USO Y PREFERENCIA DE MCROHABITAT DE REPTILES Y ANFIBIOS EN UN GRADIENTE DE VEGETACION EN UN AGROECOSISTEMA DEL
DEPARTAMENTO DEL HUILA.
EDGAR ANDREA BERNAL CASTRO
PONTIFICIA UNIVERSIDAD JAVERIANA BOGOTA-COLOMBIA
USO Y PREFERENCIA DE MCROHABITAT DE REPTILES Y ANFIBIOS EN UN GRADIENTE DE VEGETACION EN UN AGROECOSISTEMA DEL
DEPARTAMENTO DEL HUILA.
EDGAR ANDRES BERNAL CASTRO
________________________________ ______________________________ Dra. Ingrid Schuler Dra. Andrea forero
USO Y PREFERENCIA DE MCROHABITAT DE REPTILES Y ANFIBIOS EN UN GRADIENTE DE VEGETACION EN UN AGROECOSISTEMA DEL
DEPARTAMENTO DEL HUILA.
EDGAR ANDRES BERNAL CASTRO
NOTA DE ADVERTENCIA
Articulo 23 de la Resolución NO 13 de Julio de 1946
AGRADECIMIENTOS
Todo mi proceso de formación académica no pudo haber sido posible sin el apoyo constante y la motivación de mis padres Edgar Antonio Bernal Prieto y Patricia castro Charry, a los cuales amo y dedico este trabajo con todo el orgullo que como hijo puedo ofrecerles, también agradezco a toda mi familia, mis tíos y mis primos que me acompañaron, apoyaron y estuvieron presentes en todo mi proceso educativo y con los cuales seguiré contando de manera incondicional siempre.
A reglón seguido este trabajo no pudo haber sido realizado sin la supervisión y el apoyo constante del doctor José Nicolás Urbina Cardona, ya que gracias a su profesionalismo y la alta calidad que imprime en sus trabajos de investigación permitió que este trabajo de grado tuviera los estándares de calidad presentes en un trabajo profesional cumpliendo con todas las normas de ética y moral pertinentes.
También agradezco a todos mi profesores en especial al doctor Andrés Rymel Acosta Galvis por sus enseñanzas en herpetología, al doctor Julio Mario Hoyos por sus consejos y recomendaciones y a todos mis colegas que compartieron conmigo la experiencia de mi trabajo de grado, esta dedicatoria es para ustedes, Dios los Bendiga y
les retribuya generosamente todas las enseñanzas y consejos que ahora me hacen ser parte del gremio profesional.
“La libertad comienza cuando te das cuenta de que no eres «el pensador». En el momento en que empiezas a observar al pensador, se activa un nivel de conciencia superior. Entonces te das cuenta de que hay un vasto reino, Un reino de inteligencia más allá del pensamiento, y de que el mismo pensamiento: es sólo es una pequeña parte de esa inteligencia…”
RESUMEN
Se hizo un estudio sobre la distribución, el uso y la preferencia de microhábitat de la comunidad de herpetos encontrada en un agroecosistema ubicado en zona de bosque seco tropical en el departamento del Huila, se tomaron variables microclimáticas y de microhábitat de cada individuo encontrado en cuatro hábitats comunes de estas zonas: potrero en uso, potero abandonado, cultivo de flores y bosque de galería, los resultados mostraron que el fragmento de bosque seco tropical mantiene la mayor variedad de herpetos de los cuatro hábitats, sin embargo poblaciones de anfibios como Hypsiboas crepitans se encuentran mayormente en el hábitat de cultivo de flores de la familia Heliconaceae, así como poblaciones de reptiles como Gonatodes albogularis, lo que demuestra la utilidad de este hábitat para soportar algunas poblaciones de reptiles y anfibios, sin embargo, especies de anfibios como Rhinella gr. Margaritifer e Hiloxalus vergeli demostró estar muy ligada a los hábitats conservados del agroecosistema por lo que es de vital importancia la conservación de estas zonas para mantener una diversidad de herpetos que no sobreviven en otros hábitats del agroecosistema.
INTRODUCCION
Actualmente el cambio en el uso en el suelo de bosques y zonas naturales por
presente estudio se caracterizo la dinámica en la estructura y composición de la herpetofauna que se genera en un agroecosistema ubicado en una zona de bosque seco tropical, en el pueblo de Campoalegre, departamento del Huila. Se identificaron los cuatro hábitat comunes en las fincas de la región como son los potreros en uso, los potreros abandonados, los cultivos de flores y los bosques de galería y en cada hábitat se muestreo la herpetofauna y se midieron las condiciones climáticas, estructurales y topográficas que determinan la presencia de las especies mas comunes. A través del protocolo anterior, el presente trabajo busca responder las siguientes preguntas de investigación: (a) ¿cuales son los patrones de abundancia y composición de reptiles y anfibios a lo largo de un gradiente de vegetación en el agroecosistema?, (b) ¿cual es la
relación que existe entre las variables ambientales, topográficas y estructurales con la riqueza y abundancia de especies de reptiles y anfibios del agroecosistema?, (c) ¿Cuáles
son las preferencias de microhábitat de reptiles y anfibios con respecto a los tipos de vegetación en las agro-zonas del agroecosistema?.
JUSTIFICACION Y PLANTEAMIENTO DEL PROBLEMA
En Colombia el ecosistema más degradado y poco estudiado es el bosque seco tropical, el cual se ha reducido a pequeños fragmentos y bosques de galería (IAvH 1998; Merlano, 2006). En el departamento del Huila, este ecosistema ha sido transformado en agroecositemas que están conformados por potreros para ganadería y cultivos con diferentes microclimas y coberturas vegetales.
El cambio antropogénico en el uso del suelo ha tenido un efecto sobre a la comunidad de anfibios y reptiles reduciendo sus poblaciones, provocando extinción de especies, y generando la dispersión de algunas especies hacia las nuevas agro-zonas creadas por el hombre, sin embargo algunas especies pueden haberse beneficiado del agroecosistema presente (Gardner et al, 2007; Cáceres-Andrade & Urbina-Cardona 2009).
a los gradientes de hábitat generados por los cambios en el uso del suelo en la región para así conocer el impacto de las acciones humanas en este grupo.
Es de vital importancia conocer el funcionamiento de los nuevos agroecositemas y los efectos que estos han tenido en la comunidad de reptiles y anfibios para establecer parámetros de conservación de especies, para tal fin se tiene que partir del análisis de la estructura y composición de las especies de una comunidad y su distribución a lo largo de gradientes ambientales y estructurales generados por un mosaico de hábitats naturales y antropogénicos (Urbina-Cardona & Londoño 2003).
Los anfibios y reptiles cumplen un papel importante en la conservación de zonas naturales de los agroecosistemas y en el desarrollo sostenible de las nuevas zonas como cultivos y potreros, dada su naturaleza de depredadores en general, los anfibios se alimentan de insectos que pueden ser plagas que consumen tanto las nuevas plántulas que nacen en las zonas conservadas, como los frutos y hojas de los cultivos, también sirven como indicadores biológicos al ser sensibles a los químicos y toxinas en el agua por su piel permeable y por los dos estadios por lo que pasan en su ciclo de vida la mayoría de anfibios (renacuajo – adulto) (www.mma.es). Al mismo tiempo muchos
reptiles son insectívoros y otros como las serpientes sirven para combatir plagas como los roedores que pueden consumir también las pequeñas plantas y ser fuente de
enfermedades.
cambios en el uso del suelo e identificar las preferencias de microhábitat en los nuevos sistemas productivos.
MARCO TEORICO
El bosque seco tropical.
Los Bosques secos Tropicales son formaciones vegetales que se caracterizan por presentar una cobertura boscosa continua, por su distribución entre los 0-1000 m de altitud; se presenta en zonas con temperaturas superiores a los 24 C (piso térmico cálido) y precipitaciones entre los 700 y 2000 mm anuales, con uno o dos periodos marcados de sequía al año, (Espinal 1985; Murphy & Lugo 1986; IAvH 1997). El
bosque seco tropical también se le denomina bosque higrotropofítico, bosque seco Tropical de Holdridge, bosque tropical caducifolio de diversos autores, y bosque tropical de baja altitud deciduo por sequía de la clasificación propuesta por la UNESCO (IAvH 1997).
La cobertura vegetal es variada, los árboles mas grandes poseen alturas entre 15 y 25 metros y se pueden presentar hasta cuatro estratos vegetativos que incluyen estratos herbáceos, una característica de estos bosques es la escasez o ausencia de plantas
epifitas, además que el sotobosque posee una densidad de hierbas mucho menor comparada con bosques mucho mas húmedos (IAvH 1995, 1997; Gentry 1995), las
densidades de lianas e individuos de árboles y arbustos por unidad de área son similares a las de los bosques húmedos tropicales, la diferencia radica en la cantidad de madera encontrada, la cual es mucho menor en los bosques secos tropicales (Gentry 1995). En
Colombia, la familia de las leguminosas es la familia más representativa del bosque seco tropical, seguida de la familia Bignoniaceae, Sapindaceae y Capparidaceae respectivamente (IAvH 1998). Se reporto también especies vegetales del genero Acidocroton en bosque seco tropical (Fernández 1995). En el bosque seco tropical se
adaptación como son la perdida de follaje, adaptaciones estructurales como la presencia de hojas compuestas y foliolos pequeños, la corteza de los troncos es lisa y poseen aguijones y espinas para evitar la predación de hojas y tallos por parte de los mamíferos (IAvH 1995).
A si mismo, los animales presentan adaptaciones para las épocas de sequía como son las migraciones hacia bosques riparios y zonas húmedas, algunos animales cambian su dieta o acumulan lípidos y grasas para las épocas de bajos recursos alimenticios, también se producen fenómenos de estivación en diferentes especies, y las poblaciones mas propensas la deshidratación como los anfibios se restringen a los pocos cuerpos de agua permanentes (Ceballos 1995).
Anteriormente el Bosque seco Tropical representaba el 50% de las áreas boscosas en Centroamérica y el 22% en Sudamérica (Murphy & Lugo, 1986). Aunque la
información exacta de la cobertura vegetal original del Bosque seco Tropical en Colombia no se encuentra, se estima que cubría un área considerable del territorio Nacional llegando a una extensión de más de 8’146.000 hectáreas (IAVH 1994).
Conocimiento actual del bosque seco tropical en Colombia.
El Bosque seco Tropical esta considerado entre los tres ecosistemas menos conocidos y poco estudiados (Etter, 1993), en Colombia se tienen registrados 73 estudios
publicados o inéditos del bosque seco tropical, de los cuales 9 se han publicado para la región del valle seco del río Magdalena y los Departamentos de Tolima, Cundinamarca y Huila, y de los cuales solo 2 son trabajos puntuales siendo esta zona, una de las menos conocidas y con menos trabajos publicados, (IAVH, 1995; Fernández, 1995). En esta
región se desconoce la cobertura y distribución de remanentes boscosos en el departamento de Cundinamarca, sur del Tolima y norte de Huila. En cuanto a la composición herpetofaunística, existen pocos estudios de composición y no existen trabajos comparativos entre regiones. (Castro & Kattan 1991; Castro 1991), se
en la región Caribe y en donde existen zonas de bosque húmedo, seco y matorrales subxerofíticos, se reportan 13 especies de anfibios y 14 de reptiles (Moreno 1979), para
parches de bosque seco tropical ubicados en la región de Córdoba se registraron 14 especies de serpientes, 20 de lagartos y una de tortuga (Carvajal-Cogollo & Urbina-Cardona 2008), un estudio reciente reveló que en la costa Caribe colombiana se determinaron 46 tipos de herpetos (Álvarez et al, 2008). Torrente-Ruiz y colaboradores
(en prensa) reportan la distribución de anuros en un agroecosistema en el bosque seco tropical de Cambao (Cundinamarca) y reportan tres especies con una mayor abundancia en el hábitat de quebraba, seguidos por los matorrales. Posiblemente en el pasado haya existido una fauna mas diversa y rica en los Bosques secos Tropicales, pero debido el aislamiento y la fragmentación de los remanentes boscosos, se pudo haber producido diferentes procesos biológicos como migraciones que han permitieron colocar especies en peligro o que hayan desaparecido (IAvH 1998).
Situación actual del bosque seco tropical.
El bosque seco tropical es considerado actualmente uno de los ecosistemas mas amenazados del neotrópico, gracias a la fertilidad de los suelos, muchas áreas de bosque seco tropical han sido puntos de desarrollo humano y transformación del ecosistema (Janzen 1983). En Colombia, Algunos estimativos señalan que solo queda cerca del 1.5% de su cobertura original de 80.000 km2 de bosques secos (Etter 1993).
La llanura Caribe y el sur de La Guajira es la región que actualmente posee la mayor cobertura de bosque seco tropical en la actualidad, seguida de la región seca del valle del río Magdalena, en los departamentos de Tolima, Cundinamarca y Huila y finalmente el Valle geográfico del rió Cauca en donde solo existen pequeños remanentes aislados (IAvH 1998).
permanecido gracias a la carencia de tecnología para realizar cultivos comunes en las tierras planas (IAvH 1998)
Actualmente los pocos remanentes de bosque seco tropical existentes presentan una cobertura mínima y se encuentran amenazados por actividades extractivas y por sistemas productivos, por esta razón es prioritario adelantar estudios sobre el efecto de la fragmentación en las poblaciones remanentes de las especies de fauna y flora, estudios de restauración ecológica así como estudios de comportamiento de fauna, migraciones locales, y en general la caracterización de todos los hábitats riparios y dinámica del bosque seco tropical (IAvH 1998).
Recambio del uso del suelo: los agroecositemas.
Los remanentes de bosque seco tropical presentes en Colombia están rodeados en su mayoría, por agroecositemas especialmente en la región del Magdalena y los valles interandinos, donde se concentra gran parte de la producción agrícola y ganadera del país (IAvH 1998).
Un agroecosistema se define como un ecosistema el cual es sometido a frecuentes
modificaciones tanto en sus componentes estructurales, funcionales bióticos y abióticos. El problema que afrontan la mayoría de agroecositemas es la alta velocidad que existe en la razón de cambio de la cobertura vegetal y los microclimas presentes, todo ello provocado por los procesos de cosecha y potrerización (Ghersa y Martínez-Ghersa 1991; Hald 1999).
El establecimiento de agroecositemas es un fenómeno de amplia extensión siendo de todas las acciones humanas, la que más produce cambios por unidad de área en todos los ecosistemas (Solbrig 1999). Según algunas estimaciones casi la mitad de la
superficie terrestre a sido transformada para practicas agrícolas (12%), ganaderas (25%) y de aprovechamiento forestal (15%) (Solbrig 1999).
Efectos de los agroecositemas sobre la herpetofauna.
Se sabe que uno de los principales factores de extinción en anfibios y reptiles es la pérdida de hábitat, causada por el cambio del uso de la tierra por parte del hombre (www.mma. 2010), Otras causas principales de alteración, reducción y extinción de las
comunidades de herpetofauna son la contaminación ambiental y la introducción de especies, algunos de los factores que generan contaminación en ambientes naturales habitados por anfibios y reptiles son los agentes químicos utilizados en la agricultura y ganadería (www.mma. 2010).
El establecimiento de agroecositemas tiene como característica la perdida de la continuidad entre los hábitat naturales, reduciendo a fragmentos o remanentes de hábitat rodeados por una matriz antropogénica que presenta mayor extensión en el paisaje; ese fenómeno se conoce como fragmentación (Wilcox 1980), La fragmentación se ve
reflejada en cambios en la estructura de la vegetación que afectan el microclima e
influyen fuertemente en las comunidades de anfibios y reptiles que son muy dependientes de los microclimas (Urbina-Cardona et al, 2006; Gardner et al, 2007 a, b), el efecto producido sobre la herpetofauna por un agroecosistema puede entenderse no solo por la riqueza y abundancia de especies, sino también por la composición de las mismas dentro de zonas especificas del agroecosistema (Carvajal-Cogollo & Urbina-Cardona 2008), ya que algunas especies pueden estar en varios lugares pero otras pueden verse reducidas a unas pocas zonas por sus características eco-fisiológicas (Cáceres-Andrade & Urbina-Cardona 2009).
cuando eran una sola unidad, este fenómeno permite establecer una relación entre el proceso de fragmentación y la reducción local del numero de especies (Ver Bolger et al, 1997).
El impacto que pueden generar los agroecositemas sobre la flora y fauna nativa puede ser drástico, llevando incluso a muchas especies animales y vegetales a la extinción local (Prange & Gehrt 2004); sin embargo, y a pesar de que se sabe el efecto de los
agroecosistemas en la reducción de la biodiversidad, algunas especies de características generalistas pueden llegar a sobrevivir y alcanzar densidades mayores en los ambientes perturbados que en los ambientes naturales, según el caso, otras especies migran dependiendo de la época del año hacia diferentes agrozonas o hacia los remanentes de bosque cercanos (Prange & Gehrt 2004; Prange et al, 2004).
La dinámica de migraciones es compleja, y depende tanto de los grupos de especies como de las especies mismas, ejemplos son algunas especies del genero Anolis que pueden verse influenciados negativamente por la reducción del tamaño de los fragmentos de bosque (Carvajal-Cogollo & Urbina-Cardona 2008), mientras que la
densidad de serpientes puede verse aumentada en agrozonas de Colombia como los cultivos de coco en las temporadas secas del año (Urbina-Cardona et al, 2008).
Sin embargo hay que tener en consideración que para la fauna asociada tanto al bosque seco tropical como a los agroecosistemas es de vital importancia que existan otros tipos de ecosistemas aledaños que generen un mosaico en el paisaje. Debido a las migraciones locales que se producen en relación a la disponibilidad de recursos (IAVH 1998), en Colombia, la mayoría de los remanentes de bosque seco tropical se encuentran en completo asilamiento rodeados por matrices de cultivo y pastizales lo que reduce la calidad del hábitat de los remanentes y los aísla de la dispersión de especies afines al bosque (IAvH 1998).
OBJETIVO
General
Comparar el uso y preferencia de microhábitat de herpetofauna en un gradiente de vegetación dentro de un agro-ecosistema y en fragmentos de bosque seco tropical en la región del Campoalegre - Huila.
Objetivos específicos
Comparar la riqueza, composición y patrones de abundancia de especies de reptiles y anfibios presentes a lo largo de un gradiente de vegetación en un agroecosistema y en remanentes de bosque seco tropical.
Determinar la relación entre variables ambientales, topográficas y estructurales con la riqueza y abundancia de especies de reptiles y anfibios.
METODOLOGIA
Área de estudio
El estudio se realizo en la finca “Potosí” de 80 hectáreas ubicada al sur este del pueblo de Campoalegre en el departamento del Huila a 20 minutos de la ciudad de Neiva, la altura va desde 500 hasta 610 msnm, la geomorfología de la finca es plana limitando al norte con un piedemonte que llega a los 750 metros, la temperatura media es de 27 C, su precipitación media anual es de 1254 milímetros.
La finca ha sido desde hace al menos 80 años utilizada para la ganadería y la agricultura, se dan cultivos como arroz, tabaco, maíz y gran parte se destina a la cría de ganado, también se producen cultivos de flores por ser una finca que posee gran numero de riachuelos, quebradas y un riachuelo que la bordea, muchos de estos cauces han sido redirigidos hace muchos años por acequias que riegan periódicamente los cultivos presentes en la zona. Para el presente estudio se analizaron cuatro agrozonas representativas de la finca estas son:
Potrero en uso: potrero usado frecuente e intensivamente para ganadería, la estructura de la vegetación es muy simple predominando los pastos, y existe poca vegetación
arbustiva no mayor a 50 cm de altura; se observan piedras grandes, troncos y tierra suelta así como abundante estiércol de vaca, algunos de los potreros poseen zonas de sombrío en sus linderos por los árboles que se han plantado y las cercas vivas, este potrero posee dos acequias que lo atraviesan y por las cuales corre agua en diferentes horas del día durante todo el año dependiendo de la apertura y cierre de compuertas por parte de los parceleros para desviar el curso del agua con fines productivos.
trochas, la incursión del ganado es ocasional ya que solo posee una franja de pasto de aproximadamente dos metros y medio que bordea la quebrada por lo que Su uso para forrajeo es muy reducido.
Cultivo de flores: Cultivo de flores de la familia Heliconaceae, del cual se extraen flores si extraer completamente las plantas; la extracción se hace a lo largo de todo el año sin llegar a extraer mas de el 40% de las flores por ves, las plantas poseen una altura de alrededor de 2,5 metros con capas de hojas grandes cubriendo el suelo del cultivo ya que las hojas se retiran de la flor y forman cúmulos de hojarasca que llegan hasta 1 metro de altura apoyados contra el cerco de piedra, algunas hojas pequeñas caen de arboles de la familia Fabaceae los cuales generan una cobertura de dosel alta en parte del cultivo, el cultivo esta rodeado por una cerca de piedra que genera refugios para reptiles y anfibios, y se mantiene con riego durante todo el año, el suelo permanece húmedo lo que mantiene la humedad y limita con una acequia al norte con agua durante todo el año, en este cultivo no se utilizan ni herbicidas ni pesticidas, solo control biológico por los anfibios presentes.
Bosque de galería: fragmento de bosque seco tropical de alrededor de 70 metros de ancho y 800 metros de longitud que se extiende por la rivera del río y quebradas aledañas. Posee una vegetación arbórea constante con arboles de alrededor de 50 metros
de altura, abundante cobertura de hojarasca y poca vegetación herbácea, limita con una quebrada natural con agua durante todo el año de alrededor de 10 metros de rivera a rivera al este y con un potrero en uso al oeste, según información suministrada por los dueños de la finca nunca se ha talado de manera total por lo que es la parte mejor conservada del antiguo territorio de la zona.
Muestreos
Identificación de especies
especies y se tomaron los respectivos datos morfométricos para los ejemplares, con esta información se construyo una guía de campo de especies potenciales y se corroboro la información con la revisión de especímenes de el Museo Javeriano de Historia Natural (MUJ), de la Pontificia Universidad Javeriana sede Bogotá – Colombia, los organismos capturados que no se pudieron identificar directamente en campo, se preservaron y depositaron en el MUJ para su identificación.
Método de muestreo
La obtención de datos se realizo entre diciembre del 2009 y mayo del 2010, se hicieron cuatro salidas de campo, dos en temporada seca y dos al inicio de las lluvias, se ubicaron los cuatro hábitat de la finca para realizar el muestreo: potreros en uso, potreros abandonados, cultivo de flores y bosque de galería, en cada hábitat se ubicaron aleatoriamente cinco transeptos lineales (60 m de longitud) por cada salida de campo y separados por una distancia mínima de 4 metros entre ellos. Cada transepto se recorrió durante 20 minutos en la mañana y 20 minutos en la noche realizando conteos visuales y captura manual, hasta 1,60 m de altura (Crump & Scott 1994; Jaeger 1994; Tocher et al, 1997) y 2 m a la redonda (Sutherland 1996) para un esfuerzo de muestreo total de 53 horas-hombre a lo largo de 20 transeptos por hábitat (80 transeptos por la totalidad de hábitats).
Gracias a información de expertos en reconteo de anfibios y reptiles (captura – recaptura) se conoce que la tasa de recaptura es muy baja incluso en poblaciones numerosas, por lo que se capturaron los individuos por día, y se mantuvieron en un bioterio para evitar la recaptura de noche. Sin embargo los individuos encontrados en salidas diferentes se tomaron como independientes debido a la poca tasa de recapturas.
graduada; la cobertura herbácea y de hojarasca por medio de parcelas de 1x1 metros; y el porcentaje de cobertura de dosel con un densiómetro (Tocher et al, 1997; Knutson et al, 1998; Urbina-Cardona et al, 2006; Cáceres-Andrade & Urbina-Cardona 2009). Finalmente la distancia entre el individuo y el cuerpo de agua más cercano se midió con un GPS haciendo una georeferenciación de todos los transeptos y utilizando la función “Go To” desde el cuerpo de agua más cercano a cada transepto. La precipitación se midió en cada día de muestreo con ayuda de un pluviómetro presente en una estación meteorológica ubicada en la finca.
ANÁLISIS DE DATOS:
Diversidad total de especies.
Esta se tomó siendo la sumatoria de especies que se registraron en el sitio de muestreo. Para medir la riqueza especifica se utilizaron estimadores de riqueza no paramétricos Jacknife de primero y segundo orden (Magurran 1988; Colwell & Coddington 1994) en el programa estimateS 7.5 (Colwell 2005), se utilizaron estos estimadores ya que
asumen heterogeneidad en las muestras, siendo los mas indicados para estimar riqueza tanto de reptiles como de anfibios en las agrozonas de la finca. (Magurran 2004).
Diversidad espacio – temporal: Alfa.
Se tomó el número de especies tanto de reptiles como de anfibios en cada hábitat muestreado (potrero en uso, abandonado, cultivo de flores o bosque de galería) y se midió por estimadores no paramétricos, se hace una aproximación de la riqueza especifica como una función de la acumulación de especies (Magurran 1988, Colwell & Coddington 1994), utilizando los estimadores Chao 1 y 2 y Boostrap en el programa estimateS 7.5 (Colwell 2005), se aplico el Chao 1 y 2 ya que asumen homogeneidad de
El método de Bootstrap junto con Jacknife de primer y segundo orden se utilizaron para detectar diferencias en la biodiversidad entre la temporada seca y la temporada húmeda, con intervalos de confianza del 95%.
Diversidad beta de las especies
Para medir el recambio en la composición de especies entre las diferentes agrozonas se utilizo el índice de complementariedad, ya que este relaciona el número de especies en un área A con el número de especies de un área B y las especies comunes entre A y B (Colwell & Coddington 1994, Magurran 2004). Así, la riqueza total para ambos
hábitats combinado es: S AB = a + b – c
Se calcula también riqueza única para ambos hábitats: U AB = a + b – 2c
Luego, partiendo de los valores obtenidos se calculo la complementariedad de los sitios A y B así:
C AB = U AB / S AB
Estos valores de complementariedad varían, con un rango de 0 a 1, siendo cero el valor de mayor complementariedad en composición de especies (idénticos) entre los lugares, y uno el menor grado de complementariedad, cuando son totalmente distintos (Colwell & Coddington 1994).
Abundancia
Esta se calculó con la abundancia de anfibios y reptiles que se encontraron por cada hábitat y se aplico en análisis de Kruskal-Wallis, (Siegel, 1980; Zar, 1999), por medio del programa Statistica 6.0 (Stasoft 2001).
Patrones en la abundancia y composición
Se usaron las curvas de rango abundancia (Feinsinger 2001), las cuales se construyeron
ocupados por la especies de la mas a la menos abundante para observar las tendencias en cuanto a las especies encontradas en los diferentes hábitats y al número de individuos por especie para observar posibles patrones en abundancia y composición.
Preferencia del microhábitat.
Se describió el microhábitat de reptiles y anfibios considerando el tipo de sustrato donde se observaron los individuos de cada especie (tronco, rama, hojarasca, roca, hueco, tierra desnuda, hoja, cuerpo de agua y se detectaron los microhábitats mas usados, se tomara como uso la presencia o ausencia de especies en los microhábitats y como preferencia el número de especies y de individuos por especie en cada sustrato.
Correlación entre variables del microhábitat.
Se hizo un análisis de coeficientes de correlación de Spearman para relación que existe entre las diferentes variables de microhábitat para identificar las variables que no están correlacionadas, utilizando el programa Statistica 6 (Statsoft 2001).
Relación entre variables del microhábitat y la presencia de herpetofauna.
Estas se analizaron utilizando un modelo de regresión múltiple por pasos en el programa Estadística 6.0 (Statsoft 2001), para observar las variables que mas están relacionadas
con la presencia de las especies de reptiles y anfibios mas comunes; cuya abundancia sea mayor a 5 individuos en los microhábitats evaluados en las 4 agrozonas (n = 67). Las variables que no presentaron significancia en la prueba de F (p < 0.05) fueron eliminadas del modelo.
RESULTADOS
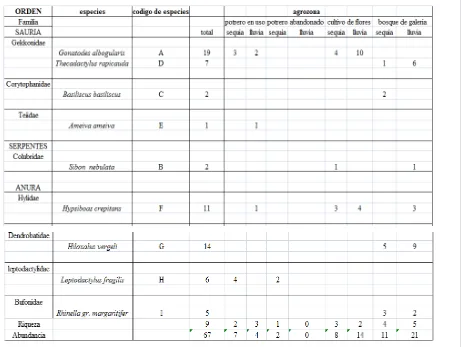
A la totalidad de muestreos que sumaron 53 horas-hombre se registraron 67 individuos pertenecientes a 5 especies de reptiles y 4 especies de anfibios pertenecientes a 9 géneros y 8 familias. La familia más representativa fue Gekkonidae teniendo el 22% de las especies colectadas (Tabla 1).
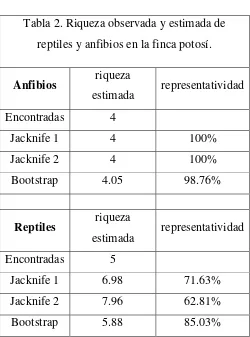
[image:22.595.89.550.402.749.2]Los estimadores de riqueza de Jacknife de primero y segundo orden mostraron un número estimado de especies de anfibios totales igual al encontrado en el área de estudio, representando el 100% de la riqueza esperada, el Bootstrap arrojo un resultado similar representado el 98% de las especies. En cuanto a los reptiles la representatividad fue menor, estando por el orden del 63 y el 68% para Jacknife, y del 65% para el Bootstrap (Tabla 2).
Tabla 2. Riqueza observada y estimada de reptiles y anfibios en la finca potosí.
Anfibios riqueza
estimada representatividad
Encontradas 4
Jacknife 1 4 100%
Jacknife 2 4 100%
Bootstrap 4.05 98.76%
Reptiles riqueza
estimada representatividad Encontradas 5
Jacknife 1 6.98 71.63%
Jacknife 2 7.96 62.81%
Bootstrap 5.88 85.03%
Diversidad espacio-temporal: Alfa
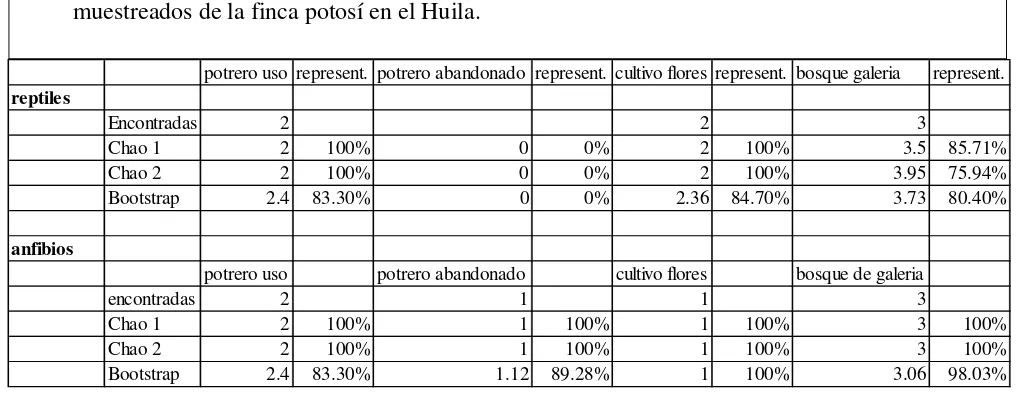
La riqueza de reptiles fue mayor en el bosque de galería con 3 especies seguida del potrero en uso y el cultivo de flores con 2 especies cada uno. Los estimadores de riqueza Chao 1 y Chao 2 y Bootstrap mostraron un numero de especies esperadas similar al numero de especies encontradas, con una representatividad entre el 83% y el 100% de las especies encontradas en el potrero en uso; de entre 84% y el 100% para el cultivo de flores; y entre el 75% y el 86% para el bosque de galería; en el potrero abandonado no se registraron reptiles (Tabla 3).
en campo con valores de entre el 83% y el 100% para el potrero en uso; entre el 89% y el 100% para el potrero abandonado; del 100% para el cultivo de flores; y entre el 98% y el 100% para el bosque de galería (Tabla 3).
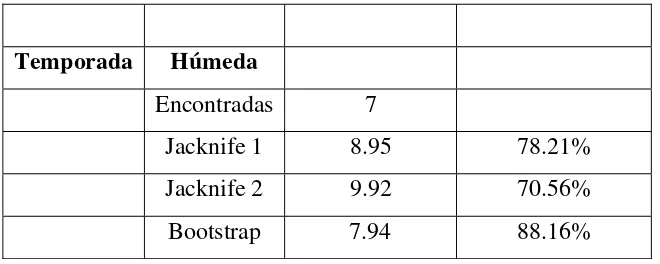
[image:24.595.54.566.356.557.2]La riqueza de herpetos fue mayor en la temporada seca que en la temporada húmeda, y se tuvo una representatividad similar entre temporadas. Para la temporada húmeda y seca los indicadores Jacknife de primer y segundo orden y el Bootstrap soportan una representatividad entre el 67% y el 83% para temporada seca; y entre el 70% y el 88% para la temporada húmeda (Tabla 4).
Tabla 3. Riqueza observada y estimada de herpetos registrados en los cuatro hábitats muestreados de la finca potosí en el Huila.
potrero uso represent. potrero abandonado represent. cultivo flores represent. bosque galeria represent. reptiles
Encontradas 2 2 3
Chao 1 2 100% 0 0% 2 100% 3.5 85.71%
Chao 2 2 100% 0 0% 2 100% 3.95 75.94%
Bootstrap 2.4 83.30% 0 0% 2.36 84.70% 3.73 80.40%
anfibios
potrero uso potrero abandonado cultivo flores bosque de galeria
encontradas 2 1 1 3
Chao 1 2 100% 1 100% 1 100% 3 100%
Chao 2 2 100% 1 100% 1 100% 3 100%
Bootstrap 2.4 83.30% 1.12 89.28% 1 100% 3.06 98.03%
Tabla 4. Riqueza observada y estimada de herpetos regitrados en temporada seca y húmeda en la finca potosí en el Huila.
Temporada Seca Riqueza estimada
Representatividad
Encontradas 8
Jacknife 1 11.9 67.22%
Jacknife 2 15.7 53.30%
Temporada Húmeda
Encontradas 7
Jacknife 1 8.95 78.21%
Jacknife 2 9.92 70.56%
Bootstrap 7.94 88.16%
Diversidad beta de las especies
El potrero abandonado por solo tener una especie registrada presenta el mayor recambio de especies con el cultivo de flores y el bosque de galería (1), sigue en orden jerárquico el recambio de especies entre el bosque de galería y el potrero en uso (0. 77) y el menor recambio de especies se dio entre el potrero en uso y el potrero abandonado
[image:25.595.134.464.83.215.2](0.5), se registró una especie única para el potrero en uso y 4 especies únicas para el bosque de galería (Tabla 5).
Tabla 5. Complementariedad de la herpetofauna presente en cuatro habitats de la finca potosí, municipio de Campoalegre – Huila.
potrero en uso potrero abandonado
cultivo de
flores bosque de galería
potrero en uso 1 1 2 1
potrero abandonado 0.5 0 0 0
cultivo de flores 0.6 1 0 2
bosque de galería 0.77 1 0.71 4
Los valores de la diagonal subrayados y en negrilla corresponden al número de especies únicas para cada Agrozona, los valores por encima de la diagonal corresponden al número de especies compartidas entre sitios. Los valores inferiores a la diagonal representan la complementariedad (rango de 0 a 1, siendo cero cuando son idénticas y 1 cuando son completamente distintas).
Se registraron 67 individuos pertenecientes a 5 especies de reptiles y 4 especies de anfibios (Tabla 1). Se determino que si existen diferencias en la composición de anfibios entre los hábitats (H = 3. 34; p = 0.34). En cuanto a reptiles también existen diferencias en la composición entre los hábitats (H = 3.76; p = 0.29).
En cuanto a reptiles se pudo determinar que no existen diferencias en la composición entre temporadas climáticas, (H = 0.04; p = 0.82); tampoco existen diferencias en la composición de anfibios entre temporada seca y temporada húmeda (H = 0; p = 1).
Patrones en la abundancia.
Las curvas de rango abundancia presentaron pendientes diferentes debido al recambio en la dominancia de especies entre hábitats (Figura 1). El bosque de galería presento la mayor equidad debido a la ausencia de especies dominantes en el ensamblaje, mientras que los cultivos de flores fueron dominados en su mayoría por dos especies. Las formas esperadas o patrones de distribución de la abundancia y el orden jerárquico de abundancias de las especies fueron diferentes entre los cuatro hábitats, salvo la especie Gonatodes albogularis que fue compartida entre el potrero en uso y el cultivo de flores como la especie mas abundante, el anfibio mas abundante en el potrero en uso fue Leptodactylus fragilis y en el cultivo de flores fue Hypsiboas crepitans el anfibio dominante en el Bosque de galería fue Hiloxalus vergeli y el reptil fue Thecadactylus rapicauda, el bosque de galería fue el que presento mayor numero de especies raras (4
Figura 1. Curvas rango-abundancia para cada una de las agrozonas, potrero en uso (triangulo), potrero abandonado (cuadrado), cultivo de flores (circulo), y bosque de galería (rombo).
Uso del microhábitat.
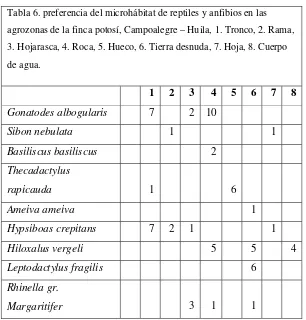
[image:27.595.146.452.327.647.2]Los microhábitats terrestres fueron los que presentaron mayor numero de especies (roca 3 sp.; tierra desnuda 3 sp.; hojarasca 3 sp.), seguidos de los microhábitats arborícolas como tronco, rama y hojas (3 sp. 2 sp. y 2sp. respectivamente). Los cuerpos de agua y los huecos fueron los que presentaron la menor riqueza de especies (una especie para cada uno; Tabla 6)
Tabla 6. preferencia del microhábitat de reptiles y anfibios en las agrozonas de la finca potosí, Campoalegre – Huila, 1. Tronco, 2. Rama, 3. Hojarasca, 4. Roca, 5. Hueco, 6. Tierra desnuda, 7. Hoja, 8. Cuerpo de agua.
1 2 3 4 5 6 7 8
Gonatodes albogularis 7 2 10
Sibon nebulata 1 1
Basiliscus basiliscus 2
Thecadactylus
rapicauda 1 6
Ameiva ameiva 1
Hypsiboas crepitans 7 2 1 1
Hiloxalus vergeli 5 5 4
Leptodactylus fragilis 6
Rhinella gr.
Margaritifer 3 1 1
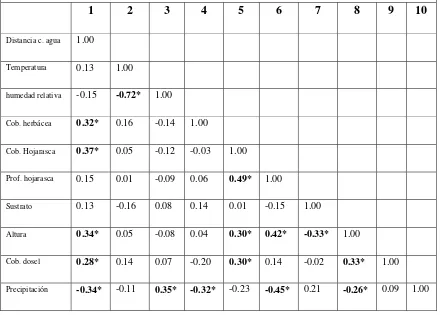
Correlación entre variables del microhábitat.
valores de correlación bajos, por lo que no presentan una relación lineal entre ellas. Las variables que más se relacionan linealmente entre si son la humedad relativa con la temperatura (-72%), cobertura de hojarasca con profundidad de hojarasca (49%) y la profundidad de hojarasca con la distancia del individuo al suelo (42%), el resto de variables tienen una correlación muy baja, esto se puede explicar debido a la heterogeneidad de zonas que se presentan en un agroecosistema, la franja de bosque de galería, los potreros abandonados y en uso y el cultivo de flores son muy homogéneos dentro de si mismos y no presentan grandes cambios en cuanto a cobertura vegetal y distancia a cuerpos de agua (Tabla 8).
Tabla 8. Coeficientes de correlación de Spearman de 10 variables ambientales medidas en la finca potosí, Campoalegre – Colombia: 1. Distancia a cuerpo de agua; 2. Temperatura; 3. Humedad relativa; 4. Cobertura herbácea; 5. Cobertura de hojarasca; 6. Profundidad de hojarasca; 7 sustrato; 8. Distancia del individuo al suelo; 9. Cobertura de dosel; 9. Precipitación.
1 2 3 4 5 6 7 8 9 10
Distancia c. agua 1.00
Temperatura 0.13 1.00
humedad relativa -0.15 -0.72* 1.00
Cob. herbácea 0.32* 0.16 -0.14 1.00
Cob. Hojarasca 0.37* 0.05 -0.12 -0.03 1.00
Prof. hojarasca 0.15 0.01 -0.09 0.06 0.49* 1.00
Sustrato 0.13 -0.16 0.08 0.14 0.01 -0.15 1.00
Altura 0.34* 0.05 -0.08 0.04 0.30* 0.42* -0.33* 1.00
Cob. dosel 0.28* 0.14 0.07 -0.20 0.30* 0.14 -0.02 0.33* 1.00
valores de p * < 0.05
Relación de la herpetofauna con las variables del microhábitat
La humedad, la cobertura de hojarasca y la distancia del individuo al suelo determinaron positivamente la presencia de Gonatodes albogularis en los microhábitats que fueron evaluados, el sustrato afecto de manera negativa la presencia de la especie (Tabla 7).
Para el caso de Thecadactylus rapicauda la distancia a los cuerpos de agua, la humedad, la cobertura de dosel y la precipitación fueron factores que afectaron de manera positiva la presencia de individuos, solo la temperatura afecto de manera negativa la presencia de la especie (Tabla 7).
La presencia de la especie Hypsiboas crepitans se ve influenciada de manera positiva
por las variables de sustrato y la distancia del individuo al suelo, las variables que afectaron de manera negativa fueron la distancia a cuerpos de agua y la precipitación (Tabla 7).
Varios factores tuvieron una relación inversa con la presencia de la especie Hiloxalus vergeli, como la distancia a cuerpos de agua, la cobertura herbácea, la cobertura de hojarasca y la distancia del individuo al suelo, la temperatura y la precipitación afectaron de manera directa la presencia de la especie (Tabla 7).
La presencia de la especie Leptodactylus fragilis se vio determinada de manera positiva por la distancia a cuerpos de agua, en cambio la temperatura, la humedad, la profundidad de la hojarasca y la cobertura de dosel, afectaron de manera negativa la presencia de la especie (Tabla 7).
Especie Modelo de regresión R múltiple F gl P
Reptiles
Gonatodes
albogularis Y=5.181+0.335*C+0.250*E-0.37*G+0.205*H 76% 8.30 9.56 <0.0005
Thecadactylus
rapicauda
Y=0.187+0.443*A-0.53*B+ 0.30*C+0.255*I
+0.474*J 73% 8.25 8.57 <0.0005
Anfibios
Hypsiboas
crepitans Y=-4,033-0,26*A+0.279*G+0.276*H-0,35*J 49% 3.02 6.59 0.01200
Hiloxalus
vergeli
Y=2.743-0.28*A+0.197*B-0.25*D-0.31*E-0.31*H+0.228*J 80% 13.37 8.57 <0.0005
Leptodactylus
fragilis Y=2.667+0.213*A-0.31*B-0.29*C-0.28*F-0.52*I 78% 9.36 9.56 <0.0005
DISCUSION.
Tanto los anfibios como los reptiles que habitan zonas transformadas por actividades humanas, responden de manera diversa y compleja a los cambios de microhábitat generados a lo largo del gradiente pastizal, borde e interior de bosques tropicales (Urbina-Cardona et al, 2006), Según los estimadores se encontró una muestra representativa (alrededor del 98% entre los dos) tanto de reptiles ( 5 especies) como de anfibios ( 4 especies), sin embargo la cantidad de especies es reducida en comparación a registros observados por otros autores en lugares como bosques de galería y bosques secos fragmentados donde la representatividad de las especies esta en alrededor del 70% (35 especies) con un esfuerzo de muestreo considerable (Carvajal-Cogollo & Urbina-Cardona 2008), esto sugiere una condición en la reducción de la riqueza de especies (Prange & Gehrt 2004) de herpetos en el agroecosistema del Huila ubicado en bosque
El tipo de muestreo empleado permite generar datos comparativos entre zonas pero al mismo tiempo genera un sesgo en cuanto a las especies encontradas en vista de que restringe el muestreo a los transectos, en otras zonas de la finca que no se muestrearon como la casa de la finca y las trochas se encontraron dos especies de serpientes (Micrurus mipartitus, Leptodeira anulata), una de lagarto, (Iguana iguana) y una de anfibio (Rhinella marina) que no se encontraron en ninguno de las agrozonas muestreadas, por lo que incluir estas zonas en futuros análisis y aumentar el esfuerzo de muestreo es recomendable para incrementar el numero de especies en el inventario y generar datos mas robustos que generen resultados mas precisos en cuanto a la riqueza de especies.
Haciendo comparaciones, el potrero en uso posee mayor riqueza de especies en relación al cultivo de flores, sin embargo las abundancia de individuos por especie es mayor en el cultivo, las especies como Hypsiboas crepitans y Ameiva ameiva están representadas por un individuo en el potrero en uso, mientas que las especies encontradas en el cultivo de flores están representadas por mas de dos individuos salvo Sibon nebulata la cual es una serpiente y se conoce que el grupo de serpientes por ser grandes depredadoras es un grupo reducido en cuanto a abundancia por especies (Rugiero & Luiselli 1996) y son difíciles de estudiar en campo (Seigel & Collins 1993). La alta abundancia de individuos de las especies en el cultivo de flores refleja una alta calidad
de hábitat para las especies que allí habitan y funciona como un ecotono por cuanto allí se comparten especies de potrero y de bosque.
La baja abundancia por especie encontrada en el potrero en uso puede deberse a que esta agrozona posee cambios a nivel de microclima y microhábitat altos por lo que las especies que tienen la capacidad (ecofisiológica y de dispersión) de atravesar esta matriz, pueden usarla como una zona de transición en la cual se encuentran solo por breves periodos de tiempo y luego migran hacia otras zonas donde las condiciones sean mas estables (Cáceres-Andrade & Urbina-Cardona 2009). La alta presencia de
controlar la territorialidad a los machos (Sprackland 1977), además se encontraron
varios juveniles en la cobertura de hojarasca acumulada junto al cerco de piedra en la cual los animales pueden encontrar alimento (e.g insectos) y refugio (Sprackland 1977).
En el presente trabajo se demuestra que la presencia de G. albogularis se ve influenciada de manera positiva por la humedad relativa y la cobertura de hojarasca, siendo el hábitat de cultivos el que le provee estabilidad en estas variables. Por otro lado, en el hábitat de cultivos, la cobertura vegetal presenta hojas anchas troncos delgados y una altura no mayor a los tres metros lo que genera condiciones de microhábitat aptas para Hypsiboas crepitans (e.g humedad relativa), además en el suelo del cultivo abundan variedad de insectos los cuales fueron vistos siendo consumidos por esta especie, lo que genera alimento constante para población. De manera general el hábitat de cultivos de heliconias provee heterogeneidad de sustratos idóneos para el establecimiento de estas dos especies, que como se demostró través de las regresiones, requieren del sustrato y de perchas sobre el suelo para poder estar presentes en los microhábitats.
Solo se encontró una especie de anfibio Leptodactylus fragilis y ninguna de reptiles en el potrero abandonado, la especie que se encontró también estaba presente en el potrero en uso y solo se encontró en la temporada seca, posiblemente se hayan encontrado mas
individuos de esta especie conforme hubieran pasado los días de temporada húmeda, esta especie se encontró en los transeptos mas cercanos a los cuerpos de agua, y su dependencia a la cercanía a estos cuerpos de agua se ve reflejada en los modelos de regresión múltiple (tabla 7). L. fragilis ha sido catalogada como una especie de zonas abiertas (Savage & Villa1986), en el presente estudio se complementa esta información
determinando que la especie evita zonas boscosas debido a que requiere temperaturas y humedades bajas, así como baja cobertura de dosel y poca profundidad en la capa de hojarasca.
hylidos); la segunda es la ausencia de plantas con estratos arbóreos, cobertura de hojarasca en el suelo y ausencia de sustratos que generen refugios, esto evita que especies de naturaleza trepadora se encuentren en la agrozona y gran parte de las especies encontradas en el agroecosistema se ubican en sustratos que permitan una altura de los individuos al suelo (e.g. cerco de piedra, sustratos arbóreos), aunque se observan algunos insectos en esta agrozona, gran parte son insectos voladores o que presentan un peligro adicional por su defensa a partir de veneno como avispas y escorpiones. Por ello, es necesario incrementar el esfuerzo de muestreo en estas áreas e incorporar los análisis de ítems alimenticios en los análisis.
La temporada seca presentó mayor número de especies que la temporada húmeda, la especie Basiliscus basiliscus se encontró solo en temporada seca, en la temporada húmeda los días por lo general son mas fríos por lo que esta especie pueden reducir sus actividades ecológicas (Van Devender 1975), otra causa probable es que la especie solo
se encontró en el cuerpo de agua principal, el cual es natural y se puede aumentar su cauce de manera abrupta en temporada de lluvias (húmeda), por lo que la especie puede migrar hacia otro hábitat cercano, sumado a la poca abundancia de la especie en la zona puede provocar la falta de registros de individuos en temporada húmeda.
El recambio mas alto de especies se da en dos agrozonas que poseen características muy diferentes; los potreros por su característica de poca cobertura de dosel albergan
especies terrestres o arborícolas de porte pequeño que habitan las cercas y los arboles presentes en los linderos del potrero, las especies que lo frecuentan poseen una afinidad hacia ambientes mas cálidos y no se ven afectados en gran medida por la baja humedad, además toleran condiciones fluctuantes de temperatura y humedad como Leptodactylus fragilis (Savage & Villa1986), Ameiva ameiva (Echternacht 1971) y Gonatodes albogularis (Sprackland 1977); el bosque por el contrario posee una cobertura de dosel
agua mas grande, y el que suministra agua a las acequias que bordean o atraviesan los potreros y los cultivos, sin embargo la presencia de esta especie en cuerpo de agua cercanos a otras agrozonas es nulo, posiblemente se deba a que esta especie utiliza sustratos de piedra, arena y es afín a coberturas vegetales relativamente grandes y estas condiciones no se encuentran en las acequias.
Hiloxalus vergeli al ser una especie de anfibio diurno, mostro estar afectada positivamente por la temperatura, esta especie no se encuentra en lugares ajenos a cuerpos de agua en ninguna parte del agroecosistema lo que se confirma por el efecto negativo a distancia a cuerpos de agua, cobertura herbácea y de hojarasca. La especie al igual de B. basiliscus se encontró exclusivamente en el cuerpo de agua natural, este posee sustratos de piedra, al igual que una cobertura vegetal mas compleja que las acequias, las cuales no poseen rocas ni la vegetación necesaria para la presencia de la especie, por lo que la conservación de hábitats como los bosques de galería son necesarios para que esta especie se mantenga en el agroecosistema, incluso los bosques de galería de pequeño tamaño como este que funcionan a modo de efecto de borde poseen la capacidad de mantener especies que solo habitan estas agrozonas incluyendo la especie Rhinella gr. Margaritifer la cual posee poblaciones en declive y posee cierto grado de amenaza según la IUCN por la perdida de hábitat (La Marca 2004).
Para el caso de Thecadactylus rapicauda la distancia a los cuerpos de agua, la humedad,
la cobertura de dosel y la precipitación fueron factores que afectaron de manera positiva la presencia de individuos, solo la temperatura afecto de manera negativa la presencia de la especie (Tabla 7).
rapicauda en el bosque de galería, solo uno se encontró en el sustrato de tronco, los demás individuos se encontraron en las ruinas de un antiguo acueducto que generaba condiciones de humedad y refugios apropiados para esta especie, por lo que las acciones de tipo antrópico no solo generan condiciones desfavorables para las especies, si no que también generan en algunos casos condiciones de microhábitat que favorecen a algunas especies, incluso Hiloxalus vergeli también se encontró en estas ruinas, siendo una especies que se influenciada un conjunto amplio de condiciones microclimáticas y de microhábitat.
La herpetofauna en un agroecosistema mantiene una dinámica compleja que incluye la dispersión espacio-temporal entre hábitat, preferencias de hábitat y reducción de poblaciones a las zonas conservadas, así como adaptaciones a nuevas agrozonas, las especies mantienen formas constantes o patrones de diversidad y abundancia marcados y diferentes entre las agrozonas de la finca y el uso del microhábitat define la diferencia en la riqueza de las diferentes agrozonas que componen el agroecosistema, es importante en fincas ubicadas en zona de bosque seco tropical, la conservación de al menos una parte del territorio original ya que esta aunque sea una zona relativamente pequeña puede ayudar como zona amortiguadora del impacto antrópico, manteniendo poblaciones de herpetos que solo sobreviven en estas zonas.
Para la conservación de las especies más sensibles, es decir aquellas afines al bosque remanente se recomienda mantener y ampliar las zonas conservadas de la finca para que tengan la capacidad de soportar una mayor variedad de especies y se produzca un aumento en las poblaciones, al mismo tiempo puede provocar efectos en el microclima de zonas aledañas como el aumento en la cobertura de hojarasca y la humedad generada en el bosque lo que beneficiaria a especies tanto de reptiles como de anfibios en los hábitats aledaños.
CONCLUSIONES
especies de los agroecosistemas se relegan hacia las zonas mayor conservadas de los agroecosistemas, las cuales son pequeñas y fragmentadas lo que genera aun mas estrés sobre las poblaciones que solo sobreviven en los fragmentos de bosque.
Algunos hábitats de las agrozonas poseen condiciones de microhábitat adecuadas para mantener poblaciones de herpetos incluso más grandes que las que se encuentran en los fragmentos de bosque y puede ser útil para conservar poblaciones estables de estas especies. Incluso dentro de los hábitats conservados, algunas especies han migrado hacia sustratos creados por el hombre ya que proveen condiciones de microhábitat más idóneas para la supervivencia de los individuos.
Es importante la conservación de parte del territorio original dentro de un ecosistema ya que este actúa como una zona de resguardo de muchas especies de herpetos que no pueden sobrevivir en otros hábitats además de realizar mas estudios relacionados con el uso y preferencia de microhábitat en otros hábitats presentes en los agroecosistemas como cultivos variados, trochas, lagos artificiales entre otros.
RECOMENDACIONES
Se recomienda ampliar el número de hábitats a muestrear en trabajos posteriores además de ampliar el número de variables e incluir variables de alimentación, presencia de
depredadores entre otras.
También se recomienda realizar un esfuerzo de muestreo mas alto debido a la poca cantidad de individuos encontrados en el agroecosistema y por los cambios actuales a nivel climático que modifican la estacionalidad de las temporadas seca y húmeda provocando aumentos o disminución en el tiempo que duran estas temporadas.
BIBLIOGRAFIA
ACEVEDO, M. E; CALVO, L; VALLE, L. 2002. Diversidad y abundancia de la Herpetofauna en plantaciones de café bajo sombra en Quetzaltenango, Guatemala. Centro para la Conservación de la Biodiversidad de Guatemala (CCBG). Disponible en: http://portal.anacafe.org/portal/usercontent/Anacafe/Mercadeo/documetosportal/Reptile sCafe.pdf. Revisado el 13 de noviembre del año 2009.
ÁLVAREZ, ESTEBAN; RODRÍGUEZ, NELLY; GÓMEZ, ANA MARÍA; COGOLLO, ALVARO; MARTÍN, CARLOS; CUARTAS, CARLOS; BLANCO, ARGELINA; ORTEGA, OSCAR & BENÍTEZ, DORIS. 2008. Identificación de prioridades de conservación en remanentes de bosque seco de la costa Caribe de Colombia. Interconexión Eléctrica SA, ISA Colombia, Calle 12 Sur No. 18-168. [email protected]
BOLGER, DT; AC, ALBERTS; RM, SAUVAJOT; P, POTENZA; C, MCCALVIN; D, TRAN; S, MAZZONI & ME, SOULÉ. 1997. Response of rodents to habitat fragmentation in coastal Southern California. Ecological Applications 7: 552-563.
CÁCERES, S. & URBINA-CARDONA, J.N. 2009. Ensamblajes de anuros de sistemas productivos y bosques en el piedemonte llanero, Departamento del Meta,
Colombia. Caldasia 31(1): 175-194.
CEBALLOS, G. 1995. Vertebrate diversity, ecology, and conservation in Neotropical dry forest. En Tropical deciduous Forest Ecosystem. S. BULLOCK, E. MEDINA & H. A. MOONEY (Eds). Cambridge Univ. Press, Cambridge. pp. 195-222
CARVAJAL-COGOLLO J. E. & URBINA-CARDONA J. N. 2008. Patrones de Diversidad y Composición de Reptiles en Fragmentos de Bosque Seco Tropical en Córdoba, Colombia. Tropical Conservation Science Vol.1 (4):397-416.
DENVER, R. J; MIRHADI, N; PHILLIPS, D.M. 1998. Adaptive plasticity in amphibian metamorphosis: response of Scaphiopus hammondii tadpoles to habitat desiccation. Ecology 79(6): 1859–1872.
ESPINAL, L. S. 1985. Geografía ecológica del departamento de Antioquia. Revista de la Facultad Nacional de Agronomía, 38 (1): 24-39
ESPINAL, L.S. & E. MONTENEGRO. 1977. Formaciones vegetales de Colombia. Instituto Geográfico Agustín Codazzi, Bogotá, pp. 201
ETTER, A. 1993. Diversidad ecosistémica en Colombia hoy. En Nuestra diversidad biótica. CEREC y Fundación Alejandro Angel Escobar. P. 43-61
ECHTERNACHT, A. C. 1971. Middle American lizards of the genus Ameiva. Misc. Publ. Mus. Nat. Hist. Univ. Kansas 55: 1-86
FERNANDEZ, J.L. 1995. Hallazgo del género Acidocroton Griseb. (Euphorbiaceae) en Sur America, en un bosque seco de Colombia. Caldasia 17 (82-85): 389-394.
GENTRY, A. H. 1995. Diversity and floristic composition of Neotropical dry forest. En Tropical deciduous Forest Ecosystem. S. BULLOCK, E. MEDINA & H. A. MOONEY (Eds). Cambridge Univ. Press, Cambridge. pp. 116-194
GARDNER, T. A; BARLOW, J. & PERES, C. A. 2007A. Paradox, presumption and pitfalls in conservation biology: The importance of habitat change for amphibians and reptiles. Biological Conservation 138(1-2): 166-179
GARDNER, T. A; RIBEIRO-JUNIOR, M. A; BARLOW, J; ÁVILA-PIRES, T. C. S; HOOGMOED, M. S. & C. A. PERES. 2007B. The value of primary, secondary, and plantation forests for a Neotropical herpetofauna. Conservation biology, volume 21, No.3, 775-787.
HALD, A.B 1999. The impact of changing the season in which cereals are sown on the diversity of the weed flora in rotational fields in Denmark. J. of Applied Ecology 36: 24-32.
INSTITUTO ALEXANDER VON HUMBOLDT, IAVH. 1994. exploración ecológica a los Fragmentos de bosque seco en el Valle del Río Magdalena (Norte del Departamento del Tolima). Grupo de Exploraciones Ecológicas Rápidas, IAVH, Villa de Leyva. Pág. 56
INSTITUTO ALEXANDER VON HUMBOLDT, IAVH. 1997. Caracterización ecológica de cuatro remanentes de Bosque seco Tropical de la región Caribe
colombiana. Grupo de Exploraciones Ecológicas Rápidas, IAVH, Villa de Leyva. Pág. 76
INSTITUTO ALEXANDER VON HUMBOLDT, IAVH. 1998. El Bosque seco Tropical en Colombia. Instituto Alexander Von Humboldt Programa de Inventario de la Biodiversidad Grupo de Exploraciones y Monitoreo Ambiental GEMA. Colombia. Pág. 12 – 24pp
JANZEN , D. H. 1983. Seasonal changes in abundance of larg nocturnal Cag-beetles (Scarabaeidae) in Costa Rica deciduous forest and adyacent horse pasture. Oikos, 41 : 274-283
KNUTSON, M. J; SAUER, D; OLSEN, D; MOSSMAN, M; HEMESATH, L & LANNOO, M. 1998. Effects of Landspace Composition and Wetland Fragmentation on Frog and Toad Abundance and Species Richness in Iowa and Wisconsin, U. S. A. Conservation Biology 13(6): 1437-1446
MERLANO, J. 2006. El Bosque Seco Tropical de Colombia. Ed. Banco de Occidente.
MURPHY, P.G. & LUGO, A.E. 1986. Ecology of tropical dry forest. Annals Review of Ecology and Systematics (17): 67-68.
MORENO, L. M. 1979. Especies de vertebrados que han sido identificadas en el parque nacional natural Tayrona en los diferentes muestreos realizados. INDERENA, Documento Interno.
PRANGE, S. & GEHRT, S. D. 2004. Changes in Mesopredator-Community Structure in Responseto Urbanization. Canadian Journal of Zoology, 82(11):1804-1817.
PRANGE, S; GEHRT S. D. & WIGGERS, E. P. 2004. Influences of Anthropogenic Resources on Racoon (Procyon lotor) Movements and Spatial Distribution. Journal of Mammalogy, 85(3):483-490.
RUGIERO L. & LUISELLI, L. 1996. Ecological notes on an isolated population of Elaphe quatuorlineata. Herpetological Journal 6: 93-95.
SPRACKLAND, R. G. 1977. All about Lizards. T.F.H. Publications. 128 pp.
SOLBRIG, O.T 1999. Observaciones sobre biodiversidad y desarrollo agrícola. En: Mateucci, SD, OT Solbrig, J Morello y G Halffter (editores). Biodiversidad y uso de la tierra. Conceptos y ejemplos de Latinoamérica. Eudeba, Buenos Aires. 29-39.
TOCHER, M; GLASCON, V & ZIMMERMAN, V. 1997. Fragmentation Effects on a Central Amazonian Frog Community: A ten-Year Study, pp. 815-816. In: Laurance, W. F. & Bierregaard, R. O. (Ed.). Tropical Forest Remnants. University of Chicago Press, Chicago, III, USA pp. 616.
URBINA-C J.N. & LONDOÑO-M, M.C. 2003 Distribución de la comunidad de herpetofauna asociada a cuatro áreas con diferente grado de perturbación en la Isla Gorgona, Pacífico colombiano. Rev. Acad. Colomb. Cienc. 27(102):105-113.
URBINA-CARDONA, J.N; LONDOÑO-MURCIA, M.C. & GARCÍA-AVILA, D.G. 2008. Dinámica espacio-temporal en la diversidad de serpientes en cuatro habitats con diferente grado de perturbación antropogénica en el Parque Nacional Natural Isla Gorgona, pacífico colombiano. Caldasia 30(2):407-421.
VAN DEVENDER, R.W. 1975. The comparative demography of two local populations of the tropical lizard, Basisliscus basiliscus. Ph.D. diss., University of Michigan.
WILCOX, B.A. 1980. Insular ecology and conservation. Págs 95-117 En: Conservation Biology: an Evolutionary-Ecological Perspective (eds. ME Soulé y BA Wilcox). Sinauer Associates, Sunderland, MA
LIBROS
W., M. A., Donnelley, R. A., McDiarmid, L. C., Hayec & M. C., Foster. Smithsonian Institution Press, Washington DC.
FEINSINGER, P. 2001. Designing Field Studies for Biodiversity Conservation. Island Press, Washington, DC.
MAGURRAN, A. 1988. Diversidad Ecológica y su Medición. Primera edición. Ediciones Vedra. Barcelona.
MORENO, C. 2001. Métodos para medir la Biodiversidad. Manuales y Tesis. Sociedad Entomológica Aragonesa. 1: 1-83.
SIEGEL, S. 1980. Estadística no paramétrica. Sexta edición. Editorial Trillas. México. 346 p.
SEIGEL, R.A. & COLLINS, J.T. 1993. Snakes: Ecology and Behavior. McGrawHill, Inc., Nueva York.
SUTHERLAND, W.S. 1996. Ecological Census Techniques: A Handbook. Cambridge University Press.
PAGINAS DE INTERNET Y PROGRAMAS UTILIZADOS
COLWELL, R.K. 2005. EstimateS: Statistical estimation of species richness and shared species from samples. Version 7.5. User’s Guide and application published at: http://purl.oclc.org/estimates
MMA. 2010. Tomado de la pagina web: http://www.mma.es/secciones/biodiversid ad/especies_amenazadas/vertebrados/anfibios_reptiles/alytes/pdf/CAPITULOV.pdf Revisado el 2 de junio del 2010.