R E S E A R C H A R T I C L E
Open Access
16/6-idiotype expressing antibodies induce brain
inflammation and cognitive impairment in mice:
the mosaic of central nervous system
involvement in lupus
Shaye Kivity
1,2*†, Aviva Katzav
1,3†, Maria Teresa Arango
1,4,5, Moran Landau-Rabi
1, Yaron Zafrir
1,
Nancy Agmon-Levin
1, Miri Blank
1, Juan-Manuel Anaya
4, Edna Mozes
6, Joab Chapman
1,3and Yehuda Shoenfeld
1,7Abstract
Background:The 16/6-idiotype (16/6-Id) of the human anti-DNA antibody was found to induce experimental lupus in naïve mice, manifested by production of autoantibodies, leukopenia and elevated inflammatory markers, as well as kidney and brain involvement. We assessed behavior and brain pathology of naive mice injected
intra-cerebra-ventricularly (ICV) with the 16/6-Id antibody.
Methods:C3H female mice were injected ICV to the right hemisphere with the human 16/6-Id antibody or commercial human IgG antibodies (control). The mice were tested for depression by the forced swimming test (FST), locomotor and explorative activity by the staircase test, and cognitive functions were examined by the novel object recognition and Y-maze tests. Brain slices were stained for inflammatory processes.
Results:16/6-Id injected mice were cognitively impaired as shown by significant differences in the preference for a new object in the novel object recognition test compared to controls (P= 0.012). Similarly, the preference for spatial novelty in the Y-maze test was significantly higher in the control group compared to the 16/6-Id-injected mice (42% vs. 9%, respectively,P= 0.065). Depression–like behavior and locomotor activity were not significantly different between the16/6-Id-injected and the control mice. Immunohistochemistry analysis revealed an increase in astrocytes and microglial activation in the hippocampus and amygdala, in the 16/6-Id injected group compared to the control. Conclusions:Passive transfer of 16/6-Id antibodies directly into mice brain resulted in cognitive impairments and histological evidence for brain inflammation. These findings shed additional light on the diverse mosaic
pathophysiology of neuropsychiatric lupus.
Keywords:Systemic lupus erythematosus, 16/6 idiotype, Anti-DNA, Neuropsychiatric lupus, Cognitive impairment
Background
Neuropsychiatric systemic lupus erythematosus (NPSLE) refers to a complex set of syndromes involving the central nervous system (CNS) in up to 56% of lupus patients [1-5]. Due to the varied diagnostic criteria applied to define NPSLE, the American College of Rheumatology has
proposed a standard nomenclature of case definitions, reporting standards and diagnostic testing recommen-dations for the 19 neuropsychiatric Systemic lupus erythematosus (SLE) syndromes [6]. While some of the focal manifestations (for example, stroke) can be explained by vasculitic or thrombotic lesions, the pathogenicity of more diffuse manifestations of NPSLE (for example, cogni-tive impairment, depression and psychosis) remains rela-tively obscure. Nevertheless, studies have demonstrated the importance of various factors involved in the development of diffuse neuropsychiatric manifestations, such as the presence of autoantibodies, inflammatory mediators (for
* Correspondence:[email protected]
†
Equal contributors
1
The Zabludovicz Center for Autoimmune Diseases, Sheba Medical Center, Tel-Hashomer, 2 Derech Sheba St., Ramat Gan 52621, Israel
2Rheumatic Disease Unit, Sheba Medical Center, Tel-Hashomer, 2 Derech
Sheba St., Ramat Gan 52621, Israel
Full list of author information is available at the end of the article
example, cytokines, matrix metalloproteinases), neuropep-tides and endocrine factors [7-10]. Other factors, such as medications and primary neurologic and psychiatric disor-ders, may play a major role as well.
More than 20 brain specific and non-specific autoanti-bodies have been proposed to be involved in the mechanism of NPLSE [11], including anti-neuronal [12], anti-ribosomal–P [13,14], phospholipid [15] anti-bodies, as well as anti NR2/anti-DNA antibodies that cross react with N-methyl-D-aspartate (NMDA) recep-tors [3,16]. During the last two decades, anti-DNA idiotypes were characterized, and found to play an im-portant role in systemic lupus erythematosus and NPSLE [17]. The 16/6 idiotype (Id) antibody is a human anti-single-stranded-DNA (anti-ssDNA) monoclonal antibody (mAb) originated from a patient with cold agglutinin dis-ease [18]. The 16/6-Id was found to be polyspecific [19], cross reacting with cytoskeletal proteins (vimentin), platelets, lymphocyte membranes, pathogens such as Klebsielapolysaccharides andMycobacterium tuberculosis glycoproteins, brain glycolipids and tumor cells [20-22]. The presence of 16/6-Id was detected in 30% of lupus patients, and their levels were found to correlate with disease activity [23,24]. Elevated titers of 16/6-Id were also detected in NPSLE patients [25]. Deposits of 16/6-Id were found in the skin, kidney and brain tissue [21,26,27], and were found to bind human cortical brain tissue sections ex vivo. The presence of circulating 16/6-Id was detected in patients with other autoimmuine diseases as well (for example, polymyositis, systemic sclerosis) [28,29]. Immunization of naïve mice with the human anti-DNA 16/6-Id mAb was shown to induce experimental lupus manifested both serologically and clinically. A wide profile of mice autoantibodies (for example, mouse 16/6-Id, and antibodies against dsDNA, ssDNA, Ro, La, RNP, Sm, histones, cardiolipin and phosphatydilserine), were detected, as well as leukopenia, elevated erythrocyte sedi-mentation rate (ESR), proteinuria and the deposition of immunoglobulins in the kidney mesangium [30-32]. In addition, recent-preliminary data showed histological brain changes in mice with experimental SLE induced by active immunization with the 16/6-Id (A. Marom and E. Mozes, unpublished results). Therefore, we hypothesized that the 16/6-Ids have a pathogenic role in neuropsychiatric lupus. In the present study we investigated the effect of 16/6-Id on behavioral and cognitive functions, as well as on the brain pathology of naïve mice injected intra-cerebra -ventricularily (ICV) with the 16/6-Id.
Methods
Mice, antibody injection and experimental design
Mice
Three-month-old, female C3H mice were obtained from Harlan Laboratories, Jerusalem, Israel, and were housed
in the animal facility at Sheba Medical Center. The mice were raised under standard conditions, 23 ± 1°C, 12-hour light cycle (6:30 am to 6:30 pm) with ad libitum access to food and water. The Sheba Medical Center Animal Welfare Committee approved all procedures.
Monoclonal 16/6-Id expressing antibodies
The human monoclonal anti–DNA antibodies were produced by a hybridoma derived from fusion of the GM4672 lymphoblastoid cell line and peripheral blood or splenic lymphocytes obtained from three lupus pa-tients. The human mAb that bears the 16/6-Id (IgG1/k) has been characterized previously [33]. The mAb was se-creted by hybridoma cells that were grown in culture and were purified by using a protein G-sepharose col-umn (Pharmacia, Fine Chemicals, Uppsala, Sweden).
The injection process is based on a detailed protocol reported by Shoenfeld et al. [34]. Mice were anesthe-tized by intra-peritoneal (i.p.) injection of ketamine (100 mg/kg) and xylazine (20 mg/kg). The skull was carefully exposed, and a small hole was drilled with a 25-gauge needle above the right lateral ventricle (2 mm lateral to the midline and 2.5 mm posterior to the bregma). A 27-gauge needle attached to a Hamilton syr-inge was inserted at this point to a depth of 2 mm, where preliminary tests had confirmed accurate ICV placement by injection of dye. Then 1μl of anti-DNA 16/6-Id mAb
or control IgG was slowly infused, the needle was with-drawn and the skin over the scalp was sutured. All anti-body solutions used contained 6 mg protein/ml. Each mouse received only a single injection.
Experimental design
Twenty-one CH3 mice were injected ICV to the right hemisphere, 11 with human 16/6-Id antibodies and 10 with commercial human IgG antibodies (control). The forced swimming test (FST) was performed at Days 16 and 23 after antibody injection, the staircase test at Days 14 and 26, the novel object recognition at Days 19 and 20 and the Y-maze test at Day 21. At Day 24, under anesthesia, a systemic perfusion was performed, and the brains were collected. Immunofluorescence staining was performed to detect markers of inflammation or neur-onal degeneration (see below).
Cognitive and behavioral tests
Forced swimming test
re-exposed to the swimming condition in a similar envir-onment, and then subjected to a test session for six mi-nutes. The behavioral measure scored was the duration (in seconds) of immobility, defined as the absence of escape-oriented behaviors, such as swimming, jumping, rearing, sniffing or diving, recorded during the six-minute test. A depression-like behavior was considered as an increased immobility time.
Staircase test
Locomotor and explorative activity was evaluated by the staircase test, as described previously by Katzav et al. [15]. This test analyzes locomotor and exploratory activ-ities (stair-climbing) and anxiety (rearing). The staircase maze consisted of a polyvinyl chloride enclosure with five identical steps, 2.5 × 10 × 7.5 cm. The inner height of the walls was constant (12.5 cm) along the whole length of the staircase. The box was placed in a room with constant lighting and isolated from external noise. Each mouse was tested individually. The animal was placed on the floor of the staircase with its back to the staircase. The number of stairs climbed and the number of rears were recorded during a three-minute period. Climbing was defined as each stair on which the mouse placed all four paws; rearing was defined as each in-stance the mouse rose on hind legs (to sniff the air), ei-ther on the stair or against the wall. The number of stairs descended was not taken into account. Before each test, the animal was removed and the box cleaned with a diluted alcohol solution to eliminate smells.
Novel object recognition test
This is a visual recognition memory test based on a method described by Tordera et al.[36]. The apparatus, an open field box (50 × 50 × 20 cm), was constructed from plywood painted white. Three phases (habituation, training and retention) were conducted on two separate test days. Before training, mice were individually habitu-ated by allowing them to explore the box for one hour. No data were collected at this phase. During training sessions, two identical objects were placed into the box in the northwest and southeast corners (approximately 5 cm from the walls), 20 cm away from each other (sym-metrically) and then the individual animal was allowed to explore for five minutes. Exploration of an object was defined as directing the nose to the object at a distance of ≤1 cm and/or touching it with the nose; turning
around or sitting near the object was not considered as exploratory behavior. The time spent in exploring each object was recorded. The animals were returned to their home cages immediately after training. During the reten-tion test, the animals were placed back into the same box after a four-hour interval, and allowed to explore freely for five minutes. One of the familiar objects used
during training was replaced by a novel object. All ob-jects were balanced in terms of physical complexity and were emotionally neutral. The box and the objects were thoroughly cleaned by 70% alcohol before each session to avoid possible instinctive odorant cues. A preference index, a ratio of the amount of time spent exploring any one of the two items (old and new in the retention ses-sion) over the total time spent exploring both objects, was used to measure recognition memory. Individual ani-mals demonstrating insufficient task performance were excluded from later specific statistical analyses for the fol-lowing reasons: (1) non-exploration, which was defined as no objection interaction or (2) technical malfunctions dur-ing data collection.
Y maze test
The Y maze test was used to assess spatial memory. It was comprised of three arms, built of black Perspex. Each arm was 8 × 30 × 15 cm at an angle of 120° from the others. One arm was randomly selected as the start arm. Each mouse was placed twice in the start arm. On the first trial, lasting for five minutes, one of the other two arms was randomly chosen to be blocked whereas on the second trial, lasting for two minutes, both arms were open. The two trials were separated by a two-minute interval, during which the mouse was returned to its home cage. The time spent in each of the arms was measured. Between each trial and between each mouse, the maze was cleaned with a 70% alcohol solu-tion and dried. Discriminasolu-tion of spatial novelty was assessed by a preference index [37]: time in the new arm - time old arm/time in the new arm + time in the old arm, assessing spatial memory. The mouse is expected to recognize the old arm as old and spend more time in the new arm.
Immunofluorescence staining
Brain perfusion and fixation
The mice were anesthetized by an i.p. injection of keta-mine (100 mg/kg) and xylazine (20 mg/kg) and sacrificed by transcardiac perfusion with phosphate buffered saline (PBS) followed by perfusion with 4% paraformaldehyde (PFA, Sigma-Aldrich Israel Ltd., Rehovot Israel) in phos-phate buffer (PO4, pH 7.4). After perfusion, the brain was quickly removed and fixed overnight in 4% PFA (in PO4, pH 7.4) at 4°C. On the following day, the brain was cryoprotected by immersion in 30% sucrose in 0.1M PO4 (pH 7.4) for 24 to 48 hours at 4°C before brain cutting.
Brain cutting and preservation
Frozen coronal sections (30 to 50 μm) were cut on a
follows. Six mice (three IgG control and three 16/6 Id) were used for immunohistochemistry. Brain sections were stained free-floating, incubated with the first anti-bodies overnight at 4°C. The slices were then washed in PBS + 0.1% Triton X-100, and incubated at room temperature for one hour with the corresponding fluor-escent chromogens-conjugated secondary antibody. Sec-tions were stained for specific antigens with antibodies against activated microglia (anti-Iba1, pAb, Abcam, Cambridge, UK) and astrocytes (anti-GFAP mAb, Dako, Carpinteria, CA, USA). Counter staining was performed with Hoechst (Sigma-Aldrich Israel Ltd., Rehovot Israel).
Statistical analysis
Results are expressed as the mean ± SEM. The differ-ences in mean for average immobility time in the FST, the staircase test parameters (number of rearing and stair-climbing events), novel object recognition and Y-maze tests were evaluated by T-test. Significant results were determined asP<0.05.
Results
Cognitive and behavioral performance
The results of cognitive performance in the novel object recognition test are presented as the proportion time spent near objects (new and old) in both groups (Figure 1). There was a significant preference for attention to the new object in the control group (64% time spent near the new object compared to 36% time spent near the old ob-ject,P= 0.012), while no difference in the preference was seen in the mice injected with 16/6-Id (56% vs. 44% time
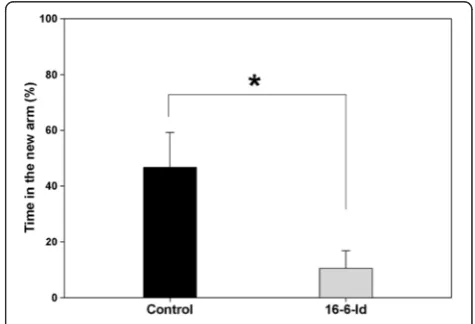
spent near the new object vs. old object,P= 0.655). This suggests a specific visual recognition memory impairment in the 16/6-Id mice. Similarly, cognitive performance in the Y-maze test is presented as a preference index for new (additional percent time spent in the novel arm) in both groups (Figure 2). The control IgG mice spent 46% add-itional time in the new lane while the mice injected with 16/6-Id spent 9% additional time in the new lane (P = 0.015 byt-test).
In the forced swimming test there was no significant difference between 16/6-Id injected and control mice in depression-like behavior at Days 16 and 24 after injec-tion. Average immobility times of the control mice vs. 16/6-Id injected mice were 117.6 ± 65.9 vs. 160 ± 72.8 (P = 0.159 by t-test) and 182.5 ± 45.4 vs. 205.7 ± 42.7 sec (P= 0.238 byt-test) on Days 16 and 24, respectively.
In the staircase test, there was no significant difference between the average rearing and stair-climbing counts, among mice from control-IgG vs. 16/6-Id (23.7 ± 2.6 vs. 21.8 ± 2.5 rearings, and 24.5 ± 2.3 vs. 16.5 ± 4.4 stair-climbing events, respectively,P >0.016). The results also did not change from Day 14 to 26.
Brain pathology
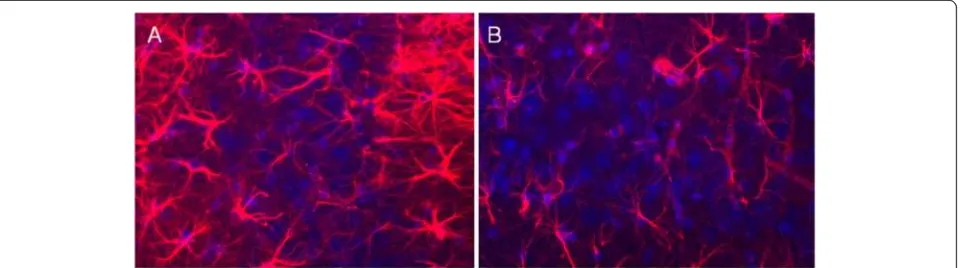
[image:4.595.302.540.457.619.2]Brain sections were stained for activated microglia and astrocytes (as markers for inflammation). The 16/6-Id injected mice demonstrated increased microglial activa-tion (Iba-1 staining), at the hippocampus (CA1, CA3, dentate gyrus, stratum radiatum) as well as the amyg-dala, compared to IgG control (Figure 3). The difference
Figure 116/6-Id injected mice displayed impaired performance in the novel object recognition test.Results are presented as the proportion of time spent near the old and new objects by the 16/6-Id (gray bars) and IgG control (black bars) injected mice. The control mice (IgG) significantly preferred the new object (64% vs. 36% for the proportion time near the new vs. old objects respectively;P= 0.01), while the 16/6-Id injected mice had no significant preference to either objects (56% vs. 44% new vs. old;P= 0.5). Results presented as mean ± SEM. * Statistically significant (P <0.05).
[image:4.595.59.290.475.635.2]in microglial activation staining was not seen in the neucortex and piriform cortex, between 16/6-Id and control-IgG mice. Increased staining for astrocytes (GFAP staining) was also noted in the CA3 hippocampal region in the 16/6-Id injected mice compared to controls (Figure 4).
Discussion
In the present study we have observed that passive transfer of 16/6-Id antibodies directly to mice brains resulted in a selective cognitive impairment, expressed as visual recognition and spatial memory deficits. De-pressive behavior (FST) and locomotor activity (staircase test) were not altered in the 16/6-Id injected mice, when compared to the control group. Our findings suggest that 16/6-Id antibodies may have a role in the pathogen-esis of cognitive impairment observed in some patients with SLE [8].
Immunostaining of brain sections from both groups revealed increased presence of activated microglia and astrocytes, in the hippocampal region of the 16/6-Id injected mice, compared to the controls. The hippocam-pus has an important function in memory processing, therefore, its damage by an inflammatory processes may affect cognitive performance in the 16/6-Id injected mice. Astrocytes in steady state conditions are mainly responsible for biochemical support and several other
[image:5.595.57.540.90.352.2]blood–brain barrier, facilitating entry into the brain by in-flammatory factors, including circulating cells of the im-mune system, cytokines, imim-mune-complex mediated small vessel inflammation, and complement components. The inflammatory reaction may induce cognitive changes ob-served in the injected mice.
We have extensively studied the pathogenesis of differ-ent autoantibodies and their influence on the brain. In-jection of anti-ribosomal-P antibodies ICV to naïve mice resulted in depressive-like behavior in these mice [42,43]. In another study, we found that injection of antiphospholipid syndrome patients with antibodies in-duced memory deficits and hyperactivity [15,44]. This suggests that a certain antibody is linked with each spe-cific disease manifestation. The presence of numerous autoantibodies, at least 174 in SLE and 20 in NPSLE, which might have a role in the mechanism of the disease were reported during the past years [11,45]. This may explain the diversity of 19 neuropsychiatric manifesta-tions which can be demonstrated in more than 50% of SLE patients [46]. We propose a hypothesis, that in NPSLE patients different manifestations are the result of an interplay among various auto-antibodies and genetic and environmental factors. For this process to occur, auto-antibodies produced in the body must be able to cross the blood–brain barrier (BBB). It is presumed that the BBB can become transiently“unlocked”following an inflammatory insult, an immune complex damage or ex-posure to infectious endotoxins (for example, lipopoly-saccharide, LPS), allowing antibody penetration. In addition, different auto-antibodies may attach to differ-ent epitopes, expressed unevenly in differdiffer-ent brain areas or neuronal networks. In the studies of Diamond et al., anti-DNA antibodies which can cross-react with the NR2 - anti-NMDA receptor were found in the sera, CSF and brains of SLE patients [16,47]. These antibodies were shown to alter brain cell function and to mediate apoptotic death in vivo and in vitro [16,47]. In their
experiments, the BBB was breached temporarily by in-jection with LPS to imitate an infection [48], while others used noradrenalin to imitate a stressful condition; both conditions were implicated in triggering disease flare-ups in SLE and NPSLE patients. The studies of Diamond et al. added another layer to the current un-derstandings regarding the role of different auto-antibodies in the pathogenesis of NPSLE. Another technique to by-pass the BBB was used by us in several experiments. In the ICV technique, antibodies were injected directly into the lateral ventricle in the mouse brain, allowing antibody dis-persal throughout the brain tissue. In our previous studies, an experimental NPSLE was induced by passive transfer of anti-ribosomal-P antibodies directly to mice brains [43]. The intra-cerebra-ventricularly injected mice exhibited a depression-like behavior, not associated with motor or cog-nitive deficits, and was significantly attenuated by prolong treatment with an anti-depressant (fluoxetine), but not with anti-psychotic drug (haloperidol). Interestingly, the anti-ribosomal-P antibody specifically stained neurons which are related to limbic and olfactory brain areas: the hippo-campus, cingulate cortex and the primary olfactory piriform cortex [43]. The depressed mice also exhibited a decreased smell threshold capability [42], as well as olfactory and lim-bic imaging alterations, when manganese-enhanced-mag-netic resonance imaging (MRI) was performed [49].
[image:6.595.60.540.90.224.2]Another issue of this puzzle was stressed almost two de-cades ago when the importance of the idiotypic network in the induction of various autoimmune diseases was ac-knowledged [22,50]. One proposed mechanism of action of the 16 /6-Id isviathe idiotypic network, in which injec-tion of human anti-DNA 16/6-Id mAbs induces the generation of anti-Id, and anti-anti-Id, and so on. The pro-duction of 16/6-Id was found to be induced also by several infectious agents (for example, Klebsiella pneumonia [51,52] and Mycobacterium tuberculosis [53]); this could point to the role of infections in initiating the disease in a genetically susceptible individual [54].
The finding, that 16/6-Id antibodies were detected in other autoimmune diseases, such as PM/DM and sclero-derma without them expressing central nervous symp-toms is interesting. Perhaps, in some diseases (for example, SLE) a variety of systemic factors enable the al-tering of BBB permeability. These factors may include other circulating antibodies, inflammatory elements, as well as vasogenic agents, growth factors and free radicals. This phenomenon is not unusual in the autoimmunity field, for instance, anti-Ro antibodies are associated with myositis or sub-acute skin manifestations in some SLE pa-tients and not in Sjogren papa-tients.
The current finding, that the 16/6-Id is related to spatial novelty and visual recognition memory impair-ments in mice, may attest for immune-mediated damage to brain areas relevant for these functions. There is a wide agreement that spatial long-term memory and ob-ject recognition is dependent on the functioning of the hippocampal region [55]. Taken together, these concepts may promote the idea for a treatment for NPSLE via blocking or inhibiting the 16/6-Id. This can be done per-haps by treatment with intravenous gamma-globulin, which harbors anti-idiotypic antibodies itself, and has shown some efficacy in the treatment of NPSLE patients [56]. Other therapeutic means may involve the utilization of inhibitory peptides based on the complementarity de-termining region of anti-DNA antibodies. Indeed, such a peptide was shown to be effective in animal models and in a limited number of lupus patients [57-59].
Conclusions
Passive transfer of anti-DNA 16/6-idiotype directly to mice brains resulted in cognitive impairment, supported by cognitive testing impairments, and changes in brain histological analysis. Therefore, the 16/6-idiotype may have a role in cognitive decline, as well as other neuro-psychiatric manifestations, which are found in lupus patients.
Abbreviations
anti-ssDNA: Anti-single-stranded-DNA; BBB: Blood–brain barrier; CNS: Central nervous system; FST: Forced swimming test; ICV: Intra-cerebra-ventricularly; i. p: Intra-peritoneal; LPS: Lipopolysaccharide; mAb: Monoclonal antibody; MRI: Magnetic resonance imaging; NMDA: N-methyl-D-aspartate; NPSLE: Neuropsychiatric systemic lupus erythematosus; PBS: Phosphate buffered saline; PFA: Paraformaldehyde; SLE: Systemic lupus erythematosus; 16/6-Id: 16/6-idiotype.
Competing interests
The authors declare that they have no competing interests.
Authors’contributions
SK participated in the immunohistochemical and behavioral studies and drafted the manuscript. AK participated in the immunohistochemical and behavioral studies and helped to draft the manuscript. MTA participated in the immunohistochemical studies and performed the statistical analysis. MLR and YZ participated in the behavioral studies. NAL participated in the behavioral studies and helped to draft the manuscript. MB participated in the design and helped to draft the manuscript. JMA participated in the
design and coordination of the study. EM and JC participated in the design and coordination of the study and helped to draft the manuscript. YS conceived of the study, participated in its design and coordination, and helped to draft the manuscript. All authors read and approved the final manuscript.
Acknowledgments
We would like to thank Itzik Sehayek for his substantial contribution to this research.
Author details
1
The Zabludovicz Center for Autoimmune Diseases, Sheba Medical Center, Tel-Hashomer, 2 Derech Sheba St., Ramat Gan 52621, Israel.2Rheumatic
Disease Unit, Sheba Medical Center, Tel-Hashomer, 2 Derech Sheba St., Ramat Gan 52621, Israel.3Department of Neurology, Sagol Neuroscience
Center, Sheba Medical Center, Tel-Hashomer, 2 Derech Sheba St., Ramat Gan 52621, Israel.4Center for Autoimmune Diseases Research (CREA), School of
Medicine and Health Sciences, Universidad del Rosario, Kr 24 N°63C-69, Bogotá, DC 111221, Colombia.5Doctoral Program in Biomedical Sciences,
Universidad del Rosario, Kr 24 N°63C-69, Bogotá, DC 111221, Colombia.
6Department of Immunology, The Weizmann Institute of Science, 234 Herzl
Street, Rehovot 76100, Israel.7Incumbent of the Laura Schwarz-Kip Chair for Research of Autoimmune Diseases, Sackler Faculty of Medicine, Tel-Aviv University, Haim Lebanon St., Ramat Aviv 69978, Israel.
Received: 28 September 2012 Accepted: 13 December 2012 Published: 4 April 2013
References
1. Ainiala H, Loukkola J, Peltola J, Korpela M, Hietaharju A:The prevalence of neuropsychiatric syndromes in systemic lupus erythematosus.Neurology
2001,57:496–500.
2. Brey RL, Holliday SL, Saklad AR, Navarrete MG, Hermosillo-Romo D, Stallworth CL, Valdez CR, Escalante A, del Rincon I, Gronseth G, Rhine CB, Padilla P, McGlasson D:Neuropsychiatric syndromes in lupus: prevalence using standardized definitions.Neurology2002,58:1214–1220.
3. Lapteva L, Nowak M, Yarboro CH, Takada K, Roebuck-Spencer T, Weickert T, Bleiberg J, Rosenstein D, Pao M, Patronas N, Steele S, Manzano M, van der Veen JW, Lipsky PE, Marenco S, Wesley R, Volpe B, Diamond B, Illei GG: Anti-N-methyl-D-aspartate receptor antibodies, cognitive dysfunction, and depression in systemic lupus erythematosus.Arthritis Rheum2006,
54:2505–2514.
4. Greenwood DL, Gitlits VM, Alderuccio F, Sentry JW, Toh BH:Autoantibodies in neuropsychiatric lupus.Autoimmunity2002,35:79–86.
5. Borchers AT, Aoki CA, Naguwa SM, Keen CL, Shoenfeld Y, Gershwin ME:
Neuropsychiatric features of systemic lupus erythematosus.Autoimmun Rev2005,4:329–344.
6. The American College of Rheumatology nomenclature and case definitions for neuropsychiatric lupus syndromes.Arthritis Rheum1999,
42:599–608.
7. Rekvig OP, Putterman C, Casu C, Gao HX, Ghirardello A, Mortensen ES, Tincani A, Doria A:Autoantibodies in lupus: culprits or passive bystanders?Autoimmun Rev2012,11:596–603.
8. Kozora E, Hanly JG, Lapteva L, Filley CM:Cognitive dysfunction in systemic lupus erythematosus: past, present, and future.Arthritis Rheum2008,
58:3286–3298.
9. Kapadia M, Sakic B:Autoimmune and inflammatory mechanisms of CNS damage.Prog Neurobiol2011,95:301–333.
10. Oikonomou KG, Zachou K, Dalekos GN:Alpha-actinin: a multidisciplinary protein with important role in B-cell driven autoimmunity.Autoimmun Rev2011,10:389–396.
11. Zandman-Goddard G, Chapman J, Shoenfeld Y:Autoantibodies involved in neuropsychiatric SLE and antiphospholipid syndrome.Semin Arthritis Rheum2007,36:297–315.
12. Weiner SM, Klein R, Berg PA:A longitudinal study of autoantibodies against central nervous system tissue and gangliosides in connective tissue diseases.Rheumatol Int2000,19:83–88.
13. Bonfa E, Golombek SJ, Kaufman LD, Skelly S, Weissbach H, Brot N, Elkon KB:
14. Toubi E, Shoenfeld Y:Clinical and biological aspects of anti-P-ribosomal protein autoantibodies.Autoimmun Rev2007,6:119–125.
15. Katzav A, Pick CG, Korczyn AD, Oest E, Blank M, Shoenfeld Y, Chapman J:
Hyperactivity in a mouse model of the antiphospholipid syndrome.
Lupus2001,10:496–499.
16. DeGiorgio LA, Konstantinov KN, Lee SC, Hardin JA, Volpe BT, Diamond B:A subset of lupus anti-DNA antibodies cross-reacts with the NR2 glutamate receptor in systemic lupus erythematosus.Nat Med2001,
7:1189–1193.
17. Shoenfeld Y, Isenberg DA, Rauch J, Madaio MP, Stollar BD, Schwartz RS:
Idiotypic cross-reactions of monoclonal human lupus autoantibodies.
J Exp Med1983,158:718–730.
18. Shoenfeld Y, Hsu-Lin SC, Gabriels JE, Silberstein LE, Furie BC, Furie B, Stollar BD, Schwartz RS:Production of autoantibodies by human-human hybridomas.J Clin Invest1982,70:205–208.
19. Shoenfeld Y, Rauch J, Massicotte H, Datta SK, Andre-Schwartz J, Stollar BD, Schwartz RS:Polyspecificity of monoclonal lupus autoantibodies produced by human-human hybridomas.N Engl J Med1983,308:414–420. 20. Andre-Schwartz J, Datta SK, Shoenfeld Y, Isenberg DA, Stollar BD, Schwartz
RS:Binding of cytoskeletal proteins by monoclonal anti-DNA lupus autoantibodies.Clin Immunol Immunopathol1984,31:261–271. 21. Amital-Teplizki H, Avinoach I, Coates AR, Kooperman O, Blank M, Shoenfeld
Y:Binding of monoclonal anti-DNA and anti-TB glycolipids to brain tissue.Autoimmunity1989,4:277–287.
22. Blank M, Shoenfeld Y:The story of the 16/6 idiotype and systemic lupus erythematosus.Isr Med Assoc J2008,10:37–39.
23. Isenberg DA, Shoenfeld Y, Madaio MP, Rauch J, Reichlin M, Stollar BD, Schwartz RS:Anti-DNA antibody idiotypes in systemic lupus erythematosus.Lancet1984,2:417–422.
24. Galeazzi M, Bellisai F, Sebastiani GD, Morozzi G, Marcolongo R, Houssiau F, Cervera R, Levy Y, George J, Sherer Y, Shoenfeld Y:Association of 16/6 and SA1 anti-DNA idiotypes with anticardiolipin antibodies and clinical manifestations in a large cohort of SLE patients. European Concerted Action on the Immunogenetics of SLE.Clin Exp Rheumatol1998,
16:717–720.
25. Amital-Teplizki H, Bearman JE, Miele PW Jr, Sukenik S, Shoenfeld Y:A multidimensional autoantibody analysis specifying systemic lupus erythematosus patients with neuropsychiatric symptomatology.Isr J Med Sci1992,28:422–427.
26. Isenberg DA, Collins C:Detection of cross-reactive anti-DNA antibody idiotypes on renal tissue-bound immunoglobulins from lupus patients.
J Clin Invest1985,76:287–294.
27. Isenberg D, Dudeney C, Wojnaruska F, Bhogal BS, Rauch J, Schattner A, Naparstek Y, Duggan D:Detection of cross reactive anti-DNA antibody idiotypes on tissue-bound immunoglobulins from skin biopsies of lupus patients.J Immunol1985,135:261–264.
28. Konikoff F, Isenberg DA, Kooperman O, Kennedy RC, Rauch J, Theodor E, Shoenfeld Y:Common lupus anti-DNA antibody idiotypes in chronic liver diseases.Clin Immunol Immunopathol1987,43:265–272.
29. Shoenfeld Y, Livne A, Blank M, Argov S, Krup M, Fleishmakher E, Sukenik S, Teplizki H:A human monoclonal anti-DNA antibody derived from a patient with polymyositis having the common lupus 16/6 idiotype.
Immunol Lett1988,19:77–83.
30. Mendlovic S, Brocke S, Shoenfeld Y, Ben-Bassat M, Meshorer A, Bakimer R, Mozes E:Induction of a systemic lupus erythematosus-like disease in mice by a common human anti-DNA idiotype.Proc Natl Acad Sci U S A
1988,85:2260–2264.
31. Shoenfeld Y, Mozes E:Pathogenic idiotypes of autoantibodies in autoimmunity: lessons from new experimental models of SLE.FASEB J
1990,4:2646–2651.
32. Shoenfeld Y:Idiotypic induction of autoimmunity: a new aspect of the idiotypic network.FASEB J1994,8:1296–1301.
33. Waisman A, Shoenfeld Y, Blank M, Ruiz PJ, Mozes E:The pathogenic human monoclonal anti-DNA that induces experimental systemic lupus erythematosus in mice is encoded by a VH4 gene segment.Int Immunol
1995,7:689–696.
34. Shoenfeld Y, Nahum A, Korczyn AD, Dano M, Rabinowitz R, Beilin O, Pick CG, Leider-Trejo L, Kalashnikova L, Blank M, Chapman J:Neuronal-binding antibodies from patients with antiphospholipid syndrome induce cognitive deficits following intrathecal passive transfer.Lupus2003,
12:436–442.
35. Porsolt RD, Bertin A, Jalfre M:Behavioral despair in mice: a primary screening test for antidepressants.Arch Int Pharmacodyn Ther1977,
229:327–336.
36. Tordera RM, Totterdell S, Wojcik SM, Brose N, Elizalde N, Lasheras B, Del Rio J:Enhanced anxiety, depressive-like behaviour and impaired recognition memory in mice with reduced expression of the vesicular glutamate transporter 1 (VGLUT1).Eur J Neurosci2007,25:281–290.
37. Dix SL, Aggleton JP:Extending the spontaneous preference test of recognition: evidence of object-location and object-context recognition.
Behav Brain Res1999,99:191–200.
38. Berry A, Vacirca D, Capoccia S, Bellisario V, Malorni W, Ortona E, Cirulli F:
Anti-ATP synthase autoantibodies induce neuronal death by apoptosis and impair cognitive performance in C57BL/6 mice.J Alzheimers Dis2013,
33:317–321.
39. Sun KH, Tang SJ, Lin ML, Wang YS, Sun GH, Liu WT:Monoclonal antibodies against human ribosomal P proteins penetrate into living cells and cause apoptosis of Jurkat T cells in culture.Rheumatology (Oxford)2001,
40:750–756.
40. Koscec M, Koren E, Wolfson-Reichlin M, Fugate RD, Trieu E, Targoff IN, Reichlin M:Autoantibodies to ribosomal P proteins penetrate into live hepatocytes and cause cellular dysfunction in culture.J Immunol1997,
159:2033–2041.
41. Matus S, Burgos PV, Bravo-Zehnder M, Kraft R, Porras OH, Farias P, Barros LF, Torrealba F, Massardo L, Jacobelli S, Gonzalez A:Antiribosomal-P autoantibodies from psychiatric lupus target a novel neuronal surface protein causing calcium influx and apoptosis.J Exp Med2007,
204:3221–3234.
42. Katzav A, Ben-Ziv T, Chapman J, Blank M, Reichlin M, Shoenfeld Y:Anti-P ribosomal antibodies induce defect in smell capability in a model of CNS-SLE (depression).J Autoimmun2008,31:393–398.
43. Katzav A, Solodeev I, Brodsky O, Chapman J, Pick CG, Blank M, Zhang W, Reichlin M, Shoenfeld Y:Induction of autoimmune depression in mice by anti-ribosomal P antibodies via the limbic system.Arthritis Rheum2007,
56:938–948.
44. Menachem A, Chapman J, Katzav A:Hyperactivity induced by antiphospholipid syndrome serum.Ann N Y Acad Sci2009,
1173:422–426.
45. Sherer Y, Gorstein A, Fritzler MJ, Shoenfeld Y:Autoantibody explosion in systemic lupus erythematosus: more than 100 different antibodies found in SLE patients.Semin Arthritis Rheum2004,34:501–537.
46. Unterman A, Nolte JE, Boaz M, Abady M, Shoenfeld Y, Zandman-Goddard G:
Neuropsychiatric syndromes in systemic lupus erythematosus: a meta-analysis.Semin Arthritis Rheum2010,41:1–11.
47. Faust TW, Chang EH, Kowal C, Berlin R, Gazaryan IG, Bertini E, Zhang J, Sanchez-Guerrero J, Fragoso-Loyo HE, Volpe BT, Diamond B, Huerta PT:
Neurotoxic lupus autoantibodies alter brain function through two distinct mechanisms.Proc Natl Acad Sci U S A2010,107:18569–18574. 48. Kowal C, DeGiorgio LA, Nakaoka T, Hetherington H, Huerta PT, Diamond B,
Volpe BT:Cognition and immunity; antibody impairs memory.Immunity
2004,21:179–188.
49. Kivity S, Tsarfaty G, Agmon-Levin N, Blank M, Manor D, Konen E, Chapman J, Reichlin M, Wasson C, Shoenfeld Y, Kushnir T:Abnormal olfactory function demonstrated by manganese-enhanced MRI in mice with experimental neuropsychiatric lupus.Ann N Y Acad Sci2010,1193:70–77.
50. Shoenfeld Y, Ben-Yehuda O, Messinger Y, Bentwitch Z, Rauch J, Isenberg DI, Gadoth N:Autoimmune diseases other than lupus share common anti-DNA idiotypes.Immunol Lett1988,17:285–291.
51. Isenberg DA, Shoenfeld Y:An analysis of autoimmunity through studies of DNA antibody idiotypes.Autoimmunity1988,1:67–75.
52. El-Roiey A, Gross WL, Luedemann J, Isenberg DA, Shoenfeld Y:Preferential secretion of a common DNA idiotype (16/6 Id) and anti-polynucleotide antibodies by normal mononuclear cells following stimulation with Klebsiella pneumoniae.Immunol Lett1986,12:313–319. 53. Shoenfeld Y, Vilner Y, Coates AR, Rauch J, Lavie G, Shaul D, Pinkhas J:
Monoclonal anti-tuberculosis antibodies react with DNA, and monoclonal anti-DNA autoantibodies react withMycobacterium tuberculosis.Clin Exp Immunol1986,66:255–261.
54. Kivity S, Agmon-Levin N, Blank M, Shoenfeld Y:Infections and autoimmunity–friends or foes?Trends Immunol2009,30:409–414. 55. Broadbent NJ, Squire LR, Clark RE:Spatial memory, recognition memory,
56. Milstone AM, Meyers K, Elia J:Treatment of acute neuropsychiatric lupus with intravenous immunoglobulin (IVIG): a case report and review of the literature.Clin Rheumatol2005,24:394–397.
57. Mozes E, Sharabi A:A novel tolerogenic peptide, hCDR1, for the specific treatment of systemic lupus erythematosus.Autoimmun Rev2010,
10:22–26.
58. Lapter S, Marom A, Meshorer A, Elmann A, Sharabi A, Vadai E, Neufeld A, Sztainberg Y, Gil S, Getselter D, Chen A, Mozes E:Amelioration of brain pathology and behavioral dysfunction in mice with lupus following treatment with a tolerogenic peptide.Arthritis Rheum2009,60:3744–3754. 59. Sthoeger ZM, Sharabi A, Molad Y, Asher I, Zinger H, Dayan M, Mozes E:
Treatment of lupus patients with a tolerogenic peptide, hCDR1 (Edratide): immunomodulation of gene expression.J Autoimmun2009,
33:77–82.
doi:10.1186/1741-7015-11-90
Cite this article as:Kivityet al.:16/6-idiotype expressing antibodies induce brain inflammation and cognitive impairment in mice: the mosaic of central nervous system involvement in lupus.BMC Medicine
201311:90.
Submit your next manuscript to BioMed Central and take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution