iii
Dedicatoria
A mis hijos Joel y Gabriela,
Los amores más grandes de mi vida.
En memoria de mis padres Otilia y Santiago,
por su gran amor y ejemplo de dedicación y
trabajo.
iv
Agradecimientos
A mis asesores: Dr. Fausto Cueva Cueva (†)
y Manuel Fukushima Nagaoka, por sus sabios
consejos y asesoramiento en el diseño y ejecución
de la tesis.
A los profesores Walter Reyes Avalos y Zulita
Prieto Lara, por su apoyo y estímulo constante
en la ejecución y culminación de la tesis.
v
PRESENTACIÓN
SEÑORES MIEMBROS DEL JURADO:
Dando cumplimiento a las disposiciones legales vigentes
contenidas en el Reglamento de Grados y Títulos de la
Escuela de Postgrado de la Universidad Nacional de
Trujillo, someto a vuestra consideración el presente
informe de tesis:
Efecto del nivel de proteína dietaria
sobre la respuesta inmunológica, ganancia de peso y
supervivencia
de
Cryphiops
caementarius
,
en
condiciones de laboratorio
, con el propósito de
obtener el
Grado de Doctora en Ciencias Biológicas.
Expreso a ustedes mi sincero agradecimiento por la
revisión
del
presente
informe
y
sus
valiosas
contribuciones.
Trujillo, febrero de 2011
vi
MIEMBROS DEL JURADO DICTAMINADOR
---
Dr. FEDERICO GONZALES VEINTEMILLA
Presidente
---
Dr. ANDRÉS RODRIGUEZ CASTILLO
Secretario
---
Dr. MANUEL FUKUSHIMA NAGAOKA
vii
INDICE
Pág.
DEDICATORIA…. ……….. iii
AGRADECIMIENTOS………. iv
PRESENTACIÓN…..………... v
MIEMBROS DEL JURADO DICTAMINADOR……… vi
INDICE……….. vii
INDICE DE TABLAS………..…. viii
INDICE DE FIGURAS………. ix
RESUMEN ………... x
ABSTRACT ……….. xi
I. INTRODUCCIÓN……… 1
II. MATERIAL Y MÉTODOS……… 10
2.1 Material biológico..………... 10
2.2 Preparación de las dietas …………..………... 11
2.3 Determinación de los parámetros de la calidad de agua ..……… 12
2.4 Extracción de la hemolinfa ………. 12
2.5 Determinación de la hemocianina……… 12
2.6 Conteo de hemocitos……….... 13
2.7 Actividad fenoloxidasa………... 13
2.8 Determinación de proteínas totales………... 13
2.9 Ganancia de peso y supervivencia……… 14
2.10 Análisis de los resultados……… 14
III. RESULTADOS………... 15
IV. DISCUSIÓN……… 25
V. CONCLUSIONES……….. 35
PROPUESTA……….… 37
VI. REFERENCIAS BIBLIOGRÁFICAS……… 38
viii
INDICE DE TABLAS
Pág.
TABLA 1. Composición de las dietas experimentales..………... 11
TABLA 2. Parámetros fisicoquímicos del agua de cultivo de Cryphiops
caementarius según nivel de proteína dietaria………. 16
TABLA 3. Número Total (NTH) y Diferencial (NDH) de hemocitos según los niveles proteicos de las dietas en la hemolinfa de Cryphiops
caementarius………... 19
TABLA 4. Ganancia de peso y supervivencia de Cryphiops caementarius al término del período experimental de 50 días, alimentados con
ix
INDICE DE FIGURAS
PágFIGURA 1. Variación de la temperatura del agua de los acuarios por
semanas de cultivo…..……… 16
FIGURA 2. Efecto del nivel de proteína dietaria (%) sobre la concentración de hemocianina de Cryphiops caementarius alimentado durante
50 días……….. 17
FIGURA 3. Número Total de Hemocitos (NTH) en la hemolinfa de Cryphiops
caementarius, según nivel de proteína dietaria proporcionada
durante 50 días………... 19
FIGURA 4. Número Diferencial de Hemocitos (NDH) en la hemolinfa de Cryphiops caementarius, según nivel de proteína dietaria
proporcionada durante 50 días……… 20
FIGURA 5. Efecto del nivel de proteína dietaria (%) sobre la actividad fenoloxidasa de Cryphiops caementarius alimentado durante
50 días ……….………...……… 21
FIGURA 6. Efecto del nivel de proteína dietaria (%) sobre la concentración de proteínas totales de Cryphiops caementarius alimentado durante 50 días……….. 22
FIGURA 7. Efecto del nivel de proteína dietaria sobre la ganancia de peso de
Cryphiops caementarius durante 50 días de experimentación…. 23
FIGURA 8. Efecto del nivel de proteína dietaria sobre la supervivencia
x
RESUMEN
En el presente trabajo, se evaluó el efecto del nivel de proteína dietaria sobre la respuesta inmunológica (hemocianina, hemocitos, fenoloxidasa y proteínas totales), ganancia de peso y supervivencia del camarón de río Cryphiops
caementarius. Para lo cual, camarones de peso y talla promedio 1, 90 ± 0,76g y
3,90 ± 0,47cm, fueron alimentados con tres dietas con concentración creciente de proteína total (10%, 30% y 50%) durante 50 días. Los camarones fueron distribuidos en 9 acuarios con 10 ejemplares en cada uno y alimentados dos veces al día considerando el 2% de su biomasa. Los análisis se realizaron en la hemolinfa, extraída del seno pericárdico y sólo se utilizaron camarones en estadio de intermuda (estadio C). Se observó un incremento significativo en la concentración de hemocianina en función al incremento del nivel de proteína dietaria (10%, 30% y 50%), con valores de 0,238 ± 0,111 mmol l-1; 0,297 ± 0,099 mmol l-1 y 0,450 ± 0,139 mmol l-1 respectivamente, lo cual indica que el camarón acumuló proteína como hemocianina. El número total de hemocitos (NTH) según los niveles proteicos de las dietas fue de 26,828 ± 12,370 x 105 cél.ml-1; 39,700 ± 26,177 x 105 cél.ml-1 y 41,131 ± 20,976 x 105 cél.ml-1 respectivamente. De igual forma, hubo un incremento en el número diferencial (NDH) de granulocitos, semigranulocitos y hialinocitos, aun cuando no hubo diferencias significativas entre los tratamientos (P > 0,05). Una disminución en la actividad fenoloxidasa y el contenido de proteínas totales en la hemolinfa se observó cuando el camarón fue alimentado con 10% de proteína en la dieta (P < 0,05). En todos los parámetros inmunológicos evaluados hubo diferencias significativas con respecto a los datos iniciales (T0). La ganancia en peso osciló entre 0,012 y 0,800 gramos y la supervivencia entre 76,66% y 86,66%. Estos resultados indican una estrecha relación entre el nivel de proteína dietaria y los parámetros evaluados.
xi
ABSTRACT
The presente study was designed to evaluate the effect of dietary protein level on immunological response (hemocyanin, hemocytes, phenoloxidase and total proteins) weight gain and survival of river prawn Cryphiops caementarius.
Prawns with average weight and height of 1,90 ± 0,76g and 3,90 ± 0,47cm were fed with three protein levels (10%, 30% and 50%). Prawns were distributed in nine tanks with ten specimens each and fed twice a day considering the 2% of their biomass during 50 days. Analysis were performed on hemolymph extracted from the pericardial sinus and only prawns in intermolt stage (C stage) were used. An increase in the hemocyanin concentration was observed with increasing dietary protein level (10%, 30% and 50%), with values of 0,238 ± 0,111 mmol l-1; 0,297 ± 0,099 mmol l-1 and 0,450 ± 0,139 mmol l-1 respectively, indicating that prawn accumulated protein as hemocyanin. The total number (NTH) hemocytes by protein levels was 26,828 ± 12,370 x 105 cél.ml-1; 39,700 ± 26,177 x 105 cél.ml-1 and 41,131 ± 20,976 x 105 cél.ml-1 respectively . Similarly there was an increase in the differential number hemocytes (NDH) even if these were not significantly different between treatments (P> 0, 05). A reduction of phenoloxidase activity and total proteins was observed when prawns were fed with dietary protein level 10%.In relation to the initial data (T0), it was observed significant differences in all immunological parameters evaluated. Weight gain ranging between 0,012 y 0,800 g and survival between 76, 66% y 86, 66%. These results show a near relation between the dietary protein level and the evaluated parameters.
1
I. INTRODUCCION
Uno de los grandes problemas que afronta la humanidad es la escasez de alimentos, especialmente de aquellos ricos en proteínas de buena calidad. Esta situación, se agudizará en el futuro debido a razones tales como el crecimiento poblacional, disminución de tierras de cultivo, cambios climatológicos, entre otros.
En este contexto, la acuicultura se ha convertido en una actividad muy prometedora en la producción de alimentos. En la década de los 1990s fue una de las industrias que mayor crecimiento tuvo en el mundo; reportándose en el año 1997 una producción total de productos acuícolas, incluido algas, de 766 000 TM. De las cuales 150 000 correspondieron a Sudamérica (De Silva, 2000).
2
El mayor crecimiento de esta industria fue durante los años 1980, pero la producción fue seriamente afectada por problemas ligados a degradación del ambiente y a enfermedades infecciosas y no infecciosas (Bachere, 2000), lo que trajo consigo pérdidas económicas significativas. En Asia, la mortalidad del camarón cultivado debido al síndrome del virus de la mancha blanca (WSSV) y del virus cabeza amarilla (YHV) dejó pérdidas cercanas a $1 billón por año desde 1994 (Lightner et al., 1998). Similarmente en Ecuador – que había llegado a ser el segundo país a nivel mundial con 253 000 toneladas en 1998 y 206 000 toneladas en 1999 - solo el virus del síndrome de Taura (TSV) causó la pérdida de $400 millones en divisas por año (Brock, 1997).
Esto es un ejemplo de los efectos de las enfermedades virales en esta actividad y ponen de manifiesto las vulnerabilidades existentes en este aspecto; por lo que el control de las enfermedades es una prioridad a nivel mundial si se quiere que la producción sea ecológica y económicamente sustentable. Para ello, debe considerarse un equilibrio entre (1) la calidad del ambiente, (2) la prevención de enfermedades mediante el diagnóstico y estudios epidemiológicos de los patógenos y (3) el estado de salud del camarón (Bachere, 2000). Esto significa que la prevención y control de las enfermedades del camarón necesita una investigación integrada en la que se consideren aspectos inmunológicos, ambientales, patológicos, fisiológicos, genéticos y nutricionales.
3
incluye melanización por activación del sistema profenoloxidasa, un proceso de coagulación, fagocitosis, encapsulación de material foráneo, acción antimicrobiana y aglutinación celular (Ratcliffe, 1985 en Chun-Hung et al., 2006). Estos mecanismos inmunitarios celulares y humorales operan de manera conjunta para mantener la integridad del organismo.
La profenoloxidasa es un cimógeno de la fenoloxidasa que cataliza tanto la hidroxilación de los monofenoles como la oxidación de los fenoles a quinonas conduciendo a la síntesis de melanina (Sritunyalucksana y Soderhall, 2000). El sistema puede ser activado por componentes celulares microbianos tales como – 1,3-glucanos, lipopolisacáridos (LPS) y peptidoglicanos (PG) por lo que se considera parte importante del proceso de inmunoreconocimiento del mecanismo de defensa. En los crustáceos, se ha demostrado que este sistema se halla en los gránulos de los hemocitos (Ashida, 1984 y 1997 en Chun –Hung et al., 2006), células análogas a los glóbulos blancos y que forman parte del sistema sanguíneo.
4
ocurren cuando los camarones son expuestos a contaminantes ambientales (Le Moullac y Haffner, 2000).
Los aspectos nutricionales también han sido ampliamente estudiados en algunas especies de crustáceos; y en lo que respecta a los requerimientos proteicos varios estudios han evaluado el nivel óptimo de proteína dietaria para obtener un máximo crecimiento y los resultados son diversos. Así, para Litopenaeus
vannamei se encontró un rango óptimo de proteína cruda ente 25% y 35%
(Teichert, 1988 y Cousin, 1995 en Pascual et al., 2004) y según Kureshy (2002) el máximo crecimiento en juveniles de esta misma especie lo obtuvo con porcentajes superiores a 32%, encontrando una mejor eficiencia alimenticia con 48% de proteína dietaria.
En términos de requerimiento absoluto, se ha reportado una tasa de crecimiento máximo, con 43 g de proteína/ kg de peso corporal/ día para juveniles de L.
vannameicon un óptimo sobre 32% (Kureshy, 2002). Cousin (1995) en Pascual
5
encontró mas hemocianina y proteína en los camarones alimentados con bajos niveles de almidón, lo cual demuestra la capacidad de L. vannamei para usar proteína dietaria como fuente de aminoácidos y almacenar proteína en la hemolinfa como hemocianina.
El sistema inmunológico de los camarones tiene una sólida base proteica y la hemocianina juega un rol importante en su funcionamiento. Estudios recientes han demostrado que además de su rol multifuncional (transportador de oxígeno, proteína de almacenamiento, portador de carotenoides, transportador de ecdisona) tiene función de profenoloxidasa (Adachi et al., 2003), propiedades antivirales (Zhang et al., 2004) y acción fungistática (Destoumieux et al., 2001). En el proceso de melanización, una reacción defensiva de los camarones, también están involucradas varias proteínas como la profenoloxidasa, fenoloxidasa, -2
macroglobulina (Sritunyalucksana y Soderhall, 2000; Perazzolo y Barracco, 1997). Otras proteínas involucradas en procesos inmunológicos son las proteínas de reconocimiento celular como las proteínas que se unen a los glucanos y lipopolisacáridos de las bacterias; asimismo los péptidos antimicrobianos llamados peneidinas que actúan sobre un gran número de patógenos, esencialmente sobre bacterias Gram positivas (Destoumieux, 2000 en Pascual et al., 2004).
6
hemocitos ocurre cuando los niveles proteicos son muy bajos, lo que trae consigo una disminución de los cimógenos que contienen, por ejemplo el sistema profenoloxidasa, las peneidinas y su actividad de fagocitosis y coagulación (Pascual et al., 2004).
En lo que respecta al estado fisiológico del camarón, uno de los factores que tiene gran influencia es el nivel de proteína dietaria y las investigaciones se han concentrado básicamente en determinar sus niveles óptimos; no se han hecho estudios, excepto el de Pascual et al. (2004), para evaluar el efecto de la proteína dietaria sobre potenciales marcadores de la respuesta inmune como son los hemocitos y la fenoloxidasa entre otros. A través de la dieta se puede modular el sistema inmunológico, ya que un organismo con deficiencias nutricionales es más susceptible a las enfermedades, por la ausencia de factores necesarios para el sistema de defensa. En peces, se reporta que variaciones en la composición de ácidos grasos o proteínas afectan a la respuesta inmune (Plat, 2005 en Cuesta et al., 2006).
7
Existen diversos trabajos sobre C. caementarius, relacionados con su morfología externa (Chávez et al., 1971), preferencias alimenticias por las microalgas en sus dietas (Norambuena, 1977; Viacava et al., 1978; López et al., 1986), aspectos bioecológicos (Valdeiglesias, 1983), biología reproductiva (Guerra et al., 1983; Reyes et al., 2002; Reyes et al., 2005), efecto de la salinidad en el crecimiento y supervivencia en postlarvas (Reyes et al., 2006). Algunos aspectos metabólicos como el consumo de oxígeno también han sido estudiados y se reporta que es afectado por la temperatura, salinidad y la interacción de ambas variables; asimismo, se señala que la respuesta metabólica a cambios de temperatura determinaría la migración ascendente de la especie en el río (Zuñiga y Ramos, 1990).
En nutrición, se destaca el uso de alimento vivo como microalgas, nauplios de
Artemia salina (Meruane et al., 1996), comunidades epipetrales (Ponce et al.,
1982), el rotífero Brachionus plicatilis (Portugal et al., 2003). Ayvar (1982) evaluó el valor biológico de varias dietas comerciales con diferentes niveles proteicos y encontró que las dietas con 44,9 % de proteína los camarones tuvieron mejor ganancia de peso total y tasa específica de crecimiento, y una supervivencia del 76,92% considerada dentro del rango normal para este tipo de cultivos.
8
producen una mayor ganancia de peso y según Kureshy y Davis (2002), un amplio rango de niveles de proteína dietaria podría ser usado para producir una máxima ganancia de peso en L. vannamei juveniles y subadultos. En lo que respecta a supervivencia, el cuidado es un factor importante; otro es el nutricional, cuya deficiencia afectaría la muda y consecuentemente la supervivencia. Según Infante (1985) la incapacidad para mudar en M. rosenbergii se debe a deficiencias alimentarias, a funciones orgánicas poco activas, a problemas de intoxicaciones o también a factores ambientales.
En los últimos años, la industria del cultivo del camarón ha sido seriamente afectada por la diseminación de diversas enfermedades virales y bacterianas las cuales han provocado importantes pérdidas económicas (Alday – Sanz et al., 1988; Lightner et al., 1998) y uno de los aspectos poco investigados para disminuir el impacto de las enfermedades es el nutricional; específicamente en lo que se refiere a los requerimientos proteicos necesarios para mantener óptimamente el estado fisiológico del camarón incluyendo el sistema inmunológico.
La relación entre proteína dietaria y respuesta inmunológica no ha sido estudiada
en C. caementarius y aún en especies que se cultivan comercialmente existen
9
ganancia de peso y supervivencia de C. caementarius, en condiciones de laboratorio.
10
II. MATERIAL Y MÉTODOS
2.1 Material Biológico
Se utilizaron 90 camarones adultos de la especie C. caementarius (Fig.1. Anexo), con peso promedio de 1,90 ± 0,76 g y longitud total promedio 3,90 ± 0,47 cm (escotadura post orbital – extremo posterior del telson), capturados en la zona baja del río Lacramarca (09º 07’41” LS y 78º 33’30” LW) de la provincia del Santa, Departamento de Ancash.
Los camarones fueron criados durante 50 días en acuarios con un volumen de agua de caño de 70l (10 camarones por acuario), aireada constantemente y alimentados con diferentes niveles de proteína dietaria (10%, 30% y 50%). Tres acuarios fueron aleatoriamente asignados para cada tratamiento (Fig.2 y 3. Anexo). Un grupo de camarones (n=9) al que se le denominó tiempo cero (T0) fueron evaluados (concentración de hemocianina, número de hemocitos, proteínas totales y fenoloxidasa) antes de iniciar el experimento.
11
2.2. Preparación de las dietas
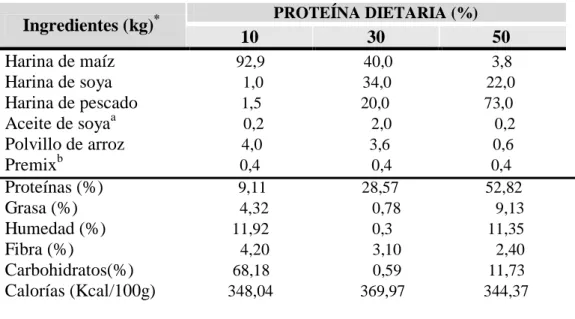
La composición de las dietas ensayadas se muestra en la Tabla 1. Se preparó tres dietas con diferentes niveles proteicos: 10%, 30% y 50%, éstas fueron formuladas empleando el software ALITE (Pezzato, 1996) y preparadas mediante la mezcla de los ingredientes secos, luego se adicionó el aceite, las vitaminas y agua hasta formar una pasta suave. Ésta fue pasada a través de una peletizadora y los spaghetti resultantes fueron secados en estufa a 36 ºC por 48 h. Después de secar, el material fue trozado y pasado a través de una malla de 1 mm de diámetro (Fig.4 y 5. Anexo)
Tabla 1. Composición de las dietas experimentales
Ingredientes (kg)* PROTEÍNA DIETARIA (%) 10 30 50
Harina de maíz 92,9 40,0 3,8 Harina de soya 1,0 34,0 22,0 Harina de pescado 1,5 20,0 73,0 Aceite de soyaa 0,2 2,0 0,2 Polvillo de arroz 4,0 3,6 0,6 Premixb 0,4 0,4 0,4 Proteínas (%) 9,11 28,57 52,82 Grasa (%) 4,32 0,78 9,13 Humedad (%) 11,92 0,3 11,35 Fibra (%) 4,20 3,10 2,40 Carbohidratos(%) 68,18 0,59 11,73 Calorías (Kcal/100g) 348,04 369,97 344,37
a
Litros; b mezcla de vitaminas
*
12
2.3. Determinación de los parámetros de calidad de agua
La calidad de agua de los acuarios se analizó semanalmente determinando oxígeno disuelto, CO2, dureza total, alcalinidad total, pH, nitritos y temperatura
según Fukushima et al. (1982).
2.4. Extracción de la hemolinfa
Se utilizaron sólo camarones en estadío de intermuda C (Reyes y Luján, 2003). La hemolinfa (aproximadamente 100 -150 μl por camarón) fue extraída del seno pericárdico utilizando jeringas descartables de 1 ml conteniendo solución Alsever como anticoagulante (Van de Braak, 2002) (Fig.6. Anexo).
2.5. Determinación de la hemocianina
13
2.6. Conteo de hemocitos
Para el conteo del número total (NTH) y diferencial (NDH) de hemocitos, se midió 20 µl de hemolinfa y se adicionó a un microtubo (Eppendorf) conteniendo 80 µl de solución Alsever (Van de Braak, 2002). El conteo se hizo utilizando una cámara de Neubauer y un microscopio Leica DM LS2 de luz convencional y campo luminoso provisto de contraste de fases. Para determinar el número de hemocitos se utilizó la fórmula de Jussila (1997) y las diferentes clases se reconocieron según Azañero et al. (2006) (Fig 8. Anexo).
2.7. Actividad Fenoloxidasa.
Una muestra de 50 µl de hemolinfa diluida al medio fue incubada por 3 minutos a 25ºC con 50 µl de tripsina tipo IX (0,1 mg/ml). Luego se adicionó 50 µl de L-DOPA (3mg/ml en buffer Cacodilato), seguido de una incubación por 10 minutos. La absorbancia se midió a 490 nm (Hernandez-López et al., 1996).
2.8. Determinación de Proteínas Totales.
14
2.9. Ganancia de peso y supervivencia.
Al final del experimento se pesaron (por desplazamiento de volumen de agua) (Fig. 9. Anexo) y contaron todos los ejemplares a fin de determinar la ganancia en peso y supervivencia.
2.10. Análisis estadístico de los resultados.
15
III.
RESULTADOS
1. Calidad de agua
Las temperaturas promedio semanal para los tres tratamientos empleados mostraron escasa variación. En la figura 1, se observa un incremento a partir de la semana 3, debido a que se colocó una cocina eléctrica en el ambiente a fin de incrementar la temperatura y favorecer el consumo de alimentos artificiales. La temperatura mínima fue de 18,5 º C y la máxima de 23,08 º C; siendo el promedio general de 21,91 ± 1,36 º C.
El promedio de oxígeno disuelto fue de 8,43 ± 0,21 mg l-1, observándose un valor ligeramente bajo en el tratamiento con 30% de proteína dietaria. Los valores de pH para los tratamientos oscilaron entre 7,92 ± 0,07 y 7,98 ± 0,06 lo cual puede considerarse ligeramente alcalino. El contenido de CO2 del agua fue bajo (1,27 ±
0,43). Respecto a la dureza y alcalinidad del agua, los promedios fueron 208,38 ± 10,03 mg l-1 y 58,07 ± 9,08 mg l-1 respectivamente. El contenido promedio de nitritos fue de 13,58 ± 12,52 mg l-1.
16
Fig. 1. Variación de la temperatura del agua de los acuarios por semanas de cultivo de C. caementarius.
Tabla 2. Parámetros fisicoquímicos del agua de cultivo de C. caementarius según nivel de proteína dietaria.
Parámetros
Proteína
Dureza (mg l-1)
Alcalinidad (mg l-1)
CO2
(mg l-1) pH
Oxígeno (mg l-1)
Nitritos (mg l-1)
Temperatura (ºC)
10% 210,8 58,5 1,2 7,98 8,49 0,76 21,5
± 13,76 ± 4,80 ± 0,54 ± 0,06 ± 0,21 ± 0,58 ±1,2
30% 204,75 54,77 1,08 7,92 8,37 15,74 22,1
± 10,31 ± 12,81 ± 0,35 ± 0,07 ± 0,19 ± 12,32 ±1,5
50% 210,5 60,75 1,44 7,93 8,45 16,76 21,8
± 6,99 ± 6,52 ± 0,41 ± 0,11 ± 0,25 ± 13,77 ±1,4
2. Hemocianina
17
en el tratamiento de 10%; 0,297 ± 0,099 mmol l-1 en el de 30% y 0,450 ± 0,139 mmol l-1 en el de 50% (Fig.2) (Tabla 1.Anexo).
Fig. 2. Efecto del nivel de proteína dietaria (%) sobre la concentración de hemocianina de C. caementarius alimentado durante 50 días. Promedio ± D.E. Letras diferentes denotan diferencias estadísticas (p< 0,05).
3. Número total (NTH) y diferencial (NDH) de hemocitos
18
41,131 ± 20,976 x 105 cél.ml-1 respectivamente; se observó un incremento significativo sólo en relación a los datos iniciales (T0) (14,398 ± 5,372 x 105 cél.ml-1), no al nivel de proteína dietaria (Tablas 2, 3, 4 y 5. Anexo). El NDH promedio de granulocitos, semigranulocitos y hialinocitos para el tratamiento con 10 % de proteína dietaria fue de 4,281 ± 2,995 x 105 cél.ml-1; 21,766 ± 11,289 x 105 cél.ml-1 y 0,781 ± 0,556 x 105 cél.ml-1, respectivamente. Para el tratamiento con 30% de proteína dietaria los granulocitos fueron 6,319 ± 4,669 x 105 cél.ml
-1
, los semigranulocitos 32,744 ± 21,859 x 105 cél.ml-1 y los hialinocitos 0,638 ± 0,581 x 105 cél.ml-1. Con 50% de proteína dietaria se encontraron 8,642 ± 5,490 x 105 cél.ml-1 granulocitos; 31,017 ± 16,630 105 cél.ml-1 semigranulocitos y 1,472 ± 1,268 x 105 cél.ml-1 hialinocitos.
19
Tabla 3. Número Total (NTH) y Diferencial (NDH) de Hemocitos según los niveles proteicos de las dietas en la hemolinfa de C. caementarius.
Fig. 3. Número Total de Hemocitos (NTH) en la hemolinfa de C. caementarius
según nivel de proteína dietaria proporcionada durante 50 días. Promedio ± D.E. Letras diferentes denotan diferencias estadísticas (p< 0,05).
Hemocitos
Proteína
Nº Total de Hemocitos (NTH) x 105 cél. ml-1
Nº Diferencial de Hemocitos (NDH) x 105 cél. ml-1
Granulocitos Semigranulocitos Hialinocitos
20
Fig. 4. Número Diferencial de Hemocitos (NDH) en la hemolinfa de C.
caementarius según nivel de proteína dietaria proporcionada durante 50
días. Promedio ± D.E. Letras diferentes denotan diferencias estadísticas (p< 0,05).
4. Actividad Fenoloxidasa.
21
Fig. 5. Efecto del nivel de proteína dietaria (%) sobre la actividad fenoloxidasa de
C. caementarius alimentado durante 50 días. Promedio ± D.E. Letras
diferentes denotan diferencias estadísticas (p< 0,05).
5. Concentración de Proteínas Totales.
22
Fig. 6. Efecto del nivel de proteína dietaria (%) sobre la concentración de proteínas totales de C. caementarius alimentado durante 50 días. Promedio ± D.E. Letras diferentes denotan diferencias estadísticas (p< 0,05).
6. Ganancia de peso
Los pesos promedio iniciales oscilaron entre 1,626 y 1,905 gramos, no habiendo diferencia significativa entre ellos, lo mismo que en los pesos promedio finales que oscilaron entre 1,819 y 2,426 gramos (Tabla 4).
23
Tabla 4. Ganancia en peso y supervivencia de C. caementarius al término del período experimental de 50 días, alimentados con tres niveles de proteína dietaria.
Parámetros
Proteína Dietaria
10% 30% 50% Peso inicial (g ) 1,807 ± 0,103 1,905 ± 0,151 1,626 ± 0,133 Peso final ( g ) 1,819 ± 0,169 2,213 ± 0,257 2,426 ± 0,959 Ganancia de peso 0,012 ± 0,388 0,308 ± 0,629 0,800 ± 1,016 Supervivencia (%) 86,66 86,66 76,66
Fig. 7. Efecto del nivel de proteína dietaria sobre la ganancia de peso de C.
24
7. Supervivencia
Los porcentajes de supervivencia se consignan en la Tabla 4, en ésta se puede apreciar que los tratamientos con 10% y 30% de proteína dietaria son los que mejor supervivencia tienen (86,66%), mientras que la mayor mortalidad se registró en el tratamiento con 50% de proteína dietaria, obteniéndose una supervivencia de 76,66%.
El canibalismo fue la principal causa de mortalidad registrada en todo el experimento para los distintos tratamientos. Éste es practicado principalmente por los animales más grandes, sobre animales que habían mudado recientemente siendo los pleópodos y periópodos los primeros órganos en ser devorados por sus congéneres.
Fig. 8. Efecto del nivel de proteína dietaria sobre la supervivencia de C.
25
IV. DISCUSIÓN
Las características físicas y químicas del agua de cultivo de C. caementarius se encontraron dentro de los rangos normales requeridos para esta especie y reportados por Venturi (1972) y Viacava et al. (1978), por lo que no hubo influencia de la calidad de agua sobre el contenido de hemocianina, hemocitos, actividad fenoloxidasa y proteínas totales en la hemolinfa; asimismo, sobre la ganancia de peso y supervivencia.
Después de 50 días de crianza, la concentración de hemocianina en la hemolinfa
de C. caementarius fue significativamente más alta en los camarones alimentados
con 50 % de de proteína dietaria, comparada a la de camarones alimentados con 30% y 10%. Estos resultados son similares a los reportados para L. vannamei en los que se observó un incremento significativo en la concentración de hemocianina y otros parámetros inmunológicos (número total de hemocitos y explosión respiratoria) en concordancia al incremento del nivel de proteína dietaria (5%, 15% y 40%) (Pascual et al., 2004). Un aumento en hemocianina y proteínas sanguíneas también fue observado en pre-adultos de Litopenaeus
setiferus alimentados con niveles proteicos de 35%, 45% y 55% (Goimier et al.,
26
al., 1980; Pascual et al., 2004), tiene funciones de fenoloxidasa (Adachi et al., 2003; Terwilliger, 2007), acción fungistática (Destoumieux et al., 2001; Adachi et al., 2003) y, recientemente se ha demostrado también actividad hemolítica (Zhang et al., 2009).
La disminución del nivel de proteína en la dieta, repercute no sólo en el crecimiento (Galindo et al., 2003); sino también en las capacidades inmunológicas a través de una reducción de la hemocianina, capacidad fagocítica y concentración de hemocitos, entre otras. En L. vannamei alimentado con 5% de proteína dietaria, un nivel subóptimo, Pascual et al. (2004) reportaron valores significativamente bajos de hemocianina comparados a los de camarones alimentados con 15% de proteína. En el presente estudio, el nivel proteico más bajo fue 10%, y si bien se observa una disminución en la concentración de hemocianina, la diferencia con el siguiente nivel 30% no es significativa; probablemente, debido a que el primero estaría sobre el nivel proteico mínimo requerido para esta especie. En el caso de L.
vannamei, por ejemplo está entre 1,8 y 3,8g de proteína dietaria / kg de peso
corporal/día (Kureshy y Davis, 2002).
27
mejor nivel proteico donde se obtuvo mayor concentración de hemocianina fue 50%, valor cercano al reportado por Ayvar (1982) cuya dieta con 44,9 % de proteína fue la de mayor ganancia de peso total y tasa específica de crecimiento; sin embargo, Venturi (1973) obtuvo buenas tasas específicas de crecimiento con dietas cuya proteína total osciló entre 22% y 34%. Al igual que en los camarones marinos, cuyos niveles óptimos de proteína dietaria son muy variados (Arantakananda y Lawrence, 1993; Kureshy y Davis, 2002), los resultados obtenidos demostrarían que los de agua dulce presentan un comportamiento similar.
Muchos factores celulares y humorales han sido estudiados con el propósito de ser usados como indicadores para medir la eficacia de potenciales inmunoestimulantes (Sritunyalucksana et al., 1999; De Philippis et al., 1998 en Campa – Córdova et al., 2002); sin embargo, hay poco conocimiento acerca de la respuesta celular y humoral de los camarones a estas sustancias.
28
La calidad de la dieta es un factor importante que influye sobre la respuesta inmunológica del camarón, se han hecho estudios utilizando dietas con diferentes concentraciones de proteína y se ha demostrado que una mayor concentración de proteína en la dieta incrementa el número de hemocitos así como también la concentración de proteínas plasmáticas (Rodríguez et al., 2000; Pascual et al., 2004).
Los resultados obtenidos sobre número total de hemocitos (NTH), indican un aumento progresivo en relación al incremento del nivel de proteína dietaria y al T0, con diferencias significativas solamente respecto a este último, más no entre los tratamientos; observándose el valor más bajo en los camarones alimentados con 10% de proteína. Las actividades de los hemocitos como fagocitosis y coagulación (Omori et al., 1989; Gargioni y Barracco, 1998 en Bachere, 2000) también habrían sido las más reducidas en este nivel proteico. Resultados similares fueron reportados por Yaemsooksawat et al. (2009) quienes trabajando con post larvas de L. vannamei no encontraron diferencias significativas en el NTH entre los tratamientos con 36%, 32%, 28% y 24% de proteína en la dieta, sólo un ligero aumento correspondiendo al nivel de proteína dietaria; lo mismo fue reportado por Rodríguez et al. (2000) en esta especie con dietas de 22% y 50% de proteína, asimismo una disminución en el NTH con respecto a los valores obtenidos en el T0.
29
proteína dietaria (Pascual et al., 2004; Goimier et al., 2006). Las diferencias en resultados podrían obedecer a múltiples factores, por ejemplo se menciona que los hemocitos circulantes pueden ser afectados por factores extrínsecos como la temperatura, pH, salinidad, oxígeno disuelto y amonio (Truscott y White, 1990; Le Moullac et al., 1998; Le Moullac y Haffner, 2000; Cheng y chen, 2001); sin embargo, no fue el caso nuestro ya que estos factores estuvieron dentro del rango normal para esta especie (Venturi, 1972; Viacava et al., 1978). Probablemente, el origen de los componentes proteicos (Smith et al., 1985; Mente et al., 2002) y de los otros constituyentes de las dietas así como de las condiciones de cultivo (Pascual et al., 2003) influyan en la variabilidad de los resultados. Asimismo el tamaño de los individuos (Smith et al., 1985), el origen de la muestra si ésta es del medio natural o de granjas, en nuestro caso se extrajo del medio natural y se observó una alta contaminación del río principalmente por desechos de fábricas pesqueras, lo cual facilita la presencia de protozoarios tipo Zoothamnium sp y bacterias como L. mucor (Azañero et al, 2009) que en grandes cantidades compiten por oxígeno con sus hospederos y pueden producir disminución del consumo de alimento (López-Tellez et al., 2003 en Azañero et al., 2009).
30
significancia entre los tratamientos. Similares resultados fueron reportados por Hose et al. (1990) quienes examinaron los hemocitos de crustáceos decápodos y encontraron una amplia variación en la proporción del número diferencial de hemocitos (NDH).
El efecto del nivel de proteína dietaria sobre el NDH ha sido demostrado en L.
vannamei, observándose una reducción en las células granulares y hialinas con la
disminución del nivel de proteína dietaria (Pascual et al., 2004). En C.
caementarius, se observó una tendencia al incremento, en todas las clases de
hemocitos, en relación a un mayor nivel proteico en la dieta; sin embargo, las diferencias no fueron significativas. No obstante, con respecto al T0 tanto los granulocitos como los semigranulocitos mostraron diferencias significativas en los niveles proteicos 30% y 50%. Estos resultados sugieren que de alguna manera sí hubo influencia del nivel de proteína dietaria en la producción de hemocitos. En todos los tratamientos los granulocitos y semigranulocitos se encontraron en mayor proporción que los hialinocitos, lo cual coincide con lo aportado por Azañero, et al. (2006) para esta misma especie.
31
niveles de fenoloxidasa, siendo significativamente más bajos en los camarones alimentados con 10% (P< 0,05). Yaemsooksawat et al. (2009) no encontró variaciones en la actividad fenoloxidasa en postlarvas de L. vannamei
alimentados con niveles proteicos de 24%, 28%, 32% y 36% durante 100 días. Estudios realizados en L. setiferus y L. vannamei demuestran que una disminución del número de hemocitos producida por valores extremos de temperatura o niveles proteicos muy bajos, puede compensarse con un incremento de fenoloxidasa/ célula.
32
en relación a los demás tratamientos, aun cuando no se observó diferencias significativas en relación a este parámetro de calidad de agua.
La ganancia de peso es un parámetro importante porque permite el incremento de biomasa; asimismo, desde el punto de vista comercial o para establecer un plantel de reproductores. Tal como lo reporta Ayvar (1982) las dietas altas en proteínas producen una mayor ganancia de peso. Según Kureshy y Davis (2002), un amplio rango de niveles de proteína dietaria podría ser usado para producir una máxima ganancia de peso en L. vannamei juveniles y subadultos.
33
Chauca (1979), citado por Ayvar (1982), trabajando con raciones alimenticias desde 34,84% a 46,56% de proteína para juveniles de C. caementarius, reportó incrementos de peso altos, entre 172,00% y 236,44%. En general, y tal como sucede en otras especies de camarones los resultados son muy variados aun trabajando con el mismo estado de crecimiento y tal como lo menciona Ayvar (1982) parece que C. caementarius no es una especie muy exigente en proteínas, pudiendo lograrse buenos resultados con niveles bajos de proteína.
Respecto a la supervivencia, se obtuvo un mayor porcentaje en los tratamientos con 10% y 30% de proteína dietaria y el menor con el de 50% (Tabla 4), señalándose al canibalismo como la causa principal de la mortalidad. Chauca (1979) citado por Ayvar (1982) reporta que las mayores mortalidades registradas
en C. caementarius se debieron a las fugas y en menor proporción al canibalismo;
la supervivencia registrada por este autor osciló entre 84% y 96%.
Nuestros resultados coinciden con los obtenidos por Ayvar (1982) en el sentido de que hay una relación inversa en el porcentaje de supervivencia y el nivel de proteínas en la dieta. Este autor obtuvo supervivencias entre 84,62% y 76, 92% para niveles proteicos en la dieta de 34,78% y 55,02% respectivamente, trabajando con juveniles de C. caementarius.
34
(28% y 35%) la mortalidad fue mayor, lo cual obedeció al canibalismo. Algunos estudios no muestran diferencias en el crecimiento y supervivencia de juveniles de
M. rosenbergii en rangos de 15% – 35% de proteína (Boonyaratpalin y New, 1980
en Luna (2007).
Los resultados obtenidos, aun cuando las diferencias encontradas entre los diferentes niveles proteicos utilizados es muy escasa, sugieren que una disminución de las proteínas de la dieta va a producir una reducción de las capacidades inmunológicas, así como en la ganancia de peso de Cryphiops
caementarius. Sin embargo, es necesario realizar más estudios a fin de determinar
35
V.
CONCLUSIONES
1. Los niveles de proteína dietaria 10%, 30% y 50% produjeron cambios significativos en el contenido de hemocianina en la hemolinfa de C.
caementarius, siendo sus valores: 0.238 ± 0.111 mmol/l; 0.297 ± 0.099 mmol/l
y 0.450 ± 0.139 mmol/l, respectivamente.
2. El número total de hemocitos (NTH) incrementó en relación al aumento de proteína dietaria (10%, 30% y 50%), siendo sus valores 26,828 ± 12,370 x 105 cél.ml-1; 39,700 ± 26,177 x 105 cél.ml-1 y 41,131 ± 20,976 x 105 cél.ml-1, respectivamente, estadísticamente diferentes con respecto a los valores iniciales (T0).
3. El número diferencial de hemocitos (NDH): granulocitos, semigranulocitos y hialinocitos no registró diferencias significativas por efecto del nivel de proteína en la dieta.
4. Los valores más bajos de actividad fenoloxidasa y proteínas totales en la hemolinfa se observaron cuando el camarón fue alimentado con 10% de proteína en la dieta.
36
6. Los porcentajes de supervivencia de los diferentes tratamientos oscilaron entre 86,66 y 76,66 %. La mortalidad obedeció principalmente al canibalismo de los animales.
37
PROPUESTA
En los últimos años, la industria del cultivo de camarón ha sido seriamente afectada por la diseminación de diversas enfermedades virales y bacterianas las cuales han provocado importantes pérdidas económicas. Uno de los aspectos poco investigados para disminuir el impacto de las enfermedades es el nutricional; específicamente, en lo que se refiere a los requerimientos proteicos necesarios para mantener óptimamente el estado fisiológico del camarón, incluyendo el sistema inmunológico.
Las proteínas son muy importantes para el funcionamiento de este sistema, puesto que permiten la formación de los componentes humorales como fenoloxidasa, peneidinas, hemocianina y proteínas anticoagulantes. Por ello, en la presente investigación se han evaluado diferentes niveles proteicos y los resultados muestran la importancia de estos nutrientes para la respuesta inmunológica, medida en términos de hemocitos, hemocianina, fenoloxidasa y proteínas totales; asimismo, para la ganancia de peso de Cryphiops caementarius.
Se han hecho algunos estudios; sin embargo, no se han definido los requerimientos nutricionales para la especie; tampoco se conoce sobre enfermedades, por lo que es necesario emprender acciones tendientes a:
Determinar los requerimientos óptimos de proteína dietaria en las diferentes etapas de su desarrollo que le permitan un fortalecimiento de su sistema inmunológico.
Realizar estudios para determinar el tipo de patógenos que afectan la salud del camarón y una vez identificados realizar estudios de desafío previa inmunomodulación a través de las proteínas de la dieta.
Promover la participación de las empresas y las Universidades para realizar investigaciones que generen nuevo conocimiento sobre otros marcadores de la respuesta inmune como son el anión superóxido, actividad antibacteriana entre otros.
Involucrar al Estado no solo en la legislación de su captura sino en la promoción del cultivo de esta especie de gran aceptación en el mercado.
38
VI. REFERENCIAS BIBLIOGRÁFICAS
Adachi, K., T. Hirata, T. Nishioka y M. Sakaguchi. 2003. Hemocyte components in crustaceans convert hemocyanin into a phenoloxidase – like enzyme. Comp. Biochem. Physiol., 134 B: 135 – 141.
Alday – Sanz, V., S.Thaikua, A. Yousif, L. Albright y T. Flegal. 1988. Studies on Ig Y for passive immunization of shrimp against white spot syndrome virus. In: Flegel, T.W. (ED.), Advances in shrimp biotechnology, Center for Genetic Engineering and Biotechnology, Bangkok, pp. 141-143.
Amaya, J. y A. Guerra. 1976. Especies de camarones de los ríos norteños del Perú y su distribución. Ministerio de Pesquería. Dirección de Investigación Científica y Tecnología, 24: 1-60.
Arantakananda, P. y A. Lawrence. 1993. Dietary protein and energy requirement of the white-legged shrimp, Penaeus vannamei and the optimal protein to energy ratio. From Discovery to Commercialization. European Aquaculture Society, Oostende, Belgium 21 pp.
Ayvar, F. 1982. Pruebas Comparativas de raciones balanceadas de diferentes niveles de proteínas en la crianza de camarones de río (Cryphiops
caementarius) en ambientes cerrados. Tesis para optar el Título de Ingeniero
Pesquero. Lima – Perú, 111 pp.
39
(Provincia del Santa, Departamento de Ancash, Perú). Civa 2006 (http:// WWW.civa 2006. org), 1-8.
Azañero, C., W. Reyes, F. Saucedo y R. Rodríguez. 2009. Parásitos asociados al camarón de río Cryphiops caementarius Molina, 1872 (Crustacea, Palaemonidae), procedentes del río Lacramarca (Prov. del Santa, Dpto. Ancash, Perú). Informe final de Proyecto de investigación. Cód. PIC. FF.CC. 030-2008. Universidad Nacional del Santa, Perú.
Bachere, E., 2000. Shrimp immunity and disease control. Aquaculture, 191: 3 – 11.
Berger, C. y O. Rubio. 2005. Cultivo de langostinos en el Perú: una industria
exitosa. Disponible en
URL:http://www.adexperu.org.pe/exposiciones/acuicultura/cultivo_del_lango stino_ALPE.pdf.
Bradford, M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem., 72: 248 – 254.
Brock, J. 1997. Special topic review: Taura síndrome, a disease important to shrimp farms in the Americas. World J. Microbiol & Technol. 13: 415 – 418. Campa-Córdova, A., N. Hernández-Saavedra, R. De Philippis, y F. Ascencio.
40
Cuesta, A., I. Salinas, L. Martínez, F.J. Martínez, J. Meseguer y M. Angeles. 2006. Efecto de diferentes dietas en la respuesta inmunitaria humoral innata de la lubina ( Dicentrarchus labrax L.). En: IV Congreso Iberoamericano Virtual de Acuicultura. CIVA 2006. 2006:171-178. Disponible en URL: http://www.civa2006.org.
Cuzon, G., C. Cahu, F.J. Aldrin, L. Messager, G. Stephan y M. Mével. 1980. Starvation effect on metabolism of Penaeus japonicus. Proc. World Maric. Soc. 11: 410-423.
Chávez, R., E. De Parodi y J. Villegas. 1971. Revista de Investigación de la Universidad Nacional San Agustín de Arequipa, 2 (1): 13 – 34.
Chen, J., C. Chen y S. Cheng. 1994. Nitrogen excretion and changes of hemocyanin, protein and free amino acid levels in the hemolymph of Penaeus
monodon exposed to different concentrations of ambient ammonia - N at
different salinity levels. Mar. Ecol., Prog. Ser. 110: 85-94.
Cheng, W. y J. Chen. 2001. Effects of intrinsic and extrinsic factors on the haemocyte profile of the prawn Macrobrachium rosenbergii. Fish & Shellfish Immunology, 11: 53-63.
41
De Silva, S. 2000. A global perspective of aquaculture in the new millennium. En Book of Synopsis in the International Conference on Aquaculture on the third millennium. Department of fisheries. Bangkok, Thailand, pp : 51 – 100. Destoumieux, D., D. Saulnier, J. Garnier, C. Jouffrey, P. Bulet y E. Bachere.
2001. Antifungal peptides are generated from the C terminus of shrimp hemocyanin in response to microbial challenge. J. Biol. Chem, 276: 47070-47077.
El-Sayed, A. 1977. Growth rates and feed efficiency of the freshwater prawn
Macrobrachium rosenbergii fed varying protein and energy levels. Bull. Nat.
Inst. Oceanog. Fish. Egy. 23: 439-448.
Fukushima, M., G. Sifuentes, G. Saldaña, G. Castillo, J. Reyes y L. Shimokawa. 1982. Métodos Limnológicos. Universidad Nacional de Trujillo. Perú, 180 pp. Galindo, J., I. Fraga, M. Artiles, M. de Arazoza, S. Alvarez y E. Pelegrin. 2003.
Efecto de niveles de proteína en la dieta sobre el crecimiento de juveniles de camarón rosado (Farfantepenaeus notialis). En: II Congreso Iberoamericano Virtual de Acuicultura. CIVA 2003. 2003:575-586. Disponible en URL: http://www.civa.org.
Goimier, Y., C. Pascual, A. Sánchez, G. Gaxiola, A. Sánchez y C. Rosas. 2006. Relation between reproductive, physiological and immunological condition of
Litopenaeus setiferus pre-adult males fed different dietary protein levels
(Crustacea; Penaeidae). Animal Reproduction Science 92: 193-208.
Guerra, A., A. Gómez y J. Montez. 1983. Desarrollo post embrionario de
42
condiciones de laboratorio. Informe final de proyecto de investigación. Cod. PI 558.79 – d. BIO. Universidad Nacional de Trujillo, Perú.
Hernández – López, J., T. Gollas – Galván y F. Vargas – Albores. 1996. Activation of the prophenoloxidase system of the brown shrimp (Penaeus
californiensis Holmes). Comp. Biochem. Physiol, 113: 61 – 66.
Hose, J. E., G.G. Martin y A. S. Gerard. 1990. Decapod hemocyte classification scheme integrating morphology, citochemistry and function. Biol. Bull 178: 33-45.
Jori, D. “Estimados de Producción de Peces y Camarones”. Industriaacuicola.com. 2008. Global Aquaculture Alliance. 8 de oct. 2009.
Disponible en URL:
Http://www.industriaacuicola.com/PDFs/5.3%20_EstimadosPecesCamarones. pdf.
Jussila, J. 1997. Physiological responses of astacid and parastacid crayfishes (Crustacea: Decapoda) to conditions of intensive culture. Doctoral dissertation. Department of Applied Zoology & Veterinary Medicine. University of Kuopio. Perth, Western Australia.
Infante, M. 1985. Camarao gigante da Malásia, um bom negócio. Reproducao. Criacao. Recría e engorda. Comercializacao. Edit. Livraria Nobel S.A. Sao Paolo. Brasil, 120 pp.
43
Le Moullac, G., C. Soyez, D. Saulmer, D. Ansquer, J. Avarre, y P. Levy. 1988. Effect of hypoxia stress on the immune response and the resistance to vibriosis of the shrimp Penaeus stylirrostris Fish & Shellfish Immunol. 8: 621 – 629. Le Moullac, G., M. Le Groumellec, D. Ansquer, S. Froissard, P. Levy y Aquacop.
1997. Haematological and phenoloxidase activity changes in the shrimp
Penaeus stylirostris in relation with the moult cycle: protection against
vibriosis. Fish & Shellfish Immunol.. 7: 227-234.
Le Moullac, G.y P. Haffner. 2000. Environmental factors affecting immune response in Crustacea. Aquaculture, 191: 121 – 131.
Ligtner,D., K. Asno, B.White y R. Redman. 1998. Experimental infection of western Hemisphere penaeid shrimp with Asian White Spot Syndrome virus an Asian Yellow Head Virus. Journal of Aquatic Animal Health, 10: 271 – 281.
López, M., E. Segovia y A. Alfaro. 1986. Microalgas: Su importancia como recurso alimentario del camarón de río del norte de Chile, (Cryphiops
caementarius, Molina 1872). Medio Ambiente, 8(1): 39 – 47.
Luna, M., C. Graziani, E.Villarroel, M. Lemus, C. Lodeiros, y G. Salazar. 2007. Evaluación de tres dietas con diferente contenido proteico en el cultivo de postlarvas del langostino de río Macrobrachium rosenbergii. Zootecnia Tropical 25 (2): 111 – 121.
Mente, E., P. Coutteau, D. Houlihan, I. Davidson y P. Sorgeloos. 2002. Protein turnover, amino acid profile and amino acid flux in juvenile shrimp
Litopennaus vannamei: effects of dietary protein source. The Journal of
44
Meruane, J., M. Rivera, M. Morales y C. Galleguillos. 1996. Desarrollo de una tecnología para la producción de larvas y post larvas del camarón de río del norte Cryphiops caementarius (Decápoda: Palaemonidae) en hatchery. IX Congreso Latinoamericano de Acuacultura en Chile. 158- 163.
Molina-Vozzo, R., J. Heinen y L. D'Abramo. 1995. Supplementation of commercials feeds with beef liver for indoor nursery culture of freshwater prawn Macrobrachium rosenbergii. J. world Aquacult. Soc. 26(1): 103-106. Norambuena, R. 1977. Antecedentes biológicos de Cryphiops caementarius
(Molina 1872) en el estro El Culebrón (Crustácea, Decápoda, Palaemonidae) Bol. Pesq. Chile, 9 : 7 – 19.
Pascual, C., A. Sánchez, F.Vargas – Allbores, G. Le Moullac y C. Rosas. 2003. Haemolymph metabolic variables and immune response in Litopenaeus
setiferus adult males: The effects of an extreme temperature. Aquaculture, 218: 637 – 650.
Pascual, C., E. Zenteno, C. Gerard, A. Sánchez, G. Gaxiola, G.Taboada, J.Suarez, T. Maldonado y C. Rosas. 2004. Litopenaeus vannamei juveniles energetic balance and inmunological response to dietary protein. Aquaculture: 236, 431-450.
Perazzolo, L. y M. Barraco. 1977. The prophenoloxidase activating system of the shrimp Penaeus paulensis and associated factors. Dev. Comp. Immunol., 21: 385 – 395.
45
Ponce, J., P. Jiménez y C. Mejía de Ponce. 1982. Efecto de las Comunidades Epipetrales sobre el camarón de río Cryphiops caementarius (Molina, 1872) Universidad Nacional San Agustín de Arequipa.
Portugal, S., J. Vargas y E. Vega. 2003. Utilización del rotífero Brachionus
plicatilis en los primeros estadios del cultivo larval del camarón de río
Cryphiops caementarius. En: II Congreso Iberoamericano Virtual de
Acuicultura.CIVA 2003. 2003:837-850. Disponible en URL: http://www.civa2003.org.
PRODUCE. 2008. Evolución Del Sector Pesquero. Boletín Estadístico Mensual de la Oficina General de Tecnología de la Información y Estadística. Ministerio de la Producción. Lima, Perú.
Rameskumar, G., S. Ravichandran, G. Kaliyavaranthan y T.T. Ajithkumar. 2009. Comparison of Protein Content in the Haemolymph of Brachyuran Crabs. Middle-East Journal of Scientific Research 4(1): 32-35.
Reyes, W., M. Pilco y K. Olórtegui. 2002. Efecto de la ablación unilateral del pedúnculo ocular en la maduración ovárica y en el ciclo de muda de
Cryphiops caementarius (Molina 1872) (Decápoda, Palaemonidae) en
laboratorio. Civa ( http://WWW.civa 2002. org), 681 – 687.
Reyes, W. y H. Luján. 2003. Estados y subestados del ciclo de muda del camarón de río (Cryphiops caementarius Molina, 1872) (Crustácea Decápoda Palaemonidae. CIVA 2003 (http://www.civa2003.org), 808-817.
46
caementarius Molina, 1872 (Crustácea, Decápoda, Palaemonidae), en
condiciones de laboratorio. Informe final de Proyecto de Investigación. Código PIC - FF - CC -005 – 2005. Universidad Nacional del Santa, Perú. Reyes, W., S. Bacilio, M. Villavicencio y R. Mendoza. 2006. Efecto de la
salinidad en el crecimiento y supervivencia de postlasvas de Cryphiops
caementarius Molina, 1872 (Crustácea, Palaemonidae), en condiciones de
laboratorio. CIVA 2006 (http://www.civa2003.org), 341-346.
Rodríguez, J., R. Cedeño, C. Molina, V. Otero, E. Valenzuela y M. Sotomayor. 2000. Efecto de la calidad de la dieta sobre la respuesta inmune del camarón
Penaeus vannamei, In: Cruz-Suarez, L., Ricque-Marie, D., Tapia-Salazar, M.,
Olvera-Novoa,
Rosas, C., G. Cuzon, G. Gaxiola, C. Pascual, G.Taboada, L. Arena y A. Van Wormhoudt. 2002. An energetic and conceptual model of the physiological role of dietary carbohydrates and salinity on Litopenaeus vannamei juveniles, J. Exp. Mar. Biol. E col. , 268: 47 – 67.
Shinn- Pyng, Y., S.Tzeng-Gan, C. Chin-Chyuan, Ch.Winton y K. Ching-Ming. 2005. Effects of an organophosphorus insecticide, trichlorfon, on haematological parameters of the giant freshwater prawn, Macrobrachium
rosenbergii (de Man). Aquaculture, 243: 383 – 392.
Smith, L. L., P. G. Lee, A. Lawrence y K. Strawn. 1985. Growth and digestibility by Three sizes of Penaeus vannamei Boone: effects of dietary protein level and protein source. Aquaculture 46: 85-96.
47
Sritunyalucksana, K. y K. Soderhall. 2000. The proPO and clotting system crustaceans. Aquaculture, 191: 53 – 69.
Terwilliger, N. 2007. Hemocyanins and the immune response: defense against the dark arts. Intgrative and Comparative Biology 47(4): 662-665.
Truscott, R.y K. White. 1990. The influence of metal and temperature stress on the immune system of crabs. Funct. Ecol. 4: 455 – 461.
Valdeiglesias, F. 1983. Observaciones sobre la supervivencia del camarón de río
Cryphiops caementarius, Molina 1872 en diferentes refugios artificiales. Tesis
para optar el Título Profesional de Ingeniero Pesquero, Lima-Perú, 83 pp.
Van de Braak, K. 2002. Haemocitic defence in black shrimp (Penaeus monodon). Tesis PHD. Wagenningen Institute of animal science, the Netherlands.
Vargas – Albores, F. 1995. Sistemas de defensa del camarón café (Penaeus
californiensis). Cience, 46: 33 – 45.
Venturi, V. 1972: Cultivo de camarones en estanques, bioecología – crianza - repoblación. 1era. Ed. Universidad Nacional Agraria La Molina. Lima – Perú, 39 pp.
Viacava, M., R. Aitken y J. Llanos. 1978 . Estudio del camarón en el Perú 1975, 1976. Bol. Inst. Mar. Perú, 3 (5): 161-232.
Yaemsooksawat, N., O. Jintasataporn, N. Areechon, S. Puntuma-o-pas y Ch. Thongtuak. 2009. Songklanakarin J. Sci. Technol. 31(1): 15-20.
Yépez, V. y R. Bandini. 1997. Evaluación del recurso camarón de rio Cryphiops
caementarius en los rios Ocoña, Majes - Camaná y Tambo, Inf. Prog. Inst.
48
Zhang, X. B., C. H. Huang y Q. W. Qin. 2004. Antiviral properties of hemocyanin isolated from Penaeus monodon. Antiviral Res. 87: 96-99.
Zhang, Y., F. Yan, Z. Hu, X. Zhao, S. Min, Z. Du, S. Zhao, X. Ye y Y. Li. 2009. Hemocyanin from Shrimp Litopenaeus vannamei shows hemolytic activity. Fish & Shellfish Immunology 27: 330-335.
Zúñiga, O. y R. Ramos. 1990. Tasa Respiratoria de Cryphiops caementarius
49
50
Fig. 1.
Individuo hembra de
Cryphiops caementarius
capturado en el río Lacramarca, Provincia del Santa
– Departamento de Ancash.
51
52
53
Fig. 4.
Preparación de las dietas con 10%, 30% y 50% de proteína.
A. Insumos utilizados, B. Mezcla de ingredientes, C. Secado,
D. Trozado para obtener el tamaño adecuado del pelet.
A
C D
54
Fig. 5.
Dietas con 10, 30 y 50% de proteína para alimentación de
55
56
Fig.7.
Hemocitos observados en la hemolinfa de
Cryphiops
caementarius
después de 50 días de alimentación con 10, 30
y 50% de proteína en la dieta.
Fig. 8.
Hemocitos en frotis de hemolinfa. Coloración Giemsa.
Granulocitos (a), semigranulocitos (b) y Hialinos (c)
Fuente: Azañero, et al.(2006).
Hialinocito
57
Fig. 9.
Medida del peso de
Cryphiops caementarius
después
58
Tabla 1.
Análisis estadísticos de los resultados obtenidos sobre
hemocianina en la hemolinfa de
Cryphiops caementarius
después de 50 días de experimentación.
Medias con 95.0 intervalos HSD de Tukey
--- %
PD Frec. Media Desv. típica Límite inf. Límite sup. --- 10 12 0.23825 0.110901 0.178233 0.298267 30 12 0.2965 0.099337 0.236483 0.356517 50 12 0.44975 0.139306 0.389733 0.509767 T0 9 0.146333 0.067972 0.077031 0.215635 --- Total 45 0.2918
Análisis de la Varianza
--- Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor --- Entre grupos 0.5245 3 0.174833 14.50 0.0000 Intra grupos 0.494268 41 0.0120553
--- Total (Corr.) 1.01877 44
Contraste Múltiple de Rango
--- Método: 95.0 porcentaje HSD de Tukey
% PD Frec. Media Grupos homogéneos
--- T0 9 0.146333 X
10 12 0.23825 XX 30 12 0.2965 X 50 12 0.44975 X
59
Tabla 2.
Análisis estadísticos de los resultados obtenidos sobre
número total de hemocitos (NTH) en la hemolinfa de
Cryphiops caementarius
después de 50 días de
experimentación.
Medias con 95.0 intervalos HSD de Tukey
--- %
PD Frec. Media Desv. típica Límite inf. Límite sup. --- 10 12 2.68281E6 1236955.0 1.66308E6 3.70255E6 30 11 3.97E6 2617682.9 2.90492E6 5.03508E6 50 11 4.11307E6 2097596.1 3.04799E6 5.17815E6 T0 8 1.43984E6 537156.6 190928.0 2.68876E6 --- Total 42 3.15777E6
Análisis de la Varianza
--- Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor --- Entre grupos 4.36126E13 3 1.45375E13 4.21 0.0116 Intra grupos 1.31372E14 38 3.45716E12
--- Total (Corr.) 1.74985E14 41
Contraste Múltiple de Rango
--- Método: 95.0 porcentaje HSD de Tukey
% PD Frec. Media Grupos homogéneos
--- T0 8 1.43984E6 X
10 12 2.68281E6 XX 30 11 3.97E6 X 50 11 4.11307E6 X
60
Tabla 3.
Análisis estadísticos de los resultados obtenidos sobre
número de granulocitos en la hemolinfa de
Cryphiops
caementarius
después de 50 días de experimentación.
Medias con 95.0 intervalos HSD de Tukey
--- %
PD Frec. Media Desv. típica Límite inf. Límite sup. --- 10 12 428125.0 299484.6 205332.0 650918.0 30 11 631875.0 466941.9 399175.0 864575.0 50 11 864205.0 548973.8 631504.0 1.0969E6 T0 8 281250.0 108716.1 8384.9 554115.0 --- Total 42 567723.0
Análisis de la Varianza
--- Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor --- Entre grupos 1.90257E12 3 6.3419E11 3.84 0.0170 Intra grupos 6.27094E12 38 1.65025E11
--- Total (Corr.) 8.17351E12 41
Contraste Múltiple de Rango
--- Método: 95.0 porcentaje HSD de Tukey
%PD Frec. Media Grupos homogéneos
--- T0 8 281250.0 X
10 12 428125.0 XX 30 11 631875.0 XX 50 11 864205.0 X