r-
i .
PRIMERA
HOJA DEL
REPORTE
FINAL
DEL SERVICIO
SOCIAL
<--
J
1.
-
NOMBRE
DELALUMNO:
@-- MATRICULA
< -
J
TELEFONO:
Jk?i%?
TRIMESTRE:
c-,
I-
HORAS
SEMANA.,/
TITULO DEL TRABAJO PI...-.
c.
*- /NOMBREDELTUTOR:
LUGAR DE REALIZACION DEL SERVICIO
SOCIAL.:
FECHA DE TERMINACION:
-
-
FECHA DE INICIO
/FECHA DE ENTREGA:
cz
I. CLAVE
e.
M'
Alejandra Ramíra Iuia774 31 35, 723 64 78 8724u53
Lic. en HidmbioIogia
Cicneiu Bid6gicas y de la S a d
97-0
20 Hrs.
~ ~ _ A R I A C I & N DEL P A T R ~ N DE
PERFOWCI&N INTEGUMENTAL EN" POSTLARVAS DEL CAMAR6NIPenaeus
Giiiamei, (BqPNE 1931) DE LAS REGIONES
M. en C. Carlor
iikrnz
SaraLab. de Biaiogia dc &encar Hidrdúgieu
UAM
-
IztapalapaO2 de Septiembre de 19%
02 de A p f o de 1997 17 de Septiembre de
H.012.96 I
-
DE
PANAMA
Y
MEXICO~Casa
abierta
al
tiempo
I
' -
UNIVERSIDAD AUTONOMA METROPOLITANA
DMSION DE CIENCIAS BIOLOGICS Y DE LA SALUD
SERVICIO SOCIAL
I,.-
-
P"
" Ii"
A QUIEN CORRESPONDA:
Por medio de la presente se hace constar que el (la):
del Departamento de: Hidrobiología
de la División de Ciencias Biológicas y de la Salud asesoro el siguiente Servicio Social:
TITULO:
camarón Peneeus vannamei,(Boone 1931
1
de las regiones de Panamá y México ALUMNO: María Alejandra Ramírez RuizMATRkULA: 8724T453
LICENCIATURA: Yidrobiologia
PERIODO:
Se extiende la presente para los fines que al interesado convengan, en la Ciudad de México, D.F. a diez de Octubre de mil novecientos noventa y siete.
M. en C. Carlos Alvárez Silva
"Variación del patrón de perforación integumental en postlarvas del
Septiembre 2 , 1996 a ---Septiembre 17, 1997
-
A T E N T A M E N T E
M. EN C. ARTURO PRECIADO LOPEZ SECRETARIO ACADÉMICO
r
r"
\ "UNIDAD IZTAPALAPA
Av. Michoacán y La Puriaima lztapalapa 09340 México, D.F. A.P. 55-535 % .
r"'"'
Ilia
UNiVERSlDAD
AUTONOMA
METROPOLITANA
M.
on
C. CARLOS ALVARET SILVAOlVlSlON DE CIENCIAS BIOLOGICAS Y DE
LA
SALUDDEPARTAMENTO OE HlDROBlOLOGlA
Mtkico,
D.
P.,
2de
agostode
1997DR JOSE LUIS ARREDONDO FIGUEROA
PXESlDE"E
DCL
CONSEJO ACADEMIC0 YBIOLÓGICAS Y DE LA SALUD. UAM-IZTAPALAPA
PRESENTE.
Por
mediode la
pnsente comunico a usteú que estoyde
acuerdocon
el contenidodel
iniormefinal
del a l m ode
la carrefade
Hidmbiología RamlnzRuiz
Maria
Alejandra con número de matricula: 87242453, cuyo irabap se titiiló "VNUACl6N DEL PATRÓN DE PERPORAQÓN INTEGUMENTAL EN POSLARVAS DE CAMAXbNPcnrrnu
V
í
(BOONE 1931) DELAS
REGIONES DE P.W& Y MlkXICO? Que tuvo como fechade ierminacidn el dia 02 de agosto
de
1997en
el laboratorio deCuencas
Hihlógica$ UAM-Iztapalapa. DIRECTOR DELA
DMSI~NDE
CIENCIASAgradeciendo de antemo la atencibn que
se siiva
prestar a la presente, quedando a sus ordenes.ATENTAMENTE
M. EN C. CARLOS ALVAREZ SILVA
PROFR
TITULAR "6" TIEMPO COMPLETOGRUPO DE
TRABAJO
BIOLOGU DE CUENCAS HIDROLOGICASUNIDAD IZTAPWPA
Pag.
PRIMERA HOJA DEL REPORTE FINAL
CARTA DE
TERMINACI~N
ML SERWCD
SOCWL
INTRODUCCI~N
...
ANTECEDENTES
...
JUSTlFlCACIdN Y NATURALEZA DEL PROYECTO...
...
*
OBJETIVOS
...
METOWLOGb UTILIZADA
... ...
,
...
,
...
OBJETIVOS Y METAS ALCANZADOS
...
RESULTADOS
Y
DISCUSIONES
...
CONCLUSI~N
...
RECOMENDACIONES
...
LITERATURA CITADA
...
,
...
APÉNDICE
TABLAS
...
FIGURAS
...
1
3
7
8
9
I 1
12
18
19
20
i
El camarón Poria- -i (Boonm,1931), se distribuye a lo largo de la costa este del Océano Pacifico desde Sonora, en el Golfo de California, México, hasta las costas de Tumbes, en Perú.
Esta especie es muy apreciada por los acuicuitores, no solo por sus excelentes condiciones de crecimiento y sobrevivencia, al tolerar amplios rangos de salinidad y temperatura, lo que facilita el manejo de estos organismos; sino adeds, por su demanda y el alto valor en el 4 mercado nacional
e internacional que posee. (Martinez-Córdoba, 19931.
En la identificación de especies del género p.liiriu a nivel de postlarvas se tienen ciertas dificultades, ya que algunos caracteres morfológicos utilizados en la separación de especies, no son permanentes como es el caso de los cromatóforos y la espina antenal, por lo que se tiene que recurrir a otros caracteres; como la longitud del rostro y del pedúnculo ocular; presencia o ausencia de espinas en la carena dorsal del sexto somito abdominal entre otras. (Calderón-Pérez, 1989).
En algunos crustáceos, como es el caso especifico de los copépodos, se presentan patrones de perforación en su tegumento que generalmente resultan de glándulas o ductos, algunos pueden estar asociados con órganos de los sentidos,
o formar órganos integumentales, también están relacionados con el número de espinas y setas, pero en muchos de los casos se desconocen sus funciones.
En los copépodos este patrón de perforación es constante en cada especie, como se ha comprobado en los estudios realizados al respecto, por lo que, se pude utilizar como un caracter más para la identificación de especies e incluso se ha utilizado para separar subespecies. (Koomen, 1992).
Tanto el patrón de perforación integumental como el número y el tipo de perforación, forman un grupo de caracteres que son de gran importancia para
estudios taxonómicos y filogenéticos.
De igual manera, en los camarones del género Penaens, existen patrones de perforación integumental, los cuales pueden ser establecidos como un' caracter
más para su identificación. Aunque hasta ahora no se habían determinado, siendo
éste un motivo para l a realización del presente trabajo.
En el trabajo realizado se llevó a cabo un estudio sobre el patrón de perforación integumental con larvas de camarón de la especie Panrrru -i
reproducidas en cautiverio, en el laboratorio de producción de postlarvas de camarón de La Paz, B.C. Sur, México, en donde se cultivan nauplios obtenidos de
sus propios reproductores y nauplios importados de Panamá, con el fin de determinar dicho patrón de perforación, de donde se obtuvieron interesantes resultados.
Para la presentación de los resultados se anexaron tablas y esquemas, donde se describen los patrones de perforación integumental encontrados en la especie mencionada.
9
ANTECEDENTES
En México, el esfuerzo pequero a que ha sido sometida la captura del camarón, ha llegado a su limite, inclusive lo ha llegado a rebasar, ocasionando un volumen de producción constante en los 70s (Anónimo, 1990a), debido a la gran demanda del producto, lo anterior propició un importante desarrollo en la camaronicultura. En 1985 representó el 7% del total producido, para 1989 ya representaba el 26% con un volumen de 56,500 toneladas y se calcula que para el afio 2000 el 50% del camarón producido a nivel mundial provendrá de granjas de cultivo (Anónimo, 1990b).
Dentro del desarrollo de la acuicultura como actividad productiva
y
rentable, el cultivo de camarón es considerado el de mayor importancia económica. Lo anterior, se debe a la fuerte demanda mundial que tiene dicho crustáceo, principalmente en los Estados Unidos y Japón, cuyas exportaciones se han incrementado anualmente de manera significativa, a un volumen de 21,076 toneladas, con un valor de 263,450 miles de dólares para el año de 1991. Con un promedio de captura anual entre 60 y 65 mil toneladas, colocándonos entre los diez primeros paises productores a nivel mundial. (SEPESCA, 1992).
Una de las condiciones que presenta la camaronicultura y de la cual depende el tamaño de la producción es el abasto de la materia prima denominada postlarva. La mayoría de las granjas obtienen la postlarva del medio natural, razón por la cual el monto de la siembra es imppedecible, azaroso e insuficiente, asi como restringido a una época muy corta del año, provocando en casos extremos la inactividad de las granjas y por último el fracaso de las cooperativas. Por esta razón, se hace imprescindible la producción de las postlarvas a gran escala y de manera controlada en los laboratorios.
(De la Lanza, 1992).
Debido a la importancia de las postlarvas de camarón, es que se pensó eti encontrar un nuevo caracter taxonómico para una mejor identificación de las especies y evitar posibles problemas de tipo genético y de reproducción a la hora de cultivar las especies.
*
Este trabajo se hizo con base a los estudios realizados sobre patrones de perforación integumental en copépodos con fines taxonómicos y filogenéticos.
c.
'-' "'d
El conocimiento de los caracteres de identificación a nivel postlarval del género plluriu, que se agrupan en dos subgéneros: Fdmtrpriumr y
Litopauona, se adquirió gradualmente. Macias-Regalado (19731, encontró que,
las postlarvas del genero lufrntrpuurru desarrollan espínulas en la carena dorsal del sexto somito abdodnal y en la región adrostral, cerca del diente epigástrico, a diferencia de las de Iítop.nrcnu que no las presentan.
Posteriormente Mair (1979), para separar a las especies del género
3.lurru utiliza las siguientes características morfológicas: la coloración de cromatóforos, longitud del rostro y del pedúnculo ocular, dientes dorsales del rastro y espínulas en el sexto segmento abdominal; en 1981 otro estudio del mismo autor, seiíala las características anteriores como las m6s confiables para separar taxonómicamente estos organismos.
Cabrera-Jiménez (1983), implantó como nuevo caracter de valor taxonómico, la espina antenal, que es particularmente Útil cuando existe ambigüedad en la distinción de las dos especies de Farfant.praciriu.
Hasta ahora no hay nada escrito sobre
el
patrón de perforación integumental en postlarvas de camarón de ninguna especie.Es por eso que para la realización de este trabajo, se revisó literatura sobre patrones de perforación en copépodos, ya que tanto estos como las postlarvas de camarón son crustáceos.
Una contribución importante respecto a la importancia de los poros integumentales en copépodos, es el de Fleminger (1973), en el que describe la firma de poros en 17 especies del génaro hcalanru y divide a estas especies en cuatro grupos, de acuerdo a su patrón de poros, tal división corresponde con la derivada de otras característlcas morfológicas de las especies.
Ferrari y Bowman ( 9 9 8 0 ) se basaron en el valor taxonómico de la firma de poros, entre otros caracteres, para elaborar un estudio sistemático sobre
12 especies de copépodos pelágicos de la familia Oithonidae. Se reconocieron patrones de poros integumentales característicos de las especies estudiadas, proponiendo debido a su importancia, llegar a un mejor conocimiento de la
función de las estructuras integumentales en la biología de los aithonidos.
Mauchline (1987) describió el patrón de poros a nivel genérico, de familia y de superfamilia; derivado de un estudio del metasoma de 249 especies representando 89 géneros adscritos a 28 familias de copépodos calanoides. Su estudio mostró que los cambios en el patrón de poros del metasoma, a nivel superfamilia, correspondió con las relaciones filogenéticas
en
boga de las superfamilias de calanoides presentadas por diferentes autores que revisaron el grupo basándose en otros caracteres morfológicos.Hulsemann y Fleminger (1990) r e e x d n a n la topografía del segmento genital en hembras de Pontollin8 .pp. y determinan la distribución de los órganos integumentales del segmento y de otras estructuras epicuticulares, encontrando que el número y arreglo de órganos integumentales difieren entre las especies y delinean una morfoclina de acuerdo a las relaciones filogenéticas sobre las bases de otros caracteres, llevándolos al reconocimiento de cuatro especies de Ponteiiizu
e.
Con esto confirman la utilidad de la firma de poros en investigaciones taxonómicas, filogenéticas yevolutivas en los copépodos.
Considerando la firma de poros en los copépodos, algunos investigadores (Huselman y Fleminger, 1997; 1990) se han restringido al urosoma en ia suposición de que la mayoria de los componentes de las especies residen ahí, (Park y Mauchline, 1994), prueban esa suposición en ocho especies del género Pleuromamma evaluando la firma de poros del cefalosoma, metasoma y urosoma separadamente en cada especie. Los autores reportan que la mayoría de la información específica de la especie está en el urosoma, pero .una porción significativa también reside en el cefalosoma y un componente menor en el metasoma. El agrupamiento de las especies en relación a su firma de poros se ajustó al agrupamiento derivado del análisis de caracteres morfológicos convencionales, como previamente se habia demostrado. para copépodos del género
Eucalanus (Fleminger, 1973). De esta manera se confirma el patrón de poros urosomaies como una herramienta adecuada y rápida para l o s estudios filogenéticos.
+
Finalmente (Park, 19951, teniendo en cuenta que la mayoría de los
estudios sobre la firma de poros en copépodos calanoides han sido hechos .sobre hembras adultas, realiza un trabajo sobre el desarrollo ontogenético.de la
firma de poros y sus diferencias sexuales, examinando a l o s machos, hembras y
copepoditos de cinco especies del género Pleuroamma.
I
'-I_.
t ''-Reportando que las diferencias interespecificas en el patrón de distribución de los poros integumentales se da desde el copepodito 111. La firma de poros cefalosomal se completa en el copepodito
V
y la firma metasomaly urosotnal se completan en el copepodito Vi, en el estado adulto. Las diferencias sexuales en la firma de poros se encuentran desde el copepodito IV
y son principalmente evidentes en el urosoma del adulto.
En base a los trabajos realizados respecto a los tipos de perforación encontrados hasta ahora en copépodos Koomen, (1992) describe los siguientes:
OPoro glandular ordinario en forma de ranura,
OPoro glandular en forma de ranura parcialmente oculto,
OPoro cerrado por un largo faldón,
"Poro glandular tubular, OClavija sensitiva, OPelo sensitivo, OEspina sensitiva,
OPoro circular pequeño,
OPoro en f o r m de urna,
OPoro en forma de botón.
.
I:
I,
11:
JUSTIPICACION
Y NATURAIJ%ZA DEL PROYECTODesde el punto de vista económico, los camarones de la familia Penaeidae soportan una pesquería comercial muy valiosa en nuestro país, ya que representan una de las principales fuentes de divisas, destinando para el mercado internacional, l a mayor parte de la producción nacional. Sin embargo, los estudios del ciclo biológico de los camarones comerciales han tropezado con diversas dificultades entre las que resalta la identificación de las larvas y su separación de aquellas otras especies que aparecen mezcladas en las capturas, pero que no son comercialmente importantes.
Uno de los problemas fundamentales es la mala utilización de las postlarvas capturadas, hecho que se origina por la falta de conocimientos en el manejo adecuado y en la identificación apropiada de estos peneidos. Para ello es importante el conocimiento de l o s caracteres de identificación a nivel postlarval de todas las especies del género Pemren8. Por lo que el haber establecido el patrón de perforación integumental en postlarvas de camarón, de la especie P. vaaaamoi será en el futuro de gran utilidad, por ser Bste un caracter permanente, importante y necesarlo para una mejor identificación de las especies. Y además estos caracteres podrán ser utilizados para separar a los organismos dentro de la misma especie, cuando se encuentren diferencias importantes.
Fue en base a los trabajos realizados sobre patrones de perforación integumental en copépodos con fines taxonómicos y filogeneticos, que nació la inquietud de determinar estos mismos patrones, pero ahora en postlarvas de camarón, ya que hasta ahora no se conocía este caracter en los peneidos.
se trabajó con la especie Penaeua
vannmmi,
por ser una de las más explotadas comercialmente en nuestro país.1:
I-
(
t
.
.
.
, ,,OBJETIVO
GENERAL
Descripción d e l patrón de perforación integumental en p o s t l a r v a s de camarón con 3 y 4 espinas r o s t r a l e s de l a especie -P A- de l a s
regiones de México y P a n a d .
OñJETiVOS
ESPECIFICO8
Determinación d e l patrón de perforación integumental en postlarvas de camarón con 3 y 4 espinas r o s t r a l e s de l a especie P.p.rru vamama procedentes d e l l a b o r a t o r i o de La Paz B.C. Sur, México.
Determinación d e l patrón de perforación integumental en p o s t l a r v a s de camarón con 3 y 4 espinas r o s t r a l e s de l a especie P a n i . w
vazuummf
procedentesd e l Laboratorio e l F a r a l l ó n , en Panamá.
Señalar l a s d i f e r e n c i a s entre l o s patrones de p e r f o r a c i ó n de ambas poblaciones.
METOüOLOGiA
UTILIZADA
Se realizó una visita al laboratorio de producci6n de postlarvas de camarón de La Paz, B. C., Sur, México, de donde se obtuvieron los organismos necesarios para la realización de este proyecto.
Para tomar l a s muestras, primero se seleccionaron los tanques de cultivo que se encontraban en mejores condiciones, o sea, en donde los organismos estaban bien alimentados, sin deformaciones ni adherencias bacterianas y los
de mayor talla. Posteriormente las muestras fueron filtradas con ayuda de un
tamiz pequefio y una caja de Petri, y después se observaron las muestras bajo el microscopio estereoqcópico. Se seleccionaron larvas con 3 y 4 dientes rostrales. De tal manera fueron tornadas las muestras
y
se seleccionaron s610los organisms que cubrían las condicioncs mencionadas anteriormente, se colocaron en frascos pequeños con forrnaldehído al 4 % para conservar lo mejor posible sus estructuras y se trasladaron al laboratorio de Biología de Cuencas Hidrológicas de la UAM-IZTAPALAPA donde se llevó a cabo el estudio de la determinación de poros integumentales de las postlarvas obtenidas, para lo cual, los organisms se transparentaron y después fueron teñidos, siguiendo las técnicas descritas por Mauchline (1988) y Park 6 Mauchline (1994).
Esta técnica consiste en transparentar primero a los organismos para eliminar el tejido blando, para lo cual primero se tomaron postlarvas de 3 y 4
espinas rostrales, de ambas regiones, se lavaron perfectamente con agua destilada para eliminar el formo1 en que se encontraban y se vaciaron
tubos de ensaye que contenian 20 m l de solución acuosa de hidróxido de potasio (KOH) al 1 0 % . Los tubos de ensaye se colocaron en. "baño maria" sobre una parrilla controlada termostáticamente a 90% en un tiempo aproximado de 5
horas,' tiempo suficiente en el cual el integumento quedó libre de tejidos blandos. Despues, se vaciaron en una caja de Petri y se lavaron con agua destilada hasta eliminar lo mejor posible el hidróxido de potasio.
a los
~.
.c
El paso siguiente fue el de teñir los organismos con una solución de negro clorazol "E" al I % , en la cual los organismos permanecieron SUmergldoS hasta obtener un color uniforme en el integumento lo que tardó de 5 'a 6 dias aproximadamente. Ya teñidos los organismos se lavaron nuevamente con agua destilada para evitar la acumulación de grumos en el integumento.
Suárez-Morales (19961.
Posteriormente, para la disección de los organismos ya transparentados y teñidos, se utilizó el liquido de Hoyer; el cual sirvió también como medio de montaje. (De la Torre, et al. 1977)
La revisión y disección de organismos se llevó a cabo bajo el microscopio estereoscópico.
Los tegumentos tetiidos
y
disectados se colocaron sobre un portaobjetos de vidrio y encima de éstos un cubreobjetos, para proteger la preparación. Estas fueron observadas bajo el microscopio óptico, donde se pudieron apreciar con mayor nitidez el número y la ubicación de los poros del integumento, como puntos deluz
que atravesaban l a superficie negra de este.Después de identificados los poros integumentales bajo el microscopio y con ayuda de una cámara de luz clara se realizaron los esquemas en donde se observan los patrones de perforación integumental de organismos con 3 y 4
espinas rostrales de las regiones tanto de México y Panamá, los cuales fueron anexados al presente trabajo.
La determinación taxonónica del patrón de perforación integumental, se realizó en base los artículos de Mauchline 6 Park (1994) y Park (1995) donde describen con esquemas, los patrones encontrados en las familias de copépodos calanoides.
OBJETIVOS Y METAS ACCANZADOS
Los objetivos planteados en el anteproyecto fueron alcanzados
satisfactoriamente, puesto que como se esperaba, las postlarvas de camarón al
igual que los copépodos presentan un patrón de perforación integumental, el
cual no había sido determinado anteriormente en estos P o n d d o . y el ser pionera
en este campo de estudio es un motivo de satisfacción personal.
El haber alcanzado la meta propuesta al inicio de este trabajo, resulta
un incentivo para seguir adelante, planteando nuevos objetivos y metas a
RESULTADOS Y
DISCUSI~N
Los patrones de perforación integumental encontrados en las postlarvas de camarón de 3 y 4 espinas rostrales, de la especie pliuriu %amammi
pertenecientes a las regiones de MQxico' y Panamá, se muestran en la8 Figiiras:
(2, 3 , 4 y 5 ) .
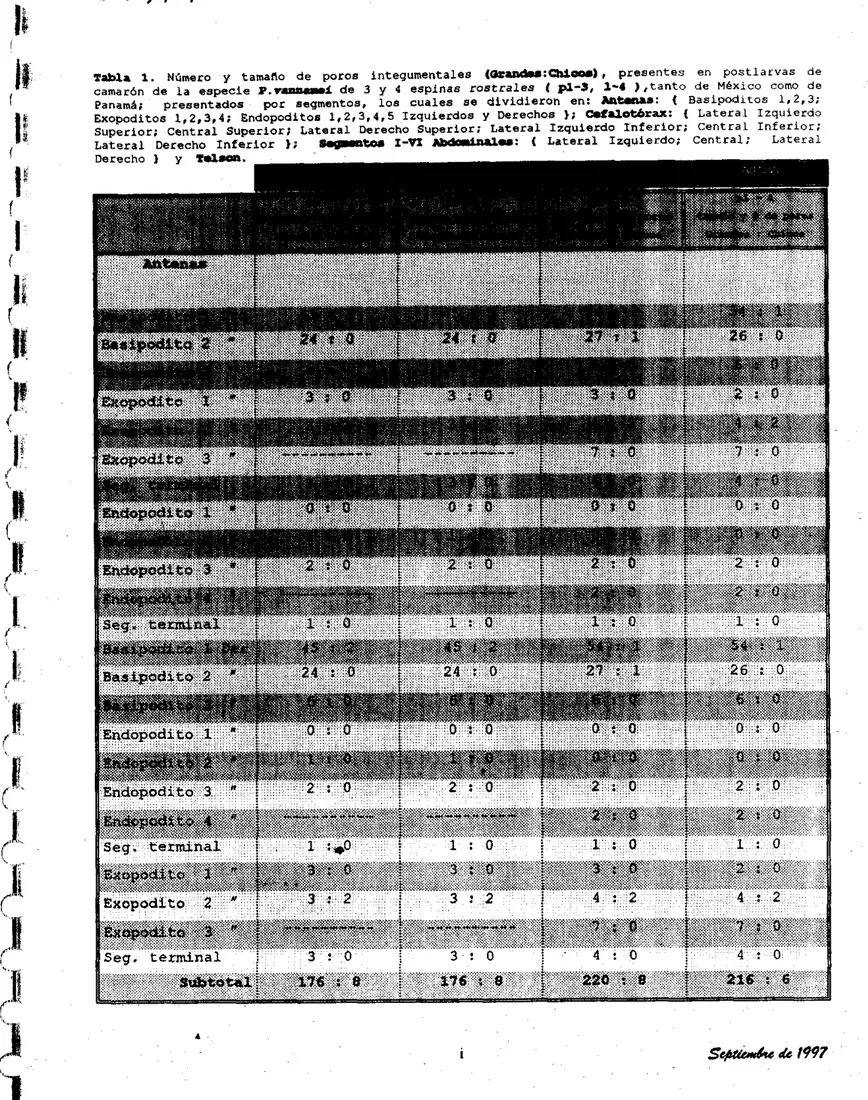
El número de poros y posición en que se encuentran, se pueden consultar en la Tabla 1 de resultados.
Los poros integumentales encontrados son todos simétricos, los cuales, se clasificaron en 2 tamaños : Grandes y Chicos (O:-); el conteo de los mismos, se realizó por segmentos: An- (basipoditos, exopoditos, endopodrtos);
C e f d o t ó r u (Lateral Izquierdo Superior (LISup. )
,
Central Superior (CSup.1,
Lateral Derecho Superior (LDSup.), Lateral Izquierdo Inferior ( L I I d . ) , Central Inferior ( C l n f . ) , Lateral Derecho Inferior (LDInf.); Srgmuitos ibdoininales
(I, 11, 111, IV, V, VI) además cada segmento se dividió en: Lateral Izquierdo
(U), Central (C)
,
Lateral Derecho (LD) y '&)@on.Ver Figura 1.
A continuación, se elabora un análisis detallado del número de poros encontrados en cada segmento por número de dientes rostrales y l a s diferencias encontradas de una región a otra.
wsTLARvAs DE 3 DIENTES ROSTRWS DE &ICO Y
~ANlllBi
Antonas
En los organismos de MÉxico se encontraron (176:8) poros: (antena izquierda, baaipoditos: 1 ( 4 5 : 2 ) , 2 (24: O ) , 3 (6: O ) : exopoditoa: 1 (3:0), 2 I3:Z) ;
segmanto trrminal:(3:0); +ndopoditos: l(0:O). 2(1:01, 3(2:0); segmcnto
terminal: ( 1 : O ) ; antena derecha, basipoditos: 1 ( 4 5 : 2 ) , 2(24:0), 3(6:01;
endopodites: l(0:Ol. 2 ( 1 : 0 ) , 3(2:0); segmento terminal: í 1 : O ) ; exopoditos:
1(3:0), 2(3:2); aegnmnto terminal: (3:O) j y en los de Panamá también hay
(176:8) poros, los cuales concuerdan exactamente en número y posición con los
de México.
Cefalotbrur
Esta p a r t e s e consideró l a más importante, ya que es l a m6s grande, (por
lo que s e d i v i d i ó en 6 regiones) y donde se encontraron d i f e r e n c i a s numéricas
mayores que en l o s o t r o s segmentos. En l o s organismos de México s e tienen
(106:18) poros: (en l a s regiones superiores, hay ( 1 8 : O ) en cada l a t e r a l y
( 3 9 : 9 ) en l a c e n t r a l ; para l a región i n f e r i o r (12:O) en cada l a t e r a l con ( 7 : 9 ) en l a c e n t r a l ) ; mientras que en l o s de Panamá hay (84:16) poros: (en l a s regiones superiores, hay (14:O) en cada l a t e r a l , ( 3 3 : 7 ) en l a c e n t r a l y en l a región i n f e r i o r (9:O) para cada l a t e r a l con ( 5 : 9 ) en l a c e n t r a l ) . Lo que nos da
una d i f e r e n c i a en l a región superior de (4:O) por cada l a t e r a l y (6:2) en l a c e n t r a l ; En l a región i n f e r i o r l a d i f e r e n c i a es d e (3:O) en cada l a t e r a l y
(2:O) en l a c e n t r a l .
saginnto I Abdolninal
En e s t e segmento, s e obtuvieron ( 4 6 : Z ) poros en l o s de México: ( ( 1 O : O ) en
cada región l a t e r a l y ( 2 6 : 2 ) en l a c e n t r a l ) y para l o s de Panama ( 3 4 : 2 ) poros:
( ( 9 : O ) en cada r e g i b n l a t e r a l y (16:2) en l a c e n t r a l ) . Por lo que l a s
d i f e r e n c i a s son de ( 1 : O ) en cada l a t e r a l y ( 1 O : O ) en l a región central.
segnvnto Ii -rial
En l a región de México, se encontraron (52:6) poros: ((8:O) en cada
región l a t e r a l y (36:6) en l a región c e n t r a l ) ; en l o s organismos de Panamá fueron (44:Z) poros: ( ( 7 : O ) en cada regrón l a t e r a l y (30:2) en l a reglón c e n t r a l ) . Entonces l a s d i f e r e n c i a s en e s t e segmento, son s ó l o de (6:4) en l a región c e n t r a l y de ( 1 : O ) para cada región l a t e r a l .
s a g w n t o 111 Abdominal
Se encontraron (40:Z) poros en l o s de Mexico: l ( 7 : O ) e n ' cada región
l a t e r a l y ( 2 6 : 2 ) para l a c e n t r a l ) . En l o s que corresponden a Panama s e tienen (34:4) poros: {(6:0) para cada región l a t e r a l y ( 2 2 : 4 ) en l a c e n t r a l ) . Aquí, se observó una d i f e r e n c i a d n más pequeña que en e l segmento a n t e r i o r , ya que l a variación fue de (1:O) en cada región l a t e r a l y (4:2) en l a c e n t r a l .
segmento IV Abdominal
Los organismos de México tlenen (36:2) poros: ((7:O) para cada reglón
Por lo que se puede observar una diferencia de (2:O) poros para cada lateral y de (6:2) en la central.
SOqXlMItOVAbdCdMl
Se observaron (34:Z) poros: ( ( 7 : O ) en cada lateral y (20:Z) en la central) en los organismos pertenecientes a México; (24:Z)poros: ((7:O) para cada lateral y (10:2) en la región central), en
los
de Panamá. Existiendo una diferencia sólo en la región central de (1O:O) poros.-to VI
-w
Por ser este segmento más grande que los d 4 s , se encontró
un
m y o r número de poros. Para los de México hay (108:O) poros: 1(18:0) en cada región lateral y (72:O) en la central); los de Panamá tienen (86:O) poros: ((17:O)para cada lateral y (52:O) en la regi6n central). La diferencia fue de ( 1 : O ) para cada región lateral y de (2O:O) en la región central.
‘fd80ll
Los organismos de México presentan (26:O) poros y los de Panamá sólo
(14:O) poros, lo que da una diferencia de (12:O) poros. En esta parte no $e encontraron poros chicos para los organismos de ninguna de las dos regiones.
Tenemos entonces que en las postlarvas de 3 espinas rostrales de México, se encontró un total de (626:40) poros; mientras que las postlarvas de Panamá, presentan (522:34) poros.
Comparando los patrones de perforación integumental, de las postlarvas de
3 espinas rostrales
,
pero de diferente región (México y Panamá), se observa que generalmente las‘ que pertenecen a México tienen un número mayor de poros que las de Panamá en todos los segmentos; excepto en las antenas, donde tienen el mismo número y el segmento V abdominal con el mismo número de poros chicos( 2 ) .
*
Anturrs
En los organismos de México se encontraron (220:8) poros: (antena
izquierda, basipoditos: 1 ( 5 4 : 1 ) , 2(27:1), 3(6:0); --tos: 1(3:0), 2(4:2), 3(7:0); s-nto h d ~ l : ( 4 : O ) ; endopoditos: l ( O : O ) , 2(0:0), 3 ( 2 : 0 ) , 4(2:0);
.-to trrminrl: (1:O); antena derecha, brsipoditos: 1(54:1), 2(27:1), 3(6:0);
uidopoditos: 1(0:0), 2 ( 0 : 0 ) , 3(2:0), 4(2:0); s q m n t o terminai: i1:Oi;
uopoditos: 1 ( 3 : 0 ) , 2(4:2), 3(7:0); s-to tmdnrl: ( 4 : O ) ) y en los de
Panamá hay (216:6) poros: l o s cuales son muy parecidos en número y p o s i c i ó n a
los de Mexico, pues l a s d i f e r e n c i a s s e encuentran s ó l o en e l hsipodito 2
(l:i), y en e l mxopdita 1 (1:O).
Cefaiotórax
A1 i g u a l que en l a a n t e r i o r , e s t a p a r t e se consideró l a más importante,
ya que e s l a más grande y donde s e encontraron d i f e r e n c i a s numéricas mayores
que en los o t r o s segmentos (por l o que se d i v i d i ó en 6 p a r t e s ) . En los
organismos de México se t i e n e n (147:28) poros: (en l a s regiones superiores, (21:O) en cada l a t e r a l , (65:7) en l a c e n t r a l y para l a r e g i ó n i n f e r i o r (15:2) en cada l a t e r a l con (10:17) en l a central); mientras que en los de Panamá
(113:16) poros: (en l a s regiones superiores, (17:O) en cada l a t e r a l , (41:7) en
l a c e n t r a l y para l a región i n f e r i o r ( 1 4 : O ) en cada l a t e r a l con (10:9) en l a
c e n t r a l ) . Lo que nos da una d i f e r e n c i a en l a r e g i ó n superior de ( 4 : O ) en l a s
l a t e r a l e s y (24:O) en l a central; En l a región i n f e r i o r l a d i f e r e n c i a es de (1:2) en l a s l a t e r a l e s y (0:8) en l a central.
segmento I Abdonir ' n d
En e s t e segmento, s e obtuvieron (58:16) poros en ambas regiones; para México { ( l l : i ) , en cada región l a t e r a l , (36:14) e n l a c e n t r a l ) y para Panamá, s e encontraron: ( ( 1 2 : O ) e n cada región l a t e r a l y (34:16) en l a c e n t r a l ) . Por l o que l a s d i f e r e n c i a s son de w s i c i ó n solamente, e l número de poros es e l mismo.
segmento 11 Abdominal
En l a r e g i ó n de México, se encontraron (68:8) poros ( ( 1 O : O ) en cada
región l a t e r a l y ( 4 8 : 8 ) en l a región c e n t r a l ) ; en l o s organismos de Panamá
fueron ( 6 0 : 1 2 ) p o r o s , ( ( 7 : O ) e n cada región l a t e r a l y l 4 6 : 1 2 ) en l a región c e n t r a l ) . . Por l o tanto l a s d i f e r e n c i a s en e s t e segmento, son s ó l o de ( 2 : O ) e n
la región central
y
de (3:O) para cada región lateral. Las de Panamá en la región central presentaron (0:4) poros m6s.8ylP.nto 111 -1181
Se encontraron (56:14) poros en los de México ((1l:O) en cada región lateral y (34:14) para la central). En los que corresponden a Panamá se tienen (56:16) ( ( 1 O : O ) para cada región lateral y (36:16) para la región central) Aqui, la pequeña diferencia fue de (0:Z) poros más para Panamá,; y también en
s u posición.
s y p n t o fv A b d o d m i
Los organismos de México tienen (50:6) poros ((1O:l) para cada región lateral y (30:4)en la central); a diferencia de loa de Panamá que presentan (48:lZ) poros ((11:Z) en cada lateral y (26:8) centrales). Por lo que se puede observar una diferencia de (1:l) para cada lateral y de (4:4) centrales.
8-nto V Abdonu ' nal
Se observaron (42:6) poros ( ( 1 O : O ) en cada lateral y (22:6) centrales) en
los organismos pertenecientes a México y (38:6)poros: ((1O:O) para cada lateral
y (18:6) en la región central), en l o s de Panamá. Existiendo diferencia sólo en
la región central de ( 4 : O ) poros.
Segmento VI Abdomi ' n i l
Por ser este segmento más grande que l o s demás, se encontró un mayor número de poros. Para los de México hay (132:6) poros ((22:O) en cada región lateral y (88:6) en la central}; los de Panamá tienen (11O:Z) poros ((2O:O)
para cada lateral y (70:Z) en la región central). La diferencia fue de (2:O) para cada región lateral y de (18:4J en la región central.
Telson
Los organismos de Méaico presentan ( 3 2 : 4 ) poros y los de Panamá (26:O) poros, lo que da una diferencia de ( 6 : 4 ) poros. En esta parte no se encontraron poros chicos para los organismos de Panamá.
Tenemos entonces que en las postlarvas de 4 espinas rostrales, las de la
región de México presentaron ( 8 0 5 : 9 6 ) poros, y en las de Panamá (725:86) poros.
Lo que nos da una diferencia total de ( 8 O : l O ) poros. (Tabla 21.
i
,w.
.*I
.
,I.
c1,
,
lY1 ,
-Comparando l o s patrones de perforación integumental de l a s postlarvas de
4 espinas rostrales
,
pero de diferenta región (México y Panamá), se observaque generalmente l a s que pertenecen a México tienen un número mayor de poros
que l a s de Panamá en todos los segmentos; excepto en e l basipodito 2 con una
diferencia de (1:l); en e l exopodito 1 con ( 1 : O ) ; en e l segmento I abdomhal en
donde tienen e l mismo número de poros chicos y grandes en ambas regiones; y en
e l segmento I11 abdominal en donde tienen también e l mismo número de poros
grandes pero l o s de Panamá tienen 2 poros chicos más.
Ij
,*'.La suposición de que las larvas de camarón como los copCpodos presentan
un patrón de perforación en su tegumento es verdadera, y el presente trabajo lo
demuestra.
Al termino del presente, se llegó a la conclusión de que las postlarvas
de camarón de P. vanrum& presentan un patrón de perforaci6n integumental
distinto para cada región y también cambian según su estadio, esto es que
aumentan mientras va creciendo el organismo. Por lo que se presentan más poros
en los organismos de 4 espinas rostrales comparados con los que tienen sólo 3
espinas.
En las postlarvas de 3 dientes rostrales se encontraron variaciones de
una región y otra tanto en el cefalotórax como en todos los segmentos y el
telson, presentando un mayor número de poros en las pertenecientes a la región
de México.
Mientras que en las postlarvas con
4
dientes rostrales esta variación se. .present6 desde las antenas, cefalotórax, los segmentos y el telson, de igual
manera fue en la región de México donde se obtuvo un mayor número de poros,
excepto en los segmentos abdominales I y IV, donde éstos fueron iguales para
ambas regiones.
Referente al valor taxonómico que representa el patrón de perforación
integumental en las postlarvas de camarón es muy importante, debido a que este
nuevo caracter es permanente en las especies de cada localidad y permite una
mejor identificación de estas.
Estos resultados nos dan l a pauta para posteriormente determinar la
separación de subespecies.
RECOMENMCIONES
Ai i g u a l que como lo hicieron Park 6 Mauchline, (1994) con l o s copépodos
del género ~lrrisrriyri, s e r i a importante determinar l o s patrones de perforación
integumental pero ahora con l a s postlarvas de l a s diferentes especies que existen d e l género P a m i r . De donde se puede i n f e r i r que a l determinar l o s patrones de perforación en cada una de l a s especies de Peneidos se encontraran diferencias entre é s t a s .
.
Para esto, se puede t o m r como base e l presente trabajo donde se
determinó e l patrón de perforación integumental en l a s postlarvas de l a especie
P. 9uIII.DLI.
Asi, a l tener l o s patrones de peKfOKaCi6n de cada especie, se podrán
comparar y diferenciar entre s i .
LITERATURA
CITADAANONIMO, 1990a. El cultivo de camar6n en México y la participación de Informe, 20 p.
POPESCA,
ANONIMO, 1990b. World shrimp farming. Aqurcml+urr Digrst, 28p.
CABRE:RA-JIM~EIEZ,J.A.,~~~~.
Characteres of taxonomic values of the postlarvae ofthe shrimp. P.nrrus (Farfantepenaeus) brevirostris Kinsley(Decapoda Natantia) of the Gulf the California,México, CrustaCaeuu.44(3):292-300.
CALDERÓN-P$REZ, J.A., E.MACtAS-REGAWIM>, y S.RENDÓN-RODRfGUE2, 1989. Clave de identificacih para los estadios de postlarva y primeros juveniles de camarón del género
-=
(Crustácea:DecBpoda) del Golfo de California, México.Cirnoias W i n a n , 15(3):57-70.
DE LA LANZA-ESPINO,G., S.GAMEZ-ETERNOD, 1992. AnAlisi6 del a s h d o da la
ouironiaultru. on YLXico hasta el aiio 1991. México, D.F. 48p.
DE
LA
TORRE,G.G., C.JUÁREZ., H.H.FIüüEROA.,1977. Tianiocis biológiou salaatas.da laborrtorio y Q euppo. Editorial Limusa. México. 251p.
FERRAR1,F.D. y T.E.BOWMAN, 1980. Pelagic copepods of the family Oithonidae (Cyclopoida) from the est coast of Central and South America. Cmithson.Contrib. 2001. 312:l-27.
FLEMINGER,A., 1973. Pattern, number, variability and taxonomic significance of
integumental organs (sencilla and glandular pores) in the genus Euaaianu6 (Copepoda, Calanoida). Pish. Bull., 71:965;1010.
FLEMINGER,A., y K . Huselmann, 1977. Geographical range and taxonomic divergence in North Atlantic Calanus &C. helgolandiciur, C. finmarcñicw y , C. g l a c i a l i r ) . Mar. Biol., 40:233-248.
HULSEHA",K. y A.FLEMINGER, 1990. Taxonomic value of minute StrUCtres on the genital segment of Pontellina females ( Copepoda:Calanoida ) . Mar Biol.,
105:99-108.
KOOMEN,P.,1992. The integumental perforation pattern of the Euchirella mesinensis female (Copepoda,Calanoida): corrections, additions,
variation, and checklist of pore
intraspecific sites. Cnutaarura 63(2):113-159.
tmcIAs-mGAuum,~.,
H . F E ~ D E Z y A . c A L D E R ~ N , ~ ~ ~ Z . Variación diurna de la densidad de postlarvas de camar6n en la boca del sistema lagunar Huizache-Caimanero Sinaloa, México. ( Crustácea: Decápoda: Penaeidae ) . An.Inst. Ciana. dol Y.r y -01. Univ.Nal.Aut6n.Méx.,9(1):381-386.
MAIR,J.MCD., 1979. The identification of postlarvae of four species of -8
(Crustacea: Decápoda) from the Pacific Coast of Mexico. Jour. Zool., London,
188:347-351.
MARTÍNEZ-CÓRDOVA,L.R., 1993. C . m i r o h i d t U r i : b.8-8 y d W l e f i a 8 para
a1 o u l t i ~ & crrmronr. prarido8. A.G.T.Editor,S.A. Méxic0,D.F. 23313.
MAUCHLINE,J., 1987. Taxonomic value of pore pattern in the integument of calanoid copepods (Crustacea) Jour. Zool., b a d . 211:697-149.
PARK,J.S. 1995. The Development of integumental pore signatures in the genus Pleurani.nsi.(Copepoda Calanoida). J . Mar. M o l . A s s . U.
K.
Great Britain.75:211-218.
PARK,J.S. y J.MAUCHLINE, 1994. Evaluation of integumental pore signatures of
species of calanoid copepods(Crustacea) f o r interpreting inter-species relationships. Mar. Biol. 120:107-114.
CEPESCA, 1992. Anuario Estadístico de Pesca. Dirección Gonerd da Infornitica y
Registro Peiqueros.
.
CUÁREZ-MORALEC,E.; J.W.REID; T.M.ILIFFE y F.FIERS, 1996. Catilogo de los
copépodos (Crustacea) continentales de l a Peninsula de Yucatán, Máxico. IECOSUR
y CONABIO), México, D.F. 493p.
*
Tabla 1. Nilmero y tamano d e poros integumentales (--:Woo.), presentes en postlarvas de camarón de l a especie P.v-i d a 3 y 4 espinas rostrales ( pl-J, 1-4 ),tanto de México como de Panamá; presentados por eegmentos, loe cuales se dividieron en: & W S : ( Basipod~tos 1.2.3; Exopoditos 1.2.3.4; Endopoditos 1,2,3,4,5 Izquierdos y Derechos 1; O I 8 l o t ó r u : ( Lateral Izquierdo Superior; Central Superior; Lateral Derecho superior; Lateral Izquierdo Inferior; Central Inferior; Lateral Derecho Inferior ); reiiitr>. I-Vi ( Lateral Izquierdo; Central: Lateral Derecho I y TU-.
3
2 : o
3 : 2 4 : 2 4 ' : 2
Seg. terminal
i
3 : o 3 : o 4 : o 4 : oI
!
220: 8 216 : 6
ACTIVIDADES
REALIZADAZITabla 2 . Se describen las actividades realizadas durante la elaboración d e l proyecto y l a duraci6n
w--*
Sogmonto tatmlnll
Exopodito 2
Endopodlto 2
‘EXOPOdltO,
Endopodite'
,SEGMENTO II ABDOMINAL
NTO 111
SEGMENTO
N
ABDOMINAL-SEGMENTO V ABDOMINAL
VI ABDOMINAL
TELSON
T i p u n 1. ~ s q i i ~ m a d e postlarva dp camar6n de l a r s p ~ c i e P. v-I divldidu en reglones, p a r a l a
especificocii~5n d e l p d t i ú n de prrforaciún i n t e q i i m r n t a l .
~ I
'1
.
F i g u r a
.
..
.
I:
.
. . .
, <..
< *;I
i,:
,.".
.I;.
'1
.
.
....
....
\,:
'--n-/
, ..+ 7.\ .
P. rinniiri d e
...