. .. ,'
..
.
JNOMBRE: CARLOS M A R T I N E Z MONTOYAMATRICULA: 7 8 2 1 4 9 1 4
CARRERA: B I O L O G I A
AREA D E CONCENTRACION: REPROOUCCION T R I M E S T R E : 1 2
HORAS SEMANA: 3 ü
LUGAR DONDE
SE
L L E V A R A A CABO: U . A . M . I .F E C H A D E I N I C I O : 0 2 - 1 2 - 1 9 8 2 F E C H A DE TERMINACION: 3 0 - 0 9 - 8 3
/NOMBRE D E L TUTOR INTERNO PUESTO Y A D S C R I P C I O N : BIOL. J O S E L U I S CONTRERAS M O N T I E L
PROFR. ASOCIADO 'ID"
D E P T O . E I O L O G I A D E L A REPRODUCCION
b
~ 4 1 . ~ ~ ~ 0 : DETERMINACION DE L A P A R T I C I P A C I O N DE LA I N E R V A C I O N
PENEANA EN L A R E A L I Z A C I O N D E L A COPULA EN L A RATA W I S T A R "
J
- .
ALUMNO:
TUTOR:
Arr h i d m a y Micimach. t e .Ylxiro 13. D.F..Td. 5814055
--
73s3
UNIVERSIDAD AUTONOMA METROPOLITANA.
IZTAPALAPA
C.
B.
S.*1Deterrninaci6n de l a P a r t i c i p a c i ó n de l a
I n e r v a c i ó n Peneana en l a Realización
1.5 P X n s v l I i 1- 1 - R = C a Macho Wistar"
CARLOS MARTINEZ MONTOYA
f8Determinaci6n de l a ~ a r t i c i p a c i ó n de l a Inervación Peneana en l a Realización de l a Cópula de l a Rata Macho Wlstar
'*
JUSTIFICACION Y NATURALEZA DEL PROYECTO
La reproducción en los mamíferos depende del adecuado
acoplamiento del macho y l a hembra; entre los eventos observables
para que se dé e l apareamiento y por l o tanto l a fertilizaclán están los movimientos pélvicos que r e a l i z a e l macho y que culmic
an
con l a penetración del pene en l a vagina (CÓpula).Sí los mov& mientos pélvicos copulatorios son inadecuados, l a cópula f a l l a . La actividad copulatoria depende de l a influencia hormonal sobreestructuras perifericas (textura, consistencia, coloración del p g
ne, pelo, p i e l sexual, producción de substancias que a l disolver- se en e l a i r e van a i n f l u i r en l a pareja atrayéndola y que se co-
noce con e l nombre genérico de feromonas, etc. l (1,14,21,30),como
d= c r t r i = t r r a s del Sistema ml:r--~~:z rz,trU! !Sr'C! que pnrtj.cfpv
en los eventos reproductivos, ya sea en e l aspecto motivacional
(Componente Apetitivo) de l a conducta de apareamiento, como en l a
ejecución de l a s pautas motoras que permiten l a consumación de l a
cópula (Componente Consumatorio)(1,2,4,5,14,21,22,25~. E l control
nervioso de los efectores musculares y viscerales que constituyen e l "Aparato Copulatorio" es determinante ya que alteraciones de l a s vias aferentes y eferentes son posibles c a w a s de f a l l a s re- productivas.
Lo
anterior permite abordar e l estudio de los mecanis-I
mos
endocrinos y nerviosos de l a reproduccl&n através del compor-tamiento sexual ya que alteraciones en e l l o s se reflejaran en l a
realización adecuada de éste.
I
.
Nuestro interes es conocer los mecanismos.nerviosos que regulan la actividad copulatoria de la rata, y así contar conun
modelo biológico más que contribuya a establecer una expli- cación general de los mecanismos dela
reproducción en las es-pecies de mamíferos.
ANTECEDENTES
La forma tradicional para estudiar el comportamiento
sexual, ha sido la observación directa
o
su analisis cinemato-gr¿-ffico.
En
l a de(cada de los 50s se diseño-una técnica e l e c t onica
que permite convertir los movimientos pelvicos realizadosdurante la copula en seiiales electricas.10 que permitió la cuan
-
tificación de los componentes motores del coito en conejos si-
do esta más precisa que las otras metodologías (27).
8
Dcy nuestra p r t - h - m n r r i m r s r r n i l a d ~ ~ r n a t+qic? rlr v-
gistro poligráfico que ha permitido caracterizar el patron
mo-
tor copulatorio, así como detectar alteraciones sutiles debidas
a
la manipulación experimental no detectadas por los metodosconvencionales, tanto en el conejo (8,10) corno.cn l a rata (7).
En base de los antecedentes mencionados nos proponemos determi-
nar
con dichas técnicas, el papel que juega la inervación delpene,
dada porel
nervio dorsalis penis (111, en la integraciónINTRODUCC I ON
La conducta sexual tiene dos componentes que son:
el
Apetitivo y el Consumatorio.
En el
caso
de la
rata
macho el componente Consumatorio
consiste en tres eventos, dentro de
los
cuales se realizan series
de movimientos pélvicos que tienden a mantenerse dentro de una
duración y frecuencia constante: La Montaa, en
lacual
no hay
Introducción del pene en la vagina, La Penetración, en la cual
se
presenta la introducción del pene en la vagina pero
no
hay
eyaculación y
por último, La Eyaculación, durante la cual
seda
ademas de la penetración la emisión del semen.
El estudio de los mecanismos hormonales y nerviosos que
participan en la regulación del comportamiento sexual, ha sido
realizado en su mayor parte estableciendo una correlación entre
la manipulación experimental sobre dichos sistemas y la obser-
vaci&n
de
las
posibles alteraciones del comportamiento sexual
después
de haberse realizado la manipulación
(2,4,17,21,22,24,25,29,30)
lo que implica el conocer las caracteristicas que
tipifican dicho comportamiento en el animal ontacto
(14,21).La
técnica
más
utilizada es la observación directa por
parte del investigador, registrando parámetros convencionales
(3,18,23,26,27),
las t6cnicas electrónicas han sido poco usadas
en
este tipo de estudios debido a que, la que se ha desarrollado,
a
pesar de su presición ha sido poco práctica. En el laboratorio
hemos desarrollado
un
sistema electrónico para registrar paráme-
tros temporales y motores de
laactividad copulatoria con una
mayor presición que los metodos tradicionales
(7,101lo
que peg
La
participación del Sistema Nervioso (tanto corticalcomo espinal) en e l control de varios de los eventos que consti-
tuyen e l patron motor copulatorio ha sido establecido por varios
investigadores (1,7,11,13,17,21,28). E l centro nervioso que con-
trola eventos como l a erección
,
tumescencia peneana y l a eyacu-lación se considera localizado en l a medula espinal. Se ha explg
rad0 l a participación de l a sensibilidad proveniente del pene
el
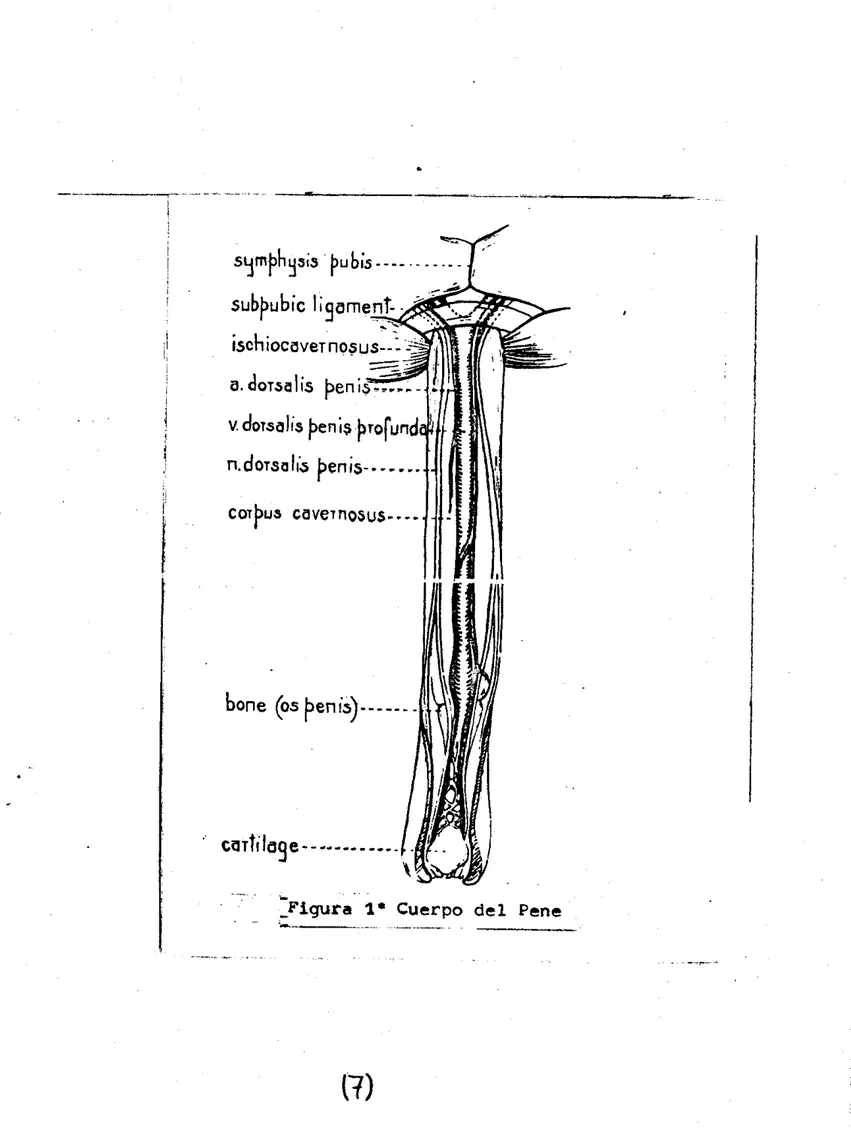
cual se encuentra inervado por ramas del nervio pélvico dor- s a l i s penis ( 1 1 ) . f i g:
1
reportandose cambios en e l patron tempo-r a l cuando son seccionados (7,9,11,13,15,20,28), sin embargo no se dan detalles de l a ejecución de l a cópula en
si,
por l o que nose sabe
si
hay alteración motriz y de haberla, cuál s e d a su con-secuencia. Por l o que es necesario demostrar formalmente s i es
importante l a inervación sensitiva peneana para l a realización de l o s movimientos pelvicos.
’
i
. ..
I
I
I
1 I
I
1
1
II
I
I
!
I
. .. -
i
-.
Figura 1. Cuerpo del Pene
MATERIAL Y METODO
La prueba de actividad sexual consistió en colocar a l macho en un redondel o c i l i n d r o de Plexiglas con un diáimetro de 53 cm
y 42
cm
de altura, con una cama de aserrín. En algunos casos pa-r a comprobar s i había erección, e l piso fu¿ de c r i s t a l , e l cual
a su vez se encQntraba L montado sobre un artefacto, que permitla
l a observación ventral.de los animales (figura 1 ) . En cuanto a
l a observación l o primero que se hacia era colocar e l macho solo,
durante 5 minutos, como periodo de acostumbramiento, a l t i h i n o
del cual se introduce a l redondel una hembra escojida por su re-
ceptividad para que e l macho l a montara iniciandose l a cronome-
tración en ese momento. La actividad sexual desarrollada por e l
macho fue! valorada por medio de dos. técnicas:
1. La observación directa, anotando e l tiempo transcurrido
y los eventos que se presentacm montas, penetraciones, eyacula-
ciones, etc.
1
2. Registrando los movimientos pelvicos copulatorios por
medio de un Acelerometro Grass SP-1 conectado a un Poligráfo
Grass en donde cada movimiento pélvico fue registrado en papel.
I I
Estos dos sistemas de valoracion de l a actividad copulatú-
r i a nos permitieron determinar e l número de eventos
a s í
como l a s caracterlsticas de l a ejecución de cada uno de e l l o s (patrones tipicos de: monta, penetración, eyaculación y ademas su amplitud, frecuencia y forma)I
i
I
i
Un grupo de doce ratas machos, con una experiencia minima
de dos eyaculaciones, f u e sometido a pruebas de actividad sexual
para determinar l o s posibles cambios en su patron de movimientos
E
p&lvicos copulatorios en diferentes condiciones de insensibilidad peneana y prepucial, para l o cual, de quince ratas macho Wistar,
sexualmente maduras, se escogieron aquellas que presentaron act&
vidad copulatoria, estando los pesos corporales de &stas entre 323 y 591 gramos ív& tabla 1).
Para seleccionar a los animales activos se hicieron
sesio-
nes de 1 5 minutos de duración y consistieron en introducir a l ma-
cho en un redondel de observación ( f i g u r a l), primero é l solo,
durante 5 minutos, como periódo de acostumbramiento, sin que
-
este tiempo se cuantificara como parte de l a duración de l a se
-
sión;
transcurridos los 5 minutos de acostumbramiento se intro-
d u d a a l redondel una hembra señuelo adecuadamente receptiva,
para incitar a l macho a montar (figura 2); en e l momento en que
se
introducía l a hembra se iniciaba l a cuenta del tiempo de dura-
ciÓn de l a sesión. S i durante e l transcurso de 15 minutos e l ma- cho no montaba, se l e retiraba y se l e repetía l a sesión trans-
curriendo un mínimo de 48 horas. Los machos que
no
fueron acti-vos en 4-6 sesiones se eliminaron.
A l o s animales activos se l e s dio dos sesiones hasta que
alcanzaban l a eyaculación, considerándolos a p a r t i r de entonces
como animales sexualmente experimentados, además, durante l a s
sesiones para adquirir experiencia copulatoria
se
l e s colocó e lames que asequra el dispositivo de registro, pero sin este Ú l -
timo, ( f i g u r a 2 ) con e l f i n de que se acostumbraran a tenerlo
puesto durante l a cópula, a s í mismo l a s sesiones se realizaron
en e l redondel de observación para que los animales se habituaran
.
Los machos seleccionados fueron hospedados desde e l primer d i a en jaulas individuales proporcionandoseles alimento y agua ad 1 ibitum.
Las sesiones experimentales se realizaron segtn l a s siguien-
tes secuencia y caracteristicas para cada macho, aunque no todos
l o s animales llegaron a completa: : ü s (ver tabla 1).
Todas l a s sesiones que a continuación se describen se dieron
5 minutos de acostumbramiento previo a l a prueba de actividad uopu-
latoria.
Control
Fueron dos sesiones con duración de 1 5 minutos s i e l macho
no
presentaba actividad copulatoria o,s t
solamente realizaba mon-tas; s$ el animal presentaba penetraciones se le daban 30 minutos
a p a r t i r de l a primera de e l l a s ; l a s sesiones se suspendfan
el
momento d e presentarse i o cyacuiabi¿n; soiv se wltslaer-Con a-quellas sesiones en que hubo eyaculaciones; cuando e l animal no l l e g o a eyacular se
le
r e p i t i ó l a sesión control correspondiente;en estas sesiones l o s animales no fueron sometidos a ninguna mani-
pulación experimental.
en
Anestesia del Pene
A l o s mismos animales se l e s sometió a 80s sesiones con du-
ración de 15 minutos alcanzaran o no l a eyaculación. La manipula-
ción
experimental consistió en anestesiar e l pene, para i o cual sesentaba a l macho sujetandolo suavemente, una vez tranquilizado se
Ie r e t r a í a el prepucio, hasta exponer completamente el pene e l cual
era masajeado durante 1 0 minutos con pomada de Xilocaina ( 5%)
cuidando que toda l a superficie del pene fuera sometida a l tra- tamiento (3igura 3 ) . A l regresar e l macho a l redoncikl de observa-
ción se introducia inmediatamente l a hembra señuelo, inicigndose
En algunos casos se suspendfa antes ya que, aparentemente, dismi-
nuia e l efecto del anestcsico y e l macho eyaculaba. En todos los
casos se tomó como tiempo de duración del efecto del anestesico
e l trascurrido hasta l a aparición de l a primera penetración.
i
c
-
-
--
--_
.-
Dencrvación del Pene
Concluida l a manipulación con Xilocaina pomada se di& un
intervalo de 1 0 días continua/ndose con dos sesiones ( de 20 minu-
tos cada una), en l a s cuales los animales tenían 4 d í a s mínimo
de haber
sido
denervados. La dcnervación se r e a l i z ó anestesiandoa los animales, hasta un nivel quir&gico, con pentobarbital sÓ-
dico (anectesal) administrado por v i a intraperitoneal; colocando
a l animal en posición supina, asequrando sus extremidades, se ti-
se hacfa un corte longitudinal en l a parte dorsal y media del pre-
pucio, y por disección roma se profundizaba hasta alcanzar e l cuer
PO del pene y localizar su extremo más profundo; se disecaban los
dos nervios dorsales del pene evitando que no quedaran f i b r a s , se
seccionaban por medio de dos cortes hechos dejando una distancia
de aproximadamente un centimetro entre uno y otro de los extremos
del corte (Figura 4 ) . Se procedia a suturar l a herida y durante los
dias siguientes se vigilaba a l animal para prevenir los problemas
-
- operación.
,. ,
.
i
<
1
.1
, .
. . ,
.-fqura 4 oenervación del Pene > - ' . . , A : '
1..4*<fG'*.'-;A
--.-
~ . ~ .
-
Denervación más Anestesia del Pene
La siguiente etapa consistio en : A los animales denerva-
dos se l e s dejo descansar de 4 a 5 dfas despues de su ultimo re- g i s t r o dentro de l a cámara de observación. A continuación a estos
mismos animales se les administro topicamente Xilocaina en e l pene
durante 10 minutos teniendo cuidado en frotarselo suave y constan-
arnes y se introduce a l a camara de observacibn, inmediatamente
despues se introsuce l a hembra señuelo y se empieza a cronometrar
e l tiempo que era d e 1 5 minutos. Se corrieron tres pruebas de
actividad . sexual ~.. ..~ ~ . por ..~ cada ~ animal.- . ..~ ~
i
- . - .
-- . .. .
-I
-~ ~.
Denervaci6n d e l Pene más Anestesia del Prepucio
A los mismos animales, transcurridos 5 días de finalizada
l a manipulación anterior se les sometio a anestesia del prepucio.
Para e l l o se sujetaba a l animal de t a l manera que se pudiese in-
yectar Xilocaina a l 1% en varios puntos del prepucio ver figuras
6 y 7. Inmediatamente inyectada l a Xilocaina se le introducia con
e l arnes montado a l a camara d e observación, se introducia l a hem
bra señuelo y se iniciaba l a sesibn. Se hicieron tres sesiones
igura 7 Denervación
RESULTADOS
A.
En
cuanto a los obletivos particulares
Una consecuencia inmediata derivada de la revisión bi-
bliográfica, fué comprender la ubicación del problema concepg
al que conforma la hipótesis del proyecto, dentro del desarrollo
histórico de
lainvestigación
dela conducta sexual, como fenó-
meno biológico
ycomo parte del esclarecimiento del significado
de los eventos que conforman
laactividad reproductiva de los
mamiferos
ysus
causas neuroendócrinas
ysignificado fisiológi-
co.
Además se ha
idofortaleciendo
el hábito de leer, comprender
lo implicit0 en las publicaclones cientificas, así como confor-
mar
la rutina periódoca para
ir
conociendo los avances en
las
areas de interes. Además se incremento
lamotivación para la
busqueda constante de inquietudes
yexplicaciones de las mis-
mas,
que es
lo queconforma
laactividad intelectual del cieR
t<fi.cn. Asf.
mismo, como
r n n q r c * i - n r + a , se a f i r m i -1 n,epirjtl.icritico
al confrontar los diferentes puntos de vista existen-
tes entre las explicaciones que diferentes investigadores prg
ponen a los mismos problemas,
ymás
aún,
los puntos de
vista
que nacieron por propia experiencia.
-
Por otro lado,
el estar en contacto
con
la actividad del
laboratorio, muy v'suis generis" en el area del comportamiento
animal, afino la disciplina
y elsentido de observación que
se
necesita para poder discernir lo importante de lo accesorio
ymantener el ambiente propicio para
la manifestación controlada
de
las conductas observadas, lo que ademds, di6 sentido al re-
sultado de la cuantificación (datos)
d elos parametros regis-
nejo estadístico de los datos.
r
B.
EHirelaciÓn al trabajo experimental:
En la tabla
1 sepuede observar que el 50%
delos animales
cubrieron satisfactoriamente
lasdiferentes etapas del diseño
ex-perimental propuesto, el
75%cubrió hasta la penúltima etapa y que
el
100%cubrió las dos primeras etapas.
Como puede verse en la fiqura
8durante la,etapa control,
los animales realizaron
SUSmovimientos péivicos
crl formasemrjan-
te
a
como
hasido reportado para el animal intacto
( 7 )coincidiendo
en cuanto
a1ñ frecuencia y duración de
l o strenes de movimientos
p&lvicos como de la monta como de
lapenetración
y laeyaculación.
Al aplicarseles Xilocaina, los animales presentaron solo
montas mientras duro
elefecto del anestésico, mismo que nosotros
consideramos que
sed6
hasta que
sepresenta la primera penetración,
después de lo cual,
ycomo puede observarse en
l afiqura 8
del lado
derecho de
labarra gruesa vertical,
los
animales
(68.33%)recuperan
la capacidad de realizar penetraciones y aún la eyaculación
(41.667;)La
duración del periódo de insensibilidad peneana producido por el
anestésico local
es deaproximadamente
15''y con una media de
9'33"ademas con una
Moday Fediana de
1 5 ' .La denervación del pene tiene como consecuencia un incremen-
to en el número de m m t a s
(aumenta del
42.46%al
96.02%,'las mon-
tas aumentan el
53.56%),sin embarqo las c-racteristicas
de dura-
ción y frecuencia no se alteran significativamente, como se observa
en la fiqura
8del
ladoderecho de
labarra gruesa vertical
yen la
tabla
2;algunos animales llegan a presentar penetración
( 7 7 % ) ein-
clusive un
(11%)presentó eyaculación; manteniéndose
10seventos
7
dentro
delas características observadas en
SUScontroles*
Se puede observar en la figura
8,del lado izquierdo de la
barra qruesa vertical, que cuando a los animales se
lesprobó de5
pugs de denervados y además aplicandoseles anestésico local en
e1pene, realizaron montas cuyos registros poiiqráficos no presenta-
ron
alteración en la ejecución al compararseles con
elcontrol.
. ,
i
i:
i -i i !... . .
.
, . .. . ~Y como se observa en l a tabla 2, e l 55% de loa animales a l menos
una vez present6 penetración y e l 33% llego a presentar eyaculación.
Como puede observarse en l a f i g u r a 8 del lado izquierdo de
l a barra vertical; cuando a l animal denervado
se
l e administro e lanestesico local inyectándosele en e l prepucio ( f i g u r a 7 )
se
obtie-ne un registro que mostraba una desorganixaci6n de l a monta en cuan
to a l a ejecución de los movimientos pelvicos, alterandose l a for- ma, frecuencia y duración del evento. Esto puede observarse en l a
figura 8 del lado izquierdo de l a barra vertical que representa e l
registro poligráfico de una monta donde solamente
se
insinfian losmovimientos pelvicos. Lo anterior se observo en e l 70% de los ani-
males, que alcanzaron esta etapa ya que e l 30% no llegaron a r e a l i - zar propiamente montas puesto que e l registro y l a observación d i -
...
I T A R L A I
""---
1
i
'
,
i
1
'!
II
I
I I I
!
I
!
j
I
t
I
1
!
x
n
x"
x
a
. . . ..
o
,
L _ ' _
DISCUSION Y CONCLUSION
Los movimientos pélvicos que r e a l i z a l a r a t a macho durante
l a cópula tienen características constantes en cuanto a l a dura-
ción de cada uno; los movimientos p&lvicos son realizados uno trás
otro en una secuencia característica y durante
un
tiempo determina-do, constituyendo trenes de movimientos pelvicos cuya duración esta
siempre dentro de ciertos lfmites a l i g u a l que e l ' número de movimi-
entos pel.vicos (frecuencia) ( 7 ) . Constituyendo patrones innatos de
l a rata. Ahora bien, estos movimientos deben tener l a s caracteris- ticas de los movimientos r e f l e j o s , cuyo centro de inteqración cs- t a r i a en Médula Espinal, a nivel lumbar y sacro, que es donde se incorporan a l C.N.C. los nervios que inervan l a región ( 1 , > 7 , 11,
1 3 , 28, 29,301. Existen evidencias de que r e f l e j o s relacionados
con l a cópula tales como: l a erección, los movimientos peneanos
que se observan durante l a erección y l a eyaculación, se inteqran
en los segmentos lumbares tanto en l a rata como en otros mamíferos
incluyenao a l homore \Ir, 13, 1 5 , L Y I . D ~ e l caso de ~a Taca, en e l
pene existen un c o l l a r de estructuras queratinizadas que asemejan espinacn) cuya base se observa una p a p i l a l a s cuales están asoci- adas con terminaciones nerviosas; estas estructuras, llamadas es-
pinas peneanas, son hormona-dependientes ( 1 3 , 29, 30) y se l e s con
sidera una función de receptores t a c t i l e s , que llevarfan a medula
espinal l a información del momento en que e l pene penetra en l a
I
vagina. La presencia de l a s espinas peneanas apoyarla l a hipótesis
de que en e l pene existen receptores que informan de los contactos del pene con l a regi6n perineal de l a hembra y que entre estos re-
ceptores se encontrarian los que generan señales para disparar
e l r e f l e j o espinal de los movimientos pélvicos. Por l o anterior
l o lógico serfa que a l bloquear l a sensibilidad del pene se alte-
r a r a l a ejecución de los movimientos p6lvicos de t a l manera que
pudieramos detectarlas con nuestro sistema de registro; sin embaf
go nuestros resultados nos indican que l a sensibilidad del pene
I
parcial ya que, en ninguno de los casos en los que se insensibilizo
e l pene, ya sea por l a administración tópica de anestásico o por
lasección de los nervios peneanos,
se
logró- que en forma consistefite se impidiera Ó se alterara l a ejecución de los movimientos pC1-
vicos y en todo caso, solo
se
postergó Ó impidio l a aparición depatrones de penetración, que están ligados aparentemente con l a
eyaculación, o l a eyaculación misma. S i n embargo, a l insensibili-
zer además del pene, e l prepucio, que cabe aclarab que a diferen-
c i a del humano, no penetra en l a vagina, dejaron de realizarse mo-
vimientos phlvicos cuando los machos montaban a l a s hembras o en
todo caso los movimientos realizados a l montar a l a hembra eran
movimientos que en poco
se
asemejaban a los del animal intacto,es decir, que l a imsensibilización del prepucio desorganizó con-
siderablemente e l patrón de ejecución de movimientos p&lvicos co-
pu l a tor io s o
Los resultados obtenidos nos llevan a concluir que:
a ) La captación de estímulos t a c t i l e s (roce, presión, calor,
etc,) posiblemente sean captados a través de l a s espinas peneanas
o
algunos otros receptores superf i c i a l e s que tendrian relación cone l r e f l e j o eyaculatorio y que en todo caso,
su
relación con los mo-vimientos p&lvicos tendría
un
efecto inhibitorio del patrón de mon-t a impidiendo los movimientos intravaginales en l a s penetraciones.
b) Por e l hecho de que animales denervados o denervados más
xilocaina lleguen a presentar e l patron de penetraci6n y e l de eya-
cuiacibn, se puede pensar en alguna de l a s siguientes posibilidades
o combinación de e l l a s :
i. Que l a sección de los nervios dorsales del pene, a pesar
del cuidado puesto, no haya sido completa.
profundos, rehcionad
de PI.
si&,
que p a r M
reflejo ayaculatoria
bilidad
delpene, CUB
que
ver
conla genera
pélvicos.
c)
Nuestros resultados
A
la rata, es
ellugar
por
dendeferente del arco
r e f l e j opara 1
mientos pelvicos.
i
con la percepción
decambios
Lpen en
el
desencadenamiento del
15) y
que
por
lo tanto, la sensl
iuiera que sea
sutipo,
no
tiene
LÓn
yejecución de
losmovimieni
jican que
e).area prepucial, en
?obablemente
seinicie la via a-
BIRLIOGRAFIA
1
.
Beach, F.A.
and Ford
C.S.;1951. Patterns
of sexual behavior
Harper Torchboo
s ;New Yor
,
London:
pp.239-242.
2.
Beach,
F.A.;1968. Factors involved
inthe control of mounting beha-
vior by female mammals. In: Reproduction and sexual behavior; edited
by
M.Diamond. Bloomington: Indiana üniversyty Press: 63-131.
3.
Bermant,
G.;1965.
Rat sexual behavior: Photographic analysis of the
intromission response. Psychon. Sci.
2 : pp.65-66.
'4.
Beyer,
C.and McDonald P.G.,1973.
Uormonal control
of sexual beha-
vior
in the female rabbit. Adv. Reprod. Physiol. 6: pp.185-219.
5. Beyer,C. Molalf,G.,Larsson,k
.
and Coderseten, P.,
1976. Steroid
regulation
of sexual behavior. J. Steroid Biochem.
&: pp.1171-1176.
6. üeyer, C., Velázquez,
J.,Larsson
.
andContreras,J.L.,
1980.Androgen
regulation of motor copulatory patterna in the male New Zealand white
rabbit. Hormones
andBehavior 14,pp. 179-190.
7,
Beyer,C. Contrerac,J.L.,
Larsson,\<
.
,
Olmedo,M.,
and Piorall,
G,1982
Patterns of motor and Seminal Vesicle activities during copulation
in the male rat.Physiology
andBehavior,29,pp. 495-500.
8.
BroKs, CH. Mc., 1973. The role
ofthe cerebral cortex and of various
sense organs in the excitation and execution of mating activity in
the rabbit.
Am.J.
Physiol., 120:pp. 544-563.
9
.
Calareso, F.R.,1970.
Experimental studies on the dorsal nerve of the
penis
of the rat.
Am J.Anat.:
10.
Contreras, J.L.,
and Beyer, C.,
1979.
A polygraphic analysis of
* . _
of Behavior, vo1.23:
pp. 939-943.
11. Chace Green E.,1963.
Anatomy of the rat. Hafner Publishing Co.,
mounting
andejaculation in the New Zealand white rabbit. Physio-
New Yor
and London; 370.
12. Dahlof,
L-Dand Larssin,
.,
1975. Interackional effects
of pudenal
nerve section
andsocial restriction
on male
rat
sexual behavior.,
.,/
13.
14.
15. 16.17.
18.
19.
20.21.
22. 23.24.
Dahlof,
L-D.,1979. Sensory stimulation
and masculine sexual beha-
vior
of the male rat. Doctoral Thesis; University of Gotbeborg.Sweden.
Dewsbury,
D.A.,1979. Description of sexual behavior in research on
hormone-behavior-interactions. ‘In: Endocrine Control of Sexual Be-
havior, edited by Beyer, C. New Yor
;
Raven Press.
Hart,B.L.,1972.
Sexual reflexes in the male rat after anesthetization
of
the glans penis.
Behav. Biol. 7:pp.
127-130.
Hart,
B.L.1978. Hormones,spinal reflex,
andsexuaX behaviour.
I
!
1
Kurt=,
R.G.and Adler, N.T.,1973.
Electrophysiological correlates
ofcopulatory behavior in the male rat: Evidence
of
an
equal inhibitory
procrss.
J.
Comp. Physiol. Physiol.
84:pp. 225-239.
Lanee.,J.W.,
Schwab R.S.
and Peterson, E.A.,
1963. Action tremor and
the cowhell phenomenon in Par inson5 disease. Brain 86:
pp. 95-110.Larsson,!!
.
1956. Conditioning
and sexual behavior in the male albino
rat.
Acta Psychologica Gothoburgensia,
1:
269.
Larsson,\:
,
and Sodersten
P.,
1973.
Mating after section of dorsal
penilenerve. Physiology
and Behavior, vol. 10: pp. 567-571.
Lis
,
R.D.Hormonal
regulation of sexual bahavior
in
polysterous ma-
mmals common
to the
laboratory. In: Handboo
of
Physiology.
Endocri-
nology 11, Part
1;
pp. 223-260.
1Malmnas, C.O.,
1973. Monoaminergic influence
of
thestosterone-acti-
vated copulatory bahavior
in
the castrated male rat. Acta Physiologi-
ca
Scandinavica Soppl. 395:
pp.1-128.
Mitsuya, H.;Asai,
J.;
Siuama,y
.,
Ushida,
T, and Hospe,Y
.,
1960.
Aplication of X-ray cinematography in urology: 1. Mechanism of eja-
culation.
J.Uro. 83:
pp.86-92.
Morall,
G.and Beyer,C., 1979. Neuroendocrine control of mammalian
I
estrous behavior. In: Beyer, C,
(Ed):Endocrine Control of Sexual
25. 26 27. 28. 29. 1
Morelí,G.,Larsson
K
andBeyer,
C.1977.
Inhibition of testosteroneinduced sexual bahavior
in
the castrates male rat by aromatase blo-c
ers.
Hormones and Behavior,9:
pp. 203-213.Peiree,
J.T.
and Nutall. 1961. Durationsof
sexual contactin
therat. J,
Comp. Physiol. Psysiol. Psychol. 54: pp. 584-587.Rubin,H.R, and
Azrin,N.H.
1967. Temporal patterns of sexual behaviorin
rabbits as determined byan
automatic recording technique.J.
Exp.I
i
Analysis Behav. 10: pp. 219-231. l
I
Sachs,
B.D.
and Garinello, L.D. 1979. Spinal pacema er controllingPenile
Re
taction and Protusionin
Rabbits. Acta Physiol. Scandina-vita, 106: pp. 199-214.
Sjostranc',
N.O.
and linge E. 1979. Principal mechanisms controllingpenile retraction and protusion
in
rabbits. : -30
Sodersten,P.
1974. Studies on the hormonal regulationof
masculines e x u a l bahavior patterns
in
female and malerats.
Doctoral Thesis.Un.l,verci$ty
of
Gateborq Sweden.31.
Stone, C.P. and Ferguson, F.W. 1940. Temporal relatioshipsin
thecopulatory acts
of
adult male r a t s . J. Comp. PhysioL. Psychol. 30:pp. 416433.
32.
C.Bcyer,L.J. Contreras, G.Mora11 and\!. Larsson. 1981.
Effects ofttern of the Rat. Physiology
and
Behavior.Vol.
27
pp. 727-7304
Castration and Sex Steroid Treatment on the Motor Copulatory Pa-
l
i
i
;
33. J.L. Contreras, P, Santos, C w l m
M,
Montoye, J.E.Yteyra
y C , Beyer 1983"Participación de l a Sensibilidad Peneana y Prepucial en l a
Realization
de los.
4Movimientos
Pélvicos CQpulatori.os de la Rata MachoWistar,eq
XXVI Congreso Nacio-
nal de la Sociedad Mexicana de Cienciw FisiQ16gicas. Colima, Col.