Rasgos funcionales y plasticidad fenotípica de plantas exóticas invasoras
138
0
0
Texto completo
(2) Palabras clave: Especies exóticas invasoras, rasgos funcionales, plasticidad fenotípica, eficiencia de la plasticidad, integración fenotípica, luz, nutrientes, hojarasca, ciclo de nutrientes, fenología de floración, eficacia biológica, comparaciones filogenéticas, modelos de ecuaciones estructurales, estadística circular..
(3) Abstract This PhD thesis addresses the study of the functional traits and phenotypic plasticity that are associated to the invasibility of alien plant species and their impact in mediterranean ecosystems. The main objectives were: i) to search for differences between invasive and native phenotypes for an ample group of morphological and physiological traits, ii) to determine whether high phenotypic plasticity is associated to the invasibility of alien plant species, iii) to establish whether leaf traits that are related to the performance of invasive species influence nutrient cycling through litter decomposition, and iv) to study whether the flowering phenology differences between invasive and native species is a trait that confers invasiveness. Morphological and physiological responses of 31 traits to two gradients of light and nutrient availability were studied in 20 invasive-native pairs (chapter 2). For the third objective, initial litter trait, litter decomposition rates and the relative importance of initial litter traits in litter decomposition were compared within 19 invasive-native pairs (chapter 3). Finally, we compared the flowering phenology of invasive and native species in three mediterranean-type ecosystems searching the flowering phenology data of 227 invasive-native pairs (chapter 4 and 5). Results showed that high light and nitrogen use efficiencies confer invasibility to alien species, resulting in a enhance fitness when they are grown in light and nutrient gradients. However the water use efficiency of alien species was low, which could limit their invasion of Spanish mediterranean ecosystems to areas where the abiotic filter of summer drought is minimised. Contrary to expectations, invasive species displayed the same or even lower levels of phenotypic plasticity compared to natives. However, a highly competitive phenotype allows invasive species to obtain a higher fitness increment per unit of plastic response compared to natives. Therefore, high plasticity efficiency seems to be a trait that confers invasibility to alien plant species. Invasive species had higher lignin content in leaf litter. Invasive species were more nitrogen- and phosphorus-dependent for their decomposition due to their higher lignin content. Because lignin is difficult to degrade, the rate of litter decomposition was lower in invasive species compared to natives, suggesting that under natural conditions nutrient cycling can be slow down upon invasion. The later flowering phenology of invasive species compared to natives in Spanish mediterranean ecosystems is not a common pattern in other mediterranean-type ecosystems such as the Cape Region of South Africa and California, where invasive species flower earlier or at the same time than natives, respectively. Thus, a different flowering phenology between invasive and native species is not a trait associated with invasiveness. However, it determines the most likely growth form of invasive species. In this sense, summer-flowering limits the invasion of alien species to habitats with water availability during summer where fast-growing herbaceous are adapted to live. Key words: Invasive alien species, functional traits, phenotypic plasticity, plasticity efficiency, light, nutrient, litter, nutrient cycling, flowering phenology, fitness, phylogenetic comparisons, structural equation modelling, circular statistics..
(4) ..
(5) Índice Abstract Resumen Capítulo 1. Introducción general...................................................................................................................1 Capítulo 2. Phenotypic plasticity of exotic invasive plant species compared to native species Resumen en castellano........................................................................................................... 17 2.1. Introduction.............................................................................................................................. 21 2.2. Material and methods.............................................................................................................. 23 2.3. Results..................................................................................................................................... 32 2.4. Discussion............................................................................................................................... 38 2.5. Conclusions............................................................................................................................. 44 Capítulo 3. Leaf litter traits of invasive alien species slow down decomposition compared to Spanish native: a broad phylogenetic comparison Resumen en castellano........................................................................................................... 59 3.1. Introduction.............................................................................................................................. 62 3.2. Material and methods.............................................................................................................. 63 3.3. Results..................................................................................................................................... 66 3.4. Discussion............................................................................................................................... 69 Capítulo 4.Different flowering phenology of alien invasive species in Spain: evidence for the use of an empty temporal niche? Resumen en castellano........................................................................................................... 79 4.1. Introduction.............................................................................................................................. 81 4.2. Material and methods.............................................................................................................. 84 4.3. Results..................................................................................................................................... 85 4.4. Discussion............................................................................................................................... 88 Capítulo 5. Flowering phenology of invasive alien plant species compared with native in three Mediterranean-type ecosystems Resumen en castellano.......................................................................................................... 101 5.1. Introduction.............................................................................................................................. 104 5.2. Material and methods.............................................................................................................. 105 5.3. Results..................................................................................................................................... 108 5.4. Discussion............................................................................................................................... 111 Capítulo 6. Conclusiones generales y líneas futuras.................................................................................. 131 Apéndice 1 ................................................................................................................................................ 141 Apéndice 2 ................................................................................................................................................ 147.
(6) ..
(7) Capítulo 1 Introducción general Se denomina especie invasora a aquella que o bien de manera natural o bien por la acción del ser humano se ha introducido en nuevas regiones donde antes no existía, siendo capaz de reproducirse por ella misma y llegando a formar un conjunto de poblaciones estables con tendencia expansiva (Richardson et al. 2000). Las especies invasoras con mayor relevancia para los ecosistemas nativos son aquellas que tienen la capacidad de modificar los diferentes niveles de organización y estructura de la biosfera, desplazando a la vegetación nativa (Rejmanek et al. 1999), modificando el ciclo del agua, de nutrientes, el régimen de perturbaciones (Brooks et al. 2004; Ehrenfeld 2003; Mack et al. 2000) y alterando la estructura y el funcionamiento de los ecosistemas (Dukes y Money 2004). Lejos de disminuir, el impacto negativo sigue aumentando por el creciente número de nuevas especies invasoras que llegan a nuestros ecosistemas (Hobbs et al. 2006) y amplificándose por las sinergias que resultan de la interacción con otros motores del cambio global (Dukes y Mooney 1999; Theoharides y Dukes 2007). De hecho, a las especies invasoras se les considera junto a la pérdida de hábitat los dos motores del cambio global que más han influido en la acelerada tasa actual de extinción de especies (Mack 1996; Mack 2003; Williamson 1996). Por tanto, identificar los mecanismos que determinan el potencial invasor de las especies exóticas es fundamental para establecer protocolos de control y planes de gestión, cómo así lo indican los programas internacionales de conservación de la biodiversidad (UICN, GSIP, FAO). La introducción de especies exóticas en la Cuenca Mediterránea no es un fenómeno nuevo (Groves y Di Castri 1991). El comercio desde la antigüedad ha transportado especies vegetales que ya consideramos como nativas después de más de dos mil años de introducción. Tales son los casos del pino piñonero (Pinus pinea), el algarrobo (Ceratonia siliqua), o la higuera (Ficus carica). El año 1492 supuso un nuevo referente con el descubrimiento de América (Fig. 1a): el número de especies exóticas introducidas aumentó brucamente y a partir de este momento se establece una clasificación arbitraria entre arqueófitos (introducidas antes de 1500) y neófitos (introducidas después de 1500). El interés comercial por las nuevas especies descubiertas introdujo de forma voluntaria en España algunos neófitos invasores como la opuntia (Opuntia ficus-indica), el ágave (Agave americana), el jacinto de agua (Eichhornia crassipes) o la lantana (Lantana camara), mientras que en otros casos como la ageratina (Ageratina adehophora), varias especies de amaranto (Amaranthus albus, A.hybridus, A.muricatus, A.retroflexus) o la coniza (Conyza bonariensis) la introducción fue involuntaria (Sanz Elorza et al. 2004). Sin embargo, no es hasta principios del siglo XX cuando el comercio a nivel global genera tales modificaciones en el planeta que algunos lo consideran como el inicio de una nueva era, el Antropoceno (Fig. 1b, Kareiva et al. 2007). Así pues hoy en día resulta indiscutible que la acción humana ha aumentado de forma exponencial el movimiento de especies y materias primas a todos los lugares del planeta, produciendo lo que algunos autores llaman un proceso global de homogeneización biótica (Lockwood et al. 2007; Olden 2005; Olden y Rooney 2006). En este proceso se estima que han sido introducidas más de 10.000 especies invasoras de plantas, 5000 de invertebrados, 1000 de microorganismos y 300 de vertebrados (DAISIE 2006; Pimentel 2002).. 1.
(8) Capítulo 1. Figura 1a: Rutas comerciales a nivel mundial tras el descubrimiento de América.. Figura 1b: Rutas comerciales marítimas y terrestres en la actualidad (Kareiva et al. 2007). Puede observarse como el comercio entre Europa, América y Asia es el mayoritario aunque otras rutas como las que cruzan el pacífico y el atlántico sur también son importantes.. 2.
(9) Introducción general. La evaluación ecosistémica del milenio (Millenium Ecosystem Assessment, United Nations, 2005) revela que como resultado de la presente globalización el elevado número de especies invasoras liberadas en nuevas regiones ha tenido un impacto importante en general y sobre los ecosistemas mediterráneos en particular. Este impacto ha sido muy importante en los últimos 50-100 años y la tendencia actual es a incrementarse significativamente (Fig. 2). Pero predecir el incremento futuro es difícil, ya que la interacción de las especies invasoras con los ecosistemas mediterráneos raramente es aditiva sino sinérgica, y en cualquier caso muy compleja (Lavorel et al. 1998; Sala et al. 2000). Por ejemplo, el efecto conjunto del cambio climático y los cambios de uso del suelo pueden disparar el impacto de las especies invasoras (Peters et al. 2006; Rejmánek et al. 2005; Vilà et al. 2003). Sin embargo, la creciente concienciación de los problemas derivados de las especies invasoras podría ayudar a mitigar sus impactos tanto ecológicos como económicos en el futuro. De hecho, existe un gran número de programas nacionales e internacionales que tienen por objeto mejorar el conocimiento sobre las especies invasoras, y con él aumentar la eficacia de su control y, en la medida de lo posible, erradicación y prevención de futuras introducciones. Cabe destacar a nivel internacional GISP (Global Invasive Species Program, www.gisp.org), a nivel europeo DAISIE (Delivering Alien Invasive Species Inventories for Europe, www.europe-aliens.org), en Sudáfrica Working for Water Program (www.dwaf.gov.za/wfw/), en Estados Unidos Centre for Invasive Species and Ecosystem Health (www.invasive.org), y en California CAL-IPC (California Invasive Plant Council, www.cal-ipc.org). Si se traducen estas políticas a costes económicos poco se conoce con detalle excepto que los costes son muy elevados (Pimentel et al. 2001; Pimentel et al. 2003). En Estados Unidos, se ha calculado que el gasto para controlar las especies invasoras asciende a 120 billones de dólares por año (Pimentel et al. 2000; Pimentel et al. 2003) y se tiene constancia de que muchas de estas especies son vectores de enfermedades peligrosas para el ser humano, como la gripe aviar, la malaria o la tuberculosis (Pimentel et al. 2007). Por eso no es de extrañar que cada vez más se computen los costes asociados a las especies exóticas invasoras en el diseño de políticas económicas (Binimelis et al. 2007; Maguire 2004; Zavaleta 2000).. Etapas del proceso de invasión Las invasiones biológicas son el resultado de un proceso espacio-temporal continuo a nivel global que implica una serie de fases, que son en esencia transporte, introducción, establecimiento y propagación sobre las nuevas áreas invadidas (Richardson y Pyšek 2006; Richardson et al. 2000; Sakai et al. 2001) (Fig. 3). Para que una planta se considere exótica en una nueva región primero tiene que sobrepasar la barrera geográfica que impide su dispersión. Aunque hay especies que han demostrado ser capaces de dispersar propágulos a larga distancia (Mack 2004; Vermeij 2005), es la acción humana la que principalmente está rompiendo esta barrera (Lockwood et al. 2007). Esta acción humana hace que las tasas de transporte estén siendo más aceleradas y de mayor alcance que los procesos naturales (Brown y Sax 2004; Cassey et al. 2005; Lockwood et al. 2007; Vermeij 2005), influyendo directamente en los patrones geográficos de invasión a gran escala de muchas especies exóticas (Thuiller et al. 2006).. 3.
(10) Capítulo 1. Figura 2: Entre los distintos motores de cambio global se específica el impacto pasado y actual de las especies invasoras sobre los ecosistemas. Un impacto alto o muy alto (tono de color verde oscuros) implica que las especies invasoras han alterado significativamente la biodiversidad en los últimos 50 a 100 años. Modificado de Millenium Ecosystem Assessment (2005).. 4.
(11) Introducción general. Una vez introducidas, las plantas exóticas tienen que hacer frente a la barrera ambiental que les impone el nuevo ecosistema. Estas barreras se componen de dos tipos de filtros: abióticos y bióticos. Los filtros abióticos severos (escasez de recursos, condiciones ambientales extremas) reducen la probabilidad de supervivencia de las especies exóticas introducidas, mientras que los más benignos favorecen su establecimiento. Aunque hay regiones donde la vida vegetal es difícil (p. ej. desiertos, regiones polares), una especie exótica encontrará mayor o menor dificultad de asentamiento dependiendo del grado de similitud entre el clima en la región de origen y la introducida. Pero también hay evidencias de plantas exóticas capaces de sobrevivir en condiciones ambientales que nunca antes habían experimentado. Diversos mecanismos como una elevada plasticidad fenotípica, una rápida evolución o la combinación de ambos (Dudley 2004) se han propuesto para tratar de explicar esta capacidad de sobrevivir en ambiente nuevos. La plasticidad fenotípica, o la capacidad de un genotipo/especie para dar lugar a una serie de fenotipos diferentes según el ambiente (Pigliucci 2001; Sultan 2000; Valladares et al. 2000), favorece la capacidad de persistencia de las plantas introducidas en las primeras fases de invasión, ya que permite adaptar el fenotipo a una diversidad de ambientes. Además, una elevada plasticidad fenotípica aumenta la probabilidad de supervivencia ante fenómenos estocásticos de extinción (Ghalambor et al. 2007; Sultan 2001). El mayor o menor grado de plasticidad fenotípica está en parte positivamente relacionado con la amplitud climática en la zona de origen de las plantas exóticas (Atkin et al. 2005; Williams et al. 1995; Willis y Hulme 2002). Por otra parte, una rápida evolución frente a las presiones de selección que impone el ambiente, también favorece una rápida adaptación al medio (Agrawal 2001; Donohue et al. 2005; Yeh y Price 2004) y permite una expansión acelerada en la nueva región invadida, lo cual ya ha sido observado en algunas plantas invasoras (Maron et al. 2004).. Fig. 3: Modelo conceptual de las diferentes barreras que las especies exóticas tienen que sobrepasar para ser ocasionales, naturalizadas o invasoras en la nueva región donde se han introducido. Modificado de Richardson et al. (2006).. 5.
(12) Capítulo 1. Alternativamente, la presión de propágulos se ha propuesto como la forma más sencilla de explicar la invasión en una nueva región. Bajo este marco conceptual la respuesta de las plantas exóticas al ambiente queda en segundo plano y es la acción humana mediante la capacidad para introducir grandes cantidades de propágulos la que tendría mayor importancia (Colautti et al. 2006). Se cree que una planta exótica introducida será una invasora exitosa cuanto mayor sea la presión de propágulos (Levine y D'Antonio 1999; Levine et al. 2003). Por un lado, a mayor número de individuos introducidos menor será la probabilidad de una extinción local. Y por otro lado, la probabilidad de introducir un genotipo que se adapte mejor a las nuevas condiciones ambientales será también mayor. La presión de propágulos varía según el vector de transporte utilizado por el hombre, pero en general tiende a ser mayor en aquellas áreas donde se concentran las rutas de transporte y donde la cantidad de mercancías transportadas es elevada (Kareiva et al. 2007; Lockwood et al. 2007). En cuanto a los filtros bióticos, los componentes que más influyen en el éxito de las plantas exóticas son la competencia y los herbívoros (Levine et al. 2004). Las plantas exóticas pueden ver limitada su supervivencia al no tener defensas adaptadas a los nuevos herbívoros que componen la comunidad de la región invadida, pero el razonamiento puede ser el contrario y las especies exóticas pueden ver superada la barrera biótica al escapar de los enemigos naturales que afectaban a sus poblaciones en el lugar de origen (Vilà et al. 2005; Wolfe 2002). A largo plazo, esta situación puede propiciar un cambio evolutivo, fomentando una menor inversión de recursos en defensas frente a los herbívoros y competidores en favor de un mayor crecimiento y reproducción (Joshi y Vrieling 2005). Cabe destacar que los ambientes productivos son los que aceleran ese cambio evolutivo rápido (Blumenthal et al. 2009). El siguiente paso para que una planta exótica se convierta en invasora es reproducirse por si sola y dispersarse a nuevas áreas. Plantas con flores de polinización generalista y/o autocompatibles, abundante producción de néctar y polen y largos periodos de floración tienen mayor éxito reproductivo (Traveset y Richardson 2006). Una elevada reproducción aumenta la presión de propágulos y por tanto la persistencia a largo plazo. Asociado a la reproducción de las plantas exóticas, las plantas nativas sufren fenómenos de disrupción de sus interacciones bióticas con los polinizadores. Como resultado, las plantas nativas ven reducido su éxito reproductivo porque disminuye el número de semillas producidas, ocurren cambios en el flujo génico y se dan fenómenos de hibridación (Moragues y Traveset 2005; Traveset y Richardson 2006), siendo estos procesos especialmente importantes en aquellas especies nativas que se asemejan fenotipicamente a las invasoras (Morales y Traveset 2009). Para que ocurran estos procesos de competencia tanto especies invasoras como nativas tienen que florecer al mismo tiempo. Sin embargo, muchas especies invasoras florecen antes que las nativas (Pyšek et al. 2003) mientras que otras después (Celesti-Grapow et al. 2003; Lloret et al. 2005). Esta diferencia en el periodo de floración se ha entendido como una estrategia adaptativa de la especies invasoras para evitar la competencia con las especies nativas por los polinizadores, pero este argumento aún queda muy especulativo porque no se ha comprobado experimentalmente si esta hipótesis es correcta o si influyen otros aspectos en el periodo de floración, como la historia de introducciones, el hábitat invadido o las condiciones climáticas en el lugar de origen. En general, a no ser que las presiones de selección en el lugar invadido hayan sido tan importantes como para que haya ocurrido cambio evolutivo, se puede hipotetizar que las plantas exóticas 6.
(13) Introducción general. florecen en función de los filtros bióticos y abióticos del lugar de origen. Por tanto, especies invasoras con diferentes presiones de selección sobre la floración en sus lugares de origen, poseerán fenologías de floración diferentes. Distinguir que procesos hay detrás de la fenología de floración de las especies invasoras se revela como un hecho importante para predecir si habrá competencia o no por la reproducción entre especies invasoras y nativas. En cuanto a la dispersión, la literatura señala que ésta es más exitosa en las especies exóticas ligadas a la acción humana. Estas especies exóticas transportadas a regiones nuevas se consideran naturalizadas (Pyšek et al. 2004), pero no serán consideradas como invasoras hasta que muestren una expansión sobre el territorio por si mismas tanto en los habitats perturbados como en los naturales.. Ecosistemas invadidos En general, las especies invasoras ocupan primeramente los hábitats perturbados porque son más fáciles de invadir y de ahí pueden saltar a los hábitats naturales con menor perturbación (Fig. 3). En el caso de España, al igual que en muchas otras partes del mundo, es en estos hábitat perturbados (bordes de carreteras, caminos, vías férreas, campos de cultivo, riberas perturbadas) donde la riqueza de especies invasoras es mayor (Gassó et al. 2009; Lake y Leishman 2004; MacDougall y Turkington 2005). La razón de porqué los hábitats perturbados son más invadidos que los no perturbados tiene que ver con la mayor presión de propágulos que se registran en esos ecosistemas, con la mayor disponibilidad del espacio y los recursos, y con la menor competencia con las especies nativas (Davis y Pelsor 2001; Levine y D'Antonio 1999; Lockwood et al. 2007; Recasens et al. 2005; Vilà et al. 2003). Cambios rápidos en el uso del suelo modifican las presiones de selección y por tanto generan oportunidades de asentamiento facilitando además la expansión por el territorio de las especies exóticas (Theoharides y Dukes 2007). La proporción de especies introducidas que llegan a naturalizarse y de ahí a invadir depende de los rasgos de las plantas exóticas, de las características del ecosistema receptor y de la frecuencia de las introducciones. En general, la combinación de rasgos biológicos que confieren una elevada eficacia biológica a las plantas invasoras y regiones que han sufrido mayor número de introducciones son las que también contienen mayor número de especies invasoras entre su flora (Bergstrom y Chown 1999; Lockwood et al. 2007). Además, las islas son ecosistemas menos resistentes a la invasión que las zonas continentales por su menor resistencia biótica. Hasta hace poco se estimaba que según la regla del 10% (Williamson y Fitter 1996), solo el 10% de las especies introducidas llegaban a naturalizarse y de ese 10% solo otro 10% llega a ser invasor. Sin embargo estudios más en detalle varían considerablemente esas proporciones. Por ejemplo en el caso de los mamíferos más del 60% de las especies introducidas llegan a ser invasoras mientras que en el caso de las plantas exóticas ornamentales esa cifra es menor del 1% (DAISIE 2006; Vilà y Pujadas 2001). Sin embargo, una vez que la especie invasora está ampliamente distribuida, lo importante es saber el impacto que va a producir sobre los ecosistemas (Levine et al. 2003). La intensidad del impacto de la planta invasora está relacionada con los rasgos que le han conferido invasibilidad y con el grado de diferencia que muestran con la flora nativa. Por tanto, estudiar los rasgos de las plantas 7.
(14) Capítulo 1. invasoras no solo es importante para predecir qué especies introducidas van a ser invasoras sino también para saber qué impactos sobre el ecosistema van a provocar las especies invasoras. Un nexo claro entre rasgos e impactos es el estudio de la influencia de la descomposición de hojarasca sobre el ciclo de nutrientes (Ehrenfeld 2003).. Rasgos de las especies de plantas invasoras Siguiendo los primeros pasos de Baker en 1965, muchos ecólogos han tratado de discernir cuales son los rasgos que confieren invasibilidad a las plantas exóticas. Para ello se ha utilizado el método comparativo, discerniendo qué rasgos de la flora invasora no están presentes en la flora nativa o en la flora exótica no invasora, e implícitamente se ha asumido que el patrón observado en esas diferencias es el que explica el potencial invasor de las especies exóticas. Sin embargo, existen numerosos sesgos en estos estudios que pueden condicionar los patrones observados. Entre los sesgos más comunes se puede citar el tiempo de residencia que lleva una especie exótica en el nuevo ecosistema, su historia evolutiva, las presiones selectivas en el lugar de origen, la facilidad de transporte, la razón de la introducción y la relación filogenética entre las especies (Pyšek y Jarošík 2005; Pyšek y Richardson 2007; Wilson et al. 2007). Hasta el momento, la mejor recopilación de rasgos que confieren invasibilidad a las plantas exóticas es la elaborada por Pyšek y Richardson (2007) (Fig. 4). En ella se condensan 59 estudios donde se han tenido en cuenta las relaciones filogenéticas entre especies.. Figura 4: Resumen de 59 estudios donde se han comparado características vegetales entre pares de especies filogenéticamente próximas, donde cada par estaba constituido por una especie invasora y una nativa o una exótica con bajo o nulo grado de potencial invasor. Entre paréntesis se señala el número de estudios recopilados para cada rasgo. Modificado de Pyšek y Richardson (2007) en el libro Biological Invasión Ed: Caldwell, MM.. 8.
(15) Introducción general. En general, se observa que las especies invasoras son más competitivas y fértiles que las nativas y que las especies exóticas no invasoras. Más allá de estos resultados esperables, lo que pone en relevancia la revisión de Pyšek y Richardson (2007) es que los rasgos involucrados en esa mayor eficacia biológica no son siempre los mismos, lo que señala que no hay una única estrategia que explique la capacidad invasora de las especies exóticas. Valga como ejemplo que los rasgos que confieren potencial invasor a una especie de acacia australiana en un clima mediterráneo de Europa seguramente no son los mismos que una gramínea americana invasora en la sabana africana debido a las formas de crecimiento y estrategias de vida distintas. En la naturaleza, las plantas al ser organismos sésiles tienen que soportar una amplia variedad de condiciones ambientales para sobrevivir tanto en el espacio como en el tiempo. Por tanto, más allá de poseer uno y otro rasgo, lo que se necesita conocer es el conjunto de rasgos que permite a las especies invasoras adaptarse a una serie de condiciones ambientales muy diversas. Se puede hipotetizar que aquellas especies exóticas que soporten mejor la variabilidad ambiental mostrando siempre una elevada eficacia biológica serán muy probablemente las que mayor éxito invasor tengan. Sin embargo, existe una gran falta de conocimiento en este aspecto ya que no existen hasta la fecha trabajos en los que se estudien los rasgos morfológicos y ecofisiológicos adaptativos de un elevado número de especies invasoras a gradientes ambientales amplios y se relacionen explícitamente con una mayor capacidad competitiva y una mayor eficacia biológica de las especies invasoras respecto a sus análogas nativas. Como mecanismo alternativo pero no excluyente, las especies invasoras pueden también poseer un elevado potencial invasor mostrando una elevada plasticidad fenotípica. En este caso no es el valor medio de los rasgos lo que le permite tolerar el ambiente, sino la capacidad de variación de los rasgos. Teóricamente se cree que cuanto mayor sea las plasticidad fenotípica de las especies mayor será su capacidad de adaptación al ambiente (Alpert et al. 2000; Richards et al. 2006), pero esa plasticidad tiene unos límites tanto internos a la propia planta, como externos, debidos al ambiente en el que se encuentra la planta (Valladares et al. 2007; van Kleunen y Fischer 2005). Experimentalmente, de los trabajos realizados hasta la fecha no se puede concluir si esta teoría sobre las ventajas de una mayor plasticidad fenotípicas es correcta o no. Mientras algunos trabajos muestran una mayor eficacia biológica de las especies invasoras gracias a su mayor plasticidad fenotípica (Gerlach y Rice 2003; Niinemets et al. 2003; Richards et al. 2006) otros no (Funk 2008; Peperkorn et al. 2005). La diferencia entre resultados puede ser debida a razones ecológicas ya que la plasticidad fenotípica puede ser importante solo bajo determinadas condiciones ambientales, o a razones metodológicas porque el número de especies estudiadas ha sido muy bajo como para tener suficiente poder estadístico para observar un patrón significativo. Menos aún se sabe de la relación entre tolerancia al estrés y plasticidad, y de la posible limitación que la integración fenotípica pueda inducir sobre la plasticidad, a pesar de ser cruciales para entender de manera realista el potencial invasor de las especies exóticas. La tolerancia al estrés y la plasticidad están muy relacionadas, ya que el valor medio de los rasgos adaptativos en un ambiente y su plasticidad covarían (Valladares et al. 2007). En ese sentido una especie invasora no necesitaría ser muy plástica si ya posee un fenotipo muy competitivo en un determinado ambiente, lo cual le permite evitar a su vez los costes de plasticidad que pueda ser maladaptativa. En este sentido una especie con un fenotipo muy competitivo tendría una mayor eficiencia de la plasticidad si por un cambio fenotípico dado en un rasgo adaptativo, incrementa más su eficacia 9.
(16) Capítulo 1. biológica que otra. Por tanto, un estudio amplio midiendo el valor medio de un número de rasgos elevados su valor medio y la plasticidad de esos rasgos en especies invasoras y nativas permitiría comprender mejor el fenómeno de las invasiones biológicas.. Objetivos de la Tesis Doctoral Esta Tesis Doctoral se encuadra dentro del seno del proyecto RINVE (Determinantes biológicos del riesgo de invasiones vegetales, CGL2004-04884/BOS) y más especificamente dentro del subproyecto RASINV (Rasgos de las especies invasoras, CGL2004-04884-C02-02) cuyos objetivos eran identificar los factores que determinan el riesgo de invasiones biológicas en comunidades vegetales, prestando particular atención tanto a los determinantes del potencial invasor de las especies exóticas como a su modulación por los elementos que determinan la invasibilidad de los ecosistemas receptores. La presente Tesis Doctoral tiene por objetivo conocer la importancia relativa de determinados rasgos funcionales y de su plasticidad fenotípica en el potencial invasor de las plantas exóticas, así como investigar el impacto de estos rasgos diferenciales sobre el funcionamiento de los ecosistemas. El estudio se enmarca en ecosistemas mediterráneos ibéricos, comparando un elevado grupo de especies invasoras y nativas de diversas formas de crecimiento y hábitat invadido (Fig. 5).. Figura 5: Algunas especies exóticas catalogadas como invasoras en España. De izquierda a derecha Acacia melanoxylon, Ipomoea indica, Schinus molle y Lantana camara.. Estructura de la Tesis Doctoral y objetivos específicos de cada capítulo Esta Tesis Doctoral se estructura en torno a cuatro capítulos, que se corresponden con los correspondientes artículos científicos, que han sido escritos originalmente en inglés para su publicación en revistas de ámbito internacional. Además de esta introducción, se incluye un capítulo de conclusiones generales y consideraciones adicionales de líneas futuras de investigación a partir de los resultados obtenidos más dos apéndices relacionados con algunas técnicas estadísticas utilizadas. A continuación se enumeran los objetivos específicos de la Tesis Doctoral: 1. Determinar la importancia de los rasgos funcionales y la plasticidad fenotípica que confieren potencial invasor a las especies de plantas exóticas. 10.
(17) Introducción general. En el capítulo 2 se evalúa si el éxito de las especies invasoras está relacionado con una elevada plasticidad fenotípica, la cual se espera que sea mayor que en las especies nativas. Para ello, se comparó mediante una aproximación experimental la respuesta de 31 rasgos morfológicos y fisiológicos de 20 pares de especies invasoras y nativas a dos amplios gradientes de luz y nutrientes. Además mediante el empleo de modelos de ecuaciones estructurales se determinó para ambos grupos: i) bajo qué condiciones ambientales la plasticidad fenotípica es el factor principal de la eficacia biológica de las especies invasoras, ii) cuáles son los límites internos y externos a la plasticidad y iii) si los resultados obtenidos difieren entre especies invasoras y nativas. Este trabajo aborda la relación entre plasticidad fenotípica y potencial invasor de las especies exóticas dentro de un contexto más amplio, incluyendo otros aspectos tan relevantes como la relación entre los valores medios del fenotipo, la eficiencia de la plasticidad y la integración fenotípica como posible límite a la expresión plástica. 2. Analizar si existen rasgos diferentes entre especies invasoras y nativas que influyan posteriormente en el ciclo de nutrientes a través de la descomposición de hojarasca. En el capítulo 3 se evalúa el impacto que producen las especies invasoras en el ciclo de nutrientes a través de la descomposición de hojarasca. Es esperable que si hay diferencias en los rasgos funcionales entre las especies invasoras y nativas estos queden reflejados en la hojarasca e influyan en el proceso de su descomposición, afectando a su vez al ciclo de nutrientes a nivel de comunidad. De nuevo, una aproximación experimental sirvió para caracterizar los rasgos físicoquímicos iniciales y las tasas de descomposición de la hojarasca de 19 pares de especies invasora-nativa. Por otra parte, la clasificación de modelos de regresión según la teoría de información ayudó a discernir qué rasgos iniciales influyeron en las diferencias de descomposición observadas entre especies invasoras y nativas. 3. Estudiar los factores evolutivos, ecológicos, humanos e históricos que determinan la fenología de floración de las especies invasoras en ecosistemas mediterráneos a nivel mundial. La evaluación de la fenología de floración en un grupo muy amplio de especies invasoras y nativas a nivel de ecosistemas mediterráneos de todo el planeta ha derivado en la búsqueda bibliográfica de los datos necesarios para realizar dos trabajos que componen dos capítulos. En el capítulo 4 se analiza si la fenología de floración es un rasgo de las especies invasoras que conservan del lugar de origen o por el contrario responde a las nuevas condiciones ambientales del lugar invadido. El trabajo indaga en los mecanismos que determinan el periodo de floración de las especies invasoras y explora la importancia de tener en cuenta las propiedades del ecosistema invadido para esclarecer por qué las especies invasoras en España florecen en un periodo distinto que las especies nativas. En el capítulo 5 se analiza si el diferente patrón de floración encontrado entre especies invasoras y nativas en España se mantiene en otros dos ecosistemas mediterráneos del mundo (California y la Region del Cabo de Sudáfrica), como adaptación a los filtros abióticos que impone la nueva región invadida. El trabajo nos ayuda a entender la importancia de la acción humana en los rasgos funcionales que muestran las especies invasoras y la interacción entre la historia de introducciones y las condiciones ambientales del lugar invadido. Una vez desarrollados los objetivos, en el capítulo 6 se enumeran las conclusiones más relevantes en función de de su carácter metodológico, teórico y de aplicación del conocimiento 11.
(18) Capítulo 1. adquirido. Posteriormente se discuten las líneas futuras que se derivan de los resultados obtenidos de esta memoria de Tesis Doctoral. Ya por último y como anexo, en los apéndices 1 y 2 se detallan el uso de los modelos de ecuaciones estructurales y la estadística circular utilizados como herramientas estadísticas en los trabajos de investigación. Aspectos relevantes de esta Tesis Doctoral Comparación de un grupo de especies amplio El gran número de especies invasoras y nativas comparadas en los trabajos de esta Tesis Doctoral refuerzan los resultados obtenidos y lo diferencian significativamente de estudios anteriores. Este amplio número de especies permite apoyar con seguridad estadística las conclusiones obtenidas de los resultados y suponen una novedad frente a anteriores trabajos experimentales en los que el número de especies estudiadas fue siempre muy bajo (no superando nunca 10 pares de especies invasora-nativa y estando en general en torno a 4-5) (Pyšek y Richardson 2007). Este hecho supone principalmente dos ventajas. Por un lado, mejora el conocimiento de las invasiones biológicas al aumentar la probabilidad de encontrar patrones de diferencias entre especies invasoras y nativas. Y por otro lado, reduce la probabilidad de que las diferencias no significativas entre especies invasoras y nativas sean debidas a una falta de poder estadístico de los análisis.. Relaciones filogenéticas y ecológicas entre las especies En el estudio comparativo de especies es necesario tener en cuenta las relaciones filogenéticas, ya que los resultados obtenidos pueden no estar relacionados directamente con las diferencias ecológicas entre especies sino más bien con sus diferencias filogenéticas (Felsenstein 1985). Pero a su vez en este enfoque filogenético no se debe perder el sentido ecológico para poder realizar conclusiones ecológicas de los resultados (Lambdon y Hulme 2006). En todos los trabajos de esta Tesis Doctoral se han tenido en cuenta las relaciones filogenéticas entre las especies tanto a la hora de seleccionar las especies invasoras y nativas de estudio como a la hora de aplicar las herramientas estadísticas necesarias para tener en cuenta esas relaciones de parentesco. Además, no se ha perdido en enfoque ecológico al realizar los diseños experimentales teniendo en cuenta las relaciones ecológicas entre especies. Enfoque mecanicista Aunque las diferencias entre los rasgos funcionales de las especies invasoras y nativas pueden estar relacionados con el potencial invasor de las especies exóticas, no siempre es así (Lockwood et al. 2007). Muchos de los rasgos funcionales de las especies exóticas pueden tener un valor nulo en su potencial invasor o incluso ser una desventaja para invadir nuevos ambientes. Por tanto, inferir causalidad entre potencial invasor y diferencias en rasgos entre especies invasoras y nativas puede dar lugar a conclusiones erróneas. Para evitar ese problema se ha elegido un enfoque más conservador en el cual primero se ha comprendido el funcionamiento y los mecanismos dinámicos de los rasgos estudiados y luego se han interpretado dentro del marco ecológico de las invasiones biológicas.. 12.
(19) Introducción general. Bibliografía Agrawal AA (2001) Ecology: Phenotypic plasticity in the interactions and evolution of species. Science 294:321-326. Alpert P, Bone E, Holzapfel C (2000) Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspectives in Plant Ecology, Evolution and Systematics 3:52-66. Atkin OK, Loveys BR, Atkinson LJ, Pons TL (2005) Phenotypic plasticity and growth temperature: understying interspecific variability. In: Meeting on Phenotypic Plasticity and the Changing Environment held at the Society for Experimental Biology Plant Frontiers Meeting, Sheffield, ENGLY, pp 267-281. Bergstrom DM, Chown SL (1999) Life at the front: history, ecology and change on southern ocean islys. Trends in Ecology & Evolution 14:472-477. Binimelis R, Monterroso I, Rodriguez-Labajos B (2007) A Social Analysis of the Bioinvasions of Dreissena polymorpha in Spain and Hydrilla verticillata in Guatemala. Environmental Management 40:555-566. Blumenthal D, Mitchell CE, Pysek P, Jarosik V (2009) Synergy between pathogen release and resource availability in plant invasion. Proceedings of the National Academy of Sciences of the United States of America 106:7899-7904. Brooks ML et al. (2004) Effects of invasive alien plants on fire regimes. Bioscience 54:677-688. Brown JH, Sax DF (2004) An essay on some topics concerning invasive species. Austral Ecology 29:530-536. Cassey P, Blackburn TM, Duncan RP, Chown SL (2005) Concerning invasive species: Reply to Brown and Sax. Austral Ecology 30:475-480. Celesti-Grapow L, Di Marzio P, Blasi C (2003) Temporal niche separation of the alien flora of Rome. In: Child LE et al. (eds) Plant Invasions: Ecological Threats and Management Solutions. Backhuys, Leiden, pp 101111. Colautti R, Grigorovich I, MacIsaac H (2006) Propagule Pressure: A Null Model for Biological Invasions. Biological Invasions 8:1023-1037. DAISIE (2006) Delivering alien invasive inventories for Europe. EU project 6th FP. www.europe-aliens.org Davis MA, Pelsor M (2001) Experimental support for a resource-based mechanistic model of invasibility. Ecology Letters 4:421-428. Donohue K et al. (2005) Environmental and genetic influences on the germination of Arabidopsis thaliana in the field. Evolution 59:740-757. Dudley SA (2004) Plasticity y the Functional Ecology of Plants In: De Witt TJ, Scheiner SM (eds) Phenotypic Plasticity: Functional and Conceptual Approaches. Oxford University Press. Dukes JS, Mooney HA (1999) Does global change increase the success of biological invaders? Trends in Ecology & Evolution 14:135-139. Dukes, J.S. & Mooney, H.A. (2004) Disruption of ecosystem processes in western North America by invasive species Revista Chilena de Historia Natural 77: 411-437. Ehrenfeld JG (2003) Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 6:503523. Felsenstein J (1985) Phylogenies and the Comparative Method. American Naturalist 125:1-15. Funk JL (2008) Differences in plasticity between invasive and native plants from a low resource environment. Journal of Ecology 96:1162-1173. Gassó N et al. (2009) Exploring species attributes and site characteristics to assess plant invasions in Spain. Diversity y Distributions 15:50-58. Gerlach JD, Rice KJ (2003) Testing life history correlates of invasiveness using congeneric plant species. Ecological Applications 13:167-179. Ghalambor CK, McKay JK, Carroll SP, Reznick DN (2007) Adaptive versus non-adaptive phenotypic plasticity and the potential for contemporary adaptation in new environments. Functional Ecology 21:394-407. Groves RH, Di Castri F (1991) Biogeography of Mediterranean invasions Cambridge University Press, Cambridge. Hobbs RJ et al. (2006) Novel ecosystems: theoretical and management aspects of the new ecological world order. Global Ecology y Biogeography 15:1-7. Joshi J, Vrieling K (2005) The enemy release and EICA hypothesis revisited: incorporating the fundamental difference between specialist y generalist herbivores. Ecology Letters 8:704-714. 13.
(20) Capítulo 1. Kareiva P, Watts S, McDonald R, Boucher T (2007) Domesticated nature: Shaping landscapes and ecosystems for human welfare. Science 316:1866-1869. Lake JC, Leishman MR (2004) Invasion success of exotic in natural ecosystems: the role of disturbance, plant attributes and freedom from herbivores. Biological Conservation 117:215-226. Lambdon PW, Hulme PE (2006) How strongly do interactions with closely-related native species influence plant invasions? Darwin's naturalization hypothesis assessed on Mediterranean islands. Journal of Biogeography 33:1116-1125. Lavorel S, Canadell J, Rambal S, Terradas J (1998) Mediterranean terrestrial ecosystems: research priorities on global change effects. Global Ecology y Biogeography 7:157-166. Levine JM, D'Antonio CM (1999) Elton revisited: a review of evidence linking diversity and invasibility. Oikos 87:15-26. Levine JM, Vilà M, D'Antonio CM, Dukes JS, Grigulis K, Lavorel S (2003) Mechanisms underlying the impacts of exotic plant invasions. Proceedings of the Royal Society of London Series B-Biological Sciences 270:775-781. Levine JM, Adler PB, Yelenik SG (2004) A meta-analysis of biotic resistance to exotic plant invasions. Ecology Letters 7:975-989. Lloret F et al. (2005) Species attributes and invasion success by alien plants on Mediterranean islands. Journal of Ecology 93:512-520. Lockwood J, Hoopes M, Marchetti M (2007) Invasion Ecology. Blackwell publishing. MacDougall AS, Turkington R (2005) Are invasive species the driver or passangers of change in degraded ecosystems? Ecology 86:42-55. Mack RN (1996) Predicting the identity and fate of plant invaders: Emergent and emerging approaches. Biological Conservation 78:107-121. Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz FA (2000) Biotic invasions: Causes, epidemiology, global consequences, and control. Ecological Applications 10:689-710. Mack RN (2003) Phylogenetic constraint, absent life forms, y preadapted alien plants: A prescription for biological invasions. International Journal of Plant Sciences 164:S185-S196. Mack RN (2004) Global plant dispersal, naturalization, and invsion: pathways, modes and circumstances. In: Ruiz GM, Carlton JT (eds) Invasive species: vectors y management strategies Isly Press, Washington, DC, pp 3-30. Maguire LA (2004) What can decision analysis do for invasive species management? Risk Analysis 24:859868. Maron JL, Vilà M, Bommarco R, Elmendorf S, Beardsley P (2004) Rapid evolution of an invasive plant. Ecological Monographs 74:261-280. Morales CL, Traveset A (2009) A meta-analysis of impacts of alien vs. native plants on pollinator visitation and reproductive success of co-flowering native plants. Ecology Letters 12:716-728. Moragues E, Traveset A (2005) Effect of Carpobrotus spp. on the pollination success of native plant species of the Balearic Islys. Biological Conservation 122:611-619. Niinemets U, Valladares F, Ceulemans R (2003) Leaf-level phenotypic variability and plasticity of invasive Rhododendron ponticum and non-invasive Ilex aquifolium co-occurring at two contrasting European sites. Plant Cell and Environment 26:941-956. Olden JD (2005) Biotic homogenization: a new research agenda for conservation biogeography. In: 2nd Biennial Conference of the International-Biogeography-Society, Shepherdstown, WV, pp 2027-2039. Olden JD, Rooney TP (2006) On defining y quantifying biotic homogenization. Global Ecology & Biogeography 15:113-120. Peperkorn R, Werner C, Beyschlag W (2005) Phenotypic plasticity of an invasive acacia versus two native Mediterranean species. Functional Plant Biology 32:933-944. Peters DPC, Yao J, Gosz JR (2006) Woody plant invasion at a semi-arid/arid transition zone: importance of ecosystem type to colonization y patch expansion. Journal of Vegetation Science 17:389-396. Pigliucci M (2001) Phenotypic Plasticity: Beyond Nature y Nurture John Hopkins University Press, Baltimore. Pimentel D, Lach L, Zuniga R, Morrison D (2000) Environmental and economic costs of nonindigenous species in the United States. Bioscience 50:53-65. Pimentel D et al. (2001) Economic y environmental threats of alien plant, animal, and microbe invasions. Agriculture Ecosystems & Environment 84:1-20. 14.
(21) Introducción general. Pimetel D (2002) Biological invasions. CRC Press, Boca Raton. Pimentel D, Zuniga R, Morrison D (2003) Update on the environmental and economic costs associated with alien-invasive species in the United States. In: Conference on Bioinvasions, Laramie, WY, pp 273-288. Pimentel D, Pimentel M, Wilson A (2007) Plant, animal, and microbe invasive species in the United States and World. In: Nentwig N (ed) Biological Invasions vol 193. Springer-Verlag, Berlin Heidelberg, pp 315-330. Pyšek P, Sadlo J, Mandak B, Jarosik V (2003) Czech alien flora and the historical pattern of its formation: what came first to Central Europe? Oecologia 135:122-130. Pyšek P, Richardson DM, Rejmánek M, Webster GL, Williamson M, Kirschner J (2004) Alien plants in checklist y floras: towards better communication between taxonomist y ecologist. . Taxon 53:131-143. Pyšek P, Jarošík V (2005) Residence time determines the distribution of alien plants. In: Invasive Plants: Ecological & Agricultural Aspects, pp 77-96. Pyšek P, Richardson DM (2007) Traits Associated with Invasiveness in Alien Plants: Where do we stay? In: Caldwell MM (ed) Biological Invasions, vol 193. Springer, Berlin Heidelberg, pp 97-125. Recasens J, Calvet V, Cirujeda A, Conesa J (2005) Phenological and demographic behaviour of an exotic invasive weed in agroecosystems. Biological Invasions 7:17-27. Rejmánek M (1999) Invasive plant species and invisible ecosystems. In: Sylund, O.T. Schei, P.J. & Vilken, A. (eds) Invasive species and biodiversity management. Kluwer, Dordrecht pp 79-102. Rejmánek M, Richardson DM, Pyšek P (2005) Plant invasions and invasibility of plant communities. In: Van der Maarel E (ed) Vegetation ecology, Oxford, pp 332-355. Richards CL, Bossdorf O, Muth NZ, Gurevitch J, Pigliucci M (2006) Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions. Ecology Letters 9:981-993. Richardson DM, Pyšek P, Rejmanek M, Barbour MG, Panetta FD, West CJ (2000) Naturalization and invasion of alien plants: concepts y definitions. Diversity & Distributions 6:93-107. Richardson DM, Pyšek P (2006) Plant invasions: merging the concepts of species invasiveness and community invasibility. Progress in Physical Geography 30:409-431. Sakai AK et al. (2001) The population biology of invasive species. Annual Review of Ecology & Systematics 32:305. Sala OE et al. (2000) Biodiversity - Global biodiversity scenarios for the year 2100. Science 287:1770-1774. Sanz Elorza M, Dana Sanchez D, Sobrino Vesperinas E (2004) Atlas de las Plantas Alóctonas Invasoras en España. Ministerio de Medio Ambiente, Madrid. Sultan SE (2000) Phenotypic plasticity for plant development, function & life history. Trends in Plant Science 5:537-542. Sultan SE (2001) Phenotypic plasticity for fitness components in Polygonum species of contrasting ecological breadth. Ecology 82:328-343. Theoharides KA, Dukes JS (2007) Plant invasion across space and time: factors affecting nonindigenous species success during four stages of invasion. New Phytologist 176:256-273. Thuiller W, Richardson DM, Rouget M, Proches S, Wilson JRU (2006) Interactions between environment, species traits, and human uses describe patterns of plant invasions. Ecology 87:1755-1769. Tylianakis JM, Didham RK, Bascompte J, Wardle DA (2008) Global change and species interactions in terrestrial ecosystems. Ecology Letters 11:1351-1363. Traveset A, Richardson DM (2006) Biological invasions as disruptors of plant reproductive mutualisms. Trends in Ecology & Evolution 21:208-216. Valladares F, Gianoli E, Gomez JM (2007) Ecological limits to plant phenotypic plasticity. New Phytologist 176:749-763. Valladares F, Wright SJ, Lasso E, Kitajima K, Pearcy RW (2000) Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology 81:1925-1936. van Kleunen M, Fischer M (2005) Constraints on the evolution of adaptive phenotypic plasticity in plants. New Phytologist 166:49-60. Vermeij GJ (2005) Invasions as expectations: a historical fact of life. In: Sax DF, Stachowicz JJ, Gaines SD (eds) Species invasions: insights into ecology, evolution y biogeography. Sinauer Press, Suderly, MA. Vilà M et al. (2003) Association between Opuntia species invasion and changes in ly-cover in the Mediterranean region. Global Change Biology 9:1234-1239.. 15.
(22) Capítulo 1. Vilà M, Maron JL, Marco L (2005) Evidence for the enemy release hypothesis in Hypericum perforatum. Oecologia 142:474-479. Vilà M, Pujadas J (2001) Ly-use and socio-economic correlates of plant invasions in European y North African countries. Biological Conservation 100:397-401. Williams DG, Mack RN, Black RA (1995) Ecophysiology of the introduced Pennisetum setaceum on HawaiiThe role of phenotypic plasticity. Ecology 76:1569-1580. Williamson M (1996) Biological invasions. Chapman & Hall, London. Williamson M, Fitter A (1996) The Varying Success of Invaders. Ecology 77:1661-1666. Willis SG, Hulme PE (2002) Does temperature limit the invasion of Impatiens glyulifera and Heracleum mantegazzianum in the UK? Functional Ecology 16:530-539. Wilson JRU et al. (2007) Residence time and potential range: crucial considerations in modelling plant invasions. Diversity y Distributions 13:11-22. Wolfe LM (2002) Why alien invaders succeed: Support for the escape-from-enemy hypothesis. American Naturalist 160:705-711. Yeh PJ, Price TD (2004) Adaptive phenotypic plasticity y the successful colonization of a novel environment. American Naturalist 164:531-542. Zavaleta E (2000) Valuing ecosystem services lost to Tamarix invasion in the United States. In: Mooney HA, Hobbs RJ (eds) The impact of glo bal change on invasive species. Isly Press, Washington, DC, pp 261300.. 16.
(23) ..
(24) ..
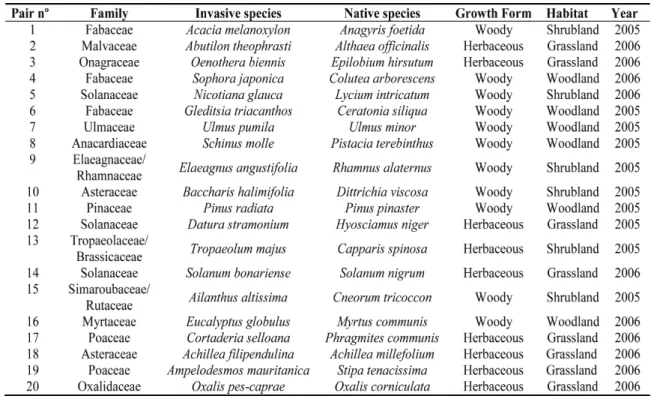
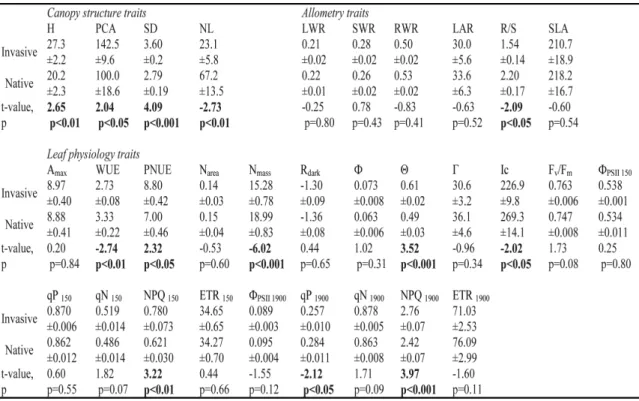
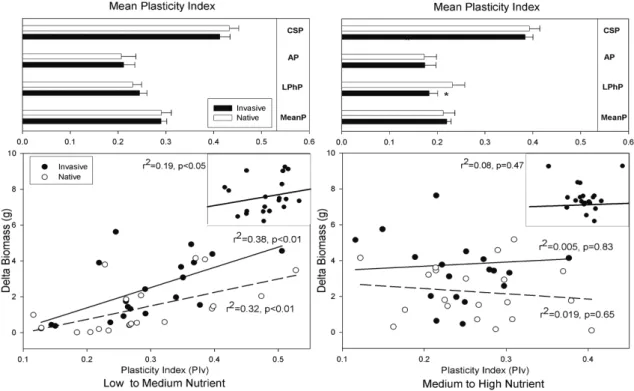
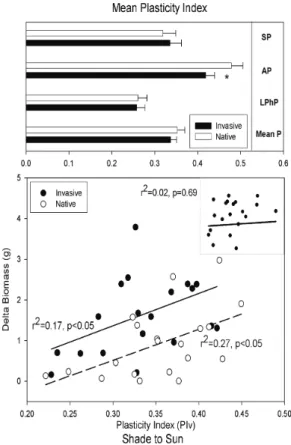
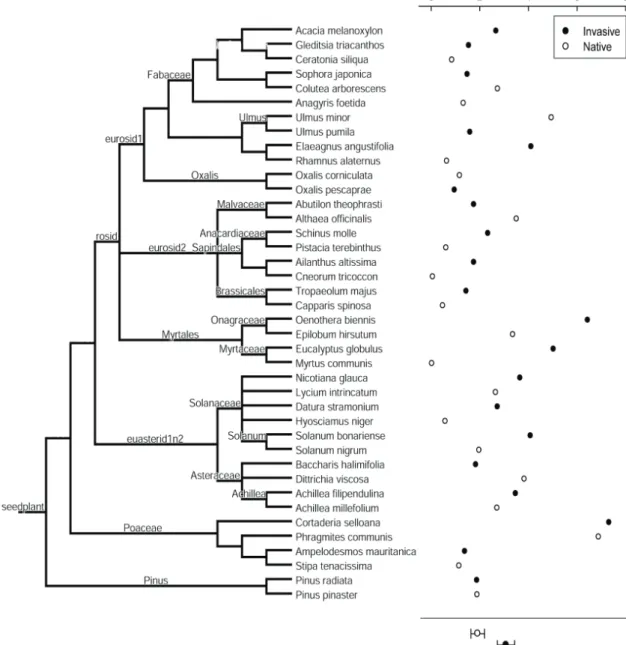
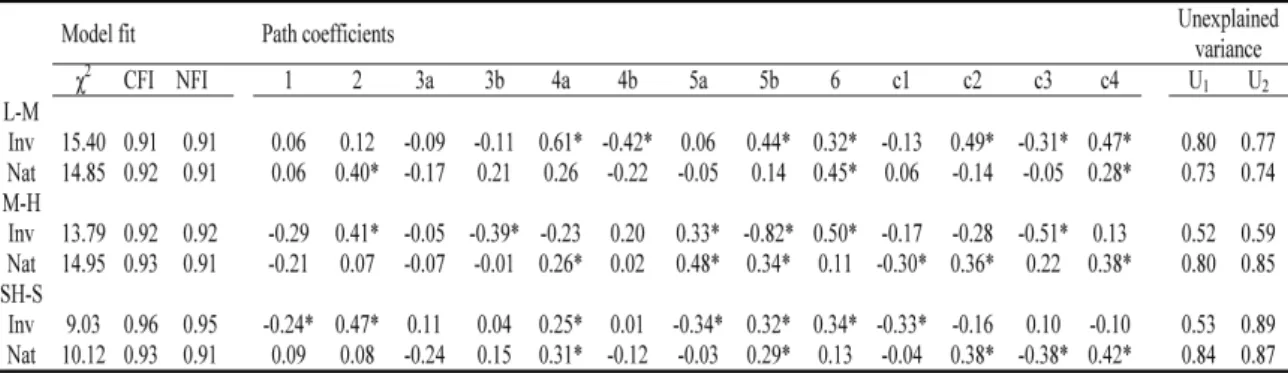
(25) Capítulo 2 Plasticidad fenotípica de las plantas invasoras comparada con las nativas Este capítulo reproduce íntegramente el texto del siguiente manuscrito: Oscar Godoy, Fernando Valladares & Pilar Castro-Díez (In preparation). Phenotypic plasticity of invasive plant species compared to natives species. Resumen Las respuestas plásticas de las especies a la variación espacio-temporal influyen fuertemente en su distribución. Se ha afirmado que las especies ampliamente distribuidas poseen una elevada plasticidad fenotípica. Los ecólogos han relacionado está elevada plasticidad fenotípica con la invasibilidad de las especies exóticas, ya que puede facilitar la colonización y rápida dispersión por el territorio de las especies exóticas (Ventajas de la Plasticidad para la Invasión, PAI hipótesis). Sin embargo, la mayoría de las afirmaciones se basan en conceptos teóricos y se tiene una total falta de conocimiento experimental. Para determinar el papel de la plasticidad fenotípica en la invasibilidad de las plantas exóticas, primero se caracterizó el fenotipo de 20 especies invasoras y nativas relacionadas filogenéticamente y ampliamente distribuidas en España, mediante la medida de 31 rasgos vegetativos, para posteriormente comparar su plasticidad, la eficiencia de la plasticidad, y la integración fenotípica a dos amplios gradientes experimentales. Además, se llevaron a cabo modelos de ecuaciones estructurales para determinar la importancia relativa de la plasticidad adaptativa bajo diferentes situaciones ambientales y los límites que la integración fenotípica impone a la plasticidad fenotípica. En media, las especies invasoras estuvieron caracterizadas por fenotipos más competitivos que los nativos. Diversos rasgos fisiológicos fueron los que influyeron en la mayor tolerancia de las especies invasoras desde los niveles limitantes, no limitantes hasta los saturantes en la disponibilidad de recursos. Las especies invasoras y nativas respondieron a las variaciones ambientales con una alta plasticidad en sus rasgos. Pero contrario a las predicciones de la hipótesis PAI, las respuestas plásticas fueron similares entre ambos grupos. Sin embargo, la invasibilidad de las especies exóticas parece estar más relacionado con la mayor eficiencia de sus respuestas plásticas. En este sentido, las especies invasoras incrementaron en mayor medida su eficacia biológica por cada incremento en plasticidad en comparación con las especies nativas. Las especies invasoras obtuvieron esa mayor eficiencia de la plasticidad por mecanismos opuestos a las nativas. Mientras que la plasticidad de los rasgos fue el factor principal en la mayor eficiencia de la plasticidad de las especies invasoras de ambientes limitantes a no limitantes, en las nativas fue el valor del rasgo. El patrón opuesto se encontró cuando los recursos variaron de no limitantes a saturantes. La integración fenotípica influyó positivamente en la plasticidad de los rasgos fisiológicos de la hoja pero negativamente en los alométricos. Por tanto, la integración fenotípica ayudó a las especies invasoras ser exitosas en ambientes limitantes, porque están basan la eficiencia de la plasticidad en sus respuestas fisiológicas. Palabras clave: Plasticidad fenotípica, especies exóticas invasoras, integración fenotípica, eficiencia de la plasticidad, plasticidad adaptativa, modelos de ecuaciones estructurales, contrastes filogenéticos independientes, gradientes de recursos, luz, nutrientes, rasgos ecofisiológicos.. 17.
(26) Plasticidad fenotípica de plantas invasoras. Phenotypic plasticity of invasive plant species compared to natives species Oscar Godoy*1,2, Fernando Valladares1,3,4 & Pilar Castro-Díez2 1. Instituto de los Recursos Naturales, Centro de Ciencias Medioambientales. CSIC. Serrano 115 dpdo E28006 Madrid Spain. *e-mail: [email protected] 2. 3. Departamento de Ecología, Universidad de Alcalá. Edificio de Ciencias, Campus universitario, Ctra. Madrid-Barcelona, km. 33.6, E-28871, Alcalá de Henares, Madrid, Spain.. Departamento de Biología y Geología. Área de Biodiversidad & Conservación, Universidad Rey Juan Carlos, ESCET, Tulipán s/n E-28933, Móstoles, Madrid, Spain. 4. Laboratorio Internacional de Cambio Global (LINCGlobal), UC-CSIC, Departamento de Ecología, Facultad de Ciencias Biológicas, PUC, Alameda 340, PC 6513677, Santiago, Chile. Abstract Plastic responses to overall space-temporal environmental variation highly influence the species distribution. Widespread species have been claimed for having high phenotypic plasticity. Ecologist are currently linking such a trait (high phenotypic plasticity) to plant invasiveness because enable exotics colonize and spread rapidly over large and environmental heterogeneous new areas (Plasticity Advantages for Invading, PAI hypothesis). However, the majority of the assumptions rely on theoretical concepts and a complete lack of experimental knowledge remains. To determine the role of phenotypic plasticity in plant invasiveness, we thoroughly compared well-known exotic invasive species with widespread native congeners. With this in mind, we first characterized the phenotype of 20 invasive-native ecological and phylogenetic related pairs from the mediterranean region of Spain by measuring an ample set of 31 different plant level traits, to consecutively compare their plasticity, the efficiency of their plastic responses and their phenotypic integration to broad experimental resources gradients. We also performed structural equation modeling to determine the relative importance of adaptive plasticity under different environmental situations and the limits that phenotypic integration imposes to phenotypic plasticity. In average, invasive species were characterized by phenotypes more competitive than natives. Several physiological traits account for the higher tolerance of invaders from limiting, non-limiting to saturating resource availability. Invasive and native species respond to environmental variations with high trait plasticity. But contrary to PAI hypothesis predictions, plastic responses were similar between both groups. However invasiveness of exotic species seems to be more related to the higher efficiency of their plastic responses. In this sense, invaders incremented higher their fitness compared to natives by each plasticity increment. Plasticity efficiency was achieved by opposite mechanisms between invasive and native species. From limiting to nonlimiting environments trait plasticity account higher in the plasticity efficiency of invaders whereas to natives the trait values were more relevant. The opposite pattern was found when resources availability was from non-limiting to saturating.. 20.
(27) Capítulo 2. Phenotypic integration rules the trait variation to positive effect in leaf physiological traits but negative in allometric ones. Since invasive species base their plasticity efficiency on physiological responses, phenotypic integration help invaders be successful in limiting environments. Keywords: Phenotypic plasticity, exotic invasive species, phenotypic integration, phenotypic efficiency, adaptive plasticity, structural equation modeling, phylogenetic independent contrast, resources gradient, light, nutrient, ecophysiological traits.. 21.
(28) Plasticidad fenotípica de plantas invasoras. 1. Introduction Linking phenotypic plasticity to invasiveness of exotic species is a burning topic in the study of biological invasion (Richards et al. 2006, Funk 2008, Hulme 2008). Phenotypic plasticity measured as the genotype property to render different phenotypes in different environments (Sultan 1995, Valladares et al. 2000, Pigliucci 2001) has been largely seen as a continuous key trait basic for explaining why exotic species are so successful in their recipient communities (Plasticity Advantages for Invading, PAI hypothesis) (Williams et al. 1995, Sakai et al. 2001, Sexton et al. 2002, Gerlach and Rice 2003, Niinemets et al. 2003, Peperkorn et al. 2005). Because high plasticity allows species express advantageous adapted phenotypes in a broad range of environments, it is likely to influence their invasiveness potential, their ecological distribution and their impact over the ecosystems (Alpert et al. 2000, Daehler 2003, Richards et al. 2005, Hulme 2008). For instance, it has been postulated that plasticity would reduce the stochastic risk of local extinction after the arrival of few genotypes and augment the likelihood of a species to become an invasive in habitat conditions where they were never before pre-adapted (Sultan 2001, Ghalambor et al. 2007). A high plasticity can be determined either by an adaptive strategy to cope with space-temporal resources fluctuation in the native region (Williams et al. 1995, Willis and Hulme 2002, Atkin et al. 2005) or by rapid evolution in novel sites after colonization (Agrawal 2001, Yeh and Price 2004, Donohue et al. 2005). In both cases, the highest the capacity to vary a broad group of morphological and ecophysiological traits the highest the probability of adaptive phenotypic plasticity to invasion, since the relative importance of any single trait is probably dependent on the invaded ecosystem (Lloret et al. 2005). Different. environmental. conditions. may. influence fitness homeostasis derived from specific-trait plasticity (Hulme 2008), especially under low resources environments (Davis et al. 2000). For instance, this “Jack-ofall-trades” strategy (sensu Richards et al. 2006) were found under low light conditions when invasive species varied largely the specific leaf area to maximize light capture (Funk 2008), when invasive tree species shifted their biomass allocation from transpiring tissue to water-accessing and water transporting tissue with an increase in temperature (Säumel and Kowarik, submitted manuscript) or when high root/shoot ratio plasticity enhanced water uptake under drought treatment (Dudley 1996). On the other hand, if invasive species track temporal pulses of high resources availability via phenotypic plasticity, “Master-of-some” strategy (sensu Richards et al. 2006), longterm effects on ecosystem impacts due to invasive persistence should be expected (Davis and Pelsor 2001, Sultan 2001). Although arguments in favor of linking phenotypic plasticity to plant invasion seem reasonable, no general pattern between phenotypic plasticity and invasive success have emerged so far and while some studies support PAI hypothesis (Gerlach and Rice 2003, Niinemets et al. 2003, Richards et al. 2006) others not (Bossdorf et al. 2005, Peperkorn et al. 2005). Nevertheless and despite it is well known that trait plasticity and mean trait value covary, only trait plasticity has been taken into account. For instance, with the same adaptive phenotypic plasticity one invasive species may display higher fitness compared to a native species if the trait value linked to fitness is significantly higher. According to this, even low plasticity may be advantageous for invasive species if the difference in the trait value linked to fitness is proportionally higher than the difference in this trait plasticity displayed between invasive and 22.
Figure



+7

Outline
Documento similar
