SABINO ADRIAN RODRIGUEZ ALMARAZ
DIRECCION D E
UNIVERSIDAD AUTONOMA DE
HUEVO
LEON
FACULTAD DE CIENCIAS BIOLOGICAS
DIVISION DE ESTUDIOS
DE
POSTGRADO
•iO
POBL
A
CIONAL^
MORPOMETRÍA
Y CRECIMIENTO
Di
•>aru»
ciarkii
(GIRARD)
(CRUSTACEA; CAMeAR.IDAE:
DEL
AREA CENTRALDE NUEVO
LION MEXICO,
T E S I S
?UE PRESENTA
C O M O
REQUINTO FARCIAI
PARA OBTENER
EL GRADO Di
MAESTRO
E N C I E N C I A ?GABINO ADRIAN
R O D R I G U E Z A L M I A R A T•I ESPECIALIDAD EN E C O L O G Í A A C U A T I C A Y PESCA
/f y-í—
~ T M
r c B
UNIVERSIDAD JUJTOROMA D8 NUEVO LEON
FACULTAD DB CIENCIAS BIOLOGICAS
TAMAÑO POBLACION&L, MDRFOMETR1A T CRECIMIENTO EN Procambarua clarkli (CRUSTACEA: CAMBARIDAE) DEL
ASEA CENTRAL DE NUEVO LEO», MEXICO.
T E S I S
QUE PRESENTA CONO REQUISITO PARCIAL PARA OBTENER EL OTADO DE
MAESTRO EE CIENCIAS
SABINO ADRIAN RODRIGUES ATJtlBAE
con especialidad en Ecología Acuática y Pesca
•r
Presidente
Secretario
vocal
M. C. Roberto Mercado Hernández
l^rtdAQiilwto«
Agradezco la valiosa coloboración del Dr. Guillermo Compeán Jiménez, asesor y director de la tésis, por sus criticas y observaciones en el análisis de los resultados y conclusiones de La misma.
Al M. C. Roberto Mercado Hernández, por br i rularme sus conocimientos de bioestadistica desde el inicio de OLÍ profesión, además de enseñarme los paquetes de progresas que han sido importantes en mis actividades de investigación. Y por ofrecerme su apoyo y amistad.
A la Dr. Elizabeth Cruz Suárez, por sus consejos y criticas en la asesoría del presente trabajo; además de iluminarme con sus ideas para realizar estudios biológicos con los acociles.
No quiero pasar por alto la gran ayuda y asesoría del Dr. M. Badii para este estudio y que durante sus cursos de Ecología numérica, me ofreció diferentes oportunidades de investigación con
los artrópodos.
A la Administración de la Fac. de Ciencias Biológicas, dirigida por el M.C. Femando Jiménez Guzmán, por el apoyo y la oportunidad de iniciar otra fase de mi vida profesional.
A la D.I.G.S.A. de la S.E.P., por financiar económicamente el presente trabajo y otros estudios con acociles.
Agradezco la valiosa información escrita que me enviaron los distinguidos Carcinólogos, Dr. Horton B. Hobbs (Smithsonian Institution), Jr., Dr. Jay V. Buner (University of Southwestern Louisiana) y al Dr. ftalter Momot (Lakehead University, Ontario).
A las secretarias, Isabel (Entomología), Lupita y Roclo (Dirección y Coordinación de carrera), por su valiosa cooperación en la escritura y revisión del manuscrito.
A los maestros Bumberto, Haría Luisa, Raúl y Carlos, compañeros del laboratorio, por su amistad y comprensión hacia mi.
A los alumnos y ex-alumnos, Judith, Daniel Acosta, Roberto, Homero, José Luis, Ricardo Muñoz, María Elena y Rafael, por su invaluable ayuda en la colecta y mantenimiento de los acociles para el presente trabajo.
A la Biól. María de las Mercedes González Maltos, por su paciencia y disponibilidad en la asesoría de los programas Word perfect, Harvard graphic y SPSS.
Dedicatoria
En primer lugar quiero dedicar esta tésis a dos seres que me dieron la vida, de la cual ahora es mi profesión y placer:
A ais padres
Josefina Almaraz López, por su continua preocupación y cuidado a mi persona, que tal vez muchas veces no le tome importancia y por ser algo insípido no notes cuanto te necesito.
Javier Rodríguez Porras, por mucho tiempo hemos estado separados, sin embargo, siempre te he extrañado y querido, como cuando era niño que ansioso esperaba tu llegada.
A mis hermanos, Laura, Miriam, Javier, David y Fina, que ha pesar de ser distintos en comportamiento y vida, siempre los he querido y respetado.
A «i abuelita*
Ana Maria Porras
De niño me dejastes y empece a conocer la falta de un ser querido, el amor que siento por ti ha estado creciendo desde niño hasta hoy en mi vida adulta. Solo tu sabes de mis alegrías, satisfacciones y tristezas.
A «i «sposa*
Judith
Gracias por comprender y guardar silencio en tus momentos de tristeza y soledad, a causa de mi trabajo y durante la elaboración de esta tésis. Hoy quisiera, recompensar con amor y dedicación el tiempo transcurrido.
A «i cangrejitai
Ana Raren
INDICE
Pag.
INTRODUCCION 1
1.1. Antecedentes
1*1.1 Distribución y Taxonomía 2
1.1.2 Estructura trófica y Dinámica Poblacional 4
1.1.3 Reproducción 8
1.1.4 Crecimiento 10
1.1.5 Importancia en las pesquerías y Acuacultura..17
1.1.6 Importancia de los Factores fisicoquimicos
en la Biología de acociles 21
1.2. Area de Estudio 23
1.2.1 Climatología 23
1.2.2 Hidrología 23
1.2.3 Características ambientales generales... 23
1.3. Objetivos 25
1.3.1 Objetivo terminal 25
1.3.2 Objetivos específicos 25
1.4. Hipótesis 25
1.5. Originalidad. 25
1.6. Importancia 26
2.1. Estudio de campo 26
2.1.1 Tamaño Poblacional 28
2.1.2 Proporción sexual y estado de la muda 29
2.1.3 Relaciones biométricas y Crecimiento
relativo 30
2.1.4 Distribución de frecuencias de talla 31
2.2. Estudio de Laboratorio 31
2.2.1 Regeneración de Quelas 31
2.2.2 Crecimiento postmudal 34
2.2.3 Crecimiento individual 35
2.2.3.1 Tasa de Incremento relativo 35
2.2.3.2 Crecimiento relativo (Alometría) 36
2.2.3.3 Crecimiento usando el modelo de von
Bertalanffy 36
3.- RESULTADOS
3.1. En campo
3.1.1. Tamaño Poblacional 37
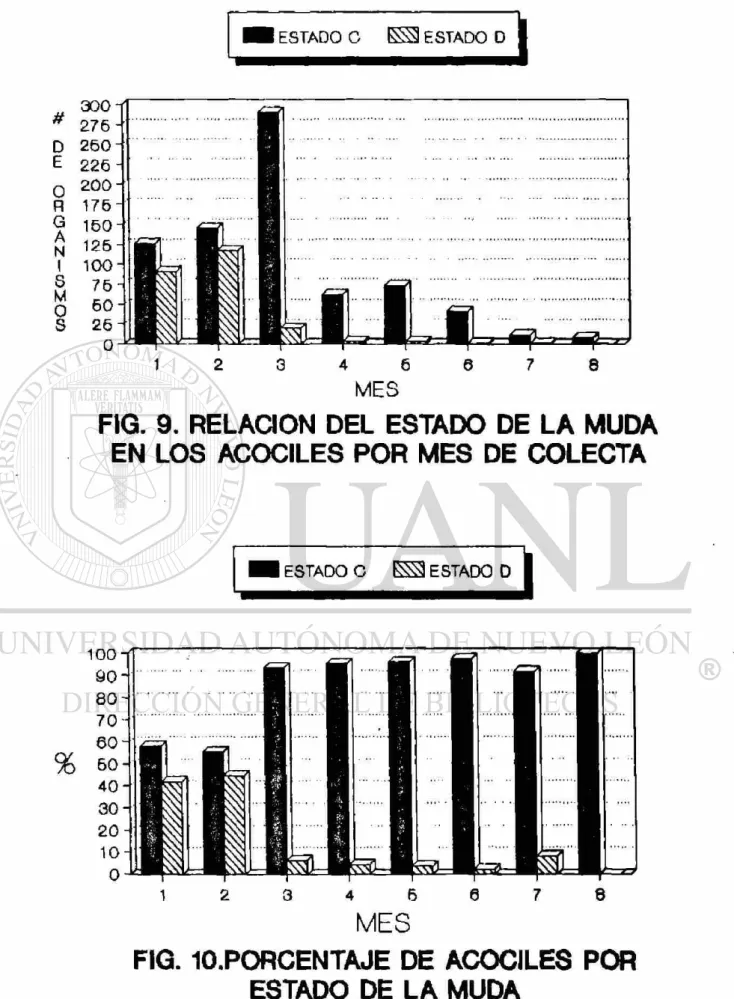
3.1.2. Estado de la muda 45
3.1.3. Porcentaje sexual 48
3.1.4. Distribución de la frecuencia de tallas...50
3.1.5. Crecimiento relativo (Alometría) de los
acociles capturados ...55
3.2. En laboratorio
crecimiento de la quela 62
3*2.2. Crecimiento de quela 65
3.2.3. Crecimiento neto por efecto de la muda 67
3.2.4. Crecimiento utilizando la relación del
crecimiento por efecto de la muda 67
3.2.5. Valores promedio de la relación del crecimiento de la quela por efecto
de la muda 68
3.2.6. Crecimiento promedio después de la segunda
muda 69
3.2.7. Comparación del crecimiento promedio en la
primera muda y segunda muda 69
3.2.8. Frecuencia de muda 70
3.2.9. Comparación de la ocurrencia de la muda en
los sexos 71
3.3. Crecimiento postmudal de acociles en laboratorio....73
3.4. Crecimiento individual en laboratorio 78
3.4.1. Tasa de Incremento relativo 78
3.4.2. Crecimiento alométrico • .80
3.4.3. Crecimiento con el modelo de von
Bertalanf fy 80
4.- DISCUSION
4.1. Tamaño Poblacional 84
4.2. Proporción sexual y estado de la muda 85
4.3. Distribución de la frecuencia de talla. 86
4.5. Regeneración de quelas
4.6. Crecimiento postmudal.
4.7. Crecimiento individual
CONCLUSIONES
RESUMEN
1.- INTRODUCCION
Los acociles del género pr^amb^rua spp. contiene 16 subgéneros y 134 especies, la mayoría se presentan en el Sureste de los Estados Unidos, con una inclinación hacia México y el Noreste de Cuba. El subgénero Scapuliy»ii^Tl1T incluye especies
como a Procambarus clarkii que tiene una amplia tolerancia ecológica (Hobbs, Jr., 1984; Hobbs, Jr*, 1989); esta especie se ha reportado en todos los Estados del Norte de México, incluyendo a Nuevo León, donde se considera que al nanos en el centro del Estado ha sido introducida (Hobbs III, et. al • 1989; Campos y Rodriguez-Almaraz, 1992). Por otro lado, recientemente se elevó a categoría específica al acocil endémico del centro de Nuevo León, como Procambarus reqiomontanus por Hobbs, Jr* y Robinson (1988); sin embargo esta especie ahora esta ausente de su localidad tipo y de otras áreas previamente reportadas y solo se conoce hacia Cerralvo, que se localiza hacia el Norte de la
localidad tipo (Monterrey). Esta reducción en las poblaciones se considera como principal factor a la presencia de Pr^aínharnn clarkii (Rodriguez-Almaraz y Campos, en preparación).
Se considera que los cangrejos de rio son los miembros más grandes y los más longevos de los crustáceos de agua dulce de Norteamérica, además constituyen una fuente de alimento para muchos peces y no pueden ser establecidos en un solo nivel trófico, ya que por sus hábitos alimenticios ocupan varios niveles tróficos y cualquier esfuerzo para discutir su bioenergia es difícil, considerándose como especies herbívoras, con alimentación suplementaria de origen animal, además representan el mejor camino para convertir la producción primaria, especialmente de material foliar en carne animal (Momot, Gowing y Jones, 1978)* Muchos acociles de bajas latitudes tienen una
longevidad muy corta, como las especies del género Procambarus sp. que viven dos años o menos, mientras que especies de altas latitudes y ambientes fríos, usualmente viven de cuatro a 16 años y la madurez sexual es más tardía (Momot, 1984)*
El ciclo de vida de los cangrejos de río tiene períodos de crecimiento y períodos quiescentes, durante estos últimos, los cangrejos de río se refugian en diversos lugares, usualmente en madrigueras, donde las hembras maduras desovan sus huevecillos, posteriormente a cada período quiescente continua un crecimiento rápido; la duración y temporalidad de ambos períodos depende de la latitud (Meredith y Schawrtz, 1960 en Huner, 1976).
calidad del alimento, principalmente larvas de insectos (Momot y Jones, 1977), en esta especie las tasas de crecimiento mensuales son mayores en machos que en hembras, debido a los periodos de incubación y cuidado de juveniles (Hazlett y Rittschof, 1985). El número mínimo de mudas para madurar en Procambarus clarkii son once, el crecimiento depende del tipo de habitat, en áreas como acequias y estanques temporales, el crecimiento y la madurez son menores que en grandes pantanos, ciénegas o estanques de cultivo
(Huner y Barr, 1984).
Los acociles son los organismos más fáciles de cultivar, ya que en comparación con otras especies de importancia comercial, las poblaciones se mantienen estables y el gasto de agua en los estanques de cultivo es menor que la requerida para cultivos de algunos peces. En el sur de Louisiana se encuentra el 90% del área dedicada al cultivo de estos organismos en los Estados Unidos, con producciones anuales de 807 kg por hectárea (Avault Jr. y Huner, 1985). Por otro lado, la producción anual bentónica en muchos lagos de Michigan fue dominada por la especie Orconectes virilis, con valores de 1000~1400 kg por hectárea, con densidades de 15 organismos por metro cuadrado (Momot, 1978). Uno de los mayores problemas en el cultivo de acociles es la reducción de la vegetación como alimento, la ausencia de ésta en los estanques ocasiona que el crecimiento se detenga en tallas subcomerciales de menos de 75 mm de longitud total (Cauge, et al., 1982; Avault Jr. et al., 1974).
1.1.ANTECEDENTES
1.1.1. Distribución y Taxonomía
Los cambarinos americanos ocupan toda la parte este de los Estados Unidos de America, desde las montañas Rocallosas hasta la vertiente Atlántica, siguiendo por todo el territorio mexicano y la parte Norte de América Central y Cuba. El cambarino Procambarus simulans reqiomontanus• se encuentra a 5 km de Monterrey, Nuevo León (localidad tipo), esta especie tiene relación con Procambarus simulans simulans de Kansas y Texas y una subespecie muy cercana a Procambarus simulans reqiomontanus. aún no descrita se encuentra en la Cd. de Rio Bravo, Tamaulipas
(Villalobos-Figueroa, 1955).
cambarino en la vertiente del Pacifico en las cuencas de los Rios Colorado y Sonora (Hobbs, Jr. 1962).
En Europa se han introducido especies de acociles de América en espera de encontrar especies que no sean afectadas por la plaga fungal. En Polonia Orconectes limosus fue introducida hace
87 años y posteriormente introducida en Alemania y Austria. Procambarus clarkii fue introducida en climas templados, como en la parte Sur de España. Pacifastacus leniusculus, es la especie más introducida en casi todo Europa (Avault, Jr.,1976).
Se han reconocido 284 especies de acociles americanos y para el estado de Nuevo León se registra a las especies Cambarellus alvarezi y Procambarus simulans reaiomontanus. estas dos son endémicas y la especie Procambarus clarkii. se localiza entre el estado de, Coahuila y Sonora (Hobbs, Jr» 1976).
La subespecie Procambarus simulans reqiomontanus se distribuye en diversos municipios del Centro y Este de Nuevo León, pero solo habita en la cuenca del Rio San Juan, una de las más importantes del Estado (Campos-González, 1982).
Los crustáceos decápodos de agua dulce de Mesoamérica, es un grupo heterogéneo y se distribuyen según su posibilidad de ocupar nichos no competidos, como es el caso de los astácidos. Esta área comprende varias zonas zoogeográficas y la fauna procede de áreas neárticas y neotropicales (Villalobos-Figueroa, 1982).
Casi la mitad de las 152 especies y subespecies de los acociles americanos son asignados al género Procambarus. habitando muchas en el Sureste de los Estados Unidos. En México hay dos especies aisladas en la vertiente del Pacifico y sobre la vertiente del Atlántico hay una amplia distribución hasta Belice, Guatemala y Noreste de Cuba e Isla Pinos. El subgénero Scapuli^<""harna que incluye a Procambarus clarkii. su rango de distribución es a tráves de la planicie costera y tienen amplia tolerancia ecológica construyendo madrigueras simples en épocas de sequía y reproducción (Hobbs, Jr. 1984).
En Tamaulipas, la presencia del acocil Procambarus clarkii se ha ubicado en el Noreste, Noroeste y Centro; que serian los primeros registros de esta especie en el Estado y su presencia se debe a posibles introducciones hechas por el hombre (Coronado-Magdaleno, 1987).
La subespecie Procambarus (Girardie 1 la) si mu lapa reqiomontanus ha sido elevada a rango especifico como Procambarus (G.) reqiomontanus. basándonos en las diferencias morfológicas, con la ahora especie Procambarus (<3.) simulans de (Texas y Kansas) y ante el desconocimiento de poblaciones entre el rango típico de esta última y la mexicana , considerándose que no hay intergrado en la distribución entre ellas (Hobbs, Jr. y Robinson, 1988).
familias, 10 géneros y 372 especies. La especie Procambarus reqiomontanus es solo conocida en Nuevo León y Tamaulipas (?) en ambientes lénticos y lóticos. La especie Procambarus clarkii, su
localidad tipo es entre San Antonio y el Paso, Texas; habitando ambientes lénticos y lóticos, y es un excavador terciario. Se considera introducida en Baja California, Chihuahua y Coahuila
(Hobbs, Jr. 1989).
Se ha establecido que las introducciones en México del acocil Procambarus clarkii, es para Baja California, Chihuahua, Coahuila y Sonora (Hobbs, III, Jass y Huner, 1989).
La especie Procambarus clarkii se considera nativa del Río Bravo central y su tributario el Río Salado, que atraviesa Chihuahua, Coahuila, Nuevo León y Tamaulipas; y es una especie exótica en el Centro y Oeste de Tamaulipas, Centro de Nuevo León, Oeste de Chihuahua, Sonora y Baja California, donde ha sido repetidamente introducida. El habitat preferido por éste acocil, es en ambientes lóticos con sustratos fangosos y la presencia de vegetación acuática (Campos y Rodriguez-Almaraz, 1992).
1.1.2. Estructura trófica y Dinámica poblaciona1
Los acociles son generalmente considerados detritívoros y su alimentación activa es cercanamente asociada con detritos vegetal. El decaimiento de material vegetal (detritos) soporta el crecimiento de bacterias, fitoplancton y zooplancton. Entre las plantas más comunes usadas en estanques de cultivo de acociles de Louisiana son Alternanthera philoxeroides Saqittarie> platyphylla y Zizania aquatica (Brunson, — — ) .
En la especie del acocil Orconectes palmeri de Tennessee, sus juveniles aparecen en Abril y la mayoría alcanzan su madurez a una longitud del caparazón de 18 a 25 mm, en el mes de septiembre u octubre. En este estudio para el mes de noviembre el 85% de todos los machos fueron machos forma I y muchos mudaron en junio para transformarse en forma II. La distribución de frecuencias de talla reveló 3 clases de edad (Payne y Price, ).
fue de 4497 kg/hectarea en la primera estación y 790 kg/hectárea en la segunda (Huner, 1978b).
En estanques de Louisiana se observó que la carpa Ctenopharynqodon idella tuvo un efecto en la. producción del acocil Procambarus clarkii. Carpas con promedios de 190 gramos de peso, en densidades de 4 por 0.01 hectárea, redujo
significativamente la producción (en número y peso) de acociles de talla cosechables (>74 mm de longitud total)• El análisis estomacal de estas carpas sugieren que la competencia por alimento (materia vegetal) fue la causa de la baja producción. Cuando la vegetación fue ausente, la cantidad de materia animal (cladóceros, acociles y girinidos) se incremento notablamente en sus estómagos (Forester y Avault, Jr., 1978).
Para determinar clases de edad en poblaciones naturales de Procambarus clarkii usando captura con trampas donde cada organismo capturado es colocado en un intervalo de clase (talla). De esta manera es fácil calcular el porcentaje de organismos en cada clase con respecto al total capturado; y así se pueden identificar clases de edad. Al diferenciar un grupo o más de otras clases de talla, se obtiene las modas. Puede haber un traslape entre las modas ya que algunos miembros de una clase de edad en particular son más pequeños o más grandes. En estanques de Louisiana durante los meses de invierno la captura fué baja, menos de una libra por trampa. El reclutamiento de Procambarus clarkii en Louisiana puede ser hasta dos generaciones en un año. En un estanque hay diferentes clases de edad y pueden ser adultos y juveniles de la temporada anterior de producción y juveniles de la temporada actual. El desove no se efectúa al mismo tiempo por eso hay diversas clases de edad en juveniles de la misma temporada. Estos últimos son los que van a formar parte del reclutamiento. En estanques se han encontrado hasta cinco clases, dos que corresponden a los que sobreviven de la temporada anterior y el resto son juveniles de la temporada, su primera clase de edad se presenta en septiembre y octubre en Louisiana y las otras dos aparecen de octubre a noviembre y de diciembre a enero (Huner, 1978a).
En los estanques de cultivo de acociles, los insectos acuáticos son en mayoría inmaduros, aunque también maduros y estuvieron representados en un 87% por la familia Corixidae. Las nayades del odonato Anax junius son depredadores de acociles; pero su comportamiento alimenticio depende de la temperatura, con tasas de depredación de 0.066 a 1.16 acociles/día (Witzing, et al. ).
clarkii tiene reclutamiento a través de todo el año en los mismos ambientes y varias generaciones por año (Huner, ).
La producción de arroz y del acocil Procaaiharna clarkii en Louisiana es significativa, ya que una alta producción de arroz se relaciona con una gran cantidad de acociles. El promedio de producción fué mayor (1059 kg/ha) que en los estanque control (800 kg/ha). También se encontró una relación inversa entre el incremento de calcio del suelo y la producción de acociles. La mortalidad natural durante las 8 semanas en estos estanque pudo haber sido enmascarada por el reclutamiento de juveniles; estos acociles son activos a temperaturas mayores a los 10 C e inactivos a temperaturas inferiores (Chien y Avault Jr,1980j.
Los acociles dependen para su alimentación del perifitón ( algas y microorganismos) y detritos (partículas de plantas en descomposición y microbios). La disminución de la vegetación natural que sirve de alimento y la pobre calidad del agua son dos de los principales problemas en el cultivo de acociles en Louisiana (Miltner y Avault,Jr 1981).
La construcción de histogramas de la distribución de frecuencias de tallas de los acociles permite evaluar y predecir la estructura demográfica de tallas y el crecimiento poblacional. Se encontró que en distintos estanques comerciales de Louisiana existió una diferencia en la estructura de las poblaciones después de seis y 12 semanas del reflujo de los estanques. Estanques con poblaciones con más del 45 % de los individuos que exceden 75 mm después de 12 semanas producen más (25 hasta el 30%) que los estanques con poblaciones con menos del 5% que exceden los 75 mm (Momot y Romaire, 1981).
La vulnerabilidad del acocil Procambarus clarkii al trampeo varía con la temperatura del agua y la captura por unidad de esfuerzo en estanques cultivados con arroz de Louisiana fué mayor que en estanques inundados tardíamente (octubre) que en los inundados en septiembre, con una diferencia promedio de 42 org. por cuatro trampas por día. Los factores que afectan la dinámica poblacional entre los dos tipos de inundación es la temperatura que favorece el crecimiento pero no la sobrevivencia, en especies como Pacifastacus leniusculus, las temperaturas de 18 a 20° C son buenas para el crecimiento pero pobres para la sobrevivencia. A temperatura de 10° C se reduce el crecimiento; pero hay alta
sobrevivencia. Otros factores son: oxígeno, disponibilidad de alimento y densidad (una alta densidad, afecta tanto la sobrevivencia y el crecimiento) (Chien y Avault, Jr. 1983).
reproducción. Los juveniles tuvieron dos máximos de actividad en verano e invierno y los adultos solamente un máximo de actividad. La migración de los acociles durante el drenado de los estanques
fue de un 32% de la población.
Los mecanismos que gobiernan el desplazamiento de la especie nativa Orconectes sanborni de Ohio por la especie invasora Orconectes rusticus son conocidos, ambas especies sin embargo tienen similiaridad en su ciclo de vida, en preferencias de habitat y hábitos alimenticios. En 0. rusticus sus juveniles crecen más rápido que los deQ. sanborni t los primeros son más grandes, por lo tanto dominan a 0. sanborni. En laboratorio 0. rusticus son menos susceptibles al pez predator Micropterus salmoides, ya que ocupan más frecuentemente refugios. Además mantienen su crecimiento poblacional alto, y una productividad mayor que 0_¡_ sanborni (Butler y Stein, 1985).
La especie Cherax tenuimanus tiene una respuesta tipo k, mientras que C. destructor tiene la estrategia tipo R. La primera
se caracteriza por una larga vida, ocupa habitats permanentes y estables y se reproducen anualmente. Además tiene una estructura poblacional estable y su "concha" relativamente dura. La segunda especie es de corta vida, ocupa habitats inestables y semipermanente, se puede reproducir varias veces al año. La estructura poblacional es inestable y su "concha" es relativamente delgada (Huner, 1986).
A una densidad de 19 /m el acocil Orconectes rusticus mantenido en jaulas experimentales durante 4 a 12 semanas, redujo el 64% de la biomasa de macrofitas, mientras una abundancia mayor de 140 gr/m eliminó todas las macrofitas.Las plantas utilizadas fueron Vallisneria americana, Juncus pelocarpus y Elodea canadensis. La abundancia y la reducción en la riqueza de especies sobre el tiempo de exposición sugieren que esta especie es un herbívoro selectivo. Macrofitas de tallo simple fueron más susceptible a la destrucción que Macrofitos con su tallo ramificado. Con estos resultados podemos sugerir que los acociles juegan un papel importante en la estructura de las macrofitas y las comunidades de invertebrados que habitan en ellas (Lodge y Lorman, 1987).
Las especies Procambarus clarkii y acutus en dos estanques comerciales del Sur de Louisiana fueron estudiadas para conocer
segundo grupo de hembras postovigeras fueron encontradas a principios de enero y se les relacionó con el reclutamiento a
finales de enero. Para P. acutus hembras postovigeras fueron observadas desde octubre a enero. Los machos forma I de P. clarkii estuvieron presentes a través de todo el año y más abundantes a finales de otoño y primavera. El promedio de talla de los machos forma I en otoño e invierno fue de 69 a 85 mm, esta especie madura en climas templados y subtropicales en dos u ocho meses, dependiendo de las condiciones ambientales (temperatura y dieta). El crecimiento de P. clarkii para otoño y primavera (1-3 mm por semana) fue la mitad del crecimiento (2-5 mm por semana) de P. acutus. Las tasas de mortalidad natural fueron de 3 a 13 % y 9% por semana, para P. clarkii y P. acutus respectivamente. La mínima talla de acociles que entran a la pesquería es de 70 mm , ya que es la talla mínima que retienen las trampas comerciales. La captura por unidad de esfuerzo en enero y febrero en ambos estanques debido a las bajas temperaturas (<10 C) ya que disminuyen las actividades alimenticias y movimientos hacia las trampas, aumentando la captura conforme aumenta la temperatura. La producción de acociles en ambos estanques fueron bajos (536-642 kg/ha), principalmente por reclutamiento secundario hasta obtener talla cosechable (>70 mm) y el mínimo esfuerzo de pesca
(Romaire y Lutz, 1989).
La producción excedente estimada usando el modelo de Schaefer para una población de Orconectes virilis en el lago Dock de Ontario, sugiere que la producción de ese "stock" es de 20 kg para hembras y 30 kg para machos con un esfuerzo de 4000 trampas por día por temporada. Un esfuerzo mayor de 6000 trampas por día produce bajos rendimientos como lo sugiere el uso de del modelo de Schaefer. Los resultados positivos en el empleo de este modelo se deben probablemente a que la capturabilidad es constante en machos (Momot, et al., 1990).
1.1.3« Reproducción
En la especie Pacifastacus leniusculus, las hembras grávidas fueron encontradas en el Rio Sacramento, California en ctubre, con una gran variabilidad en el número de huevos adheridos a los pleópodos, el promedio fue 190 huevos/hembra y la eclosión tuvo lugar entre marzo y abril (Shimizu y Goldman,-—-).
segundo grupo de hembras postovigeras fueron encontradas a principios de enero y se les relacionó con el reclutamiento a
finales de enero. Para P. acutus hembras postovigeras fueron observadas desde octubre a enero. Los machos forma I de P. clarkii estuvieron presentes a través de todo el año y más abundantes a finales de otoño y primavera. El promedio de talla de los machos forma I en otoño e invierno fue de 69 a 85 mm, esta especie madura en climas templados y subtropicales en dos u ocho meses, dependiendo de las condiciones ambientales (temperatura y dieta). El crecimiento de P. clarkii para otoño y primavera (1-3 mm por semana) fue la mitad del crecimiento (2-5 mm por semana) de P. acutus• Las tasas de mortalidad natural fueron de 3 a 13 % y 9% por semana, para P. clarkii y P. acutus respectivamente. La mínima talla de acociles que entran a la pesquería es de 70 mm , ya que es la talla mínima que retienen las trampas comerciales. La captura por unidad de esfuerzo en enero y febrero en ambos estanques debido a las bajas temperaturas (<10 C) ya que disminuyen las actividades alimenticias y movimientos hacia las trampas, aumentando la captura conforme aumenta la temperatura. La producción de acociles en ambos estanques fueron bajos (536-642 kg/ha), principalmente por reclutamiento secundario hasta obtener talla cosechable (>70 mm) y el mínimo esfuerzo de pesca
(Romaire y Lutz, 1989).
La producción excedente estimada usando el modelo de Schaefer para una población de Orconectes virilis en el lago Dock de Ontario, sugiere que la producción de ese "stock" es de 20 kg para hembras y 30 kg para machos con un esfuerzo de 4000 trampas por día por temporada. Un esfuerzo mayor de 6000 trampas por dia produce bajos rendimientos como lo sugiere el uso de del modelo de Schaefer. Los resultados positivos en el empleo de este modelo se deben probablemente a que la capturabilidad es constante en machos (Momot, et al., 1990).
1.1.3« Reproducción
En la especie Pacifastacus leniusculus, las hembras grávidas fueron encontradas en el Rio Sacramento, California en ctubre, con una gran variabilidad en el número de huevos adheridos a los pleópodos, el promedio fue 190 huevos/hembra y la eclosión tuvo lugar entre marzo y abril (Shimizu y Goldman,-—-).
66.2 mm de longitud total y en P. acutus a los 67 mm Pero se ha reportado que para la primera especie la madurez puede ser alcanzada a tallas de 50-55 mm De acuerdo a Huner ( ), el crecimiento de acociles esta en función de su habitat, en estanques comerciales donde se encuentra condiciones ideales y estables, se obtienen las tallas más grandes al madurar.
De acuerdo a Masón (1970), el comportamiento copulatorio en el acocil Pacifastacus trowbridqii es categorizado por los siguientes eventos, contacto con las quelas, toma entre ellos, colocación de las hembras en posición ventral, montaje, erección e inserción de los gonópodos, depósito de espermatotoro, separación de los gonópodos, liberación de las hembras. La copulación dura de 10 a 20 minutos.
La fecundidad de Astacus astacus en Suecia puede ser influenciada por la temperatura y la disponibilidad de alimento. El número de huevos esta claramente correlacionado con la talla de la hembra (Abrahamsson, 1972).
En la especie Procambarus clarkii de Louisiana, la cópula ocurre en mayo o en junio y hasta finales de verano o principios de otoño ocurre el desove. Los huevecillos son retenidos bajo el abdomen por una sustancia mucilaginosa. La fecundidad de ésta especie puede ser hasta de 700 huevecillos y la eclosión se presenta en dos o tres semanas. Los juveniles eclosionados permanecen con la madre hasta la segunda muda (Avault, Jr. 1972) .
La especie Procambarus clarkii en estanques comerciales de Louisiana presenta dos máximos en el desove y con un reclutamiento constante de juveniles a través del año, el primero se presenta en octubre y el segundo que es menor en diciembre y enero (de la Bretonne, Jr. y Avault Jr. 1977)
Las principales causas de mortalidad del acocil Procambarus clarkii en estanques de cultivo, puede ser la presencia de depredadores, enfermedades, tensión al mudar, bajos niveles de oxígeno, las condiciones climáticas y las actividades de pesca por el hombre. En estanques bien manejados donde no hay peces ó se controla la calidad del agua, se espera al menos que el 50-60% de juveniles de la estación sobrevivan hasta la talla comercial
(Huner, 1978a).
En la especie P. clarkii el desove ocurre en junio hasta octubre, con casos erráticos en diciembre, el máximo pico de eclosión es en Octubre. Huner (1975) estableció que puede haber más de dos generaciones por año cuando las condiciones son
favorables. La duranción de horas-luz por día tuvo una relación con la frecuencia de desove (Dendy, 1978).
eclosión a finales del verano e invierno. Los machos maduraron a tallas promedio de 35.8 mm de longitud de cef alotórax y las hembras a 36.5 mm Se encontró una relación directa entre el número de huevos producidos y la talla( Somer, 1984).
En Astacus astacus de Suecia, la reproducción en habitats naturales empieza en octubre cuando la duración del dia y la temperatura están decreciendo. La muda se efectúa durante el verano cuando hay altas temperaturas del agua. El mecanismo disparador para la reproducción fue decreciendo con la temperatura y la muda fue afectada por un incremento de temperatura. El desarrollo de los huevecillos que en la naturaleza tarda hasta nueve meses, fue acortado a cuatro meses por un incremento de temperatura. En esta especie cuando se mantiene de 18 a 20 ° C y hay una disminución por abajo de los
12°C, esto es suficiente para iniciar el proceso de cópula, en otras palabras el fotoperíodo juega un papel menos importante en estos procesos(Westin y Gydemo,1986).
El lago 223 del Noroeste de Ontario a sido experimentalmente acidificado desde 1976. La reabsorción de huevos, fertilización y tasa de desarrollo en Orconectes virilis no fué seriamente afectado por la acidificación hasta un pH del 5.1. La dureza incompleta del compuesto de la glándula del cemento que forma la membrana de la cápsula del huevo y el pedúnculo produce una pérdida de huevos de los pleópodos causando que la poblaciones decrezcan en su reproducción. El número de huevos en el ovario ha
sido correlacionado linealmente con la longitud de la hembra. La diferencia del número de huevos en el ovario y en los pleópodos se atribuye a una adherencia deficiente de los huevos; puesto que hay mortalidad, infertilización, liberación incompleta de huevos u otras causas como el canibalismo maternal (France,
1987).
El tiempo de incubación de los huevecillos de Astacus astacus es influenciado notablemente por la temperatura. Cinco lotes de huevos fueron sujetos a diferentes temperaturas, después de aproximadamente 600 CTU (unidad de temperatura celsius«°C dias). El número total de CTU para eclosionar se reduce de 1909 a 1300. No hubo diferencia sistemática en el éxito de la eclosión o supervivencia de juveniles debido a diferentes tratamientos de temperatura. Las hembras mudaron y remudaron después de cuatro a ocho semanas de la eclosión de los juveniles (Hessen, et al. 1987) ,
1.1.4. Crecimiento
Sacramento se evaluó utilizando datos de marca-recaptura, el incremento promedio de la longitud del cefalotórax fue 2.3 mm. en cada muda. Los acociles provenían del invierno, mudaron en marzo y parte de abril. La segunda y tercera muda ocurrió a principios de Junio y mitad de Agosto, respectivamente. La frecuencia de muda entre los sexos fue similar. La temperatura no solo puede afectar a la captura por trampa, sino también los períodos de muda. Las bajas temperaturas en marzo y abril corresponden a la alta actividad mudal. Con este incremento promedio es posible construir una gráfica que señale el número problable de mudas en cada edad. La tasa de mortalidad (Z) fue 0.67 que representa a todas las formas de muerte (predación, canibalismo, pesca, etc.). Hay un incremento de canibalismo durante la muda por haber una competencia intraespecífica por habitat. Las altas temperaturas incrementan la tasa de muda como en Astacus astacus además de la concentración de detritos (Shimizu y Goldman, ).
En Gecarcinus lateralis después de dos ó tres semanas de que un apéndice fué extirpado, una pequeña papila aparece de 2-4 mm. de longitud. Esta permanece en esta forma hasta que llegue el período premudal. Este crece rápidamente por 20 días aproximadamente, complementándose su crecimiento en 10 días antes de la ecdisis (Skinner, 1962).
El crecimiento de juveniles de Astacus astacus tiene una relación con la temperatura del agua. El crecimiento de los adultos en diferentes latitudes de Suecia fué diferente entre los
sexos. Los machos crecieron 9.95+/- 0.60 mm y en las hembras fue de 6.10 +/-0.38 mm. La tasa de crecimiento fué más baja en las hembras, esto se debió parcialmente a la reducción en actividades alimenticias, durante el período de incubación. El gran desarrollo de la quela de los machos, les da una posición dominante por la competencia de alimento. La temperatura, la disponibilidad de alimento y la densidad poblacional son los factores más importantes que influyen en la tasa de crecimiento
(Abrahamsson, 1972).
Pero la pérdida de todos los apéndices inhibe la muda. La talla de los apéndices regenerados en Gecarnicus es reducido por un tercio de su talla normal cuando los seis pereíopodos son perdidos y a la mitad cuando 8 apéndices extirpados están regenerando (Skinner y Graham, 1972).
La frecuencia de muda en los acociles de la especie Procambarus clarkii se incrementa por la autotomía de los quelípedos y apéndices locomotores a condiciones constantes de 18.5-20.5 °C. El incremento en la frecuencia de muda no puede ser explicado por una pérdida de biomasa ya que diversas operaciones pueden producir daño a tejidos, nervio o fibras musculares resultando en una ligera pérdida de biomasa, pero estas operaciones incrementan significativamente la frecuencia de muda
(Bittner y Kopanda, 1973).
Se ha planteado que el decremento en tiempo de una muda después de la autotomía se debe a la pérdida de masa corporal en Gecarnicus lateralis, sin embargo este procedimiento también esta relacionado con la inmovilización, que daña al exoesqueleto y al sistema neuromuscular; además de pérdida de hemolinfa. Y se considera que la pérdida de masa corporal no es la única causa del incremento en la frecuencia de muda. Los animales ablacionados de la especie Procambarus clarkii que sobrevivieron a la operación al menos 10 días, alcanzaron la ecdisis de 12-32 días; en esta especie, la frecuencia de muda en organismos con
sus 10 apéndices cortados fue del 82% comparado con el 48% a los que se les cortó solo ocho apéndices locomotores y la frecuencia de muda en el grupo control fue del 20%. El tiempo promedio de muda fué decreciendo de 88 +/- 31 días en el grupo tratado y en acociles con todos los apéndices mutilados de 45 +/- 16 días. Si la muda precoz es inducida solamente por pérdida de masa corporal, entonces la frecuencia de muda debe ser relacionada a la masa pérdida. En este estudio no hay una relación entre la masa corporal removida y el incremento de la frecuencia de la muda. Una causa puede ser el gran peso de la hemolinfa que es mayor al peso de todas las patas. En muchos crustáceos, incluyendo acociles, la regeneración de un apéndice funcional no ocurre sin una muda y la reparación del exoesqueleto no es completado hasta la muda (Bittner y Kopanda, 1973).
Para el mantenimiento de adultos reproductores del acocil Procambarus clarkii. si son depositados en pequeños recipientes no es necesario la aireación si el nivel alcanza 3.8 cm. La alimentación puede ser simple y barata; se puede utilizar las plantas lagarto íAlternanthera) o la elodea (Egeria densa), que proveen oxígeno y alimento. Bajo estas condiciones de cultivo la madurez de las crías (7.5 cm) se alcanza en seis u ocho semanas
Los acociles cavernícolas de Alabama como Orconectes virilis crecen muy lentamente y tienen gran longevidad, su crecimiento se representa por una tasa de incremento de 0.38 mm/mes y viven de 10 a 15 años. Este crecimiento fue menor que de las especies epígeas, que varía de 2.8 a 5.2 mm/mes. (Cooper y Cooper, 1976). La producción de acocil (Procambarus clarkii) de concha suave de tallas comerciales (50 a 70 mm) , se realizó en dos formas. En el primer estudio, el 92% (299 acociles) mudaron a los 10 días después de su introducción al laboratorio. La duración de intermuda en promedio fué menos de 12 días en enero, marzo y mayo, pero sobre 18 días en junio. El grupo control que provenían de otro origen mudó en 23 días. La inyección de hormonas y ligación del pedúnculo fueron usados para influir en la tasa de muda. La hormona formulada no aceleró la muda después de 4 días de la muda inicial. La ecdisesterona no acelera la muda después de inyectarse un día después de la muda inicial, pero sí la
facilita. La ligación del pedúnculo después de 4 días de la muda inicial aceleró este proceso y todas las mudas fueron exitosas. En el segundo estudio, los acociles mudaron en aguas duras (32 ppm) y agua deionizada y se determinó la dureza del caparacho a intervalos de tiempo diferentes (0-96 horas y 3 semanas). Los acociles mudaron en agua deionizada en un 1/3 de la dureza como en intermuda (Huner, 1976).
El crecimiento de juveniles Procambarus clarkii puede alcanzar tallas maduras en seis u ocho semanas despues de su liberación con las hembras en condiciones de laboratorio (Huner y Avault, Jr. 1976a).
Los subadultos de Procambarus clarkii (50-60 mm de longitud total) mudan rápidamente (13.1 +/- 5.8 días) a temperaturas de 26.7 +/- 1° C. El período de premuda (D) ocupa el 62% del ciclo entero de intermuda. La muda puede retardarse ya sea en el período de intermuda (C4) o en premuda temprana (Dq) del ciclo de
muda. El verano se considera como un periodo quiescente en las poblaciones de campo de Louisiana cuando el crecimiento cesa y baja el nivel del agua forzando a ellos a vivir en madrigueras
(Huner y Avault,Jr. 1976 b) .
Se ha observado la eficacia de la pérdida de un número crítico de apéndices ya sea pereíopodos o quelipedos en la estimulación de la muda en crustáceos. La tasa de crecimiento en Gercacinus lateralis sin pedúnculos oculares es más rápido y el crecimiento preecdisial ocurre cuando los apéndices son más pequeños que a los se les indujo la muda por autotomía (Holland y Skinner, 1976).
producen 4.5 millones de Kg valuados en 3.4 millones de dólares. Los estudios de la biología de los acociles requieren
frecuentemente la relación longitud/peso; esta es usada para determinar crecimiento, talla de madurez sexual y su comparación con datos de otras poblaciones de estos organismos. Los análisis de covarianza de los coeficientes de regresión de longitud-peso para machos y hembras para cada especie fueron significativamente diferentes. Los machos fueron más pesados, esto se debe al desarrollo acelerado de las quelas, mientras que la quela de las hembras permanecen isométricas con su longitud total através de su vida. Las dos especies de acociles presentaron un crecimiento alométrico con el peso, incrementándose más rápido que el cubo de la longitud. La longitud total y del cefalotórax son variables altamente correlacionadas para cada especie (Romaire, et al..
1976).
El crecimiento en crustáceos no puede efectuarse hasta que la vieja cutícula es reemplazada. Por un período muy corto (en horas) la nueva concha es suave, ésta absorve agua y se hincha. Virtualmente no hay incremento en peso y talla, hasta que la calcificación es lograda y hay un incremento dependiendo de la temperatura y la edad. Los fisiólogos reconocen que la ablación de los pedúnculos oculares estimula la muda pero hay complicaciones si el animal alcanza la madurez antes de que la operación sea efectuada. Para obtener acociles de concha suave es preferible obtener organismos en estado de premuda tardío pero son difíciles de capturar usando las técnicas de capturas típicas, porque no son atraídos a las trampas (Huner, 1977).
Acociles subadultos de Procambarus clarkii de 50 a 70 mm de longitud total, fueron inyectados con hormonas ecdisiales y también sujetos a una ablación bilateral del pedúnculo ocular; para comparar el acortamiento del período de intermuda. La ablación bilateral fué el mejor método para producir acociles de concha suave en comparación con la inyección de la ecdiesterona
(Huner y Avault, Jr, 1977).
Experimentos con juveniles o subadultos de Procambarus clarkii en un sistema de crecimiento intensivo, sugieren que las densidades que exceden de 20-25 acociles/m sin ninguna protección; producen baja sobrevivencia, pero no disminuyen en la tasa de crecimiento (Goyert y Avault, Jr., 1978).
son animales mucho más pequeños que aquellos que viven en ambientes más favorables, se observó también en las diferentes distribuciones de frecuencias, modas múltiples; y se debe por las presencia de dos o tres máximos de reclutamiento de juveniles, tanto en reservorios naturales, presas y estanques comerciales durante el otoño e invierno, además esta apariencia multimodal se debe al hecho que todos los acociles que están reclutándose no son necesariamente liberados hacia aguas abiertas al mismo tiempo. La diferencia de frecuencia de mudas entre juveniles de la misma edad puede influir la talla de madurez de los machos que tienen menor rapidez de crecimiento, como aquellos que son liberados previamente (Huner y Romaire, 1978).
El crecimiento de Procambarus clarkii en estanques comerciales de Louisiana puede detenerse en tallas pequeñas de 50-60 mm de longitud total, esto puede deberse al poco esfuerzo de cosecha, además del tipo de suplemento alimenticio. Acociles bien alimentados presentan hepatópancreas de color amarillo brillante
(Huner, 1978a).
El acocil Procambarus clarkii con tallas promedio de 65.8 mm de longitud total fue expuesto a dos densidades (6 y 12 organismos/m ), el crecimiento se presentó mejor cuando se les alimento con una combinación de dos plantas f Polyqonum y Jussiaea) que al alimentarlos con paja del pasto bermuda (Cynodon dactylon) en ambas densidades, pero fue mayor el crecimiento en la densidad de seis organismos. La supervivencia no fue afectada significativamente por la densidad y regimen alimenticio
(Romaire, et ¿1., 1978)
El crecimiento de Procambarus clarkii fue evaluado por la ecuación de von Bertalanffy en estanques de arroz y control de Louisiana. El crecimiento fué mayor en los estanques de arroz (longitud asintótica» 123.2 mm de longitud total) que en los estanques control (longitud asintótica» 106.9 mm de longitud total). Estos valores se obtuvieron mediante la captura de cada 15 días en estos estanques. La madurez se alcanzó a principios de marzo en los estanques de arroz; y la tasa de crecimiento fué mejor en estos por una mayor disponibilidad de alimento (Chien y Avault,Jr. 1980).
El intervalo de muda de acociles despedunculados decrece en un 25% y su tasa de crecimiento por muda se incrementa cerca de dos veces comparado con acociles intactos. Una eliminación
acociles alimentados diariamente, con respecto a los que se les alimento cada tercer o quinto día* Animales sin pedúnculo mudaron a los 11 y 16 días, aquellos sin pedúnculos y sin apéndices mudaron a los 11 y 18 días. La remoción de apéndices en acociles despedunculados también induce un decremento en la tasa de muda. La regeneración de apéndices en acociles juveniles es más rápida y alcanza la talla normal en dos mudas exitosas (Nakatani y Otsu, 1981).
Con capturas semanales de diciembre 1978 a abril 1979 se analizó el crecimiento utilizando la ecuación de von Bertalanffy
(1938), considerando solo acociles mayores de 75 mrn. El coeficiente de crecimiento y la longitud asintótica de los acociles en estanques inundados y sembrados con arroz tempranamente (septiembre) y tardíamente (octubre) fueron
k-0.050 y longitud asintótica« 92.2 mm y k- 0.0896 y longitud asintótica- 82.7 mm, respectivamente. Se considera que un valor alto de k significa una longitud asintótica menor (Chien y Avault, Jr. 1983).
El crecimiento postembriónico temprano de juveniles incluye desde la eclosión hasta la independencia de la madre, en algunas especies este periodo tarda hasta el tercer estadio. En Orconectes neglectus chaenodactylus durante este estadio tiene ya una apariencia en miniatura como los adultos. El promedio del caparacho es de 3.5 mm, éstos salen a explorar dejando la cámara maternal; pero regresan cuando hay alarma. En Procambarus clarkii, al llegar al tercer estadio son independientes de la madre de acuerdo a Penn (1943). El desarrollo de estructuras sexuales primarias se completa en tres mudas y los acociles llegan a su madurez en seis u ocho mudas (Price y Payne, 1984).
La muda de los acociles Procambarus clarkii y P. acutus es más frecuente en juveniles que viven en aguas cálidas. La muda podría ser cada 5 ó 10 días, mientras los más viejos (pero inmaduros), usualmente mudan alrededor de los 30 días. El calcio es necesario para endurecer el exoesqueleto y es inicialmente tomado de los gastrolitos, hepatopancreas y sangre. Los acociles de concha suave para su consumo deben ser capturados antes de 12 horas para obtener la mejor calidad (Culley, et al. 1985).
Juveniles de Procambarus clarkii que fueron alimentados con 3 dietas isocalóricas (harina de sardina, harina de soya, harina de acelga y agar en diferentes porcentajes) y a temperaturas de 26, 24 y 20 C, presentaron una sobrevivencia relacionada con la temperatura, el rango térmico intolerable fue de 25 a 30 C y el rango aceptable fué de 18 a 22 ° C. El tratamiento que tuvo un mejor efecto en la sobrevivencia fué la temperatura de 20 ° C y la dieta de origen animal (Denise-Re Araujo, 1985).
Procambarus clarkii fue evaluado el crecimiento, biomasa ^ productividad a densidades de 1,2,4,8 y 16 organismos por metro en estanques con arroz cultivado. El crecimiento en longitud total y peso, fue afectado significativamente por la densidad, promediando de 91.5 mm y 20.7 g en talla y peso, respectivamente, para la densidad de 1 m , y hasta 62.5 mm y 6.3 g para la densidad mayor (Lutz y Wolters, 1986).
Acociles de la especie Procambarus clarkii que fueron capturados en nueve estanques comerciales y depositados en recipientes de cultivo dentro de un "invernadero", mostraron un patrón de muda que fue predominantemente diurno. Más del 90% de las mudas ocurrió entre las 7 y 18 horas. Estos resultados concuerdan con observaciones previas de estos organismos en dos areas comerciales para cultivo y bajo condiciones de laboratorio con luz artificial (Culley y le Duobinis-Gray, 1987a).
Se examinó el efecto de extirpar una quela o ambas y dos perelopodos sobre el crecimiento y frecuencia de muda en la jaiba azul Callinectes sapidus. La pérdida de una quela no alteró el incremento por muda y frecuencia de muda comparando con animales intactos. La múltiple pérdida de apéndices, redujo significativamente el incremento de muda en la primera muda pero no afectó la duración de la intermuda. En la segunda muda después de la autotomia no fue alterado en jaibas sin cuatro apéndices de aquellos sin una quela o intactos. La regeneración de los apéndices fueron de un 85 a 88%. Hasta tres mudas fueron requeridas para regresar al 100% del tamaño de los apéndices
(David Smith, 1990).
1.1.5. Importancia en Acuacultura y Pesquerías
En la especie A. astacus de Suecia, las bajas temperaturas decrecen la actividad alimenticia e influyen en la captura. En el verano de 1961, 20 trampas produjeron 396 acociles; pero el efecto inhibitorio de bajas temperaturas decreció la captura a cuatro acociles solamente. En esta especie sus adultos mudan una vez al año; pero varia de acuerdo a las altas temperaturas y la talla de acocil (Abrahamsson, 1972).
En el Rio Sacramento de California durante 1977 y 1978, se estimó la densidad de la especie Pacifastacus leniusculus, donde se encontró valores de captura por trampa de 0.18 a 2.27 kg/trampa en diferentes sustratos; el sustrato rocoso fue el preferido por esta especie, además de que no es una especie escavadora. La densidad promedio fue 1.8 acociles adultos por m
(Shimizu y Goldman, - — - ) .
Procambarus clarkii fue evaluado el crecimiento, biomasa ^ productividad a densidades de 1,2,4,8 y 16 organismos por metro en estanques con arroz cultivado. El crecimiento en longitud total y peso, fue afectado significativamente por la densidad, promediando de 91.5 mm y 20.7 g en talla y peso, respectivamente, para la densidad de 1 m , y hasta 62.5 mm y 6.3 g para la densidad mayor (Lutz y Wolters, 1986).
Acociles de la especie Procambarus clarkii que fueron capturados en nueve estanques comerciales y depositados en recipientes de cultivo dentro de un "invernadero", mostraron un patrón de muda que fue predominantemente diurno. Más del 90% de las mudas ocurrió entre las 7 y 18 horas. Estos resultados concuerdan con observaciones previas de estos organismos en dos areas comerciales para cultivo y bajo condiciones de laboratorio con luz artificial (Culley y le Duobinis-Gray, 1987a).
Se examinó el efecto de extirpar una quela o ambas y dos pereíopodos sobre el crecimiento y frecuencia de muda en la jaiba azul Callinectes sapidus. La pérdida de una quela no alteró el incremento por muda y frecuencia de muda comparando con animales intactos. La múltiple pérdida de apéndices, redujo significativamente el incremento de muda en la primera muda pero no afectó la duración de la intermuda. En la segunda muda después de la autotomía no fue alterado en jaibas sin cuatro apéndices de aquellos sin una quela o intactos. La regeneración de los apéndices fueron de un 85 a 88%. Hasta tres mudas fueron requeridas para regresar al 100% del tamaño de los apéndices
(David Smith, 1990).
1.1.5. Importancia en Acuacultura y Pesquerías
En la especie A. astacus de Suecia, las bajas temperaturas decrecen la actividad alimenticia e influyen en la captura. En el verano de 1961, 20 trampas produjeron 396 acociles; pero el efecto inhibitorio de bajas temperaturas decreció la captura a cuatro acociles solamente. En esta especie sus adultos mudan una vez al año; pero varía de acuerdo a las altas temperaturas y la talla de acocil (Abrahamsson, 1972).
En el Río Sacramento de California durante 1977 y 1978, se estimó la densidad de la especie Pacifastacus leniusculus, donde se encontró valores de captura por trampa de 0.18 a 2.27 kg/trampa en diferentes sustratos; el sustrato rocoso fue el preferido por esta especie, además de que no es una especie escavadora. La densidad promedio fue 1.8 acociles adultos por m
(Shimizu y Goldman, - — - ) .
utilizada en el acocil Procambarus clarkii para observar su respuesta sobre el crecimiento. Al mantenerse individualmente juveniles, el incremento promedio en longitud total y peso fue 5.1 mm y 1.65 gr, respectivamente; esta se administro ad libitum. Al mantenerse en varias densidades el incremento corporal fue de 3.7 mm a 5.9 mm.
El uso de los acociles data de muchos siglos, aunque los griegos y romanos no los consumían porque consideraban que eran venenosos. Los griegos colgaban acociles vivos en árboles y cultivos para proteger a estos de las enfermedades. Los gastrolitos fueron usados como medicina. En Escandinavia se ha usado el gastrolito para la buena suerte, los juveniles de la especie Pacifastacus leniusculus son vendidos comercialmente en 12 países de Europa, utilizando estanques para obtener huevas de las hembras que producen de 100 a 400 huevecillos y en un tiempo de 16 a 18 días de edad, estos son enviados para su engorda en diferentes estanques. El costo alcanzaba precios hasta $3.00 dólares por cria. En Suecia y Finlandia, la libra de Astacus astacus alcanza $9.00 dólares (Avault, Jr., 1976)
Los estanques nuevos que son utilizados para el mantenimiento de acociles juveniles durante la primavera, posteriormente son drenados y permanecen secos en el verano. El drenado induce el enmadrigamiento y la reproducción, elimina peces predatores y permite el crecimiento de pastos anuales que sirven de alimento cuando los estanques son de nuevo llenados. Estanques secos son llenados en otoño durante las dos primeras semanas de octubre, la cosecha empieza en enero y continua hasta mayo y se vuelve a llenar otra vez. La especie más adecuada para cultivar es Procambarus clarkii, los estanques de Louisiana producen de 500 a 1000 kg/ha por estación. En veinte estanques pequeños de Louisiana se capturaron 25,873 acociles, la mayoría (88%) fueron menores de 75 mm de longitud total que es la talla mínima comercial, de estos el 37% fueron entre 5 a 7 cm que es la talla capturable para el cebo utilizado. La proporción sexual fue 1:1; de 7,928 acociles marcados de 4.5 cm o más, Be recapturó 6,548 encontrando animales que habían sido marcados en otros estanques. Las densidades promedio variaron con el período del reflujo presentando valores mayores en el reflujo de septiembre y noviembre, con promedios de 11.4 y 11.7 organismos/m , respectivamente. Existió una diferencia de acociles con respecto a las formas de captura, con trampas el 77% fueron acociles en intermuda (Huner y Avault, Jr. 1976a).
Se ha reportado que trampas para acociles con cebo protegido captura menos de la mitad del número que se podrían capturar en trampas similares con cebo expuesto, con este los acociles fueron capaces de masticar más este y libera atrayentes en el agua, además el masticado envuelve respuestas auditivas que estimulan movimientos hacia las trampas (Westman, et al», 1978).
Uno de los problemas que hay en el uso de estanques de tierra para la producción de acociles, es la migración y construcción de madrigueras, ya que esto dificulta la cosecha. Los acociles
tienen la habilidad de utilizar oxigeno atmósferico por largos períodos si mantienen húmedas sus branquias. El suelo es innecesario como sustrato y el enmadrigamiento no es esencial para la reproducción. Las plantas acúaticas como elodea (Egeria densa) y la planta lagarto (Alternanthera philoxeroides) son adecuadas para incrementar la superficie del sustrato, además son un fuente de alimento. Se ha observado que los juveniles de Procambarus clarkii recién eclosionados no pueden permanecer en concentraciones de oxígeno inferiores a una parte por millón. La densidad en las poblaciones es un factor importante que influye en el comportamiento agresivo y la sobrevivencia de los acociles
(Nelson y Dendy, 1979).
La experiencia ha demostrado en Louisiana que estanques de cultivo de Procambarus clarkii con un buen manejo,la cosecha puede alcanzar de 800-1100 kg/ha después de un año, en ausencia de depredadores, en algunos se reportan hasta 2200 kg/ha, sin embargo, algunos depredadores no se pueden eliminar y que son protegidos por la ley, como las aves acuáticas, también algunos
insectos y arañas comen acociles, y estos no se han podido eliminar pero ellos desaparecen de los estanques cuando la vegetación muere y se descompone, ya que ésta les sirve de protección (Huner, 197 8a).
Uno de los mayores problemas en el cultivo de acociles es la reducción de la vegetación como alimento. En ausencia de ésta, el crecimiento se detiene en tallas subcomerciales (<75 mm de LT). Es importante el forraje para que ofrezca un sustrato adecuado a través del período de crecimiento y poder maximizar la producción de acociles. El uso de alimento tipo "pellet" es bueno para el crecimiento; pero su uso es limitado por los altos costos (Cauge, et al., 1982).
significativamente más acociles que trampas idénticas pero de alambre galvanizado,sin embargo la captura de retención no fue relacionado con el material de las trampas. La postura de las trampas (horizontal o vertical) no afectan la eficiencia de captura ni la capacidad de retención de estas. Trampas con cebos expuestos capturó 40-47% más acociles que trampas con cebos protegidos. Un promedio de 84, 80 y 64 % de acociles marcados
fueron retenidos por las trampas después de 12, 24 y 48 dias, respectivamente (Ffister y Romaire, 1983).
Utilizando diferentes formas de mareaje, desde cortes de urópodos, uso de hilos de nylon y tintura vegetal en el acocil Procambarus clarkii se determinó que no hay un marcador efectivo para determinar el crecimiento, ya que todas las marcas fueron de corta duración. Al usar diferentes tipos de trampas y cebos, se observó que la mayor eficiencia de captura fue con trampas de trampas con cuatro entradas y el cebo más efectivo fue alimento para perros (Sanchez-Saavedra, 1983).
La producción promedio de acociles en campos de arroz de Louisiana, que son inundados al principio (septiembre) y más tarde (octubre) de la temporada fué de 1183 kg/ha y 1127 kg/ha respectivamente (Chien y Avault Jr., 1983).
Los cebos son muy importantes en la cosecha de acociles, pero es el gasto mayor en la producción de estos. Muchos productores usan a la cuchilla (Dorosoma cepedianum) pero a veces no está disponible. Han sido utilizados otros cebos menos deseables como el arenque Brevoortia pastronus, los matalotes (Catastomidae) o los bagres lIctalurus punctatus). El costo de los peces como cebo puede alcanzar precios de $0.40 dólar por kg, sin incluir costos de almacén, transporte y otros gastos. Los cebos artificiales pueden eliminar estos problemas, pero debe tener ventajas de manejo, aceptabilidad y competitividad en el costo de peces y una captura efectiva de acociles. Se han utilizado diferentes cebos artificiales que contenían el mismo acarreador (carne de pescado, harina de semilla de algodón y harina de soya); pero con diferentes atrayentes (harina de sangre, huevos pulverizados y
aceite de bagre). Los resultados mostraron que todos los cebos artificiales fueron significativamente más eficientes que la carne de la cuchilla, con excepción de la harina de sangre
(Avault, Jr., et al., 1984).
el 70-92% de los animales seria consumido. Para esto se colectarían acociles inmaduros de talla comestible que esten en intermuda y que muden en 30 días (Culley, et al., 1985).
Los cebos tradicionales usados en la captura de acociles son carne de peces, como la carpa común y mojarra, demás se han desarrollado cebos manufacturados sin embargo no son tan eficientes como los naturales. Un problema es la estabilidad que es de pocas horas a días. Hay cuatro factores fisicoquímicos que afectan la capturabilidad en los estanques; temperatura del agua, densidad de los acociles, lluvias y frentes fríos. La captura se
incrementa cuando la temperatura se eleva hasta los 30° C (Huner, 1990).
1.1.6. Importancia de los Factores Fisico-Químicos
La tolerancia a la salinidad de Procambarus clarkii fue directamente proporcional a la talla de los acociles. Los recién eclosionados fueron muertos en menos de una semana en salinidades de 15, 20 y 30 ppm; adultos de 40 a 120 mm de LT no mostraron mortalidad significativa despues de una semana en salinidades mas de 30 ppm Salinidades de 10 ppm incrementaron el crecimiento, pero de 20 ppm o más altas retardaron el crecimiento (Loyacano,
-).
El incremento de la dureza del agua (como Cloruro de Sodio) incrementa el promedio de peso ganado y el porcentaje de supervivencia. Buen crecimiento de acociles ocurre en ausencia de suelo, sí la dureza del agua es alta, el exoesqueleto de acociles en estanques sin suelo es suave ( de la Bretonne, 1969).
La dureza del agua en estanques de Louisiana es el factor más significativo que afecta el crecimiento y supervivencia de Procambarus clarkii. El aumento de la dureza se relaciona con el incremento de peso ganado y el porcentaje de sobrevivencia , por ejemplo con dureza de 200 ppm del agua, el peso aumenta hasta
12.35 g. y un 91 % de sobrevivencia (Bretonne,Jr. y Avault, Jr 1971).
Los mayores problemas en el cultivo de acociles es la reducción de oxígeno y el hacinamiento. La calidad del agua en estanques comerciales está relacionado con el origen del agua, crecimiento denso de vegetación terrestre y la pobre circulación. Cuando el oxígeno es bajo los acociles toman oxígeno atmosférico. Para aumentar la disponibilidad de alimento en los estanques el método más rápido es fertilizar a una tasa de 110 kg de
fertilizante químico (8:8:2) por hectárea ( Avault Jr., de la Bretonne y Huner, 1974).
acociles en cualquier ambiente; pero es de mayor importancia en estanques donde el control es difícil* Cherax destructor es más tolerante a las bajas de oxígeno que Ch. tenuijQanus, la primera sobrevive a condiciones de 1 ppm; la segunda especie trata de escapar a estas condiciones buscando oxigeno atmosférico como en otros acociles. Ch. tenuimanus crece mejor a 70° F y está adaptado a climas más fríos. El crecimiento en ambas especies cesa a los 58° F (Huner, 1986).
La especie nativa de Europa Astacus astacus puede tolerar bajas concentraciones de oxígeno disuelto de 2 ppm en invierno en
lagos de Finlandia (Avault, Jr., 1976).
Juveniles de Procambarus clarkii de 9 - 12 mm. de LT fueron menos tolerantes a cambios abruptos en el oxígeno que juveniles de 31-35mm LT, la LCg0 fue entre 0.75 a 1.10 ppm de 02 disuelto
para los de 9-12 mm, acociles de mayor talla en ecdisis fueron más susceptibles a bajas de oxígeno que aquellos en otro estado de muda ( Melancon Jr. y Avault Jr., 1976).
Juveniles del segundo estadio del acocil Pacifastacus leniusculus alimentados con el camarón salino (Artemia) y expuestos a diferentes gradientes de salinidad y temperatura constante de 17.5° C. El crecimiento decreció con el incremento de salinidad hasta 17.5% (mitad de la salinidad del agua de mar), además del consumo de alimento. La diferencia de crecimiento a lo largo de los gradientes de salinidad puede ser una simple consecuencia del decremento de la toma de alimento ( Rundquist y Goldman, 1978).
Los siguientes parámetros fisicoquímicos son importantes para el cultivo del acocil Procambarus clarkii y Procambarus acutus acutus. Los rangos de temperatura es de 32 - 95° F, para una actividad mínima es menos de 50 F y para el crecimiento óptimo es de 70 - 80° F. La mortalidad es a menos de 1 ppm de oxígeno disuelto. La reducción del crecimiento y stress se presenta a 1
-3 ppm y el crecimiento óptimo es a valores mayores de -3 ppm. El pH óptimo del agua es de 6.5 - 8.5 y es letal cuando hay valores mayores de 10.5 y Menores de 4. El pH del suelo debe ser de 6
-7. La dureza y alcanilidad esta en los rangos de 50 - 400 ppm (CaCo3) dureza total y 50 - 400 ppm (CaCo3) para alcalinidad y la salinidad es de 10 - 15 ppm, sobreviven pero no crecen a menos de 2 ppm, para cultivos comerciales. Toxicidad amplia a diferentes pesticidas, generalmente los más tóxicos son los piretroides sintéticos (Romaire, 1986).
la mortalidad. Acociles inmaduros más vigorosos no mudan a 72° F y si a 80 F. El patrón de muda de los juveniles es durante el día ( Culley y Duobinis-Gray, 1987b).
1.2. AREA DE ESTUDIO
Para el estudio de campo se trabajó en el lecho del Río San Juan (Fig. 1). En la localidad denominada Congregación La Boca, perteneciente al municipio de Santiago, Nuevo León. La localización exacta es en las coordenadas geográficas 25° 27' 1 1 " latitud Norte y 100° 05' 1 2 " longitud Oeste; sobre la carretera vecinal Cortinas de la Presa Rodrigo Gómez, "La Boca" a Cadereyta, km cinco. La elevación de la zona es de 400 m sobre el nivel del mar. Además, es necesario mencionar que algunos ejemplares de acociles fueron colectados del Parque Canoas, Monterrey, N.L., para estudios de regeneración de quelas.
1.2.1. Climatología
Se encuentra dentro de grupos de climas templados C (Koppen, modificado por García, 1982); además se presenta en esta área el
subtipo climático semicálido-subhúmedo. Las temperaturas más altas se registran en Junio y Julio con un promedio de 28° C y las temperaturas más bajas en Diciembre y Enero con un promedio de 13° C; la precipitación anual es del orden de 700 a 1200 mm
(S.P.P., 1981).
1.2.2. Hidrología
El Río San Juan se origina en el municipio de Santiago, N.L., a partir del arroyo "La Chueca" que desemboca en la presa Rodrigo Gómez, "La Boca" y sus aguas forman este afluente; además este Río constituye una de las principales cuencas hidrológicas del Estado y su curso llega hasta la presa Falcón, en el Estado de Tamaulipas.
1.2.3. Características ambientales generales.
mixtos. La vegetación acuática mas común fue el berro, lentegillas y el lirio acuático; los animales mas comunes fueron principalmente macroinvertebrados como larvas y adultos de insectos de los grupos de coleópteros, efemerópteros, odonatos y hemipteros. En relación a vertebrados, peces poecilidos y aterinidos. Es necesario aclarar que estas observaciones fueron cualitativas y macroscópicas.
1.3. OBJETIVOS
1.3.1. Objetivo terminal.
Contribuir al conocimiento biológico y ecológico del cangrejo rojo de río Procambarus clarkii.
1.3.2. Objetivos específicos.
Determinar el tamaño y la variación mensual de una población de Procambarus clarkii.
Analizar la estructura demográfica y el modelo o tasa de crecimiento relativo de los cangrejos de río.
Estimar las relaciones biométricas en talla y peso. Determinar el estado de la muda.
Evaluar en laboratorio el proceso de regeneración de quelas y la frecuencia de muda.
Analizar el modelo y tasa de crecimiento mensual de juveniles en laboratorio.
1.4. HIPOTESIS
El tamaño y composición de la población será diferente en cada colecta, a causa de las condiciones ambientales y el comportamiento individual de los cangrejos de río.
El crecimiento relativo será isomètrico con respecto a la longitud total y longitud del cefalótorax.
La regeneración de quelas estara correlacionada con la presencia de muda en cada acocil y la velocidad de crecimiento en talla será mayor en etapas juveniles que en organismos maduros sexualmente.
1.5. ORIGINALIDAD
de los acociles del género Procambarus sp. en la región Norte de México, que son enfocados generalmente hacia aspectos de distribución y sistemática. No obstante ser un recurso ampliamente distribuido en diversos ambientes dulceacuícolas.
1.6. IMPORTANCIA
Al analizar las características bioecológicas de los cangrejos de río de la región, contribuímos al conocimiento de una especie dulceacuícola de importancia ecológica y potencialmente económica, que podría ser cultivada y representar una fuente importante de proteína.
2. MATERIAL Y METODOS
Para el estudio biológico y ecológico de los cangrejos de río o acociles de la región central de Nuevo León, se utilizó la especie Procambarus clarkii; originalmente se había
comtemplado incluir, también la especie Prnnamhania regiomontanus, pero por la reducción y desaparición de las poblaciones de esta especie de sus áreas naturales, no se incluyó en el estudio.
2.1. Estudio de campo
Se realizaron ocho colectas mensuales en el área descrita para la captura de los cangrejos de río, utilizando trampas cilindricas con dos entradas similares (Fig. 2) a las descritas por Huner y Barr (1984); previamente a esto se hicieron pruebas de trampeo para determinar el tipo de cebo más adecuado, seleccionadose el hígado de res, entre otros cebos que incluían, visceras de pollo, carne de pescado, "cabezas" de camarón, alimento para perro tipo croqueta y alimento balanceado para camarón tipo "fideo". Se utilizó un total de 10 trampas durante todo el estudio, estas se colocaron en un transecto de 150 metros a lo largo del lecho del río durante la noche por un período mínimo de tres horas, posteriormente se procedió a la separación de los organismos capturados por trampa, para ser llevados al