Los abajo firmantes, convocados por el Programa de Doctorado en Ciencias Agrarias de la Universidad de Caldas, hemos revisado el informe final de la tesis doctoral:
ASPECTOS REPRODUCTIVOS Y ENDOCRINOS DEL CICLO
ESTRAL DE LA HEMBRA BUFALINA (
Bubalus bubalis
)
Presentada por: JORGE ALBERTO SÁNCHEZ VALENCIA Como requisito parcial para obtener el título de Doctor en Ciencias Agrarias
y certificamos su aprobación
HENRY MESA ECHEVERRY Director
AURELIANO HERNANDEZ V. Evaluador – Asesor
ASPECTOS REPRODUCTIVOS Y ENDOCRINOS DEL CICLO
ESTRAL DE LA HEMBRA BUFALINA (
Bubalus bubalis
)
Tesis Presentada a:
Programa de Doctorado en Ciencias Agrarias Facultad de Ciencias Agropecuarias
Universidad de Caldas
Como requisito parcial Para obtener el título de Doctor en Ciencias Agrarias
Por
JORGE ALBERTO SÁNCHEZ VALENCIA
Henry Mesa Echeverry., Ph.D., Director de Tesis
AGRADECIMIENTOS
A mi familia, por su apoyo incondicional.
Al Fondo Nacional de Financiamiento para la Ciencia, la Tecnología y la Innovación, Francisco José de Caldas-COLCIENCIAS, a la Vicerrectoría de Investigaciones y Posgrados de la Universidad de Caldas, a la Universidad de Caldas y al Fondo Bufalero del Centro.
Al Profesor Luis Fernando Uribe, por su orientación
A los Medicos Veterinarios: Leonardo Escobar, Fernando Meneses C, Jhoana Suarez, Santiago Meza. Por su colaboración en la elaboración de las pruebas hormonales y el desarrollo de la investigación.
DEDICATORIA
Dedico esta tesis a mi madre, mi hermana, mi sobrina, mi hijo y mi esposa quienes fueron un apoyo incondicional durante el tiempo en que escribí esta tesis, siempre alentándome para continuar.
TABLA DE CONTENIDO
LISTA DE FIGURAS ... 7
LISTA DE TABLAS ... 8
RESUMEN ... 10
CAPÍTULO I ... 12
1. INTRODUCCIÓN ... 12
1.1 HIPÓTESIS ... 13
1.2 OBJETIVOS ... 14
CAPÍTULO II ... 16
2. REVISIÓN DE LITERATURA... 16
2.1 CICLO ESTRAL EN LA HEMBRA BUFALINA ... 16
2.2 INDICADORES CELULARES ... 28
2.3 ESTACIONALIDAD REPRODUCTIVA ... 31
2.4 MORTALIDAD EMBRIONARIA ... 35
2.5 SINCRONIZACIÓN DEL ESTRO Y DE LA OVULACIÓN ... 36
BIBLIOGRAFÍA ... 46
CAPÍTULO III ... 55
ESTACIONALIDAD REPRODUCTIVA DE LA HEMBRA BUFALINA (Bubalus bubalis) ... 55
RESUMEN ... 55
3.1 INTRODUCCIÓN ... 56
3.2 MATERIALES Y MÉTODOS ... 58
3.3 RESULTADOS ... 61
3.4 DISCUSIÓN ... 66
3.5 CONCLUSIONES ... 69
LITERATURA CITADA ... 69
CAPITULO IV... 75
DINÁMICA FOLICULAR DURANTE EL CICLO ESTRAL NATURAL EN LA HEMBRA BUFALINA (Bubalus bubalis) ... 75
RESUMEN ... 75
4.1 INTRODUCCIÓN ... 77
4.2 MATERIALES Y MÉTODOS ... 79
4.3 RESULTADOS ... 82
4.4 DISCUSIÓN ... 89
4.5 CONCLUSIONES ... 92
LITERATURA CITADA ... 92
CARACTERIZACIÓN DEL DESARROLLO FOLICULAR DURANTE EL CICLO ESTRAL SINCRONIZADO CON UN PROTOCOLO OVSYNCH EN HEMBRAS BUFALINAS (Bubalus
bubalis) ... 98
RESUMEN ... 99
ABSTRAC ... 100
5.1 INTRODUCCIÓN ... 101
5.2 MATERIALES Y MÉTODOS ... 102
5.3 RESULTADOS ... 105
5.4 DISCUSIÓN ... 108
5.5 CONCLUSIONES ... 109
5.6 AGRADECIMIENTOS ... 109
LITERATURA CITADA ... 110
CAPÍTULO VI... 115
6. DISCUSIÓN GENERAL ... 115
LISTA DE FIGURAS
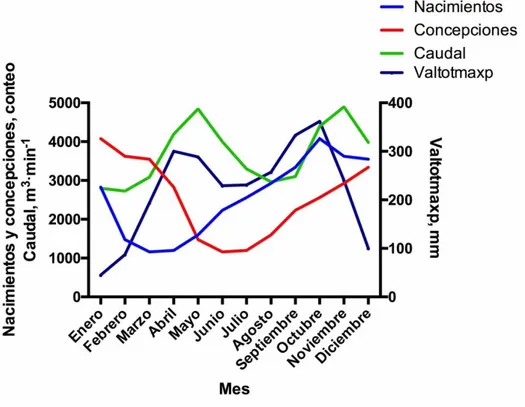
Figura 1. Relación entre los nacimientos, la concepción y el caudal máximo (m3/seg) y
valores totales máximo de precipitación (mm) en la hacienda La Suiza. 1970-2013 ... 63
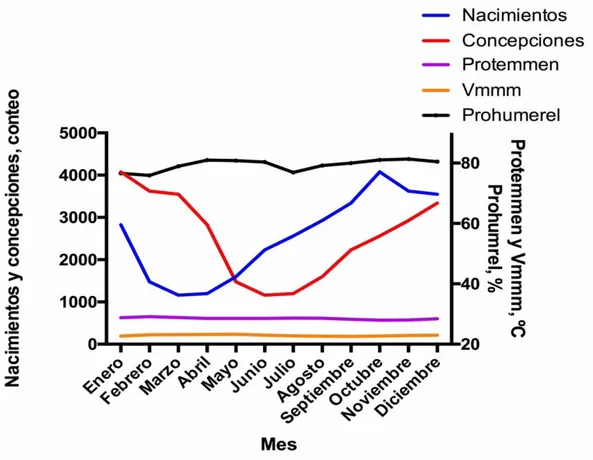
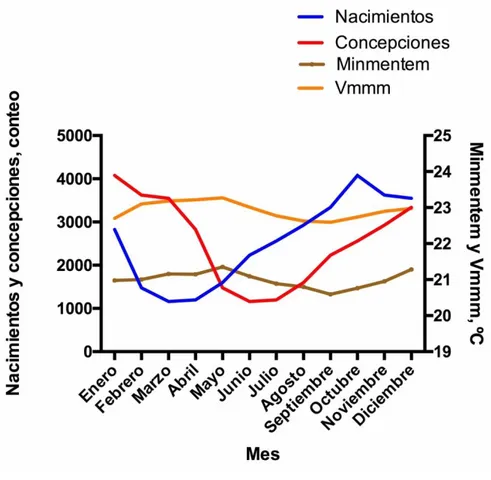
Figura 2. Relación entre los nacimientos, la concepción y los valores medios mensuales de temperatura (°C), valores medios máximos mensuales de temperatura (°C) y el promedio mensual de humedad relativa (%) en la hacienda La Suiza. 1970-2013 ... 64
Figura 3. Relación entre los nacimientos, la concepción y los valores mínimos mensuales de temperatura (°C) y los valores medios máximos mensuales de temperatura (°C) en la hacienda La Suiza. 1970-2013. ... 65
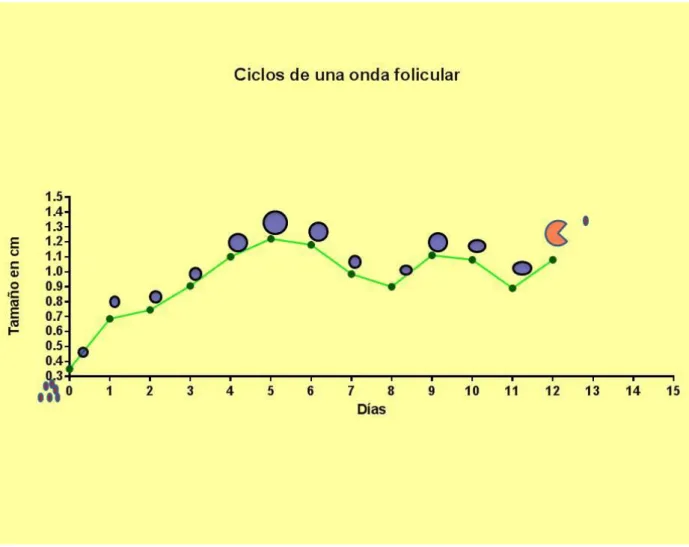
Figura 4. Crecimiento folicular en hembras bufalinas con ciclos de una onda folicular ... 83
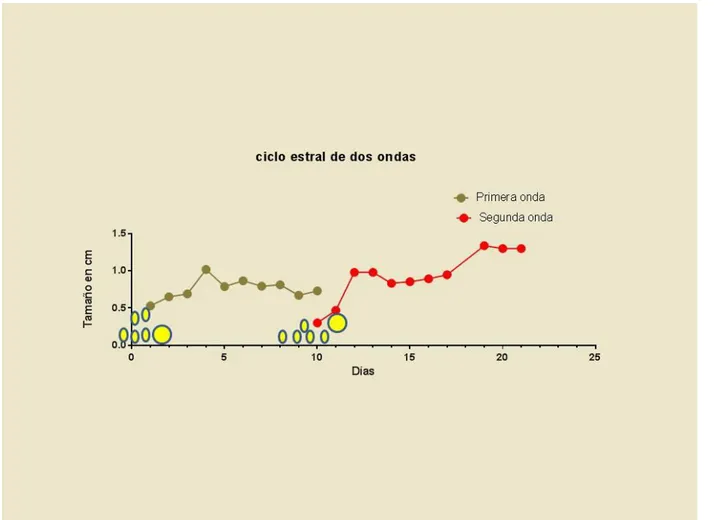
Figura 5. Crecimiento folicular en hembras bufalinas con ciclos de dos ondas foliculares ... 84
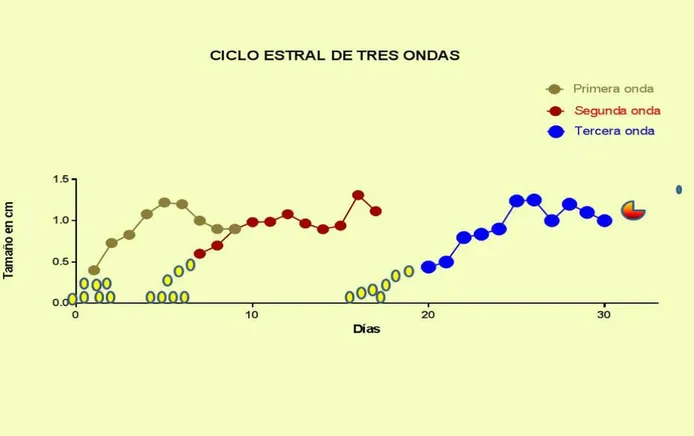
Figura 6. Crecimiento folicular en hembras bufalinas con ciclos de tres ondas foliculares ... 85
Figura 7. Representación esquemática del protocolo de sincronización Ovsynch en búfalas (Bubalus bubalis). ... 104
LISTA DE CUADROS
Cuadro 1. Descripción de las variables evaluadas en el estudio: “Estacionalidad reproductiva de la hembra bufalina (Bubalus bubalis) en Colombia” ...60
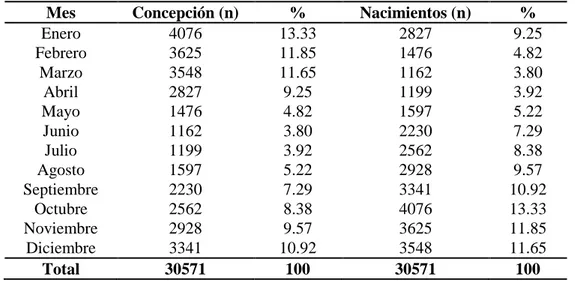
Cuadro 2. Frecuencia mensual de nacimientos y de concepciones de búfalas del Magdalena Medio colombiano (1970-2013) ...62
Cuadro 3. Correlaciones de Spearman (Rho) entre las variables climatológicas en la presentación de concepciones y nacimientos en búfala del Magdalena Medio colombiano ...62
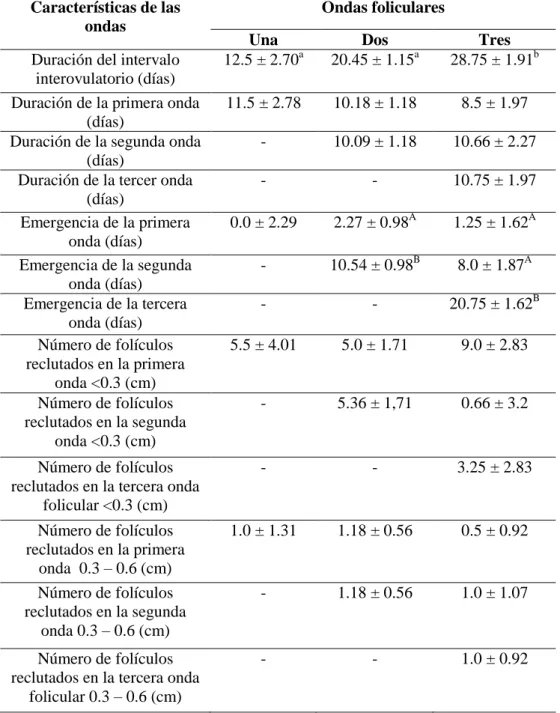
Cuadro 4. Características del ciclo estral natural y sus ondas foliculares en búfalas colombianas 86
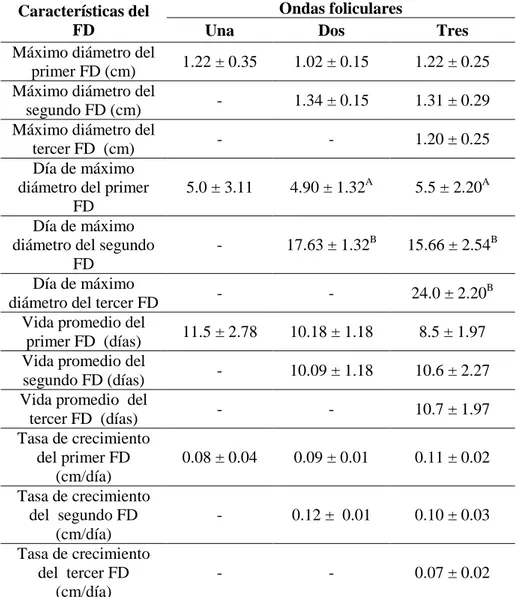
Cuadro 5. Características del folículo dominante durante el ciclo estral natural en búfalas
colombianas ...88
Cuadro 6. Respuesta ovárica en búfalas (Bubalus bubalis) sincronizadas con un protocolo
Ovsynch. ...107
ASPECTOS REPRODUCTIVOS Y ENDOCRINOS DEL CICLO
ESTRAL DE LA HEMBRA BUFALINA (
Bubalus bubalis
)
JORGE ALBERTO SÁNCHEZ VALENCIA
Universidad de Caldas Facultad de Ciencias Agropecuarias Programa de Doctorado en Ciencias Agrarias
RESUMEN
23,08% de los animales presentó una onda folicular y el 76,92% la formación de una nueva onda. El 71,15% de las búfalas ovuló a las 48 h. siguientes a la segunda aplicación de GnRH; el diámetro del FD fue 1,18 cm el día 9. La tasa de preñez fue de 46,15%.. En conclusión, los búfalos del experimento presentan un patrón reproductivo durante todo el año, con una estacionalidad relacionada con la temperatura ambiente, el régimen de lluvias y la humedad relativa. La dinámica folicular de la búfala durante el ciclo estral natural se caracteriza por presentar patrones de una, dos y tres ondas, con predominio de dos ondas foliculares, con respuesta eficiente al tratamiento hormonal con Ovsynch.
CAPÍTULO I 1. INTRODUCCIÓN
Los búfalos domésticos (Bubalus bubalis), son importantes en la economía agropecuaria de muchos países de Asia y América Latina, están adaptados a climas subtropicales y tropicales, tienen gran potencial para la producción de carne, leche y trabajo, en especial en suelos pobres de baja fertilidad (Singh y Adams 2000; Chaikhun y col., 2010; Perera, 2011). Colombia cuenta con un inventario superior a 300.000 búfalos, diseminados por todo el territorio nacional y en los diferentes hábitats del país, que comprenden variados pisos térmicos, desde el nivel del mar hasta 3000 metros de altitud (Asociación Colombiana de Criadores de Búfalos, 2009). El búfalo tiene una alta capacidad de adaptación y rusticidad lo que hace atractiva su producción.
mundial, se encuentran pocos estudios sobre el ciclo estral y la dinámica folicular en búfalas en comparación a lo reportado con el ganado bovino tanto en Bos taurus, como en Bos indicus (Presicce y col., 2002; Warriach y Ahmad, 2007; Neglia y col., 2008;
Murugavel y col., 2009; Karen y Darwish, 2010).
1.1 HIPÓTESIS
Las búfalas presentan características fisiológicas y reproductivas únicas bajo condiciones de trópico bajo.
Sobre las características de estacionalidad reproductiva:
Los búfalos en sistemas de producción en trópico bajo, muestran un patrón de
estacionalidad reproductiva la que es influenciada por factores medioambientales.
Sobre la fisiología reproductiva del ciclo estral las hipótesis son:
El ciclo estral difiere en su duración y depende del número de ondas foliculares. El patrón predominante de ondas foliculares en la búfala se comporta similar a lo
reportado en otros bovinos.
Las características del folículo dominante coinciden con lo reportado en bovinos El número de folículos reclutados en cada onda folicular es independiente del
número de ondas presentes en cada ciclo estral.
Las concentraciones de progesterona dependen del tamaño del cuerpo lúteo. En la búfala, el tratamiento hormonal, permite sincronizar la ovulación.
La dinámica folicular en la búfala es influenciada por la presencia de un Cuerpo
1.2 OBJETIVOS
1.2.1 Objetivo general
Describir el desarrollo folicular y la respuesta endocrina de la búfala Trinitaria
durante el ciclo estral natural y mediante la utilización de un protocolo de sincronización en Colombia.
1.2.1 Objetivos específicos
Determinar el patrón de estacionalidad reproductiva de un hato bufalino en el
trópico bajo colombiano.
Relacionar las variables climatológicas: valores mínimos, medios y máximos
mensuales de caudales; valores medios, máximo mensuales de nivel del río, valores totales mensuales de brillo solar, valores mínimos y medios mensuales de temperatura, valores medios mensuales de humedad relativa e índice de temperatura y humedad con la época de concepción y nacimiento en condiciones del trópico bajo Colombiano.
Caracterizar el desarrollo folicular y la ovulación durante el ciclo estral natural y
el ciclo estral sincronizado con un protocolo Ovsynch en la hembra bufalina. Determinar el comportamiento de la secreción de P4 durante el ciclo estral natural
en la hembra bufalina.
Definir la relación existente entre la concentración de los niveles de P4 y el tamaño
del cuerpo lúteo durante el ciclo estral natural en la hembra bufalina.
Establecer la relación existente entre las concentraciones plasmáticas de P4 y la
Evaluar el porcentaje de animales que presentan estro y la tasa de preñez, durante
CAPÍTULO II
2. REVISIÓN DE LITERATURA
2.1 CICLO ESTRAL EN LA HEMBRA BUFALINA
La duración del ciclo estral es definida como el período de tiempo trascurrido entre dos estros consecutivos. Los búfalos son poliéstricos que exhiben estacionalidad; diversos estudios han caracterizado el ciclo estral de la búfala con resultados disímiles, dependiendo del país, condiciones medioambientales y el tipo de animal evaluado, en general, se reporta una duración promedio del intervalo interovulatorio (IIO) entre 16 a 33 días y de 18 a 24 días (Baruselli y col., 1997; Barkawi y col., 2009; Nam, 2010; Perera, 2011).
(15-17 días) ocurren con mayor frecuencia, siendo un patrón normal de dinámica folicular que favorece las tasas de concepción y parto (Abdalla, 2003).
2.1.1 Dinámica folicular
La dinámica folicular es el proceso de continuo crecimiento y de regresión de un grupo de folículos antrales, uno de los cuales se desarrolla a folículo pre-ovulatorio y abarca las fases de: crecimiento, estática y regresión (Baruselli y col., 1997; Ireland y col., 2000; Das y col., 2013).
En general, el desarrollo de una onda folicular involucra la organización y el crecimiento simultáneo de una cohorte folicular (Peter y col., 2009). En la especie bufalina, las primeras investigaciones de la fisiología del ciclo estral las realizó Rajaloski en 1960 (Das y col., 2013) basados en observaciones histológicas de ovarios recolectados en plantas de sacrificio, lo que por primera vez describe un patrón de crecimiento folicular en ondas para la dinámica ovárica de la búfala. Kumar y col., (1997) evaluaron las poblaciones foliculares presentes en 250 ovarios de animales sacrificados en planta de beneficio y hallaron que las proporciones de áreas de folículos primordiales y estroma en ovarios de animales adultos contenían 14,4 ± 6,2 % de tejido luteal, lo que a su vez es señalada por los autores, como razón de las bajas tasas de recuperación de oocitos de ovarios que presentaban un cuerpo lúteo, el cual restringe el crecimiento folicular.
otros estadios del ciclo estral, un indicador de que la onda folicular comienza en este período. Azawi y col., (2009) al realizar morfometría de folículos en ovarios obtenidos en plantas de sacrificio concluyeron que, la búfala presenta un patrón de desarrollo folicular de dos ondas en el 64,6%, mientras 35,4% lo describen de una onda folicular.
En la década de los noventa, se inició el estudio ecográfico en la dinámica folicular de los bovinos, entre ellos los bufalinos; los estudios de Taneja y col., (1996) confirmaron los postulados de Danell en 1987, coincidiendo en que los folículos de las búfalas se desarrollan en ondas tal como sucede en los bovinos. Estas ondas foliculares se presentan, de una, dos o tres por cada ciclo estral (Campanile y col., 2010). Al describir el ciclo estral natural en búfalas de raza Murrah en la India, Taneja y col., (1996), muestran que el desarrollo de los folículos antrales en el ovario sucede en un patrón de una o dos ondas foliculares por ciclo estral, así que, durante cada onda es seleccionado un único folículo que se hace dominante, en tanto que, los demás folículos de la misma onda hacen regresión dependiendo si la fase de dominancia está asociada a la luteólisis (Taneja y col., 1996; Manik y col., 2002). Baruselli y col., (1997) describen que durante el ciclo estral en búfalas Murrah, la dinámica folicular es similar a la que ocurre en los demás bovinos, y que el patrón de dos ondas foliculares por ciclo estral es el más común, pero también, se presentan ciclos estrales con una y tres ondas foliculares.
2.1.2 Patrón de ondas foliculares
Ganguly y col., 2010). En la fase de selección, Baruselli y col., (1997) reportan un promedio de 6 a 8 folículos reclutados por cada onda folicular siendo un número menor que el presente en los demás bovinos, aunque, Barkawi y col., (2009) describen que al inicio de cada onda folicular hay una gran cantidad de folículos reclutados en vacas bufalinas y mayor porcentaje de folículos dominantes no atrésicos durante el diestro prematuro, comparado con el diestro tardío y durante el proestro / estro, eran viables el 100% de los folículos dominantes.
foliculares se asocia a un ciclo estral y fase luteal prolongadas (Baruselli y col., 1997; Singh y col., 2000; Awasthi y col., 2006; Warriach y Ahmad, 2007; Barkawi y col., 2009). El patrón de crecimiento folicular más común en las búfalas corresponde al de dos ondas foliculares, presente entre el 63 al 83 % de los casos (Taneja y col., 1996; Baruselli y col., 1997; Ali y col., 2003), seguido por los patrones de tres ondas foliculares (25 % a 33%) y 3.3% de casos de una onda folicular (Manik y col., 2002; Baruselli y col., 1997; Warriach y Ahmad, 2007; Barkawi y col., 2009; Yindee y col., 2010).
2.1.3 Emergencia de las ondas foliculares
Distintos autores han descrito ciclos estrales de una onda en búfalas con ciclos estrales con emergencia de un folículo solitario el día 1,4 ± 0,24 (ovulación día = 0) que persiste por 10,7 ± 0,66 días hasta ser el folículo ovulatorio (Ali y col., 2003; Awasthi y col., 2006); en patrones de dos ondas foliculares Baruselli y col., (1997) comunican que, el inicio de una nueva onda se presenta en los días 1,16 ± 0,5 y 10,83 ± 1,09 (ovulación = día 0) y en vacas bufalinas con ciclos estrales de tres ondas foliculares, tiene lugar los días 1,10 ± 0,32; 9,30 ± 1,25 y 16,80 ± 1,22; los ciclos de dos o tres ondas difieren entre sí (p<0,05), en lo que respecta al diámetro máximo del primer folículo dominante (15,0 ± 2,33 vs. 11,90 ± 1,68 mm) y el diámetro del folículo ovulatorio, a saber, (15,50 ± 1,60 vs. 13,40 ± 1,30 mm). Barkawi y col., (2009), reportan que para ciclos de dos ondas la primera onda folicular emerge el día 1,1 ± 0,3 y la segunda el día 11,0 ± 0,4; mientras que en búfalas de tres ondas foliculares, la primera onda curre el día 0,7 ± 0,3, la segunda el día 8,4 ± 0,4 y la tercera onda folicular sucede el día 16,3 ± 0,5.
2.1.4 Desarrollo folicular y desviación
2.1.5 Comportamiento del estro
La búfala presenta manifestaciones de estro menos evidentes que las demás hembras bovinas, con un comportamiento de monta distinto a los demás rumiantes, influenciado por diferentes factores como el clima, temperatura, fotoperíodo y nutrición, (Suthar y Dhami, 2010). Los principales signos de estro son: la inquietud, la edematización de la vulva y la secreción de moco vaginal; otros signos reportados incluyen: aumento del tono uterino, micción, el seguimiento y monta por el toro y la descarga de moco cristalino; las manifestaciones menos observadas son la posición de la cola y el apoyar la barbilla (Baruselli y col., 1997; Awasthi y col., 2007; Perera, 2008; Mohan y col., 2010; Yindee y col., 2010; Ayen y col., 2012) que por lo general, ocurren entre las 7: 00 y 20:00 (Yindee y col., 2010). El cenit se presenta en las horas nocturnas lo que constituye un limitante para los programas de IA (Suthar y Dhami, 2010). Jacomini y col., (1991), observaron el ciclo estral de 21 búfalas y demostraron que 56,45 % de los estros se iniciaron en el período nocturno. Con una observación cada cuatro horas, fueron detectados el 100% de los estros y para cada ocho y doce horas, la detección fue de 96,82 % y 82,54%; además, las manifestaciones de estro en la especie bufalina, son menos intensas comparado con los bovinos, debido a concentraciones más bajas de 17-β estradiol encontradas durante el estro normal (Carvalho y col., 2002). Sin embargo Banu y col., (2012) reportan una tasa de detección de estro en la especie que varía desde el 28% al 68% cuando son realizados por por observadores entrenados.
2.1.6 Feromonas
por el macho. Cuando el búfalo identifica los compuestos químicos durante el estro realiza el flehmen y sigue oliendo de manera repetida (Rajanarayanan y col., 2004; Rajanarayanan y col., 2011., Muthukumar y col., 2014). En la saliva de las búfalas se han identificado 37 proteínas específicas para la fase de estro, que pueden considerarse una herramienta ideal para predecir el momento de la inseminación en búfalos.
2.1.7 Estro silencioso
exposición del comportamiento neural de signos de estro, que interfiere con la estimulación de la LH (Das y col., 2013).
2.1.8 Ovulación
La ovulación ocurre en promedio a las 21,41 ± 4,56 horas después del final del estro (Jacomini y col., 1991), Baruselli y col., (1997) reporta que el intervalo entre el estro y la ovulación es en promedio 17 horas después de la aceptación de la monta y en general, ocurren durante la noche, presentando un intervalo estro ovulación en promedio de 34 horas con variaciones en períodos de 24 a 48 horas (Perera, 2008). Mohan y col., (2010) reportan que la ovulación ocurrió después de 13,4 ± 1,0 h luego del pico de estrógenos. Warriach y col., (2008) al trabajar con hembras de raza Nili-Ravi, demostraron con suficiente evidencia que el tiempo en que se presenta la ovulación es de 30 horas después de iniciado el estro.
2.1.9 Desarrollo del cuerpo lúteo
En el desarrollo del CL se evidencia la importancia del colágeno en la formación y regulación de su crecimiento; el colágeno en tejidos lúteos constituye entre el 0,9 % al 1,2 % del peso en fresco; el CL maduro entre los días 11-17 del ciclo estral de la búfala, contiene la más elevada cantidad de colágeno y la síntesis / secreción de éste es más rápido en la mitad de la fase lútea y está asociada con el rápido crecimiento del tejido lúteo (Jaglan y col., 2010).
2.2 INDICADORES CELULARES
En el desarrollo y atresia folicular del CL de la vaca bufalina, están implicadas diversas citoquinas, factores angiogénicos y linfogénicos, como el VEGF, por lo mismo, se ha reportado la coexpresión de diferentes isoformas de VEGF y sus receptores, exponiendo evidencia que contribuyen a una extensa proliferación capilar asociada con el aumento de tamaño en la desviación y maduración del folículo preovulatorio como del CL; el crecimiento asociado a VEGF evidencia una correlación positiva con las concentraciones de P4. Los resultados son consistentes con la hipótesis de que el VEGF tiene un papel esencial en el crecimiento folicular y la maduración del folículo ovulatorio, como del crecimiento y secreción de P4, tal vez, debido a la mayor cantidad de nutrientes, hormonas y otros metabolitos aportados por vía sanguínea (Feranil y col., 2005; Papa y col., 2007; Babitha y col., 2013; Ali y col., 2014).
(FP) y su crecimiento se regula en cultivo; los factores de crecimiento y β mercaptoetanol (un antioxidante) en asociación sinérgica, también mejoran el crecimiento y la supervivencia de los FP en los búfalos y regulan su crecimiento; el factor de crecimiento de fibroblastos, IGF-I y β mercaptoetanol en asociación sinérgica podrían mejorar, a largo plazo, el sistema de cultivo de FP bufalinos (Gupta y Nandi, 2010).
La Grelina es un ligando endógeno del receptor de la hormona del crecimiento, está involucrado en el desarrollo embrionario normal y se ha asociado con la fertilidad en la búfala. Además, está presente en todas las etapas del ciclo estral y es posible que actúe de forma autocrina en el endometrio; la P4, puede ser responsable de la variación en la expresión de Grelina durante las diferentes fases del ciclo estral indicando regulación de las funciones del endometrio durante el ciclo estral (Kandasamy y col., 2013). Se ha descrito el receptor de Grelina en el CL de búfalas durante la fase lútea y un efecto inhibitorio sobre la producción de P4, estas observaciones sugieren un posible papel regulador de la Grelina en el desarrollo función y regresión del CL en el ciclo estral de esta especie (Gupta y col., 2014).
de folículos antrales y el mayor porcentaje de folículos atrésicos en las búfalas (Baldrighi y col., 2014).
La concentración sérica de colesterol HDL y Fosfatasa Alcalina (ALP) son, de modo significativo, más altos en la fase lútea que en la fase folicular. Se observaron correlaciones significativas entre colesterol, sodio y potasio en el líquido folicular y la calidad y el número de ovocitos (Baki y col., 2013).
La Leptina influencia la expresión de factores angiogénicos en el cuerpo lúteo, puede actuar por acción de sus receptores favoreciendo la formación de vasos sanguíneos y la remodelación de la matriz extracelular que confirman efectos paracrinos y autocrinos sobre la esteroidogénesis para favorecer la supervivencia y desarrollo del CL (Kumar y col., 2012).
2.3 ESTACIONALIDAD REPRODUCTIVA
que la mayor frecuencia de partos se registra en el período julio a octubre, siendo septiembre el mes donde se alcanza el mayor número de partos, el mismo patrón reproductivo se ha reportado en la Provincia de Granma – Cuba (Martínez y col., 2009). Al tomar como referencia los meses en donde se evidenció la estacionalidad de la parición, se realizó la retrocuenta para establecer el período de concepción, que se presenta en los meses de septiembre a diciembre y a su vez, se manifiesta la mayor concepción en el mes de noviembre. La importancia de la duración del día en la estacionalidad reproductiva, está muy bien demostrada, donde el fotoperiodo actúa a
través de una señal de la glándula pineal (la Melatonina) desinhibiendo la secreción activa de hormona LH tónica, influencia observada cuando aumenta la longitud del día, dado que la melatonina es sintetizada durante los períodos de oscuridad y su acción es mediada por receptores a nivel del cerebro y han sido asociados a la actividad reproductiva durante los períodos influenciados por el fotoperiodo y pueden ser utilizados como marcadores genéticos para identificar búfalos que tienen predisposición a la estacionalidad reproductiva (Carcangiu y col., 2011; Di Francesco y col., 2012a; Luridiana y col., 2012). La relación entre la alta temperatura ambiental, el incremento de la temperatura rectal en el animal y el subsecuente impacto sobre la alimentación y el balance energético, son efectos conocidos del estrés térmico sobre la reproducción y la producción del búfalo de agua (Marai y Haeeb, 2010).
La estacionalidad reproductiva en el butoro ha sido abordada por diferentes estudios con disimiles resultados. La morfometría testicular y de las glándulas accesorias en el búfalo se reduce con moderación durante los meses de verano. Aunque, el fotoperíodo no aqueja los butoros ya que la calidad espermática no es afectada a lo largo de año, lo que indica que hay una tolerancia a los cambios medioambientales, por el contrario se reporta que algunos parámetros del eyaculado pueden estar alterados durante la temporada de verano, de manera particular, la velocidad de progresión de los espermatozoides siendo menor en verano que cuando se compara con las otras estaciones y con un incremento del daño en el ADN del esperma en el período de verano. (Koonjaenak y col., 2007a; Koonjaenak y col., 2007b; Arrighi y col., 2010; Khawaskar y col., 2012; Ibrahim y col., 2013).
sucede en condiciones mediterráneas como Italia donde ocurre durante la primavera y de manera informal se llama anestro de primavera. La incidencia de este anestro oscila entre el 36,6% y el 59,6% (Das y Khan, 2010; Nam, 2010).
2.4 MORTALIDAD EMBRIONARIA
mitocondrial (mtDNA) y genes relacionados con el metabolismo mitocondrial (PPARGC1A, TFAM e MT-CO1), con la apoptosis (BAX, BCL2), el metabolismo de la glucosa (SLC2A1), el estrés oxidativo (SOD1 e GPX1), el estrés térmico (HSP90AB1 e HSPA1A) y la competencia del desarrollo del oocito (BMP15 e GDF9), que impide el desarrollo apropiado del oocito disminuyendo la fertilidad (Di Francesco y col., 2012b; Macabelli y col., 2011; Macabelli y col., 2014).
2.5 SINCRONIZACIÓN DEL ESTRO Y DE LA OVULACIÓN
La manipulación del desarrollo folicular y el control de la ovulación, posibilitaron la inseminación artificial en tiempo fijo (IATF), sin necesidad de la detección del comportamiento del estro (Abdullah y col., 2001; De Araujo-Berber y col., 2002; Sartori y col., 2009). El desarrollo de ovulación e IA ponen a disposición de los productores de ganado, herramientas suficientes y eficaces para la captura selectiva de características genéticas, con su respectiva implicación económica.
El protocolo de Ovsynch consiste en una aplicación intramuscular de GnRH independiente del día del ciclo estral en que se halle la hembra, que da lugar a la ovulación del FD presente y a iniciar o coincidir con el comienzo de una nueva onda de crecimiento folicular, El suministro de PGF2α se realiza siete días después de la dosificación inicial de GnRH, que conduce a la regresión del CL presente, se aplica una segunda dosis de GnRH, 24 a 48 horas después de la dosis de PGF2α. Como los folículos
realizados por De Araujo-Berber y col., (2002) obtienen buenos resultados en búfalas con un programa Ovsynch, con tasas de ovulación después de la primera (86,6%) y segunda (93,3%,) dosis de GnRH y con una tasa de preñez de 56,1% para hembras lactantes. Rensis y col., (2005) utilizaron un protocolo de Ovsynch en búfalas con inseminación artificial 16 a 20 horas después de la segunda dosificación de GnRH y reportan tasas de concepción de 37,5% para animales con ciclicidad y 4,7% para animales en anestro; aquellos, que presentaron folículos grandes el día de la administración de GnRH, obtuvieron tasas mayores de concepción, demostrando que este tipo de protocolo es afectado por algunas variables tales como edad, intervalo posparto y estado de desarrollo folicular.
Roy y Prakash (2009) agregaron Quinagolida Hidrocloruro (Norprolac®, inhibidor de la prolactina) aceptando que las altas concentraciones de prolactina contribuye a la baja fertilidad de las novillas bufalinas durante el verano porque, disminuyen las concentraciones de estrógenos totales, 17β- estradiol y P4, el tratamiento muestra un potencial para mejorar la fertilidad durante el verano. En búfalas, Ovsynch + CIDR muestra una baja en la secreción de la señal luteolitica, PGF2α, el día 15 posovulación y
Presynch – Ovsynch (P-OV) es una modificación al protocolo original en la cual se administra dos dosis de PGF2α, con 14 días de intervalo una de otra antes de iniciar el protocolo Ovsynch, este protocolo de sincronización es efectivo para sincronizar la ovulación pero no muestra diferencias al ser comparado con el protocolo Ovsynch solo, lo que no justifica la labor previa en la pre sincronización (Orepaza y col., 2010). Karen y Darwish, (2010) realizaron un estudio para evaluar la eficiencia de un protocolo Ovsynch en búfalas egipcias (21 vacas y 8 novillas) con ciclos estrales normales y con anestro; donde después de la primera dosis de GnRH 46% (5 de 11) de las vacas con ciclos normales y 50% (5 de 10) de las búfalas en anestro, 40 % (2 de 5) de las novillas con ciclos normales y el 33% (1 de 3) de las novillas en anestro, respondieron a la ovulación o luteinización de los folículos. De la misma manera, observaron que el diámetro promedio de los folículos de los animales (vacas y novillas) que respondieron al protocolo de sincronización, fue mayor de manera significativa, comparado con los animales que no respondieron (9,7 ± 0, 4 mm vs. 6,7 ± 0,6 mm). Después de la segunda dosis de GnRH, 81% (9 de 11) de los animales con ciclos estrales regulares y el 60 % (6 de 10) de los animales con anestro ovularon; por lo que, concluyeron de esta forma que, el protocolo Ovsynch puede ser utilizado con eficacia en la sincronización de búfalas y novillas, tanto con ciclos estrales normales como en animales durante el anestro.
Heatsynch y Estradoublesynch son dos protocolos que modifican también al protocolo original Ovsynch, en el primero se administra al octavo día de iniciado el programa, benzoato de estradiol y en el segundo se sigue el protocolo Heatsynch mas la aplicación de PGF2α, dos días antes del comienzo del tratamiento. Cuando se compara ambos protocolos en búfalas cíclicas y con anestro, el Estradoublesynch demuestra que es más eficiente en la sincronización de la ovulación, después de las aplicaciones de GnRH y Benzoato de estradiol, obteniendo tasas de preñez más elevadas (62%) que las logradas por Heatsynch. De igual manera, el protocolo Estradoublesynch genera folículos ovulatorios de mayor tamaño al tiempo de la administración de Benzoato de estradiol, lo que aumenta las tasas de fertilidad comparado con el protocolo Heatsynch. Por lo tanto, el protocolo Estradoublesynch puede ser una herramienta más útil y económica que el Doublesynch y Heatsynch para la sincronización de la ovulación e inseminación artificial a tiempo fijo en búfalos (Mirmahmoudi y col., 2014a; Mirmahmoudi y col., 2014b).
Bartlomeu y col., (1999) utilizaron dos protocolos hormonales con implantes, donde, en un grupo emplearon Progesterona al 10% que contenía 1,9 mg de P4 durante 9 días, con una dosis de 1 mg de benzoato de estradiol en el momento de inserción del implante y prostaglandina (15 mg de Lupostrinol), a la retirada del implante. Otro grupo se trató por 9 días, con aplicaciones de 5 mg de valerato de estradiol y 3 mg de Norgestomet (acetato 11β-metil-17-hidroxi-19-norprogesterona) vía intramuscular en el momento de inserción
del implante, al finalizar el estudio en ambos grupos los autores reportan que para búfalos, no hubo eficiencia luteínica del valerato de estradiol.
(Lecirelina) en el día 0 (50 µg I.M) y en el día 9 (25 µg I.M), además, de una aplicación de prostaglandina (d-cloprostenol, 150 µg I.M) en el día 7. Adicionalmente, los animales del segundo grupo fueron suplementados con 60 mg de medroxiprogesterona (análogo de progesterona) entre los días 0 y 7 por aplicación de dispositivo intravaginal. La inseminación artificial fue realizada a todas las búfalas entre 16 y 20 horas después de la segunda aplicación de GnRH, independiente de la presencia de signos de estro. Los autores determinaron que la medroxiprogesterona aumenta el número de folículos pequeños y disminuye los folículos grandes retardando el crecimiento de los folículos con diámetro mayor a 5,0 mm entre los días 0 y 7. El retiro de la progesterona aceleró el crecimiento folicular del día 7 al día 9, haciendo que el diámetro de los FD aumente y supere el de los animales no tratados con progesterona. La medroxiprogesterona adicional al protocolo Ovsynch, contribuye a la ovulación de folículos mayores, consecuente con la formación de cuerpos lúteos de mayor tejido luteínico y más eficientes en el mantenimiento de la gestación.
Chaikhun y col., (2010) demostraron que es posible utilizar IATF después de la sincronización con el protocolo Ovsynch en búfalas de pantano y que la concepción y tasas de preñez son similares a las de inseminación artificial durante el estro natural, para búfalos del Mediterráneo y de raza Murrah.
Malik y col., (2011), evaluaron el efecto del implante auricular Norgestomet (3 mg) e inyección intramuscular de Norgestomet (3 mg más valerato de estradiol 5 mg) sobre la ciclicidad ovárica y la respuesta a la fertilidad en 15 búfalos durante los 90 días antes y 120 días después del período de posparto, encontrando que las concentraciones de P4 generaron la regresión de los folículos dominantes visibles hasta el día 9 de tratamiento, después con la administración de eCG (500 UI), el folículo dominante creció con rapidez hasta el día 12, cuando ocurrió la ovulación con un tamaño promedio de 12,05 ± 0,36 mm. Debido a esto, el 93.3% de los animales tratados (14/15) ovularon, y de los 14 observados por medio de ultrasonografía se dio la formación de un cuerpo lúteo después de 10 días. Las tasas de concepción en los estros inducidos de manera general hasta la tercera inseminación fueron, a saber, de 66.7% (10/15) y 86.7% (13/15). En 13 animales preñados, el promedio de inseminaciones por concepción fue de 1,31. Igualmente Malik y col., (2010) indicaron que el tratamiento Crestar (Norgestomet, 11β-methyl-17-hydroxy-19-norprogesterone acetate), suplementado con valerato de estradiol y eCG, es más eficiente en la inducción del estro y respuesta a la fertilidad comparado con el protocolo Ovsynch en búfalas Murrah en anestro posparto.
(2014), evaluaron diferentes concentraciones de P4 liberados por dispositivos intravaginales usados y nuevos durante la sincronización de la ovulación para observar el efecto sobre el desarrollo folicular y la respuesta a la preñez en búfalas en temporada de anestro. En el estudio se observa que no existe diferencia entre los tratamientos experimentales puesto que el tamaño del folículo dominante y las tasas de concepción no variaron entre ellos, a su vez, los perfiles plasmáticos de P4 si fueron distintos, no obstante, los niveles bajos de la hormona liberados por los dispositivos usados permitieron mantener un adecuado control sobre el desarrollo folicular ovárico y sin efectos indeseables sobre la respuesta a la preñez. De igual manera Neglia y col., (2008), realizaron un ensayo en búfalas sincronizadas por el protocolo Ovsynch-IATF, donde utilizaron en un grupo Cloprostenol sodico (0.524 mg) vía intravenosa (I.V) e intramuscular (I.M), y a grupos control aplicándoles solución salina fisiológica (2 ml vía I.M e I.V), observándose que los grupos tratados con PF2α en el día del estro (animales que consideraron en calor) y en el día de la IATF, obtuvieron concentraciones plasmáticas elevadas de P4 en el día 11 (Día 0: día del estro), y cuerpos lúteos de mayor tamaño, a su vez, tasas de preñez más elevadas (46.7% vs. 30.7%) comparado a los grupos control en los días 26 o 45.
BIBLIOGRAFÍA
Abdalla EB. Improving the reproductive performance of Egyptian buffalo cows by changing the management system. Anim Reprod Sci. 2003; 75(1-2):1-8.
Abdoon AS, Gabler C, Holder C, Kandil OM, Einspanier R. Seasonal variations in developmental competence and relative abundance of gene transcripts in buffalo (Bubalus bubalis) oocytes. Theriogenology. 2014; 82(8):1055-1067.
Abdullah P, Williamson NB, Parkinson TJ, Fathalla M. Comparison of estrus synchronization programmes in dairy cattle using oestradiol benzoate, short-acting progesterone and cloprostenol, or buserelin and cloprostenol. N Z Vet J. 2001; 49(5):201-210.
Ali A, Abdel-Razek AK, Abdel-Ghaffar S, Glatzel PS. Ovarian follicular dynamics in buffalo cows (Bubalus bubalis). Reprod Domest Anim. 2003; 38(3):214-218. Ali A, Fahmy S. Ovarian dynamics and milk progesterone concentrations in cycling and
non-cycling buffalo-cows (Bubalus bubalis) during Ovsynch program. Theriogenology. 2007; 68(1):23-28.
Ali I, Chouhan VS, Dangi SS, et al. Expression and localization of locally produced growth factors regulating lymphangiogenesis during different stages of the estrous cycle in corpus luteum of buffalo (Bubalus bubalis). Theriogenology. 2014; 81(3):428-436.
Angulo A, Muñoz L, González M, Álvarez LD. Sincronización de la ovulación e inseminación artificial en búfalas a tiempo fijo. Revista Mvz Córdoba. 2004; 9(2):444-450.
Arrighi S, Bosi G, Groppetti D, Cremonesi F. Morpho and Histometric Evaluations on the Testis and Epididymis in Buffalo Bulls During the Different Reproductive Seasons. Open Anat J. 2010; 2:29-33.
Atanasov A, Yotov S, Antonov A, Kolev P. Induction of oestrus and conception rates in Bulgarian Murrah buffaloes after fixedtime artificial insemination (A preliminary study). Bulgarian J Vet Med. 2011; 14(3):165-170, p.165-170.pdf. Consultado: Octubre 10 de 2014 en: http://tru.uni-sz.bg/bjvm/BJVM.
Awasthi MK, Kavani FS, Siddiquee GM, Sarvaiya NP, Derashri HJ. Is slow follicular growth the cause of silent estrus in water buffaloes? Anim Reprod Sci. 2007; 99(3-4):258-268.
Awasthi MK, Khare A, Kavani FS, Siddiquee GM, Panchal MT, Shah RR. Is one-wave follicular growth during the estrous cycle a usual phenomenon in water buffaloes (Bubalus bubalis). Anim Reprod Sci. 2006; 92(3-4):241-253.
Ayen E, Hasanzadeh S, Tabatabaei S. Defense cells profile of cervical mucous during follicular and luteal phases of estrus cycle in river buffalo. Vet Res Forum. 2012;
3(1):45-48. Consultado: Octubre 8 de 2014 en:
http://vrf.iranjournals.ir/?_action=articleInfo&article=1563.
Azawi OI, Ali AJ, Noaman UT. A study on the ovarian follicular dynamic in Iraqi Northern Buffaloes. Trop Anim. Health Prod. 2009; 41:79–83.
Babitha V, Panda RP, Yadav VP, et al. Amount of mRNA and localization of vascular endothelial growth factor and its receptors in the ovarian follicle during estrous cycle of water buffalo (Bubalus bubalis). Anim Reprod Sci. 2013; 137(3-4):163-176.
Baki Acar D, Birdane MK, Dogan N, Gurler H. Effect of the stage of estrous cycle on follicular population, oocyte yield and quality, and biochemical composition of serum and follicular fluid in Anatolian water buffalo. Anim Reprod Sci. 2013; 137(1-2):8-14.
Baldrighi J, Sá Filho MF, Batista EO, et al. Anti-Mullerian Hormone Concentration and Antral Ovarian Follicle Population in Murrah Heifers Compared to Holstein and Gyr Kept Under the Same Management. Reprod Domest Anim. 2014; 49(6):1015-1020.
Banu TA, Shamsuddin M, Bhattacharjee J, Islam MF, Khan SI, Ahmed JU. Milk progesterone enzyme-linked immunosorbent assay as a tool to investigate ovarian cyclicity of water buffaloes in relation to body condition score and milk production. Acta Vet Scand. 2012; 54(1):30.
Barile VL, Terzano GM, Pacelli C, Todini L, Malfatti A, Barbato O. LH peak and ovulation after two different estrus synchronization treatments in buffalo cows in the daylight-lenghthening period. Theriogenology. 2015; 84: 286–293.
Barile VL. Improving reproductive efficiency in female buffaloes. Livest Prod Sci. 2005; 92(3):183-194.
Barkawi AH, Hafez YM, Ibrahim SA, Ashour G, El-Asheeri AK, Ghanem N. Characteristics of ovarian follicular dynamics throughout the estrous cycle of Egyptian buffaloes. Anim Reprod Sci. 2009; 110(3-4):326-334.
Barman P, Yadav MC, Kumar H, Meur SK, Ghosh SK. Gas chromatographic-mass spectrometric analysis of chemical volatiles in buffalo (Bubalus bubalis) urine. Theriogenology. 2013; 80(6):654-658.
Bartlomeu CC, Del Rei JA, Madureira EH, Carvalho NAT, Barnabe RC, Baruselli PS. sincronizaçao do ciclo estral e da ovulaçao com utilizaçao de CIRD e CRESTAR em bubalinos. Arquivo da Faculdade de Veterinaría UFRGS, 1999; 27(1) supl:208.
Baruselli P, Madureira E, Barnabe V, Barnabe R, Visitin J, Oliveira C, Amaral R.Dinâmica folicular em búfalas submetidas á sincronizaçao da ovulaçao para artificial em tempo fixo. Arquivos da Faculdade de Veterinaria UFRGC. 1999; 27(1) Supl:210.
Baruselli P, Mucciolo R, Visintin J,et al Ovarian follicular dynamics during the estrous cycle in buffalo (Bubalus bubalis). Theriogenology. 1997; (97):1531-1547.
Baruselli P, Soares J, Gimenes L. Control of Buffalo Follicular Dynamics for Artificial Insemination, Superovulation and In Vitro Embryo Production. Buffalo Bulletin. 2013; 32(1):160-176. Consultado: Octubre 8 de 2014 en: http://ibic.lib.ku.ac.th/e-bulletin/IBBUSI201301013.pdf.
Baruselli PS, Madureira, EH, Barnabe VH, Barnabe RC, De Araujo Berber RC. Evaluation of synchronization of ovulation for fixed timed insemination in buffalo (Bubalus bubalis). Brazilian Journal of Veterinary Research and Animal Science. 2003; 40: 431-442.
Brito LFC, Satrapa R, Marson EP, Kastelic JP. Efficacy of PGF(2alpha) to synchronize estrus in water buffalo cows (Bubalus bubalis) is dependent upon plasma progesterone concentration, corpus luteum size and ovarian follicular status before treatment. Anim Reprod Sci. 2002; 73(1-2):23-35.
Campanile G, Baruselli PS, Neglia G, et al. Ovarian function in the buffalo and implications for embryo development and assisted reproduction. Anim Reprod Sci. 2010; 121(1-2):1-11.
Campanile G, Neglia G, Gasparrini B. Embryonic mortality in buffaloes synchronized and mated by AI during the seasonal decline in reproductive function. Theriogenology. 2005; 63:2334-2340.
Campanile G, Vecchio D, Neglia G, et al. Effect of season, late embryonic mortality and progesterone production on pregnancy rates in pluriparous buffaloes (Bubalus bubalis) after artificial insemination with sexed semen. Theriogenology. 2013; 79(4):653-659.
Carcangiu V, Mura MC, Pazzola M, et al. Characterization of the Mediterranean Italian buffaloes melatonin receptor 1A (MTNR1A) gene and its association with reproductive seasonality. Theriogenology. 2011; 76(3):419-426.
Carvalho NAT, Baruselli PS, Zicarelli L, Madureira EH, Visintin JA. D’Occhio MJ. Control of ovulation with a GnRH agonist after superstimulation of follicular growth in buffalo: fertilization and embryo recovery. Theriogenology. 2002; 58(9):1641-1650.
Carvalho NAT, Soares J.G, Souza D.C, Vannucci FS, Amaral R, Maio JRG, Sales JNS, Sá Filho MF, Baruselli P.S. Different circulating progesterone concentrations during synchronization of ovulation protocol did not affect ovarian follicular and pregnancy responses in seasonal anestrous buffalo cows. Theriogenology. 2014;81: 490–495
Chaikhun T, Tharasanit T, Rattanatep J, et al. Fertility of swamp buffalo following the synchronization of ovulation by the sequential administration of GnRH and PGF₂alpha combined with fixed-timed artificial insemination. Theriogenology. 2010; 74(8):1371-1376.
Chouhan VS, Dangi SS, Vazhoor B, et al. Stimulatory effect of luteinizing hormone, insulin-like growth factor-1, and epidermal growth factor on progesterone secretion and viability of cultured bubaline luteal cells. Theriogenology. 2014; 82(9):1212-1223.
Chouhan VS, Panda RP, Yadav VP, et al. Expression and localization of vascular endothelial growth factor and its receptors in the corpus luteum during oestrous cycle in water buffaloes (Bubalus bubalis). Reprod Domest Anim. 2013; 48(5):810-818.
Crudeli G, Pellerano G. Efecto de diferentes variables sobre la preñez en búfalas sometidas a sincronización del celo e inseminación artificial a tiempo fijo. Rev. Vet. 2008; 19(1):14-17.
Dadarwal D, Ghuman SPS, Honparkhe M, Singh J. Synchronization of ovulation and subsequent fertility in buffaloes following PGF2α- PGF2α protocol, with or without GnRH. Indian J Anim Sci. 2009; 79 (9): 861–865.
Danell B. 1987. Oestrous behavior, ovarian morphology and cyclical variation in follicular system and endocrine pattern in water buffalo heifers. (PhD Thesis). Department of Obstetrics and Gynecology, Faculty of Veterinary Medicine, Swedish University of Agricultural Sciences, Uppsala, Sweden. 124pp.
Das GK, Khan FA. Summer anoestrus in buffalo--a review. Reprod Domest Anim. 2010; 45(6): e483-e494.
De Araujo Berber RC, Madureira EH, Baruselli PS. Comparison of two Ovsynch protocols (GnRH versus LH) for fixed timed insemination in buffalo (Bubalus bubalis). Theriogenology. 2002; 57(5):1421-1430.
Derar R, Hussein H, Fahmy S. The effect of parity on the efficacy of an ovulation synchronization (Ovsynch) protocol in buffalo (Bubalus bubalis). Anim Reprod. 2012; 20(88):52-60.
De Rensis F, Lopez-Gatius F. Protocols for synchronizing estrus and ovulation in buffalo (Bubalus bubalis): A review. Theriogenology. 2007; 67:209–216.
Di Francesco S, Neglia G, Vecchio D, et al. Influence of season on corpus luteum structure and function and AI outcome in the Italian Mediterranean buffalo (Bubalus bubalis). Theriogenology. 2012a; 78(8):1839-1845.
Di Francesco S, Novoa MVS, Vecchio D, et al. Ovum pick-up and in vitro embryo production (OPU-IVEP) in Mediterranean Italian buffalo performed in different seasons. Theriogenology. 2012b; 77(1):148-154.
Drost M. Advanced reproductive technology in the wáter buffalo. Theriogenology. 2007; 68: 450–453.
Dubey PK, Tripathi V, Singh RP, Sharma GT. Influence of nitric oxide on in vitro growth, survival, steroidogenesis, and apoptosis of follicle stimulating hormone stimulated buffalo (Bubalus bubalis ) preantral follicles. J Vet Sci. 2011; 12(3):257-265.
El-Shahat KH, Kandil M. Antioxidant capacity of follicular fluid in relation to follicular size and stage of estrous cycle in buffaloes. Theriogenology. 2012; 77(8):1513-1518.
Feranil JB, Isobe N, Nakao T. Cell Proliferation in the Atretic Follicles of Buffalo and Cattle Ovary. Reprod Domest Anim 2004; 39(6):405-409.
Feranil JB, Isobe N, Nakao T. Immunolocalization of von Willebrand factor and vascular endothelial growth factor during follicular atresia in the swamp buffalo ovary. J Reprod Dev. 2005; 51(4):419-426.
Ganguly A, Ganguly I, Meur SK. Multiple molecular forms of inhibin in buffalo (Bubalus bubalis) ovarian follicular fluid. Res Vet Sci. 2010; 89(1):14-19.
García AR, De Souza Nahúm B, Lourenco Junior JB, et al. Associação da medroxiprogesterona ao protocolo Ovsynch para inseminaçao artificial em tempo fixo de búfalas cíclicas (Bubalus bubalis) criadas na Amazonia Oriental. Acta Amazónica. 2008; 38: 369-378.
Ghuman SPS, Honparkhe M, Singh J, Dhami DS, Kumar A, Nazir G, Ahuja C. Fertility response using three estrus synchronization regimens in lactating anestrous buffaloes. Indian J Anim Sci. 2012; 82 (2):162–166.
Gimenes LU, Carvalho NAT, Sá Filho MF, et al. Ultrasonographic and endocrine aspects of follicle deviation, and acquisition of ovulatory capacity in buffalo (Bubalus bubalis) heifers. Anim Reprod Sci. 2011; 123(3-4):175-179.
Gokuldas PP, Yadav MC, Kumar H, Singh G, Mahmood S, Tomar a KS. Resumption of ovarian cyclicity and fertility response in bull-exposed postpartum buffaloes. Anim Reprod Sci. 2010; 121(3-4):236-241.
Gupta M, Dangi SS, Chouhan VS, et al. Expression and localization of ghrelin and its functional receptor in corpus luteum during different stages of estrous cycle and the modulatory role of ghrelin on progesterone production in cultured luteal cells in buffalo. Domest Anim Endocrinol. 2014; 48:21-32.
Hassan F, Khan MS, Rehman MS, Sarwar M, Bhatti S a. Seasonality of calving in Nili-Ravi buffaloes, purebred Sahiwal and crossbred cattle in Pakistan. Ital J Anim Sci. 2010; 6(2):1298-1301.
Ibrahim N, Al-sahaf M, Alwan A. Reproductive Activity of Mature Iraqi Bull Buffaloes: Testes Dimensions and Histological Picture. Int J Anim Veter Adv. 2013; 5(1):34-37.
ICA. Censo Pecuario Nacional- 2015. Consultado: Febrero 20 de 2015 en:
http://www.ica.gov.co/getdoc/8232c0e5-be97-42bd-b07b-9cdbfb07fcac/Censos-2008.aspx.
Ireland JJ, Mihm M, Austin E, Diskin MG, Roche JF. Historical Perspective of Turnover of Dominant Follicles during the Bovine Estrous Cycle: Key Concepts, Studies, Advancements, and Terms. J Dairy Sci. 2000; 83: 1648-1658.
Jacomini J, Chow L, Andrade V, Melo P. Detecçao do estro e inseminaçao artificial em búfala (Bubalus bubalis). Revista Brasilera de Reproducao Animal. 1991; 15(1-2):19-24.
Jaglan P, Das GK, Kumar BVS, Kumar R, Khan F a, Meur SK. Cyclical changes in collagen concentration in relation to growth and development of buffalo corpus luteum. Vet Res Commun. 2010; 34(6):511-518.
Jain A, Baviskar PS, Kandasamy S, et al. Interferon stimulated gene 15 (ISG15): molecular characterization and expression profile in endometrium of buffalo (Bubalus bubalis). Anim Reprod Sci. 2012; 133(3-4):159-168.
Jan MH, Das GK, Khan F a., et al. Evaluation of follicular oxidant-antioxidant balance and oxidative damage during reproductive acyclicity in water buffalo (Bubalus bubalis) Asian Pacific J Reprod. 2014; 3(1):35-40.
Jones AL, Lamb GC. Nutrition synchronization and management of beef embryo transfer recipients. Theriogenology. 2008; 69(1):107-115.
Kandasamy S, Jain A, Baviskar P, et al. Molecular characterization and expression profile of ghrelin gene during different reproductive phases in buffalo (Bubalus bubalis). Domest Anim Endocrinol. 2013; 45(2):55-63.
Kandasamy S, Jain A, Kumar R, Agarwal SK, Joshi P, Mitra A. Molecular characterization and expression profile of uterine serpin (SERPINA14) during different reproductive phases in water buffalo (Bubalus bubalis). Anim Reprod Sci. 2010; 122(1-2):133-141.
Kandiel MMM, El-Naggar RAM, Abdel-Ghaffar AE, Sosa GAM, Abou El-Roos NA. Interrelationship between milk constituents, serum oestradiol and vaginal mucus indicators of oestrus in Egyptian buffaloes. J Anim Physiol Anim Nutr (Berl). 2014; 98(1):197-200.
Karen AM, Darwish SA. Efficacy of Ovsynch protocol in cyclic and acyclic Egyptian buffaloes in summer. Anim Reprod Sci. 2010; 119(1-2):17-23.
Karthikeyan K, Muniasamy S, SankarGanesh D, Achiraman S, Ramesh Saravanakumar V, Archunan G. Faecal chemical cues in water buffalo that facilitate estrus detection. Anim Reprod Sci. 2013; 138(3-4):163-167.
Khan FA, Das GK. Follicular fluid nitric oxide and ascorbic acid concentrations in relation to follicle size, functional status and stage of estrous cycle in buffalo. Anim Reprod Sci. 2011; 125(1-4):62-68.
Koonjaenak S, Chanatinart V, Aiumlamai S, Pinyopumimintr T, Rodriguez-Martinez H. Seasonal variation in semen quality of swamp buffalo bulls (Bubalus bubalis) in Thailand. Asian J Androl. 2007b; 9(1):92-101.
Koonjaenak S, Pongpeng P, Wirojwuthikul S, Johannisson A, Kunavongkrit A, Rodriguez-Martinez H. Seasonality affects post-thaw plasma membrane intactness and sperm velocities in spermatozoa from Thai AI swamp buffaloes (Bubalus bubalis). Theriogenology. 2007a; 67(9):1424-1435.
Kumar A, Solanki VS, Jindal SK, Tripathi VN, Jain GC. Oocyte retrieval and histological studies of follicular population in buffalo ovaries. Anim Reprod Sci. 1997; 47(3):189-195.
Kumar L, Panda RP, Hyder I, et al. Expression of leptin and its receptor in corpus luteum during estrous cycle in buffalo (Bubalus bubalis). Anim Reprod Sci. 2012; 135(1-4):8-17.
Lamb CG, Dahlen CR, Larson JE, Marquezini G, Stevenson S. Control of the estrous cycle to improve fertility for Fixed – time artificial insemination in beef cattle: A review. J Anim Sci. 2010 88(13): E181–192.
Luridiana S, Mura MC, Pazzola M, et al. Association between melatonin receptor 1A (MTNR1A) gene polymorphism and the reproductive performance of Mediterranean Italian buffaloes. Reprod Fertil Dev. 2012; 24(7):983-987.
Macabelli CH, Ferreira RM, Gimenes LU, et al. Reference gene selection for gene expression analysis of oocytes collected from dairy cattle and buffaloes during winter and summer. PLoS One. 2014; 9(3):e93287.
Macabelli CH, Gimenes LU, Barrilao AG, Chiaratti MR. Avaliação molecular da competência de desenvolvimento de oócitos de búfalas aspirados in vivo durante o inverno e o verão. 20º Simpósio Internacional de Iniciação Científica da USP. Universidade de São Paulo. 2011
Madan ML, Das SK, Palta P. Application of reproductive technology to buffaloes. Anim Reprod Sci. 1996; 42: 299-306.
Malik RK, Singh P, Sharma RK, Singh I, Phulia SK, Tuli RK. Efficacy of Norgestomet ear implant for estrus induction on postpartum anestrus Murrah buffaloes (Bubalus bubalis). Indian J Anim Sci. 2011; 81 (7): 687–690.
Malik RK, Singh P, Sharma RK, Singh I, Tuli RK. Estrus and fertility response of postpartum anestrus Murrah buffaloes to Crestar and Ovsynch treatment regimens. Indian J Anim Sci. 2010; 80 (10):982–85.
Manik R, Palta P, Singla S, Sharma V. Folliculogenesis in buffalo (Bubalus bubalis): a review. Reprod Fertil Dev 2002; 14(5):315-325.
Marai IFM, Haeeb AAM. Buffalo’s biological functions as affected by heat stress - A review. Livest Sci. 2010; 127(2-3):89-109.
Martínez A, Ray J, López R. Comportamiento de algunos indicadores productivos y reproductivos del búfalo de río en la provincia Granma. Rev Cuba de ciencias agrícolas. 2009; 43(2):127-130.
Megahed GA, Anwar MM, Wasfy SI, Hammadeh ME. Influence of heat stress on the cortisol and oxidant-antioxidants balance during oestrous phase in buffalo-cows (Bubalus bubalis): thermo-protective role of antioxidant treatment. Reprod Domest Anim. 2008; 43(6):672-677.
Mirmahmoudi R, Prakash B.S. The endocrine changes, the timing of ovulation and the efficacy of the doublesynch protocol in the Murrah buffalo (Bubalus bubalis). Gen Comp Endocrinol. 2012; 177: 153–159.
Estradoublesynch and Heatsynch protocols in Murrah buffalo cows (Bubalus bubalis). Theriogenology. 2014a; 82: 1012–1020.
Mirmahmoudi R, Souri M, Prakash B.S. Endocrine changes, timing of ovulation, ovarian follicular growth and efficacy of a novel protocol (Estradoublesynch) for synchronization of ovulation and timed artificial insemination in Murrah buffaloes (Bubalus bubalis). Theriogenology. 2014b; 81: 237–242.
Mohan K, Prakash BS. Changes in endogenous estrogens and expression of behaviors associated with estrus during the periovulatory period in Heatsynch treated Murrah buffaloes (Bubalus bubalis). Trop Anim Health Prod. 2010; 42(5):947-952.
Mondadori R, Luque M. Ultrastructural and morphometric characterization of buffalo (Bubalus bubalis) ovarian preantral follicles. Anim Reprod Sci 2007; 97(3-4):323-333.
Mondadori RG, Santin TR, Fidelis AG, Porfírio EP, Báo SN. Buffalo (Bubalus bubalis) pre-antral follicle population and ultrastructural characterization of antral follicle oocyte. Reprod Domest Anim. 2010; 45(1):33-37.
Mondal S, Prakash BS, Palta P. Endocrine Aspects of Oestrous Cycle in Buffaloes (Bubalus bubalis): An Overview. Asian-Australasian J Anim Sci. 2006; 20(1):124-131.
Mondal S, Suresh KP, Nandi S. Endocrine Profiles of Oestrous Cycle in Buffalo: A Meta-analysis. Asian-Australasian J Anim Sci. 2009; 23(2):169-174.
Murugavel K, Antoine D, Raju MS, López-Gatius F. The effect of addition of equine chorionic gonadotropin to a progesterone-based estrous synchronization protocol in buffaloes (Bubalus bubalis) under tropical conditions. Theriogenology. 2009; 71(7):1120-1126.
Muthukumar S, Rajkumar R, Rajesh D, et al. Exploration of salivary proteins in buffalo: an approach to find marker proteins for estrus. Faseb J. 2014; 28(11):4700-4709. Nam NH. Characteristics of reproduction of the water buffalo and techiques used to
improve therir reproductive performance. J Sci Dev 2010; 8(1):100-110.
Nazir G, Ghuman SPS, Singh J, et al. Improvement of conception rate in postpartum flaxseed supplemented buffalo with Ovsynch+CIDR protocol. Anim Reprod Sci. 2013; 137(1-2):15-22.
Neglia G, Gasparrini B, Di Palo R, De Rosa C, Zicarelli L, Campanile G. Comparison of pregnancy rates with two estrus synchronization protocols in Italian Mediterranean Buffalo cows. Theriogenology. 2003; 60(1):125-133.
Neglia G, Natale A, Esposito G, Salzillo F, Adinolfi L, Campanile G, Francillo M, Zicarelli L. Effect of prostaglandin F2α at the time of AI on progesterone levels and pregnancy rate in synchronized Italian Mediterranean buffaloes. Theriogenology. 2008; 69: 953–960.
Oropeza AJ, Rojas AF, Velazquez MA, Muro JD, Márquez YC, Vilanova LT. Efficiency of two timed artificial insemination protocols in Murrah buffaloes managed under a semi-intensive system in the tropics. Trop Anim Health Prod. 2010; 42(6):1149-1154.
Pandey, A. Dhaliwalb, S. Ghumana, S. Agarwalc, S. Impact of pre-ovulatory follicle diameter on plasma estradiol, subsequent luteal profiles and conception rate in buffalo (Bubalus bubalis). Anim Reprod Sci. 2011; 123(3-4): 169-174.
Paul V, Prakash B. Efficacy of the Ovsynch protocol for synchronization of ovulation and fixed-time artificial insemination in Murrah buffaloes (Bubalus bubalis). Theriogenology. 2005; 64:1049-1060.
Perera BMAO. Reproduction in domestic buffalo. Reprod Domest Anim. 2008; 43 Suppl 2:200-206.
Perera BMAO. Reproductive cycles of buffalo. Anim Reprod Sci. 2011; 124(3-4):194-199.
Peter AT, Levine H, Drost M, Bergfelt DR. Compilation of classical and contemporary terminology used to describe morphological aspects of ovarian dynamics in cattle. Theriogenology. 2009; 71(9):1343-1357.
Porto-Filho RM, Gimenes LU, Monteiro BM, et al. Detection of estrous behavior in buffalo heifers by radiotelemetry following PGF2α administration during the early or late luteal phase. Anim Reprod Sci. 2014; 144(3-4):90-94.
Presicce GA, Bella A, Terzano GM, De Santis G, Senatore EM. Postpartum ovarian follicular dynamics in primiparous and pluriparous Mediterranean Italian buffaloes (Bubalus bubalis). Theriogenology. 2005; 63(5):1430-1439.
Presicce GA, Senatore EM, De Santis G, et al. Hormonal stimulation and oocyte maturational competence in prepuberal Mediterranean Italian buffaloes (Bubalus bubalis). Theriogenology 2002;57(7):1877-1884.
Rajakoski, E. The ovarian follicular system in sexually mature heifers with special reference to seasonal, cyclical, end left-right variations. Acta Endocrinol. 1960; 52 (suppl.):1- 68.
Rajanarayanan S, Archunan G. Identification of urinary sex pheromones in female buffaloes and their influence on bull reproductive behaviour. Res Vet Sci. 2011; 91(2):301-305.
Rajanarayanan S, Archunan G. Occurrence of flehmen in male buffaloes (Bubalus bubalis) with special reference to estrus. Theriogenology. 2004; 61(5):861-866. Rakesh HB, Singh SK, Sharma GC, et al. Morphological and functional characterization
of corpus luteum during different stages of estrous cycle in buffalo. Indian J Anim Sci. 2013; 83(7):710-712.
Rensis F De, Ronci G, Guarneri P, Xuan B. Conception rate after fixed time insemination following Ovsynch protocol with and without progesterone supplementation in cyclic and non-cyclic Mediterranean Italian buffaloes (Bubalus bubalis). Theriogenology 2005; 63(7):1824-1831.
Rossi P, Vecchio D, Neglia G, Di Palo R, Gasparrini B, DOcchio MJ, Campanile G. Seasonal fluctuations in the response of Italian Mediterranean buffaloes to synchronization of ovulation and timed artificial insemination. Theriogenology. 2014; 82: 132–137.
Roy KS, Prakash BS. Changes in endocrine profiles during Ovsynch and Ovsynch plus norprolac treatment in Murrah buffalo heifers at hot summer season. Trop Anim Health Prod. 2009; 41(4):677-687.
Roy KS, Prakash BS. Seasonal variation and circadian rhythmicity of the prolactin profile during the summer months in repeat-breeding Murrah buffalo heifers. Reprod Fertil Dev. 2007; 19(4):569.
Russo M, Vecchio D, Neglia G, et al. Corpus luteum function and pregnancy outcome in buffaloes during the transition period from breeding to non-breeding season. Reprod Domest Anim. 2010; 45(6):988-991.
– bases protocols in suckled Bos indicus cows. Anim Reprod Sci 2010; 120: 23-30. Sá Filho OG, Meneghetti M. Peres RFG, Lamb GC, Vasconcelos JLM. Fixed – time artificial insemination with estradiol and progesterone for Bos indicus cows II: Strategies and factors affecting fertility. Theriogenology, 2009; (72): 210–218. Sah SK, Nakao TA. Clinical Study of Anestrus Buffaloes in Southern Nepal. J Reprod
Dev. 2010; 56(2):208-211. Sartori R, Baruselli P, Souza A. Recent advances in ovulation synchronization and superovulation in dairy cattle. Anim Reprod. 2009; 6(1): 194.
Scully S, Evans AC, Duffy P, Crowe MA. Characterization of follicle and CL development in beef heifers using high resolution three-dimensional ultrasonography. Theriogenology. 2014; 81(3):407-418.
Sertu S, Voiculescu M, Pop A, Bota A. Research Concerning the Reproduction Seasonality in Carpathian Buffalo. Sci Pap Anim Sci Biotecnologies. 2012;
45(1):253-258. Consultado: Octubre 21 de 2014 en:
http://spasb.ro/index.php/spasb/article/view/580.
Singh J, Nanda AS, Adams GP. The reproductive pattern and efficiency of female buffaloes. Anim Reprod Sci. 2000; 60-61:593-604.
Srivastava SK. Oestrus induction and conception in buffaloes after hormonal treatment during summer. Indian J Anim Sci. 2005; 75 (7):765-768.
Suthar V, Dhami A. Estrus detection methods in buffalo. Vet World. 2010; 3(2):94-96. Consultado: Octubre 8 de 2014 en: http://www.ejmanager.com/mnstemps/2/2-1297701873.pdf?t=1401423552.
Tanaeja M, Ali A, Singh G. Ovarian follicular dynamics in water buffalo. Theriogenology. 1996; 46(1):121-130.
Vecchio D, Neglia G, Di Palo R, et al. Is a delayed treatment with GnRH, HCG or progesterone beneficial for reducing embryonic mortality in buffaloes? Reprod Domest Anim. 2010; 45(4):614-618.
Waheed M. Ovarian activity and hormonal relationships in pregnant buffaloes. Buff Bulletin. 2011; 30(1). Consultado: Octubre 22 de 2014 en: http://ibic.lib.ku.ac.th/e-Bulletin/2011-55.pdf.
Warriach HM, Ahmad N. Follicular waves during the oestrous cycle in Nili-Ravi buffaloes undergoing spontaneous and PGF2alpha-induced luteolysis. Anim Reprod Sci. 2007; 101(3-4):332-337.
Warriach HM, Channa AA, Ahmad N. Effect of oestrus synchronization methods on oestrus behaviour, timing of ovulation and pregnancy rate during the breeding and low breeding seasons in Nili-Ravi buffaloes. Anim Reprod Sci. 2008; 107(1-2):62-67.
Yindee M, Techakumphu M, Lohachit C, et al. Follicular dynamics and oestrous detection in Thai postpartum swamp buffaloes (Bubalus bubalis). Reprod Domest Anim. 2011; 46(1):e91-e96.
Yindee M, Techakumphu M, Lohachit C, Sirivaidyapong S, Na-Chiangmai a., Colenbrander B. Ovarian activity and sexual behavior in the postpartum Swamp Buffalo (Bubalus bubalis). Ital J Anim Sci. 2010; 6(2s):632-635.
Yotov S, Atanasov A, Dineva J. Influence of postpartum luteal activity on the reproductive performance in bulgarian murrah buffaloes. Vet ir Zootech. 2013;
62(84). Consultado Octubre 8 de 2014 en:
CAPÍTULO III
ESTACIONALIDAD REPRODUCTIVA DE LA HEMBRA BUFALINA (Bubalus bubalis)
REPRODUCTIVE SEASONALITY OF FEMALE BUFFALO (Bubalus bubalis)
ESTACIONALIDAD EN BUFALAS
RESUMEN
El objetivo de este estudio fue describir la estacionalidad reproductiva en la búfala en condiciones del trópico bajo en el Magdalena Medio colombiano y su relación con parámetros climáticos. Se evaluaron registros de nacimientos de 30.571 animales desde 1970 a 2013 del Fondo Bufalero del Centro y los datos hidrometeorológicos de la estación Puerto Berrio-Antioquia, Colombia. Para el análisis se utilizó una regresión de Poisson y una correlación de Spearman para determinar la relación entre la época de la concepción (%) y los nacimientos (%) con las siguientes variables hidrometeorológicas: valores mínimos, medios y máximos mensuales de caudales de ríos (m3/seg), valores medios, máximo mensuales de niveles de los ríos (cms), valores totales mensuales de brillo solar (oC), valores mínimos, medios y máximos mensuales de temperatura (oC), valores medios mensuales de humedad relativa (%), valores máximos mensuales de precipitación (mms en 24 horas), valores totales mensuales de precipitación (mms) y el Índice de temperatura y humedad (ITH). El porcentaje de concepción fue mayor durante el primer trimestre del año y la mayor probabilidad de partos se presentó durante el segundo semestre, el menor número de nacimientos se observó en el mes de marzo. El análisis encontró una correlación negativa (p<0,05) entre la concepción y las variables: los valores máximos
Fuentes de financiación: Patrimonio autónomo Fondo Nacional de Financiamiento para la