II Programa de Especialización y Diplomado
en Agrobiotecnología
Quito - Ecuador
2008
Laboratorio de Biotecnología Agrícola
[
FISIOLOGIA VEGETAL
]
Respiración Celular
La respiración consiste en una serie ordenada de reacciones que permiten la degradación escalonada de la glucosa (o de otros compuestos orgánicos), hasta CO2 y H2O. La energía liberada en muchos de estos procesos se utiliza para
sintetizar ATP a partir de ADP y Pi. La molécula de ATP representa un medio transitorio de almacenaje de energía; energía que será empleada posteriormente en diversos procesos de síntesis de la célula viva.
La glucólisis y la fermentación son relativamente poco eficaces en la producción de ATP, mientras que el ciclo de Krebs asociado al sistema transportador de electrones representa la mayor aportación de ATP a la célula viva. Aunque la degradación de la molécula de glucosa por la vía de la glucólisis y del ciclo de Krebs representa la cadena metabólica más empleada por la respiración, el ciclo de la hexosa-monofosfato debe ser también considerado como un camino posible, en muchos organismos.
La intensidad de la respiración está influenciada, también por varios factores del medio, entre los cuales se cuenta la temperatura, la concentración de oxígeno, la concentración de anhídrido carbónico, la concentración de sales inorgánicas en el medio de cultivo, entre otros.
Durante los procesos de fotosíntesis la mayor parte de la energía almacenada por las plantas se encuentra en forma de glúcidos, como almidón y glucosa. La debilitación o ruptura de los enlaces carbono-carbono de compuestos de este tipo libera una enorme cantidad de energía que puede ser utilizada por la planta. Sin embargo la cantidad de energía contenida en un compuesto, como la glucosa, debe ser liberada lentamente a lo largo de una serie de reacciones escalonadas, reguladas por enzimas. En general una serie de reacciones celulares que conducen a la síntesis o a la lisis de un compuesto orgánico de denomina vía metabólica.
ATP: Reserva de energía
El ATP es un compuesto intermedio, que sirve de reserva temporal de energía. La energía liberada en la oxidación de compuestos como glúcidos, lípidos entre otros, es inmediatamente utilizada en la síntesis de ATP, dependiente del Adenosin-difosfato (ADP) y el fósforo inorgánico (Pi). El enlace que une el último fosfato al ADP recibe el nombre de enlace de alta energía, por la facilidad que
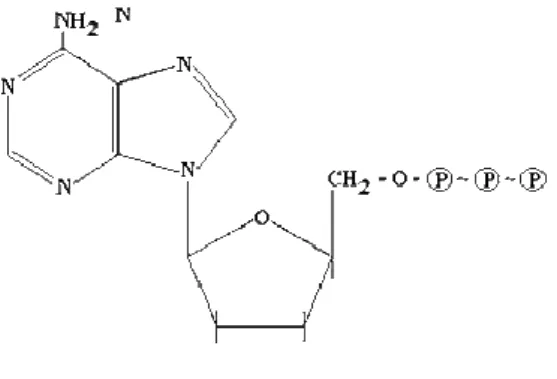
posee de ceder el último grupo fosfato. En este proceso la energía se transfiere a otro compuesto. Así tenemos un compuesto intermedio que es capaz de recibir la energía liberada de una reacción y transferirla a otra reacción que la necesite (Fig. 1).
Fig 1. Estructura molecular del ATP.
Respiración en las plantas
Las plantas se han desarrollado sin formar órganos especializados para el intercambio gaseoso y sin pigmentos transportadores de O2 y CO2. Las plantas
terrestres que viven en una atmósfera gaseosa han desarrollado espacios aéreos intercelulares que son continuos desde los estomas y lenticelas hasta cada célula viva que forma la planta. El intercambio de gases tiene lugar por difusión a través de estos espacios aéreos en respuesta a gradientes de concentración.
Glucólisis y Fermentación
La glucólisis es la secuencia de reacciones que convierten a la glucosa o glucosa-1-fosfato, procedente de la degradación fosforolítica del almidón, en piruvato con la producción de ATP. La glucólisis, que tiene lugar en el citoplasma celular es la primera de las tres fases de la respiración; siguen a continuación el ciclo de Krebs y el proceso de transporte de electrones, ambos mecanismos localizados en la mitocondria. Bajo condiciones aeróbicas, el piruvato entra en la mitocondria y es oxidado completamente hasta CO2 y H2O. En condiciones
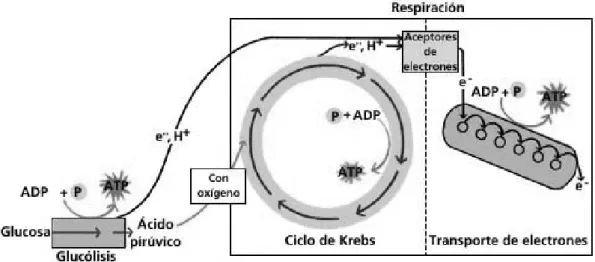
anaeróbicas, el piruvato puede convertirse en lactato o etanol mediante procesos en los que no se produce la degradación completa como en la respiración. La respiración y la fermentación están relacionadas, porque comparten los mismos procesos (fig 3).
Fig. 3. Esquema de producción de ATP derivado de la glucólisis.
Podemos considerar que la glucólisis desempeña tres funciones importantes para el metabolismo vegetal:
1. A partir del ácido fosfoenolpirúvico puede sintetizar aminoácidos aromáticos como fenilalanina y tirosina, que sirven como precursores de la lignina; como también precursores de componentes de la pared celular, etc.
2. Almacenamiento de energía en forma de ATP. Durante la glucólisis se consumen 2 ATP y se forman 4 ATP por la que el saldo final es la formación de 2 ATP.
3. Formación de 1 NADH durante la oxidación del gliceraldehido 3-fosfato en 1-3 difosfoglicerato.
Por tanto la ecuación de la transformación de glucosa en piruvato es:
Dado que el citoplasma celular tiene una cantidad limitada de NAD existen mecanismos para reoxidar el NADH formado. En condiciones aerobias el NADH en la mitocondria será reoxidado por el sistema transportador de electrones, produciendo ATP; bajo condiciones anaerobias, la reoxidación se realiza por las reacciones catalizadas por la alcohol y láctico deshidrogenadas.
En la fermentación alcohólica no se produce oxido-reducción neta en la conversión de la glucosa en alcohol. La reacción neta de fermentación es la siguiente:
Etanol: Glucosa + 2Pi + 2ADP → 2etanol + 2CO2 + 2ATP
Al igual que en la fermentación alcohólica, en la fermentación láctica no existe óxido-reducción neta. La reacción neta de la fermentación láctica es:
Lactato: Glucosa + 2Pi + 2ADP → 2lactato + 2ATP
La producción de ATP durante la glucólisis es un acontecimiento importante, ya que va a permitir que los organismos anaerobios completen sus ciclos biológicos a expensas exclusivamente de esta energía.
Mitocondrias vegetales
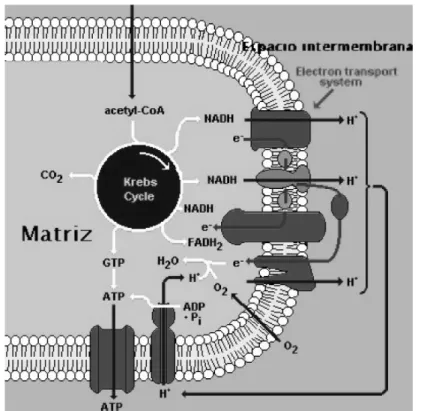
Las mitocondrias son el centro de respiración celular, y donde tienen lugar la mayor producción de energía en forma de ATP, para todas las reacciones celulares. Las células vegetales suelen poseer unas 200 mitocondrias aunque se han llegado a detectar hasta 2000 en los elementos inmaduros del metaxilema en brotes jóvenes de maíz.
La mitocondria tiene un sistema de membranas especializado. La membrana externa es menos permeable, permite el paso de sustancias cargadas como no cargadas con peso molecular menor de 10000, de esta forma, permite el paso de moléculas de bajo peso molecular, siendo igual la concentración en el citoplasma que en el espacio intermembranal. La membrana interna presenta una permeabilidad muy limitada para la mayoría de las sustancias, con excepción de moléculas no cargadas o con peso molecular menor a 150.
Fig 4. Esquema del proceso metabólico en la célula vegetal.
Fosforilación Oxidativa
Es el proceso por el cual se forma ATP, cuando se transfieren los electrones el NADH o el FADH2 al O2 mediante una serie de transportadores de electrones. La
fuerza que dirige la fosforilación oxidativa es el potencial de transferencia de electrones del NADH. El cambio en el potencial redox entre el NAD+ y el oxígeno molecular es de 1,14 voltios ya que el potencial redox del NAD+ es de -0.32 voltios y el del oxígeno de +0.82 voltios.
Formación de Acetil-coenzima A
Si bien, la degradación de la glucosa en condiciones anaeóbicas (citoplasma) produce ácido pirúvico siendo este la terminación del proceso glucolítico. Si existe suficiente oxígeno, se produce una decarboxilación oxidativa del ácido pirúvico que desemboca en la formación de acetil-CoA. Esta reacción es muy compleja y requiere, al menos, de cinco cofactores esenciales, así como de complejos de enzimas. La ecuación que resume este proceso es:
Piruvato + CoA + NAD+ → aceti-CoA + CO2 + NADH + H+
La reacción está catalizada por el complejo de la piruvato deshidrogenasa. A demás los dos electrones transportados al NAD+, para formar NADH, acaban
pasando por el sistema transportador de electrones, como resultado del cual se formarán tres moléculas de ATP.
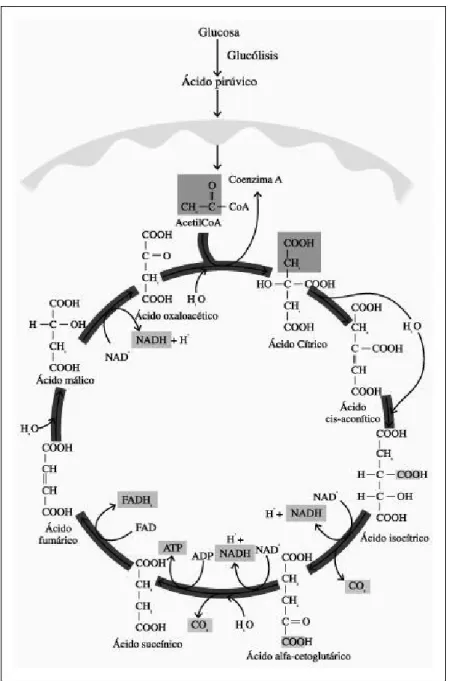
Ciclo de Krebs
También se lo conoce como el ciclo del ácido cítrico o ciclo de los ácidos tri-carboxílicos y se encuentra localizado en el interior de la mitocondria. La descarboxilación oxidativa del piruvato, producido en la glucólisis para formar Acetil CoA y que tiene lugar en la matriz mitocondrial, es el eslabón entre la glucólisis y el ciclo de Krebs.
En el ciclo de Krebs (Fig. 5) un compuesto de cuatro carbonos, el oxalacetato se condensa con el acetilo para dar lugar a un ácido tricarboxílico de seis carbonos, el citrato. Mediante la descarboxilación oxidativa de un isómero del citrato se forma un compuesto de cinco átomos de carbono, el a-cetoglutarato que vuelve a descarboxilarse oxidativamente para dar un ácido de cuatro carbonos, el succionato que regenerará de nuevo al oxalacetato. De esta forma dos átomos de carbono entran en el ciclo como una unidad acetilo y dos átomos dejan el ciclo como dos moléculas de CO2. Dado que el CO2 está más oxidado que el
grupo acetilo, deben existir algunas reacciones de óxido-reducción en el ciclo, de hecho existen cuatro de estas reacciones. Tres iones hidruro, por tanto seis electrones, son transferidos al NAD+ y un par de átomos de hidrógeno (dos electrones) al FAD que producirán un total de once moléculas de ATP en la cadena de transporte de electrones. Además de formar un enlace fosfato de alta energía en cada vuelta del ciclo. La reacción neta es:
Acetil CoA + 3NAD+ + FAD + ADP + Pi + 2H2O → 2CO2 + 3NADH + FADH2 + ATP + 2H+ + CoA
Considerando que de cada molécula de glucosa se forman dos moléculas de piruvato y, por consiguiente, dos de acetil-CoA, habría que multiplicar esta reacción y la catalizada por la piruvato deshidrogenasa por dos.
Como resumen final podemos considerar que el ciclo de Krebs cumple tres funciones principales en plantas.
1. Producción de donadores de electrones como el NADH y el FADH2 que,
al ser posteriormente oxidados en la cadena transportadora de electrones, darán lugar a la formación de una considerable cantidad de energía.
2. Síntesis de una cantidad limitada de ATP.
Fig 5. Ciclo de Krebs.
Cadena de Transporte de electrones
Todos los equivalentes de reducción originados en la glucólisis o ciclo de Krebs van a ser oxidados en la cadena mitocondrial de transporte de electrones. Esta cadena consta de una serie de transportadores de electrones que, mediante reacciones de óxido-reducción, van a llevar los electrones hasta el último aceptor de los mismos, el O2, formándose H2O como producto final. El transporte de
flavoproteínas, proteínas conteniendo hierro y azufre similares a la ferrodoxina, ubiquinona, citocromos tipo b, citocromos tipo c y citocromos tipo a.
Fig 6. Cadena transportadora de electrones.
La determinación cuantitativa de la presencia de cada uno ha llevado a la conclusión de que existen en cantidades similares, lo que lleva a pensar que todos los componentes están asociados estructuralmente, formando algún tipo de unidad funcional. La cadena respiratoria está unida a elementos estructurales de la membrana, que parecen ser necesarios para mantener a los componentes de la cadena en el orden correcto que les permite reaccionar a unos con otros. El compuesto intermedio del ciclo de Krebs es oxidado con la cesión de dos átomos de hidrógeno en la reacción. Los dos electrones que poseía el hidrógeno son transportados a lo largo de la cadena de enzimas (del grupo de los citocromos) hasta llegar al oxígeno. Por cada par de electrones transportados por esta cadena se producen tres moléculas ATP.
Rendimiento energético del proceso respiratorio
Una vez conocido el mecanismo de la glucólisis, el ciclo de Krebs y la cadena de transporte de electrones, podemos calcular cuantos ATP se forman en la oxidación completa de la glucólisis. La reacción total es la siguiente:
Glucosa + 36ADP + 36Pi + 6O2 → 6CO2 + 36 ATP + 42H2O
La relación P/O es de tres, puesto que se forman 36ATP y se consumen 12 átomos de O2. Se forman 36 moléculas de H2O que son producidas al sintetizar
las 36 moléculas de ATP. De estas 36 moléculas de ATP, 32 se formarán por fosforilación oxidativa. En la Tabla 1, podemos ver un esquema del rendimiento energético de la respiración celular.
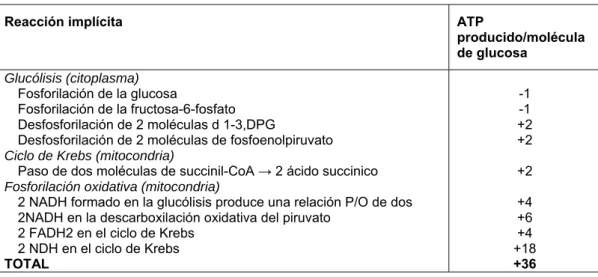
Tabla 1. Rendimiento energético de la respiración celular.
Reacción implícita ATP
producido/molécula de glucosa
Glucólisis (citoplasma)
Fosforilación de la glucosa
Fosforilación de la fructosa-6-fosfato Desfosforilación de 2 moléculas d 1-3,DPG
Desfosforilación de 2 moléculas de fosfoenolpiruvato
Ciclo de Krebs (mitocondria)
Paso de dos moléculas de succinil-CoA → 2 ácido succinico
Fosforilación oxidativa (mitocondria)
2 NADH formado en la glucólisis produce una relación P/O de dos 2NADH en la descarboxilación oxidativa del piruvato
2 FADH2 en el ciclo de Krebs 2 NDH en el ciclo de Krebs
TOTAL -1 -1 +2 +2 +2 +4 +6 +4 +18 +36
La eficiencia total de la generación de ATP es alta, ya que la oxidación de la molécula de glucosa produce 686 kcal, y la energía liberada almacenada en el ATP es de 263 kcal. Por tanto la eficiencia termodinámica de la formación de ATP a partir de glucosa es de 38% bajo condiciones estándar.
Cociente respiratorio
Cuando se mide la respiración es importante conocer el O2 consumido como el
CO2 producido, esta relación recibe el nombre de cociente respiratorio. CR= CO2/O2
En general cuando un glúcido es respirado en la célula se consume una molécula de O2 por cada molécula de CO2 producida, siendo este cociente igual
a uno. Las sustancias del ciclo de Krebs están más fuertemente oxidadas que los glúcidos y, en consecuencia, necesitan menos oxígeno para su oxidación a CO2 y agua, dando valores de CR superiores a uno. Por ejemplo: la oxidación de
Las grasas están más reducidas que los glúcidos, por lo cual es necesario más oxígeno para oxidarlas en la respiración, la respiración de una grasa puede dar valores de CR tan bajos como 0,7.
El CR es importante para la investigación porque puede dar una idea del sustrato que está siendo respirado, si embargo debe considerarse que el CR expresado solo será el promedio de los distintos sustratos que deben estar siendo respirados en un momento determinado. En plantas adultas el CR de distintos órganos se mantiene siendo casi en su totalidad derivado de glúcidos, sin embargo, si la planta es sometida a condiciones deficientes, puede faltar sustancias de reserva, mostrando valores menores a uno.
Regulación de la Respiración en las plantas
Se sugiere que la tasa de respiración fluctúa de acuerdo a las demandas energéticas celulares a través de la regulación por retroalimentación de la glucólisis y del transporte electrónico por el cociente ATP/ADP en el citosol. Sin embargo las plantas presentan otros mecanismos (no fosforilantes) para degradar la glucosa, la situación in vivo puede no ser tan simple.
Las vías no fosforilantes actúan como “válvulas de exceso de flujo” de la vía respiratoria principal y serán activadas in vivo solo cuando los niveles de azúcar
sean elevados y el flujo glucolítico sea rápido. Las vías de la glucólisis en plantas están siendo investigadas y su estudio se ve complicado debido a la duplicidad del sistema en los plastidios.
Factores que afectan la Respiración
• Temperatura: Tiene una influencia directa sobre los procesos bioquímicos, debido a que las reacciones enzimáticas están reguladas por la temperatura existen rangos estrechos entre los cuales pueden tener lugar. A temperaturas cercanas a 0ºC la intensidad de la respiración se hace muy baja. Al subir la temperatura las actividades respiratorias se aceleran hasta llegar a niveles en que por su acción se destruye la actividad de las enzimas. Se alcanza una intensidad máxima aproximadamente entre 35 y 45ºC. Se debe tener en cuanta también el tiempo de exposición a la temperatura; según parece, temperaturas mayores a 30ºC, empiezan a actuar con un efecto adverso produciéndose la desnaturalización de las enzimas. Puesto que a estas temperaturas el efecto no es inmediato primero se ve un incremento en la respiración, que con el paso del tiempo descenderá rápidamente. A mayor temperatura, menor será el tiempo para que se anule la respiración.
• Oxígeno: El oxígeno es importante para las reacciones del ciclo de Krebs, a demás es el último aceptor del sistema de transporte de electrones. En general a bajas concentraciones de O2 se espera que existan procesos
aeróbios y anaeróbios dentro del sistema, con la producción de CO2
incrementan las concentraciones de O2 la respiración anaeróbica disminuye
hasta llegar a un punto de extinción.
• Anhídrido Carbónico: Un aumento del CO2 tiene un claro efecto negativo en
en la respiración. Aunque existen estudios que señalan un efecto depresivo debido al CO2, hay pruebas de que este efecto puede ser parcialmente
indirecto. Estudios han demostrado que el CO2, puede provocar el cierre de
los estomas, limitado así el intercambio gaseoso. Esto puede repercutir en una elevación considerable de la concentración interna de CO2 y, con ello, en
una limitación de la respiración.
• Sales inorgánicas: Observaciones indican que la respiración puede aumentar cuando una planta o un tejido se hacen pasar desde agua a soluciones salinas. La cuantía de la respiración por encima de la normal se ha denominado respiración “salina”.
• Estímulos mecánicos: Estudios han demostrado que la respiración puede aumentar si las hojas se manipulan, si se golpeaban o se enrollaban. La respuesta a la manipulación decrece si se repite este proceso por cierto periodo de tiempo. Barrer, encontró que la respiración de los tubérculos de papa aumentaba después de su manipulación.
Transpiración
El agua es la sustancia más abundante en los tejidos vegetales, sin embargo las plantas retienen una pequeña porción del agua que absorben y que pasa a través de ellas durante su ciclo de vida. Están absorbiendo grandes cantidades de agua que pasa por los sistemas de transporte y sale a la atmósfera. A pesar de necesitar grandes cantidades de agua las características anatómicas de su estructura foliar son tales que causan la pérdida continua de grandes cantidades de agua.
Pierden agua sobre todo en forma de vapor, a través del proceso de transpiración. Después de ser sacada por las raíces el agua es transportada a lo largo del xilema hasta llegar a las células del mesófilo de la hoja. La disposición de estas células de membrana fina proporciona espacios intercelulares abundantes, lo que representa una disposición ideal para la evaporación del agua a partir de la superficie celular. Una gran parte de la superficie epidérmica está constituida por un gran número de poros llamados estomas. Los poros
estomáticos se abren a los espacios intercelulares de la hoja y el medio externo (fig 7).
Fig 7. Transporte de agua en las plantas.
provoquen alta transpiración. En general esta capa es muy gruesa en las hojas muy expuestas al sol y en las plantas de habitats secos. Sin embargo la cantidad de agua perdida en la transpiración cuticular y en la lenticular es insignificante a la pérdida por transpiración estomática.
Los estomas son de tamaño microscópico y están rodeados por dos células epidérmicas especializadas llamadas células oclusivas o subsidiarias, que regulan la apertura y el cierre de los estomas (fig 8). Según la especie, la superficie de una hoja puede tener entre 1.000 - 60.000 estomas por centímetro cuadrado. A pesar de su número, abiertos solo ocupan del 1 al 2% de la superficie total de la hoja. Por lo general se encuentran en el envés de la hoja, pero algunas especies se encuentran en ambas caras. Los estomas están presentes en todas las angiospermas y las gimnospermas, exceptuando las plantas acuáticas.
Fig 8. Estomas, mostrando células guardián, subsidiarias y ostiolo.
Los poros de los estomas pueden ser considerados como puertos de intercambio entre el medio externo y el interior de la hoja. Por ello los factores físicos que influyen sobre la difusión de vapor de agua a través de dichos poros son importantes en el estudio de la transpiración. Aunque, cuando están abiertos al máximo, el área total solo es del 1-2% de la superficie foliar, la difusión de vapor de agua a través de los poros supera con frecuencia el 50% de la evapotranspiración de una superficie libre. Estudios realizados indican que la difusión por un poro circular pequeño es más aproximadamente proporcionada al perímetro o al diámetro del poro que a su área.
Los factores del ambiente que tienen una mayor influencia sobre la apertura y cierre de los estomas son la luz, el agua, y la temperatura.
reduce la turgencia y, por tanto, el estoma se cierra total o parcialmente. Cuando mayor es el déficit de agua, más pronto se cierran los estomas.
Cuando los demás factores son constantes, se observa que un incremento de la temperatura provoca un aumento de la apertura estomática.
Factores que influyen sobre la velocidad de la Transpiración
Se conocen otros aspectos, a parte de la apertura estomática, que tienen influencia sobre la transpiración. La relación entre la parte aérea y la radicular, y el área y la estructura de la hoja tienen una influencia considerable sobre la pérdida de agua por las plantas. Cualquier factor del medio ambiente que afecte la intensidad del gradiente de presión de vapor entre la atmósfera interna y externa de la hoja afectará la intensidad del gradiente de presión de vapor entre la atmósfera interna y la externa de la hoja afectará de modo evidente la velocidad de transpiración. Finalmente, la disponibilidad del agua en el suelo influirá también sobre la transpiración.
En caso de existir las condiciones para la transpiración, la eficacia de la superficie absorbente (superficie radical) y la de la superficie de evaporación (superficie foliar) regulan la velocidad de la transpiración. Si la absorción de agua es menor que la transpiración, se produce un déficit de agua que reducirá la transpiración. El área foliar si es mayor, mayor será la pérdida de agua por transpiración.
La estructura foliar en plantas de climas secos presenta por lo general modificaciones estructurales, principalmente en sus hojas. La resistencia de hojas xerofíticas a la perdida de agua y a la marchitez es debida en primer lugar al espesor y la eficacia de la capa de cutina que las recubre. En tiempo seco, los estomas se mantienen cerrados y la transpiración cuticular se convierte en la única posibilidad de pérdida de agua.
Fig 9. Comparación de la transpiración con aire en movimiento y estático, según la abertura del estima..
Importancia de la Transpiración
Los estomas se abren en presencia de luz, para absorber el CO2 requerido en la fotosíntesis; aunque el balance hídrico se altere, al escaparse el agua de la planta. El flujo de agua a través de la planta inducido por la transpiración, provee un buen sistema de transporte para los minerales, que son absorbidos por las raíces y que se mueven en la corriente transpiratoria. Así mismo, la absorción de agua del suelo, tiene un efecto en la movilización de sales minerales del suelo hacia la raíz, facilitando su absorción, sin un gasto de energía adicional, que implicaría la formación de masas de raíces que exploren amplias superficies de suelo.
Otro efecto de la transpiración es la acción refrigerante de la hoja. La evaporación de agua de la superficie foliar, va acompañada por una perdida de calor. El calor de evaporación del agua es aproximadamente 600 cal. g,-1 esta pérdida de calor ayuda a mantener una temperatura adecuada de la hoja, durante días muy soleados. La reducción de temperatura foliar por transpiración esta en el orden de 2-3°C por debajo de la temperatura del aire. Podemos concluir que la transpiración ejerce un efecto de enfriamiento de la superficie foliar.
Se ha sugerido que la transpiración es necesaria para el crecimiento normal de las plantas, ya que ayuda a mantener un estado de turgor óptimo. Cuando las plantas crecen en una atmósfera saturada de humedad, presentan un aspecto suave y carnoso, que puede ser el resultado de una gran absorción de agua, que causa un mayor alargamiento celular. Las plantas terrestres casi nunca están en un estado de turgor óptimo, aunque la savia celular pueda tener una presión osmótica alta, como en algunas halófilas, la pérdida de agua por transpiración mantiene la presión de turgor por debajo de la presión osmótica.
Abertura del estoma
Transpiración
Balance Hídrico de la Planta
Los procesos básicos que determinan el balance hídrico de una planta son: la absorción, conducción y pérdida de agua. Si queremos determinar que también se equiparan la acumulación y la pérdida de agua debemos medir la absorción y la evaporación en un intervalo de tiempo determinado. La diferencia entre la absorción y la transpiración, nos indica el balance hídrico, o cualquier desviación de la condición de equilibrio. El balance hídrico de una planta oscila de valores positivos a negativos. Ocurren fluctuaciones a corto plazo determinadas por las variaciones en la apertura estomática, en tiempos cortos de pocas horas, sin embargo variaciones que se alejan más del equilibrio pueden ocurrir durante el día, especialmente durante la fase diurna y nocturna.
Transporte a través de las membranas
Las membranas biológicas son totalmente permeables a pequeñas moléculas no cargadas, como agua y O2. Estas moléculas poseen una elevada solubilidad en
lípidos y pueden pasar a través de la bicapa lipídica. Por el contrario, moléculas mayores no cargadas como la sacarosa, o moléculas cargadas, como los iones inorgánicos o los ácidos orgánicos, no pueden atravesar la bicapa lipídica y su paso a través de las membranas se produce por medio de canales o transportadores, de los que cada membrana posee su propio y único conjunto. La energía para realizar el transporte de solutos y para mantener los gradientes electroquímicos la proporciona la ATPasa.
Las células vegetales deben ser capaces de captar nutrientes a través de la membrana plasmática y de mantener fuera los solutos no deseados. Dentro de las células vegetales existen compartimientos (citoplasma, vacuola, cloroplasto, etc.), que poseen una composición única y hay un intercambio de continuo de solutos a través de sus respectivas membranas. A excepción de la vacuola, la composición del compartimiento se mantiene dentro de límites bastante estrechos, fenómeno conocido como homeostasis. Para conseguir esto deben regularse los flujos de los solutos.
Para un soluto no cargado, la fuerza motora para la difusión a través de una membrana es el gradiente de concentración a través de la membrana. En el equilibrio, la concentración de un soluto no cargado es la misma a ambos lados de la membrana, independientemente del potencial de membrana.
Para un soluto cargado (de acuerdo a la ecuación de Nernst) en equilibrio, la diferencia de concentración de un soluto a través de una membrana es contrarrestada por una diferencia de voltaje entre los compartimientos.
Una característica de los sistemas biológicos es que no están en equilibrio, los organismos vivos gastan una considerable cantidad de energía manteniendo una composición radicalmente diferente de la que los rodea. En el caso de las membranas biológicas, la energía química se convierte en un gradiente electroquímico de protones utilizado para transportar otros solutos a través de la membrana.
El gradiente electroquímico de protones consiste en un gradiente de concentración, o una diferencia de pH, y un potencial de membrana, cuyo valor relativo depende de la membrana. Por ejemplo, en presencia de luz, el gradiente electroquímico de protones en un cloroplasto es, casi exclusivamente, un gradiente de concentración con pH 4 en el lumen de los tilacoides y pH 8 en el estroma del cloroplasto.
Tabla 2. Clasificación de las proteínas de transporte basada en el mecanismo y en la energía de transporte.
_______________________________________________________________________________
1. Transportadores
1.1. Transportadores Activos
1.1.1. Transportadores activos primarios
1.1.1.1. Acoplados a reacciones redox (p ej., citocromo c oxidasa) 1.1.1.2. Acoplados a la luz (p ej., bacteriorrodopsina)
1.1.1.3. ATPasas (p ej., ATPasa-H+) 1.1.2. Transportadores activos secundarios
1.1.2.1. Simportes (p ej., transportador de sacarosa) 1.1.2.2. Antiportes (p ej., transportador de Na+)
1.2. Uniportes Pasivos (p ej., transportadores de glucosa de eritrocitos) 2. Canales (o poros)
2.1. Regulados por voltajes (p ej., canal de K+)
2.2. Regulados químicamente (p ej., mediante hormonas) 2.3. Otros (p ej., sensibles a la presión)
Tomado de: AZCON-BIETO, J. Fisiología y Bioquímica Vegetal. p.34.
Homeostasis: Regulación de pH y concentración de iones
La Homeostasis es la capacidad para permanecer constante. El medio ambiente puede cambiar constantemente pero la célula tiende a mantener las condiciones en el interior e la membrana plasmática más o menos constante. Esto es muy importante en las plantas por su imposibilidad de moverse frente a condiciones adversas.
pH
Las proteínas son sensibles a cambios en las concentraciones de protones (el pH) en su medio circundante y la actividad de las enzimas muestra a menudo una fuerte dependencia del pH, por tanto el controlarlo es muy importante para la célula. El pH en la solución del suelo varía dependiendo del tipo de suelo y de otras condiciones, pro sueñe estar en el intervalote 5-6. El pH del citoplasma es normalmente entre 7 y 8 y es relativamente constante. Por el contrario el pH de la vacuola es de 4-6 y se han observado variaciones mucho mayores. Las plantas CAM constituyen un ejemplo extremo donde el pH de la vacuola cambia desde 6,0 en la luz hasta 3,5 en la oscuridad, debido a la acumulación de ácido málico, uno de los primeros productos de la fijación del CO2 en estas plantas.
Ca2+
La concentración de Ca2+ libre se mantiene extremadamente baja en el
citoplasma. A pesar de que la concentración en el suelo y en la pared celular es superior, al menos, tres órdenes de magnitud. Existen dos razones principales por las que la concentración debe ser baja:
a. A elevadas concentraciones el Ca2+ forma complejos insolubles con
fosfato
b. Muchos procesos están regulados por la concentración de Ca2+ libre y una baja concentración puede modificase mucho más de prisa
Para mantener una concentración de Ca2+ baja la membrana tiene que tener una baja permeabilidad para el Ca2+, así como un método muy eficiente para
bombearlo hacia afuera. Uno de estos métodos es la ATPasa Ca2+ que acopla
la extracción de Ca2+ a la hidrólisis de ATP. Otro mecanismo para eliminar el
Ca2+ del citoplasma se realiza mediante un antiporte Ca2+/H+ en la membrana
vacuolar.
Se conoce relativamente poco a cerca del papel del transporte de Ca2+
mitocondrial en la homeostasis del Ca2+ citoplasmático en células vegetales. Sin
embargo, dado que uno de los puntos de entrada en la cadena de trasporte electrónico de la mitocondria vegetal, la NAD(P)H deshidrogenasa insensible a la rotenona, en la superficie de la membrana mitocondrial interna, es dependiente del Ca2+, se requiere alguna forma de regulación de la concentración de Ca2+ en
el interior de la mitocondria.
Nitrato y Fosfato
Debido a que el nitrato y el fosfato son nutrientes esenciales y a que su concentración en la solución de suelo es, a menudo, limitada por el crecimiento celular, las plantas han desarrollado mecanismos muy eficientes para acumular estos iones. Así, en suelos bien fertilizados, las plantas contienen grandes cantidades de nitrato y fosfato, encontrándose la mayor cantidad en la vacuola, de donde pueden ser movilizados cuando son necesarios. En condiciones de suelo pobre los nutrientes son movilizados, desde la vacuola hasta el citoplasma, donde son utilizados. La vacuola puede quedar totalmente desprovista de ellos, mientras que todavía se encuentran en concentraciones normales en el resto de la célula.
Hormonas Vegetales
El desarrollo normal de una planta depende de la interacción de factores externos: luz, nutrientes, agua y temperatura; e internos: hormonas. Las hormonas vegetales (o fitohormonas) son sustancias producidas en un tejido y transportadas a otro, donde producen unas respuestas fisiológicas determinadas. Son activas en muy bajas concentraciones. Presentan una función fisiológica concreta, y se transportan fácilmente a través de los vasos conductores.
Las principales hormonas vegetales o fitohormonas, se agrupan de acuerdo al tipo de receptor celular que presenten o su función. Las más importantes son:
• Auxinas
• Citoquininas
• Giberelinas
• Ácido abcísico
• Etileno
Mientras que cada fitohormona ha sido implicada en un arreglo relativamente diverso de papeles fisiológicos dentro de las plantas y secciones cortadas de éstas, el mecanismo preciso a través del cual funcionan no es aún conocido. Cabe decir que muchas respuestas de la planta, no se deben a la actuación de una hormona concreta, sino a la interacción de varias, y que actualmente, se está investigando en el descubrimiento de lo que podrían ser nuevas hormonas, pero que hoy por hoy, se les prefiere llamar reguladores del crecimiento.
A continuación el la tabla 3 se presenta un resumen de las distintas funciones que presentan las hormonas.
Tabla 3. Funciones de las fitohormonas en la planta.
Efecto fisiológico Auxinas Giberelinas Citoquininas Ácido Abscísico
Etileno
Respuestas trópicas Sí Sí No Sí Sí
Aumento del tamaño celular en
cultivos de tejidos Sí, en algunos casos Sí, en algunos casos Sí No No
Control de la diferenciación en el
cultivo de tejidos Sí Sí Sí Sí Sí
Estimula el enraizamiento Sí No Respuesta muy
variable Sí, en algunos casos Sí
Inhibe el desarrollo radicular Sí No Se desconoce Puede inhibirlo No
Estimula la división del cambium Sí Sí Sí Puede inhibirla No
Abscisión de hojas y frutos Sí No Sí Sí Sí
Activa el crecimiento de frutos Sí Sí Sí, en algunos
casos No No
Afecta al crecimiento del tallo No Sí, lo activa No Lo inhibe Lo inhibe
Interrumpe el reposo de las yemas
vegetativas No Sí Sí No, lo induce Sí, en algunos casos Favorece la germinación de
algunas semillas No Sí No No, la inhibe en general Sí, en algunos casos Favorece la síntesis de
alfa-amilasa en granos de cereal No Sí Sí No, la inhibe No Mantenimiento de la dominancia
apical Sí Sí Sí, en algunos casos Se desconoce Sí
Inhibe la degradación de proteínas
y de clorofila en la senescencia Sí, en algunos casos Sí Sí, en algunos casos No, la acelera No, la acelera
Activa el pico climatérico de la respiración de frutos en el proceso
de maduración
Se desconoce No No No Sí
Cuando la planta germina, comienzan a actuar algunas sustancias hormonales que regulan su crecimiento desde esa temprana fase: las fitohormonas, llamadas giberelinas, son las que gobiernan varios aspectos de la germinación; cuando la planta surge a la superficie, se forman las hormonas llamadas auxinas, las que aceleran su crecimiento vertical, y, más tarde, comienzan a aparecer las citocininas, encargadas de la multiplicación de las células y que a su vez ayudan a la ramificación de la planta (fig 10).
Fig. 10. Distribución, a modo amplio, del efecto de las fitohormonas dentro de la planta.
•
Auxinas.
Es el primer grupo de hormonas vegetales que se descubrió, y su conocimiento data de la época de Charles Darwin quien refleja la existencia de este tipo de sustancias en su libro “Power of movement in plants”, aunque por aquel entonces no se las hubiera bautizado todavía.
Fig. 11. Experimento de Went, corte del coleóptido de avena.
Efectos:
- Dominancia del brote principal e inhibición de la ramificación lateral - Estimulación del crecimiento apical de toda la planta
- Diferenciación de los vasos conductores (xilema y floema)
- Estimulación de la formación de raíces adventíceas (Importante en la plantación de esquejes)
- Promueve el crecimiento del fruto
- Inhibición de la caída de las hojas y de los frutos
- Produce el gravitropismo (crecimiento en función de la fuerza de gravedad), en combinación con los estatocitos (células especializadas en detectar la fuerza de gravedad, por contener amiloplastos)
- Puede actuar como herbicida
Si las auxinas están implicadas en la proliferación celular quiere decir que intervienen en la mitosis o división celular. Concretamente, las auxinas están implicadas en la síntesis de unas determinadas moléculas (ciclinas) sin cuya presencia las células no pueden entrar en mitosis. Si aplicamos auxinas a una concentración de 10-6 o 10-9 molar se estimula la síntesis de dicha ciclina y la
entrada de las células en mitosis, mientras que si la concentración supera los 10-6 molar se inhibe su síntesis (fig 12).
Uno de los fenómenos más curiosos observables en las plantas son los tropismos, es decir, los cambios que experimentan los vegetales ante un estímulo (luminoso, gravedad, táctil):
En el fototropismo, el mecanismo de acción de las auxinas es simple y depende de las zonas donde incida con más fuerza la radiación del azul. En estas zonas, se produce un aumento de la concentración de auxinas y es hacia esa zona donde se dirige la planta, debido a que las auxinas hemos dicho que estaban implicadas en la estimulación del crecimiento apical.
•
Citoquininas
Son hormonas derivadas de la adenina (base nitrogenada del DNA) y están relacionadas principalmente con los procesos de división celular (mitosis), aunque también actúan a otros niveles.
Efectos:
- En combinación con la auxina, regula la morfogénesis (formación de tejidos) en cultivos de tejidos.
- Retrasan la senescencia (envejecimiento de las hojas), al retrasar la inactivación del ADN permiten la síntesis de clorofila.
- Transporte de sustancias a nivel de floema
- Estimulación de la pérdida de agua por transpiración - Retraso de la senescencia (envejecimiento) de las hojas - Activación del crecimiento de las yemas laterales
- Eliminación de la dormancia que presentan las yemas y semillas de algunas especies
- Inducción a la partenocarpia de algunos frutos - Estimulación de la formación de tubérculos
Se conocen con el nombre de “hormonas juveniles”, debido a que evitan el envejecimiento (senescencia) prematuro de la planta y se empezaron a estudiar en el año 1954. Se descubrieron a partir de los tumores que provocan en las plantas la acción de determinados microorganismos como Agrobacterium tumefaciens.
Las citoquininas se sintetizan, sobre todo, en la zona meristemática de la raíz de la planta, las auxinas que lo hacían en las partes aéreas jóvenes. Desde la raíz, son transportadas en una forma llamada conjugada a través del xilema a toda la planta.
Fig. 13. Comparación entre la concentración de cuatro tipos de citoquininas (mol) en relación al porcentaje de elongación celular.
•
Giberelinas
Se encuentran en todos los órganos, pero sobre todo en las semillas inmaduras. La más conocida es el ácido giberélico.
Efectos
- Producen un incremento en el crecimiento del vástago. - Estimulan la división celular y afectan a hojas y tallos.
- En plantas con morfología juvenil diferente de la adulta, modifican esta última y vuelve a la juvenil.
- Inducen la floración en algunas plantas en roseta.
- estimulan la germinación del polen y pueden producir frutos partenocárpicos.
- Sustitución de las necesidades de frío o de día largo requeridas por muchas especies para la floración.
- Eliminación de la dormancia que presentan las yemas y semillas de numerosas especies de vegetales.
- Retraso en la maduración de ciertos frutos, especialmente los cítricos. - Inducción del alargamiento de los entrenudos en los tallos.
Se conocen en la actualidad más de 125 hormonas diferentes de este grupo. Se descubrieron realizando un estudio de un extracto del hongo (Gibberellum fugikunoi) responsable de la enfermedad bakanae en los cultivos de arroz. Dicha
enfermedad se caracteriza porque todas las plantas de arroz presentan acame. Al analizar el extracto, se encontró ácido giberélico y de ahí, que a este grupo de hormonas se las conozca como giberelinas.
Las giberelinas se encuentran en cantidades particularmente abundantes en órganos jóvenes de las plantas, especialmente en los puntos de crecimiento del vegetal (zonas apicales) y en las hojas jóvenes en proceso de formación. Algunas se mueven libremente por la planta, pero en algunos casos, parecen estar muy localizadas. El desplazamiento de las giberelinas parece ser debido a un transporte meramente pasivo.
•
Ácido abcísico
El ácido abscísico (ABA) es la última hormona descubierta por los fisiólogos en las plantas. Se caracteriza por inhibir muchos fenómenos de crecimiento en las plantas superiores, y específicamente, por estar asociado a la dormancia de yemas y semillas, así como también por causar la caída (abscisión) de las hojas. Es un compuesto derivado del ácido mevalónico y su biosíntesis tiene lugar en: frutos, semillas, raíces, hojas y tallos.
Está relacionado con la capacidad de ciertas plantas para restringir su crecimiento o su capacidad reproductora en épocas desfavorables. Se ha comprobado que las hojas de las plantas experimentan un aumento considerable en la producción de ABA cuando están ante una situación de estrés hídrico. Así como también se ha comprobado, que encharcamientos en las raíces, frío y ciertas alteraciones patológicas estimulan su síntesis.
El ABA se transporta rápidamente a toda la planta, tanto a través del xilema como del floema.
Efectos:
- Regulación de la apertura estomática, de modo que una aplicación exógena de dicha hormona comporta el cierre de los estomas.
- Dormancia de yemas y semillas en climas fríos. - Abscisión de hojas y frutos.
- Inhibición de la síntesis de RNA y proteínas.
- Inhibición del crecimiento de muchas partes de la planta. - Inhibe el crecimiento de los tallos.
- Induce la senescencia de las hojas.
Fig. 14. Giberelinas como promotores y el ácido abscísico como inhibidor de la germinación del a semilla
•
Etileno
Se conoce desde hace mucho tiempo que cantidades muy pequeñas de este gas afectan al crecimiento vegetal: senescencia y abscisión de las hojas, así como la maduración de algunos frutos. Es la única hormona vegetal conocida, hasta el momento, que se presenta en estado gaseoso en condiciones normales de presión y temperatura.
Si consideramos a toda la planta, nos daremos cuenta que la mayor cantidad de etileno se sintetiza a nivel de las flores y frutos. Precisamente en estos últimos, observamos un fenómeno, el incremento progresivo de la concentración de etileno conforme van madurando.
Se observa un incremento en la concentración de etileno cuando se somete a la planta a situaciones de estrés.
Efectos:
- Estimulación del crecimiento de las raíces.
- Inhibición del transporte de auxinas en el interior de la planta.
- Estimulación de la síntesis de algunas enzimas o la liberación de alfa-amilasa ya formada, por ejemplo, en granos de cereales durante la germinación.
- Inducción de la maduración de los llamados frutos climatéricos. Maduración anticipada de algunos frutos (plátano, tomate, cítricos, etc...) mediante la aplicación de etileno.
- Eliminación de la dormancia de yemas y de algunos órganos vegetativos, tales como bulbos y tubérculos.
- Promueve la caída de hojas, flores y frutos (abscisión). - Produce curvatura de las hojas hacia abajo (epinastia).
- Induce la formación de raíces en hojas, tallos y pedúnculos florales. - Induce la feminidad en flores de plantas monoicas (las que tienen flores
masculinas y femeninas sobre el mismo individuo).
Fotoperíodo y Vernalización
Fotoperíodo
El requisito previo para la respuesta de una planta a la luz es que esta sea absorbida. Esto implica la necesidad de un aceptor de algún tipo (generalmente un pigmento), que debe estar presente y ser capaz de absorber la longitud de onda de luz responsable de la respuesta. En muchos casos, la absorción de luz por el receptor hace que éste se vuelva más reactivo, lo cual a su vez desencadena una sucesión de reacciones químicas que conducen en último término a una respuesta general de la planta.
El término fotoperiodicidad escapa de una definición precisa. En general, se le define como respuesta de una planta a la longitud relativa de los períodos de luz y oscuridad. Sin embargo, esta definición puede modificarse en varios aspectos. La intensidad y la composición de la luz pueden ser características que influyen sobre la magnitud de la respuesta. La cantidad total de luz recibida puede tener también influencia.
Se acepta de modo general que la duración y el orden de la secuencia es especialmente importante en la iniciación de una respuesta fotoperiódica. Entonces, cualquier respuesta realizada por una planta ante la duración y el orden de alternancia de períodos de luz y oscuridad puede denominarse
respuesta fotoperiódica.
Las plantas responden a alteraciones de los períodos de luz y oscuridad de maneras distintas. La floración, el crecimiento vegetativo, el alargamiento de los entrenudos, la germinación de la semilla y la caída de las hojas constituyen algunos casos de respuestas fotoperiódicas que han sido descubiertos en las plantas.
En algunas plantas, los fotoperíodos de días largos inducen a la floración, mientras que otras parecen no responder a ellos, floreciendo indiferentemente en condiciones de día largo o corto. Otras responden a fotoperíodos situados entre los de día largo y los de día corto. Las condiciones dadas a continuación se basan en un ciclo de luz y oscuridad de 24 horas.
• Una planta de días cortos florece cuando la longitud del día es inferior a cierta longitud crítica. Las condiciones de luz que superen esta longitud crítica mantendrán a la planta de días cortos en estado vegetativo. La denominada longitud crítica del día es distinta según las especies.
• Una planta de día largo florece cuando se sobrepasa una cantidad de horas de luz crítica. También en este caso, la longitud crítica de a exposición a la luz difiere de una a otra especie.
longitud crítica. El factor que delimita los dos grupos no es el número de horas de luz recibida sino cuando florecerá una planta, si antes o después de un período de luz crítico (fig 16).
Fig. 16. Diferencias de respuesta ante el fotoperíodo de tres especies que presentan distinta respuesta al mismo.
Aunque la longitud de fotoperíodo no tenga ninguna influencia sobre la aparición de flores, puede comprobarse que presenta una influencia de tipo cuantitativo. Al aumentar la longitud del fotoperíodo se obtiene un incremento del número de primordios florales. Mientras que la longitud del período de oscuridad determina la iniciación real de los primordios florales, la longitud del periodo de luz determina el número de primordios florales que van ha ser producidos.
Un espectro de luz correspondiente a la efectividad de las distintas longitudes de onda de luz demuestra que las más eficaces de cara a la inhibición de la floración se encuentren entre 620 660 um (anaranjado-rojo) con un máximo
situado aproximadamente a 640 um. Por ello, la luz roja se considera la radiación
más efectiva para las reacciones de interrupción luminosa.
Vernalización
En muchas plantas la temperatura tiene una profunda influencia sobre la iniciación y el desarrollo de las estructuras reproductivas. Muchas plantas de día largo requieren una exposición a temperaturas entre 0 a 10 ºC (vernalización) para florecer tempranamente. Esta dependencia de la temperatura asegura que variedades o especies que comienzan su desarrollo vegetativo en otoño, no florezcan hasta pasado el invierno. Al igual que para el fotoperiodo el requerimiento de vernalización puede ser más o menos absoluto dependiendo de la especie y del genotipo de que se trate. Mediante la vernalización independiente de distintos órganos de la planta, se ha podido demostrar que únicamente la vernalización del meristema apical es capaz de provocar la inducción de la transición floral.
Esto parece indicar que el tratamiento de vernalización no conlleva la producción de un estímulo floral transmisible. En Thlaspi varvense, una especie de día largo
con requerimiento de vernalización, las plantas regeneradas a partir de discos de hoja vernalizadas florecen tempranamente sin necesidad de pasar por el proceso. Estos resultados sugieren que el efecto de la exposición a temperaturas bajas no está limitado a las células meristemáticas, aunque la aceleración de la transición floral que desencadena está limitada al meristemo apical, por ser esta la región de crecimiento de la planta.
El requerimiento de vernalización que demuestra Arabidopsis y otras especies de día largo puede sustituirse por tratamientos exógenos con giberelinas, lo que sugiere que estos reguladores de crecimiento también podrían mediar en el proceso de vernalización. En Thlaspi varvense, se ha demostrado que la
exposición a temperaturas bajas, desbloquea la síntesis de toda una familia de giberelinas, cuando las plantas retornan a temperaturas normales. Mientras muchas giberelinas como GA20 son transportadas a lo largo de la planta y podrían formar parte de un estímulo floral transmisible, existen otra que no se movilizan y podrían participar en estímulo no transmisible generado por las bajas temperaturas.
Interacción entre Fotoperíodo y Vernalización
Por lo general en especies de día largo existe una interacción entre el efecto de fotoperíodo y el efecto de vernalización. El resultado de la exposición a ambos factores ambientales suele corresponder a la suma del efecto de cada uno de los factores hasta un límite de saturación determinado, a partir del cual tratamientos más prolongados no producen un efecto mayor.
REFERENCIAS
AZCON-BIETO, J. (1993). Fisiología y Bioquímica Vegetal. Madrid. Interamericana. Mc Graw Hill. Pp.581.
BARCELO COLL, J. (2OO1). Fisiología Vegetal. Madrid. Edic. Pirámide. Pp. 141-144, 153-157, 162-163, 165-169, 177, 187-191, 203-208, 216-225.
BERNSTEIN,R. y S., BERNSTEIN (1998). Biologia. Santa Fe de Bogota. Mc Graw Hill. 10° edic. pp. 114-127.
DAVLIN, R. (1982).Fisiología Vegetal. Barcelona. Omega. Pp. 577 pp.
HORTON, R. H. (1995). Bioquímica. México D.F. Prentice Hall. Pp. 16-2 a 16-5, 16-27 a 16-29.
M. DEVLIN, R. (1982). Fisiología vegetal. Barcelona. Edic. Omega. Pp. 189-194. PANIAGUA, R. y otros (1999). Biología celular. Madrid. Mc Graw Hill. Pp. 226-229, 232-234.
PRESCOTT y otros. (1999). Microbiologia. Madrid. Mc Graw Hill. 4° edic. Pp. 187-193.
SOLOMON PEARL, E y otros. (2001). Biología. México D.F. Mc Graw Hill. 5° edic. Pp. 180-194.