante
en la infl amación 340
Las reacciones infl amatorias pueden ser localizadas
o generalizadas 340
ENFOQUE CLÍNICO DEFICIENCIADEADHESIÓNLEUCOCÍTICA (LAD)
ENSERESHUMANOSYBOVINOS 343
Se desarrolla infl amación crónica cuando el antígeno
persiste 344 Funciones de IFN- y TNF- en la infl amación crónica 344 En enfermedades infl amatorias crónicas se producen
estructuras tipo HEV 346
Agentes antiinfl amatorios 346
Los tratamientos con anticuerpos reducen la extravasación
de leucocitos 346
Los corticosteroides son fármacos antiinfl amatorios
potentes 347
Los NSAID combaten el dolor y la infl amación 347
14
Reacciones citotóxicas
mediadas por células
351
Reacciones efectoras 351
Propiedades generales de las células T efectoras 352
Las necesidades de activación de las células T son diferentes 352 Las moléculas de adhesión celular facilitan las interacciones
mediadas por el receptor de célula T (TCR) 352
Las células T efectoras expresan varias moléculas efectoras 353
Células T citotóxicas 353
Los linfocitos T citotóxicos efectores se generan a partir de
precursores propios 353
Los linfocitos T citotóxicos CD8pueden rastrearse con
tecnología de tetrámeros MHC 355
Los linfocitos T citotóxicos (CTL) destruyen células de
dos maneras 355
Células asesinas naturales 360
Las células asesinas naturales (NK) y las células T
comparten algunas características 361
La destrucción (muerte) por células asesinas naturales
es similar a la mediada por linfocitos T citotóxicos 361 Las células asesinas naturales tienen receptores de
activación e inhibición 362
ENFOQUE CLÍNICO COMBINACIONESDEGENES MHC-KIR
INFLUYENENLASALUD 364
00 MAQ. PRELI-KINDT.indd xviii
Células NKT 364
Citotoxicidad mediada por células dependiente
de anticuerpo 366
Valoración experimental de la citotoxicidad
mediada por células 366
El cultivo concurrente de células T con células extrañas
estimula la reacción de linfocitos mixtos 366
Es posible demostrar la actividad de linfocitos T citotóxicos
mediante linfólisis mediada por células 367
La reacción de injerto contra hospedador indica
citotoxicidad mediada por células 367
15
Reacciones de hipersensibilidad
371
Clasifi cación de Gell y Coombs 371
Hipersensibilidad mediada por IgE (tipo I) 372
Existen varios componentes comunes de las reacciones
tipo I 373
El enlace cruzado de IgE inicia la desgranulación 377 Sucesos intracelulares inducen la desgranulación de
los leucocitos 377
Diversos agentes farmacológicos median las reacciones
tipo I 379
Las reacciones tipo I pueden ser generales o localizadas 381 Las reacciones de fase tardía inducen infl amación
localizada 383 Las reacciones tipo I son reguladas por muchos factores 383
ENFOQUE CLÍNICO GENÉTICADELASMA 384
Se emplean diversos métodos para identifi car las reacciones
de hipersensibilidad tipo I 386
Las hipersensibilidades tipo I pueden controlarse por medios médicos 386
Hipersensibilidad citotóxica mediada por
anticuerpo (tipo II) 388
Las reacciones transfusionales son tipo II 388
La enfermedad hemolítica del neonato se debe a
reacciones tipo II 389
La anemia hemolítica inducida por fármacos es una
reacción tipo II 391
Hipersensibilidad mediada por inmunocomplejos
(tipo III) 391
Las reacciones tipo III pueden ser localizadas 392
Las reacciones tipo III también pueden ser generalizadas 392
Hipersensibilidad tipo IV o tardía (DTH) 393
Existen diversas fases de la reacción de hipersensibilidad
tardía 394 Numerosas citocinas participan en la reacción de
hipersensibilidad tardía 395
La reacción de hipersensibilidad tardía se identifi ca con
una prueba cutánea 396
La dermatitis por contacto es un tipo de reacción de
hipersensibilidad tardía 396
16
Tolerancia y autoinmunidad
401
Establecimiento y mantenimiento de la tolerancia 402
La tolerancia central limita el desarrollo de células T y B
autorreactivas 403 La tolerancia periférica regula las células autorreactivas en
circulación 404 Las células T reguladoras son un componente de la
tolerancia periférica 406
El secuestro de antígeno es un modo de proteger
antígenos propios 407
El fallo de la tolerancia causa autoinmunidad 407
Enfermedades autoinmunitarias específi cas
de órganos 407
Algunas enfermedades autoinmunitarias son mediadas
por lesión celular directa 407
Algunas enfermedades autoinmunitarias son mediadas por
autoanticuerpos estimuladores o bloqueadores 409
Enfermedades autoinmunitarias sistémicas 410
El lupus eritematoso sistémico ataca muchos tejidos 410 La esclerosis múltiple ataca el sistema nervioso central 411 La artritis reumatoide ataca las articulaciones 411
Modelos animales de enfermedades
autoinmunitarias 411
Los animales pueden desarrollar autoinmunidad de manera
espontánea 412
Puede inducirse autoinmunidad de manera experimental en animales 413
Pruebas de la participación de células T CD4ⴙ,
MHC y TCR en la autoinmunidad 413
Las células T CD4 y el equilibrio entre las células TH1 y TH2 desempeñan una función importante en la
autoinmunidad de algunos modelos animales 414
La autoinmunidad puede relacionarse con MHC o
con receptores de células T particulares 414
Mecanismos propuestos para la inducción
de autoinmunidad 414
La liberación de antígenos secuestrados puede inducir
enfermedad autoinmunitaria 415
ENFOQUE CLÍNICO ¿PORQUÉLASMUJERESSONMÁSSUSCEPTIBLES QUELOSVARONESALAAUTOINMUNIDAD? DIFERENCIASDEGÉNERO
ENLASENFERMEDADESAUTOINMUNITARIAS 416
El mimetismo molecular puede contribuir a la enfermedad autoinmunitaria 416 Se cuenta con pruebas de imitación entre MBP
y péptidos víricos 418
La expresión inapropiada de moléculas MHC clase II
puede sensibilizar las células T autorreactivas 418 La activación de las células B policlonales puede ocasionar
enfermedad autoinmunitaria 419
Tratamiento de las enfermedades autoinmunitarias 419
El tratamiento de las enfermedades autoinmunitarias del ser
humano plantean desafíos especiales 420
00 MAQ. PRELI-KINDT.indd xix
La infl amación es un blanco del tratamiento de la
autoinmunidad 420 Las células T activadas son un posible blanco terapéutico 421
Los antígenos orales pueden inducir tolerancia 421
PARTE IV El sistema inmunológico
en la salud y la enfermedad
17
Inmunología de los trasplantes
425
Bases inmunitarias del rechazo de injertos 426
El rechazo de aloinjertos manifi esta especifi cidad y memoria 426 Las células T desempeñan una función clave en el rechazo
de los aloinjertos 426
Los perfi les antigénicos similares propician la aceptación
de los aloinjertos 428
Se determinan los antígenos eritrocíticos y los MHC de los
donantes y los receptores de injertos 428
El rechazo de injerto mediado por células se produce en
dos etapas 431
Manifestaciones clínicas del rechazo de injertos 433
Los anticuerpos preexistentes en el receptor median el
rechazo hiperagudo 433
Reacciones de células T median el rechazo agudo 434 El rechazo crónico ocurre meses o años después
del trasplante 434
Tratamiento inmunosupresor general 434
Los inhibidores de la mitosis impiden la proliferación
de las células T 434
ENFOQUE CLÍNICO ¿TIENENFUTUROCLÍNICOLOS
XENOTRASPLANTES? 435
Los corticosteroides suprimen la infl amación 436
Algunos metabolitos micóticos son inmunosupresores 436 La radiación linfoide total elimina los linfocitos 436
Tratamiento inmunosupresor específi co 436
Los anticuerpos pueden suprimir las reacciones de
rechazo de injerto 437
Bloquear las señales coestimuladoras puede inducir anergia 438
Inmunotolerancia a los aloinjertos 439
Los sitios privilegiados aceptan desigualdades antigénicas 439 La exposición temprana a los aloantígenos puede inducir
tolerancia específi ca 439
Trasplante clínico 440
El órgano trasplantado con más frecuencia es el riñón 441 Se practican trasplantes de médula ósea para tratar
leucemias, anemias e inmunodefi ciencias 441
El trasplante cardíaco es una operación desafi ante 442 Los trasplantes de pulmón son cada vez más comunes 442 Los trasplantes de hígado se practican para tratar
defectos congénitos y lesiones por agentes víricos
o químicos 442
El trasplante de páncreas ofrece una curación de la diabetes mellitus 443 Se emplean injertos cutáneos para tratar a las víctimas
de quemaduras 443
El xenotrasplante puede ser la solución ante la escasez
de órganos de donante humano 444
18
Inmunorreacción a las
enfermedades infecciosas
447
Infecciones víricas 448
Muchos virus son neutralizados por anticuerpos 449
La inmunidad mediada por células es importante para
el control y la depuración víricos 450
Los virus pueden evadir los mecanismos de defensa
del hospedador 450
La gripe es la causa de algunas de las peores pandemias
de la historia 451
La reacción humoral a la gripe es específi ca de cepa 454 La cepa aviar H5N1 representa una amenaza de pandemia 454
Infecciones bacterianas 455
Las reacciones inmunitarias a las bacterias extracelulares e
intracelulares pueden diferir 455
Las bacterias pueden evadir con efi cacia los mecanismos
de defensa del hospedador 455
Las reacciones inmunitarias pueden contribuir a la
patogénesis bacteriana 457
La difteria (Corynebacterium diphteriae) puede controlarse mediante inmunización con toxoide desactivado 458 La tuberculosis (Mycobacterium tuberculosis) es controlada
primordialmente por células T CD4 458
Enfermedades parasitarias 460
Las enfermedades por protozoarios afectan a millones
de personas en todo el mundo 460
El paludismo (especies de Plasmodium) infecta a
600 millones de personas en todo el mundo 460
Dos especies de Trypanosoma causan la enfermedad
del sueño 462
La leishmaniosis es un modelo útil para demostrar
las diferencias en las reacciones del hospedador 462 Diversas enfermedades son causadas por gusanos
parásitos (helmintos) 462
Enfermedades micóticas 465
La inmunidad innata controla la mayoría de las
infecciones micóticas 466
La inmunidad contra patógenos micóticos puede ser
adquirida 466
Enfermedades infecciosas emergentes 467
Las enfermedades pueden resurgir por diversas razones 467 ENFOQUE CLÍNICO AMENAZADEINFECCIÓNPORAGENTES
POTENCIALESDEBIOTERRORISMO 468
Otras enfermedades letales han aparecido recientemente 470 El brote de SARS desencadenó una rápida respuesta
internacional 470
00 MAQ. PRELI-KINDT.indd xx
19
Vacunas
475
ENFOQUE CLÍNICO VACUNACIÓN: DESAFÍOSEN ESTADOS UNIDOS
YENPAÍSESENDESARROLLO 476
Inmunizaciones activa y pasiva 477
La inmunización pasiva consiste en la transferencia
de anticuerpos preformados 477
La inmunización activa confi ere protección prolongada 478
Diseño de vacunas para inmunización activa 481
Vacunas con microorganismos vivos atenuados 481
Vacunas de microorganismos desactivados o
muertos 484
Vacunas subunitarias 484
Algunos toxoides se emplean como vacunas 485
Se usan cápsulas bacterianas de polisacárido como vacunas 485 Podrían elaborarse vacunas con glucoproteínas víricas 485 Por medio de técnicas recombinantes se obtienen proteínas
de agentes patógenos 485
El empleo de péptidos sintéticos como vacunas ha
progresado con lentitud 485
Vacunas conjugadas 486
Un polisacárido confi ere protección contra varios hongos 486 Las vacunas subunitarias multivalentes confi eren
inmunidad celular y humoral 486
Vacunas de DNA 488
Vacunas con vectores recombinantes 488
20
SIDA y otras inmunodefi ciencias
493
Inmunodefi ciencias primarias 493
Las inmunodefi ciencias linfoides pueden incluir células T,
células B o ambas 495
Las inmunodefi ciencias del linaje mieloide afectan
la inmunidad innata 500
Los defectos del complemento causan inmunodefi ciencia o
enfermedad por inmunocomplejos 502
Los trastornos de inmunodefi ciencia se tratan mediante
restitución del elemento defectuoso 502
Los modelos experimentales de inmunodefi ciencia incluyen
animales alterados por medios genéticos 503
SIDA y otras inmunodefi ciencias adquiridas
o secundarias 504
La epidemia de VIH/SIDA ha cobrado millones de vidas
a nivel mundial 505
El VIH-1 se propaga por contacto sexual, sangre infectada
y de madre a hijo 505
ENFOQUE CLÍNICO PREVENCIÓNDELAINFECCIÓNINFANTIL
POR VIH MEDIANTETRATAMIENTOANTIRRETROVÍRICO 507
El retrovirus VIH-1 es el causante del síndrome
de inmunodefi ciencia adquirida 508
Los estudios in vitro revelaron el ciclo de multiplicación
del VIH-1 509
La infección por el VIH-1 propicia infecciones oportunistas 512 Los agentes terapéuticos inhiben la multiplicación
de los retrovirus 515
Es posible que una vacuna sea el único medio para
detener la epidemia de VIH/SIDA 518
21
Cáncer y sistema inmunitario
525
Cáncer: origen y terminología 525
Transformación maligna de células 526
Oncogenes e inducción de cáncer 527
Los genes relacionados con el cáncer tienen muchas
funciones 527 Los protooncogenes pueden convertirse en oncogenes 529 La inducción del cáncer es un proceso de múltiples etapas 530
Tumores del sistema inmunitario 530
Antígenos tumorales 531
Algunos antígenos son específi cos de tumor 532
ENFOQUE CLÍNICO UNAVACUNAQUEPREVIENEELCÁNCER 534
Los virus pueden inducir antígenos tumorales 535
Pocos antígenos tumorales son exclusivos de células
tumorales 536 Los tumores pueden inducir reacciones inmunitarias
potentes 537 Células asesinas naturales y macrófagos son
importantes en el reconocimiento de tumores 537
Evasión del sistema inmunitario por los tumores 538
Los anticuerpos antitumorales pueden intensifi car
el crecimiento de los tumores 538
Los anticuerpos pueden modular los antígenos tumorales 538 Las células tumorales expresan con frecuencia
concentraciones bajas de moléculas MHC clase I 538 Las células tumorales pueden emitir señales
coestimuladoras defi cientes 538
Inmunoterapia del cáncer 539
La manipulación de las señales coestimuladoras puede
incrementar la inmunidad 539
El incremento de la actividad de células presentadoras
de antígeno puede modular la inmunidad tumoral 540 El tratamiento con citocinas puede acentuar
las inmunorreacciones a los tumores 540
Los anticuerpos monoclonales son efi caces para
tratar ciertos tumores 542
22
Sistemas experimentales
546
Modelos animales experimentales 546
Las cepas endogámicas pueden reducir la variación
experimental 547
00 MAQ. PRELI-KINDT.indd xxi
Los sistemas de transferencia adoptivos permiten
el examen in vivo de poblaciones de células aisladas 547
Sistemas de cultivo celular 547
Los cultivos de células linfoides primarias provienen
de sangre u órganos linfoides 547
Las líneas celulares linfoides clonadas son herramientas
importantes en inmunología 549
Creación de líneas de células linfoides híbridas 550
Bioquímica de proteínas 551
Las técnicas de radiomarcado permiten la identifi cación
sensible de antígenos o anticuerpos 551
Las marcas de biotina facilitan la detección de pequeñas
cantidades de proteínas 551
La electroforesis en gel separa las proteínas por tamaño
y carga 551
La cristalografía de rayos X ofrece información estructural 553
Tecnología de DNA recombinante 555
Las enzimas de restricción escinden el DNA en secuencias precisas 555
Las secuencias de DNA se clonan en vectores 556
Los vectores de clonación son de utilidad para duplicar
secuencias de DNA defi nidas 556
La clonación del cDNA y el DNA genómico permite
el aislamiento de secuencias defi nidas 556
Las clonas de DNA se seleccionan por hibridación 558 La prueba de Southern blotting identifi ca el DNA
de una secuencia determinada 559
La prueba de Northern blotting identifi ca mRNA 559
La reacción en cadena de la polimerasa amplifi ca
cantidades pequeñas de DNA 559
Análisis de secuencias reguladoras del DNA 560
El análisis de huellas de DNA identifi ca los sitios en que
se fi jan proteínas a éste 560
El análisis de retardo en gel identifi ca complejos de DNA
y proteína 561
Los ensayos de luciferasa miden la actividad
transcripcional 562
Transferencia de genes a células de mamífero 562
Los genes clonados transferidos a células cultivadas
permiten el análisis in vitro de la función génica 562 Los genes clonados transferidos a embriones de ratón
permiten el análisis in vivo de la función génica 563 En los ratones con desactivación génica, el gen
seleccionado se daña 564
La tecnología “knock-in” permite reemplazar un gen
endógeno 565 La selección de genes inducibles por el sistema Cre/lox
tiene como fi nalidad la supresión génica 565
Microarreglos: método para analizar patrones
de expresión génica 567
ENFOQUE CLÍNICO ANÁLISISDEMICROARREGLOSCOMO
INSTRUMENTODIAGNÓSTICOPARALASENFERMEDADESHUMANAs 568 Microscopia bifotónica para visualización in vivo
del sistema inmunitario 570
Avances en la tecnología de fl uorescencia 571
Apéndice I: Antígenos CD
A-1
Apéndice II: Citocinas
A-27
Glosario G-1
Respuestas a las preguntas de estudio
R-1
Índice alfabético
I-1
00 MAQ. PRELI-KINDT.indd xxii