Susceptibilidad a los antimicrobianos y factores de virulencia
en cepas de Vibrio cholerae no-O1 aisladas de pacientes con
enfermedad diarreica aguda
Luis Enrique Cabrera-Rodríguez1, Laura Bravo-Fariñas2, María Margarita Ramírez-Álvarez2, Alina
Llop-Hernández2, Anabel Fernández-Abreu2, Luis Morier3, Graciela Borrego-Hernández4
1Laboratorio de Microbiología Clínica, Centro Municipal de Higiene y Epidemiología de Güines, La Habana, Cuba. 2 Labora-torio Nacional de Referencia de Enfermedades Diarreicas Agudas, Instituto de Medicina Tropical “Pedro Kourí”, Ciudad de la Habana, Cuba. 3Laboratorio de Cultivos de Células, Instituto de Medicina Tropical “Pedro Kourí”, Ciudad de la Habana, Cuba. 4 Laboratorio de Microbiología Clínica, Centro Provincial de Higiene y Epidemiología de Holguín, Holguín, Cuba
RESUMEN
Solicitud de sobretiros: Dra. Laura Bravo Fariñas, Instituto de Medicina Tropical “Pedro Kourí”, Autopista Novia del Mediodía Km. 6, La Lisa, Ciudad de La Habana, Cuba. E-mail: laura@ipk.sld.cu
Recibido: el 24 de abril de 2008. Aceptado para publicación: el 3 de octubre de 2008
Introducción. Las infecciones intestinales cau-sadas por Vibrio cholerae no-O1 tienen un incre-mento en el ámbito mundial.
Objetivo. Conocer la susceptibilidad bacteriana a drogas de elección y alternativas y determinar la presencia de algunos factores de virulencia como: las enzimas extracelulares (DNasa, gelatinasa, lecitinasa, elastasa y hemolisina), adherencia a células HEp-2 y presencia de fimbrias.
Materiales y Métodos. Se realizó un estudio en el Laboratorio Nacional de Referencia de Enferme-dades Diarreicas Agudas del Instituto de Medicina Tropical “Pedro Kourí”, en 65 cepas de Vibrio cholerae no-O1, aisladas de heces de pacientes con Enfermedad Diarreica Aguda, procedentes de 7 Laboratorios de Microbiología clínica de los Cen-tros Provinciales de Higiene y Epidemiología del país (Ciudad de la Habana, Camagüey, Las Tunas, Holguín, Granma, Santiago de Cuba y el Centro Municipal de la Isla de la Juventud), en el período comprendido de enero a diciembre de 2006. Resultados. El 32.3 % y el 30.7 % de las cepas presentaron resistencia a la sulfonamida y a la am-picilina, respectivamente. Se apreciaron niveles de sensibilidad superiores al 85% con fluoroquinolo -nas, tetracicli-nas, cloranfenicol y
trimetoprim-sul-fametoxazol. Las enzimas hemolisina y gelatinasa estuvieron presentes en el 100% de las cepas. Conclusiones. Las cepas estudiadas muestran baja resistencia a los agentes antimicrobianos analizados, excepto sulfonamida y ampicilina. Al menos dos factores de virulencia de los investigados estuvieron presentes en todas las cepas.
Palabras clave: Vibrio cholerae no-O1, factores de virulencia, susceptibilidad a antimicrobianos, enfermedad diarreica aguda
ABSTRACT
Antimicrobial susceptibility and virulence fac-tors of Vibrio cholerae no-O1 isolates isolated from patients with acute diarrhea.
Introduction. The intestinal infections caused for Vibrio cholerae no-O1 are increasing in the world.
Objective. To know the antimicrobial susceptibil-ity to the drug of choice and alternative ones and to determine the presence of some virulence fac-tors as: extracellular enzymes (Dnase, gelatinase, lecitinase, elastase and hemolysin), HEp-2 cells adherence and presence of fimbrias.
Este artículo está disponible en http://www.revbiomed.uady.mx/pdf/rb081933.pdf
Materials and Methods. A total of 65 isolates of
Vibrio cholerae no-O1 were obtained from patients with acute diarrhea in 7 provincial laboratories of microbiology in Cuba from January to December 2006.
Results. The figures for resistance of sulphonamide a n d a m p i c i l l i n w e r e 3 2 . 3 % a n d 3 0 . 7 % , respectively. Sensitivity levels over 85% were recorded for the antibiotics fluoroquinolones, tetracycline, chloramphenicol and trimetoprim-sulphamethoxazole. Hemolysin and gelatinase enzymes were found in 100% of the isolates. Conclusions. The studied isolates show low re-sistance towards the tested antimicrobials except for sulfonamide and ampicilline. At least two of the investigated virulence factors were found in all isolates.
Key words: Vibrio cholerae no-O1, virulence factor, antimicrobial susceptibility, acute diarrheic disease.
INTRODUCCIÓN
Los miembros del género Vibrio (V.) son bacilos Gram negativos, anaerobios facultativos y oxidasa positiva, que pertenecen a la familia Vi-brionaceae, según la segunda edición del Manual de Bacteriología Sistemática de Bergey (1). Este género está integrado por 65 especies, de las cuales 12 resultan patógenas para el hombre y de ellas las de mayor trascendencia clínica son: V. cholerae, V. parahaemolyticus y V. vulnificus (2). V. cholerae, de acuerdo con el lipopolisacárido capsular, tiene más de 200 serogrupos, divididos en O1 y no-O1. Sólo los serogrupos O1 y O139 son los respon-sables de epidemias y pandemias de cólera (3).
V. cholerae no-O1 es bioquímicamente idéntico a V. cholerae O1 y ha sido asociado con casos esporádicos y brotes de gastroenteritis aguda en diferentes países, aunque la mayoría de las cepas no producen la toxina colérica (TC), sino que sintetizan una enterotoxina termoestable (ST), constituyendo el principal mecanismo asociado a la patogenicidad (4,5). Otros factores de virulencia
presentes son: la producción de las enzimas extracelulares (gelatinasa, elastasa, lecitinasa), citolisinas, citotoxinas, hemolisinas, producción de polisacárido capsular y hemaglutininas (5,6).
En los últimos 20 años las infecciones intestinales por los microorganismos del género
Vibrio se han incrementado a escala mundial (2). Los serogrupos de V. cholerae no-O1 están involucrados en la emergencia de una nueva variante de V. cholerae. Esta hipótesis es sustentada por la génesis de V. cholerae O139, evolucionado por la transferencia horizontal de genes del serogrupo O1 a los serogrupos no-O1, lo que le confirió a este serogrupo el potencial epidémico (7,8). Por lo antes expuesto, se propone conocer la susceptibilidad a los antimicrobianos y la presencia de algunos factores de virulencia asociados a la patogenia, en cepas de V. cholerae no-O1 aisladas de heces de pacientes con Enfermedad Diarreica Aguda (EDA) en el país.
MATERIALES Y MÉTODOS
Se estudiaron 65 cepas de V. cholerae no-O1, aisladas de heces de pacientes con Enfermedad Diarreica Aguda, procedentes de 7 Laboratorios de Microbiología Clínica de los Centros Provinciales de Higiene y Epidemiología del país (Ciudad de la Habana, Camagüey, Las Tunas, Holguín, Granma, Santiago de Cuba y del Centro Municipal de la Isla de la Juventud), pertenecientes al cepario del Laboratorio Nacional de Referencia de Enfermedades Diarreicas Agudas del Instituto de Medicina Tropical “Pedro Kourí” (LNR/EDA/ IPK), en el período comprendido de enero a diciembre de 2006.
las cuales se seleccionaron, al menos, 3 colonias por placa, que fueron inoculadas en los medios de diferenciación primaria Agar Hierro y dos azúcares de Kligler y Agar Hierro-Lisina. Se escogieron aquellas colonias que después del crecimiento durante 18 a 24 horas las cepas presentaron codifi -cación genética para la enzima citocromo oxidasa. A éstas se les realizó el estudio fisiológico corres -pondiente para ubicarlas en el género Vibrio (1).
Para la identificación de especies, la serotipificación (Antígeno O) y el estudio de la sensibilidad al compuesto vibriostático O/129 se realizaron según lo descrito en la literatura especializada (1).
A las cepas se les determinó la susceptibilidad a las drogas antimicrobianas de elección y alternativas utilizando el método de difusión en agar (Kirby–Bauer) (9).
Drogas de elección: tetraciclina (TE) 30µg, doxiciclina (DO) 30µg, cloranfenicol (CL) 30µg, trimetoprim-sulfametoxazol (SXT) 1.25/23.75µg, ampicilina (AM) 10µg y sulfonamidas (SSS) 300µg.
Drogas alternativas: ácido nalidíxico (NA) 30µg, ciprofloxacina (CIP) 5µg y ofloxacina (OF) 5µg.
Para la lectura e interpretación se usó el protocolo recomendado por el Instituto de Estándares de Laboratorio Clínico (CLSI) de los Estados Unidos de América, 2005 (10).
A las cepas se les determinaron los siguientes factores de virulencia: la producción de las enzimas extracelulares (DNasa, gelatinasa, elastasa, lecitinasa y hemolisina), citotoxicidad en células Vero y presencia de fimbrias.
La actividad de DNasa fue investigada según McFaddin (11). Un halo rosado alrededor de las colonias en Agar DNasa con 0.01% de azul de toluidina indicó actividad de nucleasa.
La enzima elastasa se determinó por la metodología establecida por Scharmann (12). Se inoculó por punteo en la superficie del medio Agar elastina, incubándose hasta 7 días a 37ºC. Un halo transparente alrededor de la colonia indicó la presencia de la enzima elastasa.
El estudio de la enzima gelatinasa se realizó por la técnica descrita por Blazevic (13). Se inoculó por punteo en la superficie del medio Agar gelatina, incubándose 24 horas a 37ºC. Una opacidad alrededor de la colonia indicó producción de la enzima gelatinasa.
La presencia de la enzima lecitinasa se determinó por el protocolo recomendado por Boiko (14).
Se inoculó por punteo en la superficie del medio Agar lecitina, incubándose 48 horas a 37ºC. Un halo transparente alrededor de la colonia indicó la presencia de la enzima.
Para demostrar la beta-hemolisina, se siguió la técnica descrita por Robinson (15). Se tomó una asada del cultivo y se inoculó en estría por agotamiento en Agar Triptona-Soya suplementado con sangre de carnero al 5%, incubándose 24 horas a 37ºC. Un halo transparente alrededor de la colonia indicó la presencia de la enzima beta-hemolisina.
Para determinar la capacidad de adherencia a células HEp-2, se procedió según el método de Carrello (16). Se utilizó la línea celular HEp-2 ATCC-CCL 1992, procedente del Laboratorio de Cultivo de Células del IPK. Se preparó una suspensión bacteriana de V. cholerae no-O1, se inoculó 1 ml de la misma en cada pocillo de la placa de células HEp-2 y se incubó a 37ºC en atmósfera de 5% de CO2 durante 90 minutos.
Para determinar la presencia de fimbrias se procedió según el método de Nishikawa (17). Se preparó una suspensión bacteriana y una suspensión al 3% de glóbulos rojos humanos, grupo O de varios donantes. Se mezclaron 20µl de ambas en una lámina portaobjeto, se agitó durante 5 minutos y se consideraron positivas aquellas pruebas en las que se observó aglutinación de los glóbulos rojos.
RESULTADOS
antimicrobianas investigadas, expresándose los resultados en categoría de interpretación de: sensible, intermedio y resistente.
Cuadro 1
Susceptibilidad a los antimicrobianos de las cepas de Vibrio cholerae no-O1
Drogas antimicrobianas
Sensible Intermedio Resistente
No. de
cepas %
No. de
cepas %
No. de
cepas %
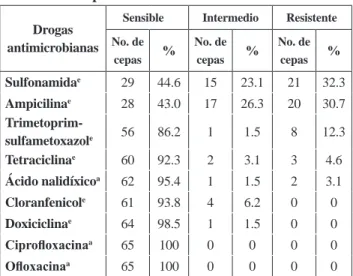
Sulfonamidae 29 44.6 15 23.1 21 32.3
Ampicilinae 28 43.0 17 26.3 20 30.7
Trimetoprim-sulfametoxazole 56 86.2 1 1.5 8 12.3
Tetraciclinae 60 92.3 2 3.1 3 4.6
Ácido nalidíxicoa 62 95.4 1 1.5 2 3.1
Cloranfenicole 61 93.8 4 6.2 0 0
Doxiciclinae 64 98.5 1 1.5 0 0
Ciprofloxacinaa 65 100 0 0 0 0
Ofloxacinaa 65 100 0 0 0 0
Leyenda: edrogas de elección, adrogas alternativas
Analizando el resultado de las cepas frente a las drogas de elección se observó que el 32.3% y el 30.7% de las cepas presentaron resistencia a la sulfonamida y a la ampicilina respectivamente. Se obtuvieron valores de sensibilidad superiores al 85% con los antibióticos: doxiciclina, cloranfenicol, tetraciclina y trimetoprim-sulfametoxazol.
En relación con la susceptibilidad a las drogas alternativas se apreció 100% de sensibilidad para la ciprofloxacina y ofloxacina y de 95.4% para el ácido nalidíxico.
De las 65 cepas, 2 (3%) resultaron multirre-sistentes (resistencia a 3 o más grupos de drogas antimicrobianas). El patrón de multirresistencia es TE-, NA-, SXT-.
Todas presentaron, al menos, un factor de virulencia. Las enzimas hemolisina y gelatinasa estuvieron presentes en el 100% de las cepas investigadas y la adherencia a células HEp-2 se observó en el 87.6% de las cepas. El resto de las enzimas, elastasa, lecitinasa y DNasa resultaron positivas en 86.1%, 80% y 73.8%, respectivamente (Figura1).
100 100
87.6 86.1
80 73.8
66.1
0 20 40 60 80 100 120
Factores de virulencia
Porcentajes de positividad
Hemolisina Gelatinasa Adherencia a células Hep-2 Elastasa Lecitinasa DNasa Hemaglutinación
Figura 1. Presencia de factores de virulencia
DISCUSIÓN
Los miembros de la familia Vibrionaceae son agentes etiológicos de EDA en áreas tropicales y subtropicales de todos los continentes. Desde las décadas pasadas, se ha incrementado a escala mun-dial la incidencia de gastroenteritis aguda de forma esporádica y en brotes por V. cholerae no-O1 (2).
Analizando en este estudio los resultados de la susceptibilidad a los antimicrobianos en cepas de V. cholerae no-O1, se observó una resistencia moderada a la sulfonamida. Similar nivel de resistencia fue obtenido por Iwanaga y cols., en Laos (18). Sin embargo, el valor de resistencia para la sulfonamida del presente estudio no coincide con el obtenido en una investigación epidemiológica hecha por Mohanthy y cols., entre los años 1998 y 2002, en un hospital de nivel terciario en el norte de la India, sobre los patrones de resistencia contra los antimicrobianos en cepas de V. cholerae, registrándose 90.2% de resistencia a la sulfonamida en estas cepas (19).
En el LNR/EDA/IPK, Bravo y cols. en 2000 y 2004 realizaron varios estudios sobre el comportamiento de la susceptibilidad a los antimicrobianos en cepas de V. cholerae no-O1 aisladas de pacientes con diarreas, observándose valores de sensibilidad por encima del 80% a tetraciclina, ácido nalidíxico y trimetoprim-sulfametoxazol (22,23), encontrándose en esta investigación correspondencia con los resultados obtenidos por los autores citados anteriormente.
El patrón de multirresistencia (TE-, NA-,
SXT-) encontrado en la presente investigación
coincide con el hallado en Perú (24), no así con el obtenido en la India y en Argentina (25,26). La diferencia pudiera deberse a la cantidad de cepas estudiadas en el presente estudio y al uso racional de los agentes antimicrobianos en Cuba.
Las infecciones intestinales por V. cholerae
no-O1 generalmente provocan gastroenteritis ligera, considerándose la diarrea autolimitada, siendo necesaria la prescripción de la terapia antimicrobiana en niños y ancianos con diarreas severas, en pacientes con factores de riesgos (diabetes mellitus, malnutrición, inmunodeficiencias, entre otros) y en las infecciones extraintestinales (septicemia, celulitis, otitis, etc.) (27).
Son escasos los estudios sobre factores de virulencia (producción de enzimas extracelulares, presencia de fimbrias y adherencia a células HEp-2) realizados en América Latina y en el Caribe, en cepas de V. cholerae no-O1 aisladas de pacientes con EDA (28).
La hemolisina producida por V. cholerae
O139 y por V. cholerae no-O1 es idéntica a la hemolisina El Tor de V. cholerae O1 y es considerada un factor de virulencia para causar diarreas, especialmente cuando las cepas carecen de toxinas bien definidas como la TC (29).
Observando el comportamiento de este factor de virulencia en nuestro trabajo, todas las cepas produjeron la enzima hemolisina. Este resultado es similar a los encontrados por Singh y cols. en la India y por Bininost y cols. en Argentina (30,28).
Cuando se revisa la bibliografía científica internacional en relación con la producción de proteasas, lipasas, citotoxinas, entre otras, en el género Vibrio (7,14), se señala que las mismas son comunes en este género, implicadas en la producción de hemorragias, edema y en la alteración de los sistemas de defensa del organismo, favoreciendo el desarrollo de los procesos infecciosos. En las cepas de este estudio, se evidenció la producción de las enzimas DNasa, gelatinasa, lecitinasa y elastasa, coincidiendo con lo señalado en las monografías revisadas sobre propiedades y mecanismos de virulencia en las especies del género Vibrio, realizada por los autores antes citados. La caracterización fenotípica permite establecer significativos avances en la fisiopatología de las infecciones intestinales causadas por este microorganismo.
La adherencia bacteriana a las células del huésped susceptible representa un paso importante para la colonización tisular y es mediada por moléculas llamadas adhesinas, destacándose en V. cholerae la proteína de membrana externa (Omp U) y la toxina reguladora de Pili (tox R) (31). En lo relativo a la alta capacidad de adherencia de las cepas de V. cholerae no-O1 de este trabajo a la línea celular HEp-2, se observa concordancia con lo publicado por Dalsgaard y cols. y por Israll y cols., apreciándose que predominó un patrón de adherencia difuso en las cepas de V. cholerae O1 y V. cholerae no-O1 (32,33).
O t r o f a c t o r d e v i r u l e n c i a e n l o s microorganismos pertenecientes al género
Vibrio es la presencia de fimbrias, estructura que
favorece la colonización y adherencia microbiana (34). En esta investigación se observó, a través de la prueba de hemaglutinación con eritrocitos humanos, la presencia de fimbrias en las cepas objeto de estudio. Nuestros resultados concuerdan con los datos obtenidos en experimentos sobre mecanismos de enteropatogenicidad, expresión de hemaglutininas y patrones de virulencia en cepas de V. cholerae O1, V. cholerae O139 y V. cholerae
diarrea aguda, en los Estados Unidos de América y en Rusia, que demostraron porcentajes de positividad para este factor de virulencia de 61.5%, 55.7% y 70%, respectivamente (35,36).
Los mecanismos de producción de virulencia no están bien esclarecidos en estos serogrupos (37). Los factores de virulencia estudiados en las cepas que incluyó esta investigación pudieran ayudar a dilucidar la patogénesis de V. cholerae
no-O1 como agente etiológico de la EDA. Los resultados de esta investigación muestran un bajo grado de resistencia a los agentes antimicrobianos analizados, excepto sulfonamida y ampicilina; estos resultados pudieran deberse a que las cepas en estudio han interactuado poco con los grupos de drogas antimicrobianas antes señalados, debido a que en Cuba se hace un uso racional de los antibióticos. Sin embargo, es necesario continuar una vigilancia estricta de la susceptibilidad bacteriana a las drogas de elección en estos microorganismos, para detectar cambios en los patrones de resistencia en las cepas circulantes en las diferentes provincias y regiones del país.
En este trabajo se describen, por primera vez, algunas propiedades de virulencia de V. cholerae
no-O1 aislados de pacientes con EDA en Cuba. Futuras investigaciones deben relacionarse con los mecanismos genéticos de virulencia y de resistencia contra los antimicrobianos en estos microorganismos.
REFERENCIAS
1. Garrity GM, Bell JA, Lilburm TG. Taxonomic outline of the prokaryotes. En: Bergey ′s Manual of Systematic Bacteriology. 2 nd ed. Release 4.00; Oct 2004. [Citado el 10 de febrero del 2005]. Disponible en URL: http:// dx.doi.org/10.1007/bergeysoutline
2. Lesmana M, Subekti DS, Tjaniadi P, Simanjuntak CH, Punjabi NH, Campbell JR, et al. Spectrum of Vibrio species associated with acute diarrhea in North Jakarta, Indonesia. Diagn Microbiol Infect Dis 2002; 43:91-7.
3. Chatterjee SN, Chaudhuri K. Lipopolysaccharides of Vibrio cholerae. I. Physical and chemical characteriza-tion. Biochem Biophys Acta 2003; 1639:65-79.
4. Nair GB, Oku Y, Takeda Y, Ghosh A, Ghosh RK, Chattopadhyay S, et al. Toxin profiles of Vibrio chol-erae non-O1 from environmental sources in Calcutta, India. Appl Environ Microbiol 1988; 54:3180-2. 5. Farfan M, Miñana D, Fusté MC, Lorén JG. Genetic
relationships between clinical and environmental Vibrio cholerae isolates based on multilocus enzyme electro-phoresis. Microbiology 2000; 146:2613-26.
6. Shinoda S. Protein toxins produced by pathogenic vibrios. J Nat Toxins 1999; 8:259-69.
7. Bhattacharya MK, Dutta D, Bhattacharya SK, Deb A, Mukhopadhyay AK, Nair GB, et al. Association of a disease approximating cholera caused by Vibrio cholerae of serogroups other than O1 and O 139. Epi-demiol Infect 1998; 120:1-5.
8. Boyd EF, Waldor MK. Alternative mechanism of cholera toxin acquisition by Vibrio cholerae: general-ized transduction of CTXФ by bacteriophage CP-T1. Infect Immun 1999; 67:5898-905.
9. Bauer AW, Kirby WM, Sherris JC, Turck M. Anti-biotic susceptibility testing by a standarized single disk method. Am J Clin Pathol 1966; 45:493-6.
10. Clinical and Laboratory Standards Institute. Per-formance standards for antimicrobial susceptibility testing: eleventh informational supplement. Wayne; 2005. CLSI Document M100-S15.
11. Mc Faddin JF. Pruebas bioquímicas para la identifi -cación de bacterias de importancia clínica. Baltimore: William and Wilkins; 2002.
12. Scharmann W. Elastase in Pseudomonas and Aeromonas Zentralbl Bakteriol 1972; 220:435-42.
13. Blazevic DJ, Ederer GM. Principles of biochemical test in diagnostic microbiology. New York, J. Wiley. 1975.
14. Boiko AV. Pathogenicity factors of various Vibrios and Aeromonas. Zh Mikrobiol Epidemiol Immunobiol 2000; 79:104-8.
15. Robinson J, Beaman J, Wagener L, Burke V. Com-parison of direct plating with the use of enrichment culture for isolation of Aeromonas spp from faeces. J Med Microbiol 1986; 22:315-7.
16. Carrello A, Solburn K, Budden J, Chong B. Adhe-sion of clinical and environment Aeromonas isolates to HEp-2 cells. J Med Microbiol 1988; 26:19-27. 17. Nishikawa Y, Kimiura T, Kishi T. Mannose-resistant
adhesion of motile Aeromonas to INT 407 cells and the differences among isolates from humans food and water. Epidemiol Infect 1991; 107:171-9.
19. Mohanthy S, Kapil A, Das BK. Seasonality and antimicrobial resistance pattern of Vibrio cholerae in a tertiary care hospital of North India. Trop Doct 2004; 43:249-51.
20. Morris JG, Tenney JH, Drusmo GL. In vitro sus-ceptibility of pathogenic Vibrio species to norfloxacin and six other antimicrobial agents. Antimicrob Agent Chemother 1985; 28:442-5.
21. Tjaniadi P, Lesmana M, Subekti D, Machpud N, Komalarini S, Santoso W. Antimicrobial resistance of bacterial pathogens associated with diarrheal patients in Indonesia. Am J Trop Med Hyg 2003; 68:666-70.
22. Bravo L, Cabrera L, Ramírez M, Castañeda N, Fernández E, Garrigó E. Sensibilidad antimicrobiana de cepas de Vibrio cholerae no-O1 aisladas de pacientes en Cuba. Rev Esp Quimioterap 2004; 17:200-1. 23. Bravo L, Ramírez M, Maestre JL, Llop A, Cabrera
R, García B. Vibrio cholerae no-O1 toxigénico. Rev Cubana Med Trop 2000; 52:106-9.
24. Ibarra JO, Flores LE, García RH, Alvarado T.
Estudio preliminar de la resistencia de Vibrio cholerae O1, brote epidémico 1998, a antimicrobianos. Rev Peru Biol 2000; 7:3-6.
25. Neelam K, Manjula M, Vikas G, Varsha G. Out-break of cholera in & around Chandigarh during two successive years (2002,2003). Indian J Med Res 2005; 122:404-407.
26. Organización Panamericana de la Salud. Resistencia antimicrobiana en las Américas: Magnitud del problema y su contención. Washington DC: OPS; 2003. 27. World Health Organization. Management of the
patient with cholera. [Citado el 20 de julio del 2005]. Disponible en URL: http://www.who.int/csr/resources/ publications/cholera/whocddser9115rev1.pdf. 28. Bidinost C, Saka HA, Aliendro O, Sola C,
Panceta-Dutttari G, Carranza P, et al. Virulence factors of non-O1 non-O139 Vibrio cholerae isolated in Córdoba,
Argentina. Rev Argentina Microbiol 2004; 36:158-63. 29. Sack D, Sack R, Nair G, Siddique A. Cholera. Lancet
2004; 363:223-33.
30. Singh DV, Matte MH, Matte GR, Jiang S, Sabena F, Shukla BN. Molecular analysis of Vibrio cholerae O1, O139, non-O1, and non-O139 strains: Clonal relation-ships between clinical and environmental isolates. Appl Environ Microbiol 2001; 67:910-21.
31. Israll AM, Balotescu C, Lazar V, Cernat R, Dinu C.
Factors influencing the capacity of cellular substrate
adherencia of Vibrio cholerae O1 and non O1 strains. Bacteriol Virusol Parazitol Epidemiol 2002; 47:119-24. 32. Dalsgaard A, Albert MJ, Tavior DN, Shimada T,
Meza R, Serichantelergs O, et al. Vibrio cholerae non-O1 serogrups obtained from an outbreak of diarrhea in Lima, Peru. J Clin Microbiol 1995; 33:2715-22. 33. Israll A, Balotescu C, Camian M, Dinu C, Bucurenci
N. Comparative study of different methods for detection of toxic and other enzymatic factors in Vibrio cholerae strains. Roum Arch Microbiol Immunol 2004; 63:63-77.
34. Chiavelli DA, Marsh JW, Taylor RK. The Mannose– sensitive hemagglutinin of Vibrio cholerae promotes adherence to zooplankton. Appl Environm Microbiol 2001; 67:3220-5.
35. Mathur J, Waldor MK. The Vibrio cholerae ToxR-regulated porin OmpU confers resistance to antimicro-bial peptides. Infect Immun 2004; 72:3577-83. 36. Telesmanich NR, Bezuglova EV, Mishankin MB,
Vinokur NI. Role of lectin (hlyA) in the hemotytic and hemaglutinating activity Vibrio cholerae. Zh Microbiol Epidemiol Immunobiol 2004; 5:12-6.