OLIMPÍADA ARGENTINA DE BIOLOGIA
AUSPICIA Y FINANCIA EL MINISTERIO DE EDUCACIÓN, CIENCIA Y
TECNOLOGÍA
UNIVERSIDAD NACIONAL DE RÍO CUARTO
FACULTAD DE CIENCIAS EXACTAS FÍSICO-QUÍMICAS Y NATURALES-
CUADERNILLO TEÓRICO
1
CONCEPTOS DE LAS CIENCIAS BIOLÓGICAS
Sobre la base de los temarios teóricos y prácticos propuestos para la OAB, y considerando la necesidad de lograr un abordaje del conocimiento actualizado es que ponemos a su disposición esta guía orientadora, según la bibliografía disponible en esta Olimpíada, la cual es consultada por los Comités académicos para lograr ideas que permitan la elaboración de los exámenes de todas las instancias de esta Olimpíada.
La ampliación de estos contenidos pueden realizarse en cualquier material que Ud. disponga y que se encuentre acorde a lo aquí especificado.
En esta primera propuesta se presentan los tres tópicos de los temarios teóricos (Biología Celular, Organismos y Etología, Ecología y Evolución), de los cuales se seleccionaron algunos puntos en función de las dificultades observadas en ediciones anteriores de la OAB, entre ellos Biotecnología, Sistemática de animales, Anatomía de vegetales, Cladismo y conceptos incluidos en Etología. Asimismo se incorporaron algunas destrezas básicas incluidas en los temarios prácticos de ambos niveles.
Compilación: Lic. y Prof. Analía Barbosa
2
Tópico: Biología Celular
Ingeniería genética
En 1970 comienza el desarrollo de la tecnología del DNA recombinante, llevando a métodos de investigación novedosos.
Las técnicas de DNA recombinante se desarrollaron en un principio como herramientas que permitirían a los científicos obtener una gran cantidad de copias de cualquier segmento de DNA específico, de manera que éste pudiera estudiarse desde el punto de vista bioquímico. Esto se realizó en un inicio introduciendo DNA ajeno en células de microorganismos. En condiciones apropiadas, éste se duplica y transmite a las células hijas cuando la original se divide. Así una secuencia de DNA específica puede ser amplificada o clonada para producir millones de copias idénticas que pueden aislarse en forma pura. En la actualidad son cada vez más importantes los métodos in
vitro.
La tecnología del DNA recombinante tiene varias aplicaciones, una de las que más avanza es la ingeniería genética, que implica la modificación del DNA de un organismo para producir nuevos genes con nuevas características. Su desarrollo se debe al descubrimiento de las enzimas de restricción y de la transformación bacteriana de los plásmidos. Las bacterias producen enzimas conocidas como de restricción, las cuales cortan las moléculas de DNA sólo en lugares específicos.
La amplificación de un fragmento de DNA puede lograrse in vitro, con el uso de
polimerasas de DNA de bacterias termorresistentes e in vivo, introduciendo moléculas
de DNA recombinante (vector + inserto DNA) en bacterias u otros microorganismos como las levaduras.
Las enzimas de restricción son “tijeras moleculares” cuya especificidad permite realizar cortes del DNA de manera controlada. Existen varias enzimas conocidas, un ejemplo es Bam HI que reconoce y corta una molécula de DNA en la secuencia de bases 5´G/GATTCC-3´, otro es EcoRI, que corta la secuencia 5´-G/AATTC-3´.
3
que se denomina “pegajosos” por la posibilidad que poseen de unirse entre sí por enlaces de hidrógeno. Además, el fragmento puede tratarse con una ligasa de DNA para lograr una molécula recombinante estable.
En la figura se muestra el mecanismo de acción de una enzima de restricción.
Luego del aislamiento, los fragmento del DNA, deben ser incorporados a un portador adecuado llamado vector para poder ser amplificado.
Vectores
Después de unirse a un vector o vehículo de clonación, un segmento de DNA puede llegar a entrar en una célula huésped y replicarse (clonarse). Los vectores son, esencialmente, moléculas de DNA transportadoras. Para servir de vector, una molécula de DNA debe tener unas determinadas características:
1. Debe poder replicarse independientemente junto con el segmento de DNA que transporta.
4
vector.
3. Debe tener algún marcador de selección, (normalmente genes de resistencia a antibióticos o genes de enzimas que la célula huésped no tenga) para poder distinguir las células huésped que transportan al vector, de las que no las contienen. 4. El vector debería poder extraerse fácilmente de la célula huésped.
Actualmente se utilizan tres tipos principales de vectores: los plásmidos, los
bacteriófagos y los cósmidos.
Los plásmidos
Un plásmido es una molécula circular separada del DNA bacteriano y mucho
más pequeña que éste pero con la capacidad de duplicarse dentro de la célula bacteriana y de brindarles facultades para desarrollarse en condiciones específicas en las que no podrían hacerlo si no estuvieran transformadas.
Son moléculas de DNA de doble cadena extracromosómicas de origen natural que tienen un origen de replicación (ori+) y que se replican autónomamente en células bacterianas. Para poder utilizarlos en ingeniería genética, se han modificado de manera que contengan un número limitado de sitios de restricción y genes de resistencia a antibióticos específicos.
El vector pBR322 fue uno de los primeros plásmidos diseñados que se utilizó.
Figura. Mapa de restricción del plásmido pBR322, que muestra las localizaciones de los sitios
de las enzimas de restricción que cortan el plásmido por un solo sitio. Estos sitios pueden utilizarse para insertar los fragmentos de DNA a clonar. También se muestran las localizaciones de los genes de resistencia a antibióticos.
5
Este plásmido tienen un origen de replicación (ori), dos genes de selección (resistencia a los antibióticos ampicilina y tetraciclina), y algunos sitios de restricción únicos. Dentro del gen de resistencia a tetraciclina hay sitios de restricción únicos para las enzimas BamHI, SpbI, SalI, XmaIII y NruI, y dentro del gen de resistencia a ampicilina hay sitios de restricción únicos para las enzimas RruI, PvuI, y PstI. Si se introduce un pBR322 en una célula bacteriana sin plásmidos y sensible a antibióticos, la célula se convertirá en resistente a la tetraciclina y a la ampicilina. Si se inserta un fragmento de DNA en los sitios de restricción RruI, PvuI, o PstI, se inactivará el gen de resistencia a ampicilina, pero el gen de resistencia a tetraciclina seguirá activo. Si este plásmido recombinante se transfiere a una célula bacteriana que no contenga plásmidos, las células que contengan el plásmido recombinante podrán identificarse, ya que serán resistentes a tetraciclina y sensibles a ampicilina.
Figura. El plásmido pUC18 ofrece varias ventajas como vector de clonación. Debido a su pequeño tamaño, puede aceptar fragmentos de DNA relativamente grandes; se replica hasta un alto número de copias y tiene un gran número de sitios de restricción en el sitio de clonación múltiple, localizado dentro del gen lacZ. Las bacterias que contienen pUC18 producen colonias de color azul si crecen en un medio que contenga X-gal. El DNA insertado en el sitio de clonación múltiple interrumpe el gen lacZ, resultando en colonias blancas, lo que permite la identificación directa de las colonias que tienen insertos de DNA clonados.
6 Los bacteriófagos lambda y M13
Lambda es un bacteriófago muy utilizado en experimentos de DNA recombinante. Se han identificado y cartografiado todos los genes de lambda, y se conoce toda la secuencia nucleotídica de su genoma. El tercio central de su cromosoma es prescindible, y puede reemplazarse por DNA exógeno sin afectar la capacidad del fago para infectar células y formar calvas. Se han desarrollado más de 100 vectores basados en el fago lambda, eliminado diversas porciones del grupo génico central. Para clonar utilizando el vector lambda, se corta el DNA del fago con una enzima de restricción (por ejemplo, EcoRI), lo que produce un brazo izquierdo, un brazo derecho y una región central. Se aislan los brazos y se ligan (utilizando la DNA ligasa) a un segmento de DNA obtenido tras cortar DNA genómico con la misma enzima de restricción (en este ejemplo EcoRI).
Las moléculas recombinantes resultantes pueden introducirse en células huésped bacterianas de dos maneras: Por transformación. Una vez en las células
huésped, se replican generando fagos infectivos, llevando cada uno de ellos el inserto de DNA. Por infección , se mezcla el DNA de lambda que contiene el inserto con los
componentes proteicos del fago (cabezas y colas); de esta mezcla se forman partículas fágicas infectivas (empaquetamiento “in vitro”). Los fagos pueden amplificarse
haciéndose crecer en placas sembradas con bacterias, donde se formarán calvas, o bien infectando células en un medio líquido y recogiendo las células lisadas.
Figura. Pasos en la
7 Los cósmidos y los vectores transbordadores
Los cósmidos son vectores híbridos construidos utilizando partes del cromosoma del fago lambda y de DNA plasmídico. Los cósmidos contienen la secuencia cos del fago lambda, necesaria para el empaquetamiento del DNA del fago dentro de su cubierta proteica, y secuencias plasmídicas de replicación (ori) y de genes de resistencia a antibióticos, que permiten identificar las células huésped que los contienen. El DNA de los cósmidos que contienen insertos de DNA se empaqueta en la cápside proteica de lambda para formar partículas fágicas infectivas.
Una vez que el cósmido entra en la célula huésped se replica como un plásmido. Puesto que la mayoría del genoma lambda se ha delecionado, los cósmidos pueden transportar insertos de DNA mucho más grandes que los que lambda puede llevar. Los cósmidos pueden transportar casi 50kb de DNA insertado; los vectores fágicos pueden acomodar insertos de DNA de unas 15 kb, y los plásmidos generalmente están limitados a insertos de 5- 10 kb.
Existen otros vectores híbridos, construidos con orígenes de replicación provenientes de distintas fuentes (por ejemplo, plásmidos y virus animales como SV40), que pueden replicarse en más de un tipo de célula huésped. Generalmente, estos vectores transbordadores contienen marcadores genéticos que permiten su
selección en los dos sistemas huésped, y pueden utilizarse para transportar insertos de DNA entre E. coli y otras células huésped como levadura y viceversa.
8
Para cartografiar y analizar genomas eucarióticos complejos, se desarrolló un vector multiuso llamado cromosoma artificial bacteriano (BAC) basado en el factor F de bacterias. Éste es un plásmido que se replica independientemente y está implicado en la transferencia de información genética en la conjugación bacteriana. Como los factores F pueden transportar fragmentos del cromosoma bacteriano de hasta 1 Mb, se diseñaron para que funcionen como vectores de DNA eucariótico. Los vectores BAC tienen los genes de replicación y de número de copia del factor F, e incorporan un marcador de resistencia a un antibiótico y sitios de restricción para insertar el DNA exógeno a clonar. Además, el sitio de clonación está flanqueado por regiones promotoras que pueden utilizarse para generar moléculas de RNA y expresar así el gen clonado, o para utilizarlas como sonda para paseo cromosómico, y para secuenciar el DNA del inserto clonado.
9
Figura. Resumen de los pasos seguidos en la clonación de un vector plasmídico. Los vectores plasmídicos se aíslan y
10 Clonación de DNA en E. coli
Para replicar el DNA clonado se pueden utilizar distintos tipos de células huésped. Uno de los huéspedes más utilizados es la cepa de laboratorio K12 de E. coli.
Ésta y otras cepas de E. coli se utilizan como huéspedes ya que están bien
caracterizadas genéticamente y pueden aceptar un amplio espectro de vectores, incluyendo los plásmidos, los fagos y los cósmidos.
Para crear moléculas de DNA recombinante se precisan varios pasos. Si se utiliza un plásmido como vector, el procedimiento es el siguiente:
1. El DNA que se va a clonar se aísla y se trata con una enzima de restricción para crear fragmentos que acaben en secuencias específicas.
2. Estos fragmentos se ligan a moléculas de plásmido que han sido cortadas con la misma enzima de restricción, obteniéndose un vector recombinante.
3. El vector recombinante se transfiere a células huésped bacterianas, generalmente por transformación, un proceso en el que las moléculas de DNA atraviesan la membrana de la célula huésped transfiriéndose a su interior.
4. La célula huésped se hace crecer en una placa de cultivo, donde formará colonias. Puesto que las células de cada una de las colonias provienen de una sola célula inicial, todas las células de la colonia, y los plásmidos que contienen, son genéticamente idénticas, o clones. Se rastrean las colonias para identificar las que han incorporado el plásmido recombinante.
Se utilizan varios métodos para seleccionar las colonias que contienen plásmidos con el inserto de DNA. Este proceso se denomina rastreo (o cribado). Para vectores como pBR322, el rastreo es un proceso que consiste en dos etapas. Por ejemplo, si el DNA que se desea clonar se inserta dentro del gen de resistencia a tetraciclina, este gen se inactivará. Después de transformar células huéspedes con el plásmido recombinante, éstas se hacen crecer en placas de cultivo que contienen el antibiótico ampicilina. Todas las células que hayan incorporado un plásmido (con o sin inserto) crecerán y formarán colonias, mientras que las células que no lo hayan incorporado morirán ya que son sensibles a la ampicilina.
11
contiene ampicilina a una placa que contiene tetraciclina, debido a que el inserto ha inactivado el gen de resistencia a tetraciclina. Entonces, el patrón de colonias de la placa con tetraciclina se compara con el de la placa con ampicilina, y se identifica las colonias de la placa con ampicilina que no han crecido en la placa con tetraciclina. Las células de estas colonias contienen vectores con el inserto de DNA, y se transfieren a un medio de crecimiento para nuevos análisis.
Figura. Selección de colonias que contienen un vector plasmídico que tiene dos genes de
resistencia a antibióticos, uno para tetraciclina y otro para ampicilina. En este experimento, la presencia del inserto de DNA en el plásmido inactivará el gen de resistencia de tetraciclina.
Otros vectores, como pUC18, tienen construcciones que producen colonias
azules cuando están en células bacterianas sembradas en un medio que contiene una sustancia denominada X-gal. Los sitios de restricción del sitio de clonación múltiple de
pUC18 están dentro del gen lac, el gen responsable de la capacidad de formar colonias
azules, y la inserción de DNA en el sitio de clonación múltiple interrumpe esta capacidad. Como se ha expuesto anteriormente, los plásmidos que contienen los segmentos de DNA insertados producen colonias blancas, mientras que los que no tienen insertos producen colonias azules.
De igual modo, los fagos que contengan DNA exógeno también pueden utilizarse para infectar células huésped de E. coli, y cuando se siembran en un medio
12
un solo fago inicial. Los cósmidos infectan a las células bacterianas como los fagos, pero luego se replican como los plásmidos dentro de la célula huésped. Las células que contiene cósmidos con DNA clonado pueden identificarse y recuperarse de la misma manera que los plásmidos.
Clonación en huéspedes eucarióticos
Hemos descripto la utilización de E. coli como huésped para la clonación.
También pueden utilizarse otras especies de bacterias como huéspedes, como B.
subtilis y Streptomyces. Estos sistemas huésped bacterianos y sus vectores son
parecidos a los descriptos para E. coli. Sin embargo, para estudiar la expresión y
regulación de los genes eucarióticos, a menudo es conveniente e incluso necesario utilizar huéspedes eucarióticos. En esta sección describiremos sistemas de clonación que utilizan células eucarióticas como huéspedes.
Vectores de levadura
Aunque la levadura es un organismo eucariótico, puede manipularse y crecer de manera parecida a las células bacterianas. Además, la genética de las levaduras se ha investigado exhaustivamente, lo que ha proporcionado un gran catálogo de mutaciones y unos mapas genéticos altamente detallados de sus cromosomas. En levadura hay un plásmido de origen natural, denominado plásmido 2 micras (o plásmido 2 µ), que se
ha utilizado para construir varios vectores de clonación para levadura. Combinando secuencias de plásmidos bacterianos con plásmidos 2 micras, se pueden producir vectores con muchas propiedades útiles.
Construcción de bibliotecas de DNA
Puesto que cada segmento de DNA clonado es relativamente pequeño, deben construirse muchos clones diferentes para incluir todas las pequeñas porciones del genoma de un organismo. El conjunto clonado de todas las secuencias genómicas de un solo individuo se denomina biblioteca. Las bibliotecas clonadas pueden provenir del
13 Bibliotecas genómicas
Las bibliotecas genómicas (o genotecas) suelen construirse utilizando vectores fágicos, que pueden contener grandes fragmentos cromosómicos. Para preparar una biblioteca en el fago lambda, se corta el DNA de lambda con una enzima de restricción para eliminar el grupo génico central. El DNA genómico que se desea clonar se corta con la misma enzima, y se purifican los fragmentos cortados de tamaño óptimo para el empaquetamiento (de 15 a 17kb) mediante electroforesis en gel o por centrifugación. Estos fragmentos de DNA se ligan con los brazos del cromosoma de lambda para formar la biblioteca.
En una biblioteca genómica están representados todos los genes de un organismo. La biblioteca es un medio para recuperar cualquier gen del genoma del organismo, pudiéndose estudiar con detalle junto con sus secuencias reguladoras adyacentes. Teóricamente, una biblioteca genómica contiene al menos una copia de todas las secuencias representadas en el genoma. Sin embargo, cada molécula de vector puede contener sólo relativamente pocas kilobases de DNA insertado, por lo que una de las primeras tareas al preparar una biblioteca genómica es seleccionar el vector más adecuado para contener el genoma completo en el menor número posible de clones. El número de clones necesarios para contener todas las secuencias de un genoma depende del tamaño medio de los insertos clonados y del tamaño del genoma a clonar. Este número puede calcularse con la siguiente fórmula:
N= ln (1-P) ln (1- ƒ)
Donde N es el número de clones necesarios, P es la probabilidad de recuperar una secuencia determinada, y ƒ representa la fracción del genoma presente en cada clon.
14 Bibliotecas de cDNA
Se puede construir una biblioteca que represente los genes que se están transcribiendo en una célula eucariótica en un momento dado, utilizando el mRNA aislado de esa célula. Casi todas las moléculas de mRNA eucariótico tiene una cola de poli-A en su extremo 3’. Primero, se hibrida la población de moléculas de mRNA que tienen colas de poli-A 3’ con oligo-dT (DNA corto de cadena sencilla formado sólo por dexositimidina). La secuencia de oligo-dT híbrida con la cola de poli –A, y sirve de cebador para la síntesis de una cadena complementaria de DNA utilizando la enzima
retrotranscriptasa (transcriptasa inversa). Esta enzima es una DNA polimerasa
dependiente de RNA que copia un molde de RNA de cadena sencilla en un DNA de cadena sencilla. El resultado es un dúplex RNA-DNA de doble cadena. La cadena de RNA se elimina y la cadena sencilla de DNA se utiliza como molde para sintetizar la cadena complementaria de DNA, utilizando la DNA polimerasa I. El extremo 3’ de la
cadena sencilla de DNA se dobla sobre sí mismo formando una horquilla (un lazo), por lo que puede servir de cebador para sintetizar la segunda cadena. El resultado es un DNA dúplex con las cadenas unidas por un extremo. La horquilla puede abrirse,
obteniéndose una molécula de DNA de doble cadena (denominada DNA
complementario o cDNA), cuya secuencia nucleotídica deriva de una molécula de
RNA.
15
Si se añade un trozo corto de DNA con sitios de restricción en cada uno de sus extremos, el cDNA puede clonarse. Este sitio de clonación puede cortarse con la enzima de restricción adecuada, produciendo extremos pegajosos, lo que permite que el cDNA se inserte en el sitio de restricción de un vector plasmídico o fágico.
Una biblioteca de cDNA es diferente de una biblioteca genómica ya que representa sólo un subconjunto de todos los genes del genoma. Otra diferencia es que las bibliotecas de cDNA no contienen las secuencias promotoras adyacentes al gen, ni tampoco contienen las secuencias intercaladas o intrones, ya que éstos han sido eliminados del pre-mRNA durante la reacción de corte y empalme y no se encuentran en el mRNA maduro.
Las bibliotecas de cDNA utilizan mRNA como punto de partida, de manera que sólo representan a las secuencias que se están expresando en un tipo celular, en un tejido, o en un estadio concreto del desarrollo embrionario. La decisión de construir una biblioteca genómica o de cDNA depende del problema planteado. Si se está interesado en un gen particular, podría ser más fácil preparar una biblioteca de cDNA del tejido en el que se expresa dicho gen. Por ejemplo, los glóbulos rojos producen grandes cantidades de hemoglobina, y la mayoría del mRNA de estas células es mRNA de la
β-globina. Una biblioteca de cDNA preparada utilizando mRNA aislado de glóbulos rojos permite aislar con facilidad un gen de la globina. Si se interesaran las secuencias reguladoras adyacentes al gen de la globina, se necesitaría construir una biblioteca genómica, ya que estas secuencias reguladoras no se encuentran en el mRNA de la globina, y por lo tanto no estarían representadas en la biblioteca de cDNA.
Identificación de secuencias clonadas específicas
16 Sondas para rastrear clones específicos
Muchos de los protocolos utilizan una sonda para rastrear la biblioteca e identificar el clon que contiene el gen de interés. A menudo, las sondas son polinucleótidos radioactivos que contienen una secuencia de bases complementaria a todo o parte del gen de interés. Otros métodos utilizan sondas que dependen de reacciones químicas o colorimétricas para indicar la localización de un clon específico. Las sondas pueden provenir de distintas fuentes; genes relacionados aislados de otras especies pueden servir de sonda si la secuencia se ha conservado suficientemente. Por ejemplo, se aislaron copias extracromosómicas de genes ribosómicos de la rana
Xenopus laevis por centrifugación, se cortaron con enzimas de restricción, y se
clonaron en vectores plasmídicos. Como las secuencias de los genes ribosómicos se han conservado enormemente durante la evolución de los eucariotas, estos genes clonados de Xenopus se marcaron con radioisótopos y se utilizaron como sonda para
aislar los genes ribosómicos humanos de una biblioteca genómica.
Si el gen que se quiere seleccionar de la biblioteca es transcripcionalmente activo en determinados tipos celulares, se puede utilizar una sonda de cDNA para encontrarlo. Esta técnica es especialmente útil cuando puede obtenerse el mRNA purificado o enriquecido. Se sabe que el mRNA de la $-globina es el RNA mensajero predominante en determinados estadios del desarrollo de los glóbulos rojos. Para hacer una sonda, se aísla el mRNA de estas células, se purifica, y se copia con la retrotranscriptasa en una molécula de cDNA. Este cDNA puede utilizarse como sonda para recuperar el gen de la $-globina de una biblioteca genómica, incluyendo los intrones y las regiones de control adyacentes.
Métodos de análisis de las secuencias clonadas
17 Cartografía de restricción
Un mapa de restricción es la recopilación del número, del orden y de la distancia entre los sitios de corte de enzimas de restricción en un segmento clonado de DNA. Las unidades de mapa se expresan en pares de bases (pb) o, para largas distancias, en pares de kilobases (kb). Los mapas de restricción proporcionan información que puede utilizarse para subclonar fragmentos de un gen, o bien para comparar la organización de un gen y de su cDNA con el fin de identificar los exones y los intrones en la copia genómica del gen.
Los fragmentos generados tras cortar el DNA con enzimas de restricción pueden separarse mediante electroforesis en gel, método que separa los fragmentos por su tamaño, y en el que los fragmentos más pequeños se mueven más rápidamente. Los fragmentos aparecen como una serie de bandas que pueden visualizarse tiñendo el DNA con bromuro de etidio e iluminándolo con luz ultravioleta (siguiente figura). El tamaño de los fragmentos individuales puede determinarse corriendo un conjunto de fragmentos marcadores de tamaño conocido en otro carril del mismo gel.
Figura. Gel de agarosa que contiene fragmentos separados de DNA, teñidos con un colorante
(bromuro de etidio) y visualizados mediante iluminación ultravioleta.
18
enzimas nombradas. Los fragmentos generados por la digestión con las enzimas de restricción se separan por electroforesis. El tamaño de estos fragmentos se estima por comparación con un conjunto de patrones de tamaño separados electroforéticamente en carriles adyacentes del mismo gel. Para construir el mapa, se analizan los fragmentos generados con las enzimas de restricción.
1. Cuando el DNA se corta con HindIII, se producen dos fragmentos, uno de 0,8 kb
y otro de 6,2 kb, indicando que sólo hay un sitio de restricción para esta enzima (y que está localizado a 0,8 kb de uno de los extremos).
2. Cuando el DNA se corta con SalI, se producen dos fragmentos, uno de 1,2 kb y
otro de 5,8 kb, lo que significa que hay un único sitio de restricción localizado a 1,2 kb de uno de los extremos.
En conjunto, estos resultados muestran que hay un sitio de restricción para cada enzima, pero se desconoce la relación existente entre estos dos sitios. Con esta
información, hay dos mapas posibles. En un modelo, el sitio HindIII está a 0,8 kb de
uno de los extremos (modelo I), y el sitio SalI está a 1,2 kb del mismo extremo. En el
modelo alternativo (modelo 2), el sitio HindIII está localizado a 0,8 kb de un extremo, y
el sitio SalI está localizado a 1,2 kb del otro extremo.
El modelo correcto puede determinarse si se consideran los resultados de la digestión de ambas enzimas, HindIII y SalI. El modelo 1 predice que la digestión con
ambas enzimas generará tres fragmentos de 0,4, 0,8 y 6,2 kb; ambas enzimas generarán tres fragmentos de 0,8, 1,2 y 5 kb. El patrón real de fragmentos observado después de la digestión con ambas enzimas indica que el modelo correcto es el modelo 1 de la Figura.
19
manera de localizar sitios de mutación en los genes.
Los mapas de restricción también pueden utilizarse para refinar los mapas génicos. En la mayoría de los casos, la exactitud de los mapas construidos por análisis genéticos se basa tanto en la frecuencia de recombinación entre marcadores genéticos como en el número de marcadores genéticos utilizados en la construcción del mapa. Si hay una gran distancia entre los marcadores y/o variación en la frecuencia de recombinación, el mapa genético puede no corresponder al mapa físico de esa región cromosómica. Por ejemplo, el genoma humano es grande (3,2 x 109 pb), y el número de genes cartografiados es pequeño (unos pocos millares), lo que significa que cada unidad del mapa está compuesta por millones de pares de bases de DNA. El resultado es una correlación baja entre los mapas genético y físico de los cromosomas. Los sitios de corte de las enzimas de restricción pueden utilizarse como marcadores genéticos, reduciendo así la distancia entre los distintos sitos del mapa, incrementando la fidelidad de los mapas, y proporcionando puntos de referencia para la correlación de los mapas genético y físico.
20 Transferencia de Southern y Northern
Los insertos de DNA clonados en vectores pueden utilizarse en experimentos de hibridación para caracterizar la identidad de genes específicos, para localizar regiones codificantes o regiones reguladoras flanqueantes en secuencias clonadas y para investigar la organización molecular de las secuencias genómicas.
Edward Southern desarrolló la utilización de segmentos de DNA clonado, separados por electroforesis, transferidos a filtros, y rastreado con sondas.
21
Conocido como transferencia de Southern (Southern blot), este
procedimiento tiene muchas aplicaciones. En la transferencia de Southern, el DNA clonado se corta en fragmentos con una o más enzimas de restricción, y estos fragmentos se separan por electroforesis en gel (Figura 20). El DNA se desnaturaliza dentro del gel en fragmentos de cadena sencilla, y éstos se transfieren a un filtro de un material, normalmente nitrocelulosa o un derivado de nylon, que une el DNA. La transferencia se hace colocando la hoja de la membrana encima del gel, y provocando que el tampón pase por el gel y por la hoja de nitrocelulosa o de nailon por capilaridad. El tampón pasa por el gel y por la membrana, arrastrando al DNA fuera del gel e inmovilizándolo en la membrana.
En la práctica, esto se hace poniendo el gel con el DNA desnaturalizado sobre una gruesa esponja que actúa de mecha. La esponja está parcialmente sumergida en una cubeta con tampón. Se pone una membrana encima del gel, y se cubre con hojas de papel secante o absorbente y un peso. La acción capilar arrastra el tampón de la cubeta a través de la esponja, del gel, de la membrana, y del montón de papel secante. Al pasar el tampón por el gel, los fragmentos de DNA se transfieren a la membrana, a la que se unen. Después de la transferencia, el DNA de cadena sencilla se fija a la membrana calentándola a 80 ºC o exponiéndola a la luz ultravioleta para que se hagan uniones entre los fragmentos y la membrana.
Entonces, los fragmentos de DNA del filtro se hibridan con una sonda. Sólo formarán híbridos los fragmentos de DNA de cadena sencilla presentes en la membrana que sean complementarios a la secuencia nucleotídica de la sonda. Se lava la sonda no unida, y se visualizan los fragmentos hibridados. Si se utiliza una sonda radioactiva, la posición de la sonda se determina por autorradiografía, utilizando película fotográfica.
22 Figura. Transferencia de Southern.
23
complementario al segmento de DNA clonado. Esto se lleva a cabo extrayendo RNA de uno o varios tipos celulares o tejidos. Se fracciona el RNA por electroforesis en gel, y el patrón de bandas se transfiere a una membrana que une el RNA como en la transferencia de Southern. El filtro se hibrida con una sonda de DNA de cadena sencilla, proveniente de DNA genómico clonado o de cDNA. Si hay RNA complementario a la sonda de DNA, éste se detectará por autorradiografía como una banda en la película fotográfica. Como el protocolo original que utiliza DNA unido a un filtro se conoce como transferencia de Southern, el protocolo que utiliza RNA unido al filtro se denominó transferencia Northern.
La transferencia Northern proporciona información de la presencia de un transcripto de RNA complementario de un gen clonado en una célula o en un tejido determinado, y se utiliza para examinar patrones de expresión génica en tejidos embrionarios y adultos. La transferencia Northern también puede utilizarse para detectar cortes y empalmes alternativos del mRNA, y para detectar múltiples tipos de transcriptos provenientes de un solo gen. La transferencia Northern también proporciona otras informaciones de los mRNA transcriptos. Si se corre un RNA marcador en un carril adyacente, se puede calcular el tamaño del mRNA de un gen de interés. Además, la cantidad de RNA transcripto presente en una célula o en un tejido está relacionado con la densidad de la banda de RNA del film de autorradiografía. Se puede cuantificar la cantidad de RNA midiendo la densidad de la banda, lo que proporciona una medida relativa de la actividad transcripcional. De esta manera, la transferencia Northern puede utilizarse para caracterizar y cuantificar la actividad transcripcional de un gen determinado en distintas células, tejidos, u organismos.
Secuenciación de DNA
24
década de los 70 se desarrollaron métodos más eficientes de secuenciación y, actualmente, en un laboratorio de biología molecular, es posible secuenciar más de mil bases en una semana. En un futuro cercano, con la automatización de este proceso, se podrán secuenciar miles de bases en un solo día.
Un método de secuenciación de DNA diseñado por Alan Maxam y Walter Gilbert utiliza productos químicos para cortar el DNA. Este método se utilizó para determinar la secuencia de bases de los 4.362 nucleótidos del plásmido pBR322. El segundo método, que es el que generalmente se utiliza, lo desarrollaron Frederick Sanger y sus colaboradores. El método de Sanger se basa en la elongación 5’ – 3’ de las moléculas de DNA por la DNA polimerasa. Ambas estrategias generan un conjunto de moléculas de DNA de cadena sencilla de diferentes longitudes. Para determinar la secuencia de los nucleótidos en un segmento de DNA, ambos métodos utilizan una serie de cuatro reacciones, realizadas en tubos separados. Estas secuencias, cuya diferencia en tamaño es de un solo nucleótido, se separan por electroforesis en gel en cuatro carriles adyacentes. El resultado es una serie de bandas que forman un patrón parecido a una escalera. La secuencia puede leerse directamente del patrón de bandas de los cuatro carriles. Los secuenciadores automáticos de DNA utilizan colorantes fluorescentes en vez de sondas radioactivas. Se utilizan cuatro colorantes, produciendo un patrón de picos de colores que, al leerlos, proporcionan la secuencia de DNA.
25
experimentos confirmaron que el producto del locus de la fibrosis quística es una proteína de membrana que regula el transporte de iones cloro por la membrana plasmática. En la mayoría de casos de CF, el gen mutante tiene una secuencia de DNA alterada que causa la producción de una proteína defectuosa.
Además de identificar los defectos del DNA que causan los fenotipos mutantes, la secuenciación de DNA también se utiliza para examinar la organización de un gen (el número de intrones y de exones, y sus límites), para proporcionar información de la naturaleza y de la función de las proteínas codificadas por los genes, como el tamaño, el número y tipo de dominios (región transmembrana, de unión al DNA) y de la relación con proteínas similares y con proteínas de otros organismos.
Figura. La secuenciación del DNA se ha automatizado con la utilización de colorantes
fluorescentes, uno para cada base; las bases se leen en orden de izquierda a derecha.
Reacción en Cadena de la Polimerasa ( PCR)
26
Uno de los prerrequisitos para muchas técnicas de DNA recombinante es la disponibilidad de grandes cantidades de un segmento específico de DNA. Estos se obtenían a menudo después de un trabajo intensivo y tedioso de clonación y reclonación. La PCR permite la amplificación directa de segmentos de DNA específicos sin clonación, y puede utilizarse en fragmentos de DNA que estén presentes, inicialmente, en cantidades infinitesimalmente pequeñas. El método de PCR se basa en la amplificación de un segmento de DNA utilizando DNA polimerasa y cebadores, oligonucleótidos que hibridan con la cadena complementaria a la secuencia a amplificar. Hay tres pasos fundamentales en la reacción de PCR, y la cantidad de DNA amplificado producido sólo está limitado, en teoría, por el número de veces que se repiten estos pasos.
1. El DNA que se requiere amplificar se desnaturaliza en cadenas sencillas. Este DNA no necesita estar ni purificado ni clonado, y puede provenir de distintas fuentes, incluyendo DNA genómico, muestras forenses como sangre seca o semen, muestras almacenadas en registros médicos, pelos, restos momificados, y fósiles. El DNA de doble cadena se desnaturaliza por calor (a unos 90º C) hasta que se disocia en cadenas sencillas, normalmente unos 5 minutos.
2. Los cebadores hibridan al DNA de cadena sencilla. Estos cebadores son
oligonucleótidos sintéticos que hibridan con las secuencias flanqueantes del segmento a amplificar. Generalmente se utilizan dos cebadores diferentes. Cada uno de ellos tiene la secuencia complementaria a una de las dos cadenas del DNA. Los cebadores se alinean con sus extremos 3’ encarados ya que hibridan a cadenas opuestas, como se observa en la figura 22. La utilización de cebadores sintéticos significa que se debe tener alguna información de la secuencia de DNA a amplificar.
3. A la mezcla de reacción se le añade una DNA polimerasa resistente al calor, la polimerasa Taq. La polimerasa extiende los cebadores en dirección 5’ – 3’, utilizando como molde al DNA cadena sencilla unido al cebador. El producto es una molécula de DNA de doble cadena con los cebadores incorporados en el producto final.
27
del ciclo se realiza a una temperatura diferente. El ciclo puede repetirse llevando a cabo otra vez todos los pasos. Empezando con una molécula de DNA, el primer ciclo produce dos moléculas de DNA, dos ciclos producen cuatro, tres ciclos producen ocho, etc. Veinticinco ciclos amplifican varios millones de veces el DNA en cuestión. El proceso es automático y se utiliza una máquina denominada termociclador, que puede programarse para realizar un número predeterminado de ciclos, produciendo grandes cantidades de segmentos de DNA amplificado en unas pocas horas.
28
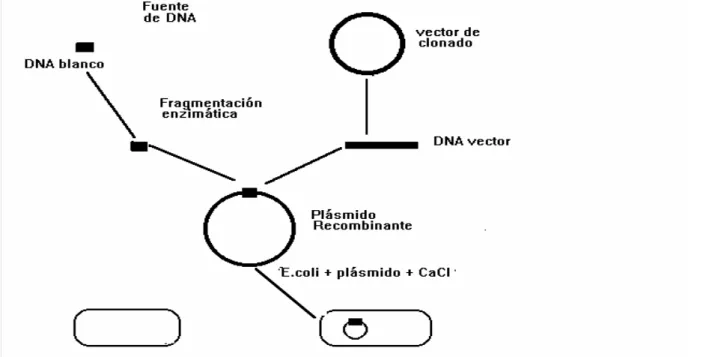
ESTRATEGIASBÁSICASPARAEL CLONADO MOLECULAR
El clonado de ADN basado en células se usa para amplificar ADN con el objeto de poder caracterizarlo físicamente y llevar a cabo estudios funcionales de los genes, de grupos de genes o de otras secuencias de ADN de interés.
Fig. Esquema de clonado molecular en plásmido.
Ejercicio 1
La figura de arriba sintetiza los distintos pasos a seguir en un experimento de clonado básico. Analiza el esquema y describe cada uno de ellos.
HERRAMIENTAS BÁSICAS PARA EL CLONADO
1. ENDONUCLEASAS DE RESTRICCIÓN.
29
Enzima Organismo fuente Sitio de restricción
EcoRI Sau3AI HindII HaeIII BamHI MsoI NotI Escherichia coli Satphylococcus aureus Haemophilus influenzae H. aegiptus Bacillus amyloliquefaciens Moraxela bovis Nococardia otitdis-caviarum I 5’ -G-A-A-T-T-C- -C-T-T-A-A-G- 5’ I I 5’ -N-G-A-T-C-N- -N-C-T-A-G-N- 5’ I I 5’ -G-T-Py-Pu-A-C- -C-A-Pu-Py-T-G- 5’ I I 5’ -G-G-C-C- -C-C-G-G- 5’ I I 5’ -G-G-A-T-C-C- -C-C-T-A-G-G- 5’ I I 5’ -N-G-A-T-C-N- -N-C-T-A-G-N- 5’ I I 5’ -G-C-G-G-C-C-G-C- -C-G-C-C-G-G-C-G- 5’ I Ejercicio 2
a)¿ Qué son las endonucleasas de restricción?
b) ¿Qué características poseen los sitios de reconocimiento de las enzimas de restricción?
c) ¿Qué tipos de corte pueden producir y qué nombres reciben las terminaciones formadas?
d) Además de ser útiles como “ tijeras moleculares” ¿qué otro uso se les da a las endonucleasas de restricción?
2. VECTORES DE CLONADO
Existen vectores naturales tales como plásmidos o bacteriófagos que han sido
30
construidos por el hombre como los cósmidos, cromosomas artificiales de levadura
(YACs) y los cromosomas artificiales de mamíferos (MACs). Los vectores empleados
más frecuentemente comparten las siguientes propiedades: 1. Molécula pequeña, bien caracterizada.
2. Origen de replicación en la molécula que permita su propia reproducción así como también la del fragmento insertado.
3. Fácil recuperación de la molécula híbrida.
Ejercicio 3
a) ¿Qué son los plásmidos y qué características naturales poseen?
b) Los plásmidos naturales han sido modificados para hacerlos más adecuados como vectores de clonado. Observa la estructura del pBR322 y deduce qué condiciones debe reunir un plásmido para que sirva como vector.
EcoR I Hind III
Origen de BamH I replicación Sph I 562 (ORI) Tetr Sal I 651
BstZ I 939
Ampr
Pst I Pvu I Sca I
Fig.2 Esquema del plásmido vector pBR322
c) Volvamos nuevamente a la figura donde se muestra el clonado en plásmidos y supone que el vector usado fue el pBR322. Las moléculas de pBR322 circular purificadas deben ser cortadas con una enzima de restricción para insertar un fragmento de ADN extraño ¿Qué enzima usarías y por qué?.
d) ¿Con qué enzima cortarías el ADN fuente? ¿ Por qué?
31
f) ¿Cómo se origina la molécula quimera cuando el ADN de interés es cortado con una enzima diferente a la empleada para cortar el plásmido?
g) ¿Qué tamaño máximo de fragmentos puede ser clonado en plásmidos?
h) ¿Cómo se introducen las moléculas recombinantes en las bacterias hospedadoras? i) ¿Cómo identificarías las colonias de bacterias que han incorporado los plásmidos con inserto?
j)¿Qué es una biblioteca genómica o genoteca? ¿Cómo se construye?
Bacteriófagos: usaremos como ejemplo el Fago lambda por ser uno de los más
conocidos.
Brazo izquierdo Región no esencial Brazo derecho
Cabeza ciclo lisogénico A B E Z J att xis N cro P S
cos W D F int red cl O Q R cos
Cola inmunidad lisis
0 10 20 30 40 50 60 70 80 90 100
Figura. Mapa sintetizado del cromosoma lineal del bacteriófago lambda mostrando la
posición de algunos genes.
Ejercicio 4
a) ¿Qué son los sitios cos?
b) ¿Qué regiones del lambda pueden ser reemplazadas por ADN exógeno sin afectar su capacidad de infectar bacterias?
c) ¿Cuál es el tamaño máximo de fragmentos que pueden ser insertados en el fago lambda? ¿por qué?
d) ¿Qué ventajas ofrece clonar usando como vector un bacteriófago en lugar de un plásmido?
32
a) ¿Qué son los cósmidos?
b) Analiza las ventajas del clonado en cósmidos en relación con el clonado en los dos vectores anteriores.
VECTORES DE EXPRESIÓN
El objetivo primario del clonado de genes para aplicaciones biotecnológicas es la expresión del gen clonado en el hospedador seleccionado. Pero, la inserción de un gen en un vector no necesariamente asegura que será expresado con éxito. La producción de una proteína a partir de un gen requiere que el mismo sea transcripto adecuadamente y que su mRNA sea traducido. En respuesta a esta necesidad se han creado varios vectores de expresión especializados como el que se muestra a continuación.
Nco I
ros Pst I
Hind III
ptac
T 1 Ampr
T2
ori
Figura. Esquema del vector de expresión pKK233-2. Ampr = gen de resistencia a la amicilina.
ptac = promotor del operon lac. rbs = sitio de unión a los ribosomas . Noc I, Pst I y Hind III = sitios de cortes para esas endonucleasas de restricción. T1 y T2 = secuencias terminadoras de la transcripción.Ori 0 origen de replicación. La flecha indica la
dirección de la transcripción.
Ejercicio 6 Compara al pKK233-2 con el pBR322 ¿Qué similitudes y qué diferencias
33
TÉCNICAS DE RASTREO (SCREENING) POR HIBRIDACIÓN DEL ADN.
Una vez construida la biblioteca genómica, se puede localizar dentro de ella un gen o bien algún segmento de ADN de interés. Se han diseñado numerosas técnicas de sondeo: Hibridización Southern, hibridización Northern, hibridización Western entre otras.
Ejercicio 7
a) ¿Qué es una sonda, qué tipos de sondas existen y para qué sirven?
b) Describe brevemente cada una de las técnicas de sondeo mencionadas arriba.
ADN COMPLEMENTARIO
Ejercicio 8
a) ¿A qué se denomina cADN o ADN copia y cómo se obtiene?
b) ¿Cómo se construye una biblioteca de cADN?
CLONADO POR PCR (REACCIÓN EN CADENA DE LA POLIMERASA)
El PCR es un procedimiento efectivo para obtener grandes cantidades de una secuencia específica de ADN in vitro. Se puede obtener una amplificación de hasta
más de un millón de veces. Para que el PCR ocurra se necesita:
• Dos cebadores o primers (repasar transcripción del ADN) complementarios a regiones sobre cadenas opuestas del ADN a ambos lados de la secuencia a amplificar.
• Una ADN polimerasa termoestable que soporte temperaturas de 95ºC o más (Taq,
ADN polimerasa de Thermus aquaticus). • Los cuatro desoxirribonucleótidos trifosfato.
34
Esta técnica consiste en alrededor de unos 30 ciclos de replicación del ADN. Cada ciclo contiene 3 etapas:
1. Desnaturalización: 95ºC
2. Renaturalización: ~55ºC
3. Síntesis: ~75ºC
Ventajas del clonado por PCR.
• Rapidez: Una reacción típica de 30 ciclos de 3 a 5 min cada uno. El tiempo requerido para el clonado en células es de semanas o meses.
• Sensibilidad de la reacción: Es posible amplificar secuencias a partir de diminutas cantidades de ADN, aún de una única célula.
• No es necesario que el ADN esté muy purificado: Permite amplificar secuencias específicas de material en el que el ADN está muy degradado o embebido en un medio que hace problemático su aislamiento.
Ejercicio 9
a) Esquematiza una reacción en cadena de la polimerasa (PCR) hasta el cuarto ciclo.
BIBLIOGRAFÍA:
GRIFFITHS, A. Genética. Ed. McGraw Hill Interamericana. 5ta edición.
KLUG, W y M. CUMINGS. 1999. Conceptos de Genética. Ed. Prentice Hall. 5ta. ed.
35
Tópico: Organismos
La diversidad biológica actual ha sido dividida en tres dominios basado principalmente en la evidencia molecular, dado que los organismos que pertenecen a un dominio particular han ido evolucionando separadamente de los hallados en los otros dominios durante más de 1.000 millones de años; por esto constituyen las divisiones más profundas en la historia de la vida evolutiva hasta ahora conocida.
LOS CARACTERES QUE DEFINEN A LOS TRES DOMINIOS SON:
Bacteria
Organismos: Termotogales, flavobacterias, cianobacterias, bacterias púrpuras, bacterias gram-positivas, bacterias verdes no-sulfurosas.
Características: Células procarióticas. Membranas lipídicas compuestas principalmente por diésteres de diacil-glicerol. El RNA ribosomal de la subunidad pequeña de los ribosomas (16S-rRNA) es del tipo eubacteriano, es decir, posee un bucle entre las posiciones 500-545.
Dominio Archaea
Organismos: Pyrodictium, Thermoprocteous, termococales, metanococales, metanobacterias, metanomicrobiales, halófilos extremos.
Características: Células procarióticas. Membranas lipídicas compuestas principalmente por diéteres de glicerol isoprenoides o tetraéteres de diglicerol. El RNA ribosomal de la subunidad pequeña de los ribosomas (I6S-rRNA) es del tipo arqueobacteriano, es decir, tiene una estructura única entre las posiciones 180-197 ó 405-498.
Dominio Eukarya
Organismos: Animales, protozoos ciliados, protozoos flagelados, plantas, hongos, diplomonas, algas rojas, euglenoides, microsporidias.
36 Cuadro de los tres dominios
Dominios
Característica Bacteria Archae Eukarya
Núcleo rodeado por membranas ausente ausente Presente Organelas rodadas por membrana ausente ausente presente Peptidoglucanos en la pared celular presente ausente ausente Lípidos de membrana Enlazado por éster
no ramificado
Enlazado por éter ramificado
Enlazado por éster no ramificado
ribosomas 70S 70S 80S
Iniciador del tRNA Formilmetionina Metionina Metionina
Operones Sí Sí No
Plásmidos Sí Sí Raros
RNA polimerasas uno varios Tres
Sensibles a cloranfenicol y a la estreptomicina
Si No No
Ribosomas sensibles a la toxina diftérica
No Sí Sí
Algunos son metanógenos No Sí No
Algunos fijan nitrógeno Sí Sí No
Algunos conducen una fotosíntesis basada en la clorofila
Sí No Sí
(P(
37
Figura. La estructura filogenética más profunda de la diversidad biológica obtenida por Carl
Woese a partir de la secuenciación de rRNA.
Figura. La naturaleza jerárquica de la clasificación biológica consiste en la formación de
38
Los tres dominios Archaebacteria , Bacteria y Eukarya, incluyen a los seis Reinos tal como se muestra:
Dominio Archebacteria: Reino Archaebacteria. Dominio Bacteria: Reino Bacteria (Eubacteria)
Dominio Eukarya: Reinos: Protista, Fungi, Plantae, Animalia.
LOS CARACTERES GENERALES QUE DEFINEN A CADA REINO, SON:
Reino Características
Archaeo-bacteria
Se caracterizan por la ausencia de peptidoglucano en las paredes celulares y la presencia de lípidos de composición distintiva en las membranas celulares que no se encuentra en ninguna otra bacteria. La mayoría vive en lugares calurosos y ácidos
Eubacteria o Bacteria
Células de vida libre; diferenciación celular incipiente en algunos grupos. Incluye a todas las bacterias.
Protista o Protoctista
39
Fungi (Hongos)
Células eucariotas. Formación de esporas y ausencia de flagelos (amastigotas). Las esporas haploides germinan generando hifas que por un proceso de septación más o menos incompleto da lugar a la formación de células. El citoplasma puede fluir en mayor o menor grado a través de la hifa. Al conjunto de hifas se le llama micelio y constituye la estructura visible de la mayor parte de los hongos. Las hifas adyacentes pueden compartir núcleos por conjugación dando lugar a una célula heterocariótica cuyos núcleos se dividen por mitosis y originan una hifa dicariótica. En la reproducción sexual, ambos núcleos se fusionan y forman una célula cigótica diploide que se dividirá por meiosis y formará las nuevas esporas haploides.
Plantae (Plantas)
Organismos multicelulares eucariotas desarrollados a partir de un embrión que no produce una blástula. Las células eucariotas de la mayor parte de las plantas poseen plástidos fotosintéticos, sin embargo, ésta no es una característica exclusiva ni general de las plantas. A diferencia de los animales -cuyas células son en su mayoría diploides- y fungi -cuyas células son haploides o dicarióticas- las plantas alternan de manera ordenada un estadio haploide o de gametofito -donde se producen gametas por mitosis- y otro diploide o de esporofito -donde se producen gametas por meiosis.
40
Animalia (Animales)
Organismos multicelulares eucariotas desarrollados a partir de un embrión que pasa por un estadio de blástula. Aunque la multicelularidad ha surgido independientemente en todos los reinos, en los animales es característica ya que las células están unidas por complejas estructuras como los desmosomas uniones denominadas “gap" y septadas. A diferencia de las plantas, en los animales la meiosis es gamética, es decir, a la reducción cromosómica le sigue inmediatamente la formación de gametas sin posibilidad de originar individuos haploides como el gametofito.
Ejercicio 1
Ubicar en las categorías taxonómicas especificadas (Dominio y Reino) a: Trifolium
repens (alfalfa), Tripanosoma cruzi (agente causante de Mal de Chagas), Dipilidium
caninum (gusano parásito de perros), Ascaris lumbricoides (gusano parásito de
vertebrados), Amynthas hawayanus (lombriz de tierra), Azolla sp (planta flotante), Zea
mays (maíz).
Bibliografía:
CURTIS, H. y N. BARNES. 2000. Biología. Ed. Med. Panamericana. 6ta. Ed.
PURVES, W; D. SADAVA; G. H. ORIANS Y H. CRAIG HELLER. 2003. Vida. La ciencia
41
¿Cómo iniciar el estudio de un animal?
Existen distintos arquetipos que subyacen a la biodiversidad de formas animales con aspectos estructurales comunes que son compartidos por ellos.
Los principales atributos que definen la arquitectura corporal (plan corporal) son: la multiceluridad, la simetría, el diseño del tubo digestivo, la cavidad corporal (celoma), el desarrollo embrionario y la metamería.
42
según la relación de las células que lo componen
Según la simetría y número de capas tisulares
Según su desarrollo embrionario
Según la disposición del tubo digestivo
Según la presencia o ausencia de cavidad corporal
Según el modo de formación del celoma Según la metamería
* *1Equinodermata: es considerado por su
bilateralidad en estadíos larvales.
Animal
Metazoos (mesozoosy Parazoos) (Porifera)
Eumetazoos
Radial-diploblástico
(Cnidaria) Bilateral –
Triploblástico
En saco ciego De tubo en tubo
Acelomados
(Platyhelminthes) Pseudocelomados
(Nematoda)
Celomados
Esquizocelomados (Annelida, Mollusca, Arthropoda)
Enterocelomados
43 Glosario
Metazoos (mesozoos y parazoos): Animales multicelulares con capas celulares no
homólogas a las de los blastodermos de los eumetazoos. Presentan un mayor nivel de organización que el que se encuentra en las colonias de protozoos, no obstante se considera que poseen un grado celular de organización, en el cual la organización
celular es una agregación de células funcionalmente diferenciadas, con división de trabajo, pero con escasa tendencia a organizarse como tejido.
Eumetazoos: Animales multicelulares con una organización estructural, en la cual
combinan sus células en unidades mayores, aquí una célula es una parte especializada del conjunto del organismo que es incapaz de vivir por sí sola. Existe un grado de
organización celular-tisular, en el cual las células similares se agregan según
patrones definidos, de lo cual surgen los tejidos.
Simetría: Es el equilibrio de las proporciones, o correspondencia en tamaño y forma de
las partes o estructuras situadas en lados opuestos de un plano (plano de simetría)
Simetría radial: Cuando el organismo puede quedar dividido en mitades semejantes
por más de dos planos que contengan un eje longitudinal (plano de simetría).
Simetría bilateral: Cuando el organismo puede ser dividido en dos mitades
especulares (derecha e izquierda) en un solo plano sagital.
Diploblástico: disposición básica de tejidos embrionarios con dos capas tisulares,
endodermo y ectodermo.
Triploblástico: disposición básica de tejidos embrionarios con tres capas tisulares,
endodermo, mesodermo y ectodermo.
Sistema digestivo en saco ciego: Existe una abertura que funciona como boca y ano
44
Sistema digestivo de tubo en tubo: Tubo unidireccional, con dos aberturas, boca y
ano.
Celoma: Cavidad llena de fluido que rodea al tubo digestivo, proporcionando un diseño
del tipo “tubo dentro de un tubo”, lo que permite una flexibilidad mucho mayor de la cavidad interna. También supone la disponibilidad de espacio para los órganos viscerales y permite un mayor tamaño y complejidad al dejar mayor superficie expuesta para intercambios celulares. Funciona adicionalmente como un esqueleto hidrostático en ciertos casos, especialmente en muchos gusanos contribuyendo a actividades como traslación y excavación.
Acelomado: No existe cavidad corporal alrededor del tubo digestivo. El espacio entre
la epidermis (ectodérmica) y el tubo digestivo (Endodérmico) está completamente ocupado por una masa esponjosa de células denominada parénquima (mesodermo).
Pseudocelomado: Existe una cavidad corporal alrededor del tubo digestivo limitada
por blastocele persistente que deriva del blastocele embrionario, por ello se denomina a esta cavidad pseudoceloma o pseudocele.
Celomado: Hay una verdadera cavidad desarrollada dentro del mesodermo tapizada
por peritoneo mesodérmico.
45
Metamería (segmentación): la repetición seriada de unidades corporales a lo largo del
eje longitudinal de un organismo. Cada una de esas unidades se denomina segmento o
metámero. Esta disposición afecta a estructuras externas e internas de varios sistemas.
La segmentación del cuerpo (metamería) se llama verdadera o mesodérmica,
cuando existen tabiques internos que separan las secciones sucesivas del animal. Se opone a la llamada segmentación superficial o aparente, que se observa en algunos gusanos – Acantocéfalos, aschelmintos – y dónde sólo la cutícula o la pared corporal, incluso la musculatura, presenta las divisiones sucesivas a lo largo del eje longitudinal, pero faltan los tabiques y no hay repetición de órganos internos. También se observa el pseudomatamerismo – Platelmintos, Nenertinos – cuando en el cuerpo existe una tendencia a la repetición de las partes y de los órganos (por ejemplo, las gónadas), sin que estos sean separados por tabiques. (Novikoff, M. M. 1976.)
Se conoce como Metámero cada uno de los segmentos que se repiten en
ciertos grupos de animales, celomados de simetría bilateral (Bilateria). La
metamerización es una de las principales modificaciones del celoma. Cada
metámero tienen cavidades celómicas separadas de las de otros metámeros por tabiques, y las estructuras internas (ganglios nerviosos, nefridios, gónadas, etc.) y externas (patas, branquias, etc.) están repetidas en cada metámero.
Metamería Homómera: Los segmentos son semejantes entre sí.
Metamería Heterónoma: Los segmentos se fusionan entre sí en grupos funcionales
llamados tagmas para funciones especiales.
Esquizocelomado: El celoma surge de la división de bandas mesodérmicas originadas
a partir de células de la región del blastoporo (abertura al exterior del tubo digestivo primitivo) , las cuales proliferan y se ahuecan para formar la cavidad celómica.
Enterocelomado: El celoma se origina por invaginaciones del tubo digestivo primitivo
46
Protostomado: El blastoporo origina la boca, en tanto que el ano se forma
secundariamente.
Deuterostomado: El blastoporo origina el ano, en tanto que la boca se forma
47
Tagmosis: metamerización heterónoma en tagmas que presentan los Arthropodos
para funciones especializadas.
48
Clase Antenas Patas Partes
bucales
Tagmosis Ejemplos
Crustacea 2 pares N pares y
birrámeas Madíbulas 1 maxila 2 maxila Cefalotórax Abdomen
Insecta 1 par 3 pares y
unirrámeas Madíbulas 1 maxila 2 maxila (labium) Cabeza Tórax Abdomen
Myriapoda 1 par N pares y
unirrámeas Mandíbulas 1 maxila Variable Cabeza Tronco
Chelicerata Ausentes 4 pares y unirrámeos Quelíceros pedipalpos Cefalotórax (prosoma) Abdomen (opistosoma) Ejercicio 2
* Considerando los conceptos expuestos, realizar un cuadro comparativo entre los Phyla más conocidos.
* Elige al menos dos animales mencionados en el ejercicio 1 y descríbelos considerando todos los aspectos mencionados en este apartado.
Ejercicio 3
* Busca un dibujo de pata birrámea y unirrámea, identifica sus partes y explica a qué se debe el nombre e hipotetiza cuál será la importancia adaptativa de cada tipo.
* Completa el cuadro dado arriba mencionando dos ejemplos de cada una de las Clases.
49
BARNES, R. D. 1989. Zoología de los Invertebrados. Ed. Mc Graw Hill Interamericana 5ta.
ed.
CURTIS, H. y S. BARNES 2000. Biología. Ed. Médica Panamericana. 6ta. ed.
HICKMAN, C., L. ROBERTS y A. PARSON. 2002. Principios integrales de Zoología.
Ed. Mc Graw Hill Interamericana 11ma. ed.
PURVES, W; D. SADAVA; G. H. ORIANS Y H. CRAIG HELLER. 2003. Vida. La ciencia
de la Biología. Ed. Médica Panamericana. 6ta. ed.
SOLOMON, E. P; L. R. BERG y D. W. MARTIN, 1999.Biología. Ed. Mc Graw Hill
50
El estudio de los vegetales, como cualquier ser vivo, implica abordarlo desde diferentes perspectivas, entre ellas su morfología externa.
ADAPTACIONES DEL CORMO
No siempre el cuerpo de las plantas superiores sigue, el modelo estructural básico (raíz,
tallo y hoja). Además de la variabilidad en el aspecto externo, que es causada por diferentes
modos de ramificación y simetría, la multitud de formas de los órganos vegetativos aumenta por
modificaciones morfológicas. Estas modificaciones o adaptaciones están generalmente
relacionadas con el ambiente en el que se desarrolla la planta, algunas son la acumulación de
reservas y la existencia de mecanismos de multiplicación vegetativa.
Este texto hace referencia a las adaptaciones del cormo no relacionadas y relacionadas
con la acumulación de reservas.
A-
Adaptaciones independientes de la acumulación de reservas
1- Hoja
a- Espinas foliares: son modificaciones agudas, muy ricas en tejidos de sostén y como
consecuencia, rígidas; pueden ser ramificadas o sencillas. Este tipo de adaptación está difundida
en vegetales típicos de zonas áridas, pero aparece también, como defensa contra herbívoros, en
algunos vegetales no xeromorfos de otras regiones climáticas y en ciertas plantas trepadoras.
Las espinas foliares se producen por transformación de hojas o de partes de éstas (fig. 1).
En Berberis sp, por ejemplo, son parte de la lámina foliar, que además presenta estípulas
modificadas. Éstas se desarrollan también en Acacia, Robinia y Prosopis (algarrobo).
51
b- Zarcillos foliares: son órganos filamentosos, simples o ramificados, haptotrópicos que
la planta utiliza exclusivamente para trepar. Tienen origen por transformación del limbo foliar o
de otra parte de la hoja. Ejemplos: zapallo (Cucurbita pepo) los zarcillos foliares resultan de
hojas reducidas a la nervadura media, arveja (Pisum sativum) los últimos folíolos de la hoja
pueden transformarse en zarcillos ramificados (fig. 2); en Lathyrus aphaca (Leguminosa) el
limbo se ha transformado en un zarcillo simple y su función originaria, es decir, la asimilación
de C2O, es desempeñada por dos grandes estípulas.
Figura 2: Zarcillos foliares en una hoja pinnada de Pisum sativum; con desarrollo de una rama lateral florífera.
c- Filodios: se le da este nombre a un pecíolo dilatado y lámina que sustituye a la lámina
de la hoja, por lo general totalmente abortada. Se presenta en algunas especies de género Acacia
(fig. 3).
52
2- Tallo
a- Espinas caulinares: aparecen como toda rama lateral en la axila de las hojas. Pueden
ser simples o ramificadas, generalmente presentan una foliación reducida, aunque en algunos
casos producen hojas normales con yemas en las axilas. Ejemplo: Crataegus (fig. 4) y Prunus
spinosa.
Figura 4: Espina de Crataegus sp.
b- Zarcillos caulinares: son semejantes en forma y función a los zarcillos foliares, difieren
en su origen. Se originan de una yema axilar, del mismo modo que las ramas laterales normales.
Figura 5: Zarcillo caulinar de Passiflora sp.
c- Braquiblastos: en las plantas superiores la intensidad de crecimiento de las ramas
laterales puede ser muy variada; en unos casos se desarrollan ramas con entrenudos largos, son
los macroblastos, y otras dan lugar a braquiblastos, ramas cortas y con aspecto de roseta (fig. 6).
Los braquiblastos tienen, con frecuencia, una vida limitada, suelen ser simples o
escasamente ramificados. En algunas plantas leñosas las hojas normales, al menos en estado
adulto, se forman únicamente en los braquiblastos por Ejemplo: Prunus cerasus (guindo), Malus
53 Figura 6: Macro y braquiblastos de Pinus sp
d- Cladodios y Filóclados. Cladodios: son ramas de forma comprimida o hasta laminar,
generalmente con hojas rudimentarias, color verde, en la que se realiza, por lo tanto la función
fotosintética.
A pesar de su semejanza con los nomófilos, con quienes coinciden funcionalmente,
difieren de ellos por el origen y por tener crecimiento ilimitado. Es frecuente su desarrollo en
plantas de lugares secos, por ejemplo Cactáceas, en las que además se acumula agua (figs. 7 a y
b).
Figura 7: Opuntia sp tallos aplanados con ramas laterales (aréolas), las hojas de dichas ramas se han transformado en espinas. Derivación de una forma suculenta (a la derecha) a partir de una forma cactácea con hojas (a la izquierda)
Si las ramas laminares fotosintetizantes son braquiblastos y presentan por ello aspecto
54
Figura 8: Rama de Ruscus aculeatus mostrando filóclados con aspecto de hoja, sobre las que aparecen las flores.
e- Estolones: ramas laterales, más o menos delgadas, a menudo muy largas que nacen de
las base de algunos tallos. Los estolones pueden desarrollarse arrastrándose por la superficie de
la tierra y constituyen estolones epígeos, por ejemplo el fresal (fig. 9); o bien pueden hacerlo por
debajo del suelo formando estolones subterráneos, por ejemplo la menta.
Figura 9: Formación de un estolón de fresa (Fragaria sp).
3– Raíz
a- Espinas radicales: entre las Monocotiledóneas, por ej. algunas palmeras
(Acanthorrhiza) producen espinas radicales en la base del tallo, originadas por transformaciones
de las raíces adventicias.
b- Raíces contráctiles: son raíces embrionarias o adventicias encargadas de enterrar más
profundamente la planta o de favorecer su dispersión. Cumplen su función por medio de
55
Se desarrollan preferentemente en las especies con tubérculos, bulbos o rizomas (fig.10).
Figura 10: Raíces contráctiles de Arum maculatum. Hundimiento progresivo del tubérculo por contracción de la raíz: I germinación, II principio del segundo año, III finales de dicho año, IV planta adulta, tubérculo a 10 cm por debajo del suelo.
B- Adaptaciones relacionadas con la acumulación de reservas
1- Hoja
Bulbos: brote subterráneo con los catáfilos o las bases foliares convertidos en órganos
reservantes.
En un bulbo de cebolla (Allium cepa) en corte longitudinal se distingue el tallo en forma
de disco con entrenudos breves, en su extremo se ubica la yema terminal de donde brota el
vástago epígeo con la inflorescencia. Las hojas cercanas al ápice tienen vaina engrosada y lámina
fotosintetizante normal. Las ubicadas en la parte media han perdido la lámina, y se reducen a la
vaina gruesa. Las más externas son muy delgadas, y constituyen las vainas foliares de hojas
viejas que han muerto después de la reabsorción de sus sustancias de reserva.
En otros casos, el bulbo está constituido por catáfilos escamosos y reservantes que se
disponen de manera imbricada, a este tipo se lo llama escamoso, por ejemplo la azucena.
56
este período se reactiva la yema, produciendo hojas y flores a expensas de las reservas
almacenadas en él.
En las axilas de las hojas se producen yemas, cuyas hojas también acumulan reservas en
la base formando nuevos bulbos; éstos al independizarse y formar raíces adventicias contribuyen
a la multiplicación vegetativa de la planta.
2- Tallo
a- Rizoma: tallo subterráneo reservante, con crecimiento horizontal; como vive fuera de
la zona de la luz, carece de nomófilos u hojas propiamente dichas capaces de asimilar o
transpirar; en su lugar hay catáfilos, la mayoría de las veces en forma de escamas membranosas.
Posee yemas y produce vástagos folíferos y floríferos; también raíces adventicias. Podría
confundirse con una raíz, difiere de ella por sus catáfilos y yemas, por no tener caliptra y
principalmente por su anatomía.
Durante el período del año desfavorable a la vegetación, en los países con inviernos fríos
o con estaciones excesivamente secas, el rizoma defiende a la planta contra los rigores del
ambiente.