Análisis de la regulación transcripcional del operón ECP en citrobacter rodentium
68
0
0
Texto completo
(2) UNAM – Dirección General de Bibliotecas Tesis Digitales Restricciones de uso DERECHOS RESERVADOS © PROHIBIDA SU REPRODUCCIÓN TOTAL O PARCIAL Todo el material contenido en esta tesis esta protegido por la Ley Federal del Derecho de Autor (LFDA) de los Estados Unidos Mexicanos (México). El uso de imágenes, fragmentos de videos, y demás material que sea objeto de protección de los derechos de autor, será exclusivamente para fines educativos e informativos y deberá citar la fuente donde la obtuvo mencionando el autor o autores. Cualquier uso distinto como el lucro, reproducción, edición o modificación, será perseguido y sancionado por el respectivo titular de los Derechos de Autor..
(3) Hoja de datos del Jurado . . 1. Datos del alumno Escalera Maurer Andrés (222) 2302535 Universidad Nacional Autónoma de México Facultad de Ciencias Biología 408024505 2. Datos del tutor M. en C. Verónica Iranzú Martínez Santos 3. Datos del sinodal 1 Dr. Alfonso Miguel Torre Blanco 4. Datos del sinodal 2 Dra. Elda Guadalupe Espín Ocampo 5. Datos del sinodal 3 Dra. Bertha María Josefina González Pedrajo 6. Datos del sinodal 4 Dra. Luisa Alvarina Alba Lois 7. Datos del trabajo escrito Análisis de la regulación transcripcional del operón ecp en Citrobacter rodentium. 66 pp. 2012 . 1 . .
(4) El presente trabajo se desarrolló en el Departamento de Microbiología Molecular del Instituto de Biotecnología de la Universidad Nacional Autónoma de México, en Cuernavaca, Morelos, México. En el laboratorio del Dr. José Luis Puente García bajo la asesoría de la M. en C. Verónica Iranzú Martínez Santos. Durante la realización del trabajo se me proporcionó una beca de alimentación y hospedaje otorgada por el Instituto de Biotecnología de febrero 2011 a enero 2012. 2 . .
(5) Índice General 1. Resumen .................................................................................................................................................................... 5 2. Introducción. ............................................................................................................................................................ 6 2.1 Citrobacter rodentium .................................................................................................................................. 6 2.2 C. rodentium como modelo animal ........................................................................................................... 9 2.3 Fimbrias. ......................................................................................................................................................... 10 2.3.1 Regulación. ............................................................................................................................................ 13 2.3.2 Dominio EAL ......................................................................................................................................... 15 2.4 Operones fimbriales en C. rodentium ................................................................................................... 17 2.4.1. “E. coli Common Pilus” ..................................................................................................................... 19 3. Antecedentes ........................................................................................................................................................ 23 4. Justificación ........................................................................................................................................................... 26 5. Objetivos ................................................................................................................................................................. 27 5.1 General. ........................................................................................................................................................... 27 5.2 Particulares. .................................................................................................................................................. 27 6. Materiales y Métodos ......................................................................................................................................... 28 6.1 Cepas, plásmidos y condiciones de crecimiento ............................................................................... 28 6.2 Oligonucleótidos .......................................................................................................................................... 30 6.3 Purificación de DNA cromosomal .......................................................................................................... 32 6.4 Construcción de fusiones transcripcionales ...................................................................................... 33 6.6 Ensayos de actividad enzimática de la cloranfenicol acetil transferasa (CAT). ..................... 35 6.7 Purificación de RNA total .......................................................................................................................... 36 6.8 Determinación del sitio de inicio de la transcripción por “primer extension” ...................... 37 6.9 “Western immunoblot” .............................................................................................................................. 37 6.10 Construcción de la cepa mutante en el gen Rod_29251 ............................................................... 38 6.11 Complementación de las mutantes Δ ROD_29251 y Δ ecpR ......................................................... 39 7. Resultados ............................................................................................................................................................. 40 7.1 Determinación del sitio de inicio de la transcripción de ecpA ..................................................... 40 7.2 Caracterización de la región reguladora de ecp ................................................................................ 43 7.3 Cinética de expresión de ecpA ................................................................................................................. 45 7.4 Condiciones de expresión de ecpA ........................................................................................................ 46 7.5 Papel de EcpR en la expresión de ecpA ................................................................................................ 47 7.6 Papel del ROD_29251 en la expresión de ecpA ................................................................................. 50 7.7 Papel de otros reguladores en la expresión de ecpA ....................................................................... 53 8. Discusión ................................................................................................................................................................ 55 9. Conclusiones. ........................................................................................................................................................ 59 10. Perspectivas. ...................................................................................................................................................... 60 11. Bibliografía. ........................................................................................................................................................ 61 . . 3 . .
(6) Lista de Abreviaturas Ap: ampicilina Cm: cloranfenicol. DO: densidad óptica Kb: Kilo bases. Nal: ácido nalidíxico. Pb: pares de bases. PCR: reacción en cadena de la polimerasa rpm: revoluciones por minuto Stp: estreptomicina TA: temperatura ambiente Tc: tetraciclina. Tm: temperatura de fusión. Wt: silvestre . Lista de figuras Figura 1. Formación de la estructura en forma de pedestal característica de las lesiones A/E. .... 7 Figura 2. Representación esquemática de la isla de patogenicidad LEE de C. rodentium mostrando algunos genes representativos. ............................................................................................ 8 Figura 3. Representación esquemática de la biogénesis de la fimbria tipo 1 por la vía chaperona-‐ usher. ................................................................................................................................................................. 12 Figura 4. Organización general de los operones que codifican para fimbrias ensambladas por la vía chaperona-‐usher. .................................................................................................................................... 12 Figura 5. Organización de dominios en proteínas que contienen el dominio EAL caracterizadas estructural y/o funcionalmente. .............................................................................................................. 17 Figura 6. Comparación entre los operones ecp de C. rodentium y EPEC ............................................... 22 Figura. 7. Modelo de la regulación transcripcional del operón ecp en EPEC y EHEC. ....................... 25 Figura. 8. EcpA se expresa mejor en medio DME a 26ºC en estático. ..................................................... 26 Figura. 9. Identificación del sitio de inicio de la transcripción de ecpA. ............................................... 41 Figura. 10. Caracterización de la región reguladora de ecpA. .................................................................. 42 Figura. 11. “Western blot” para detectar a EcpA-‐FLAG. .............................................................................. 44 Figura. 12. Cinética de expresión de ecpA. ..................................................................................................... 45 Figura. 13. Condiciones de expresión de ecpA. .............................................................................................. 47 Figura. 14. La eliminación de ecpR no afecta expresión de ecpA. .......................................................... 48 Figura. 15. La sobre expresión de EcpR disminuye la producción de EcpA. ....................................... 49 Figura. 16. Características de la proteína codificada por el gen ROD_29251. ..................................... 50 Figura. 17. La proteína ROD_29251 es un regulador positivo de la expresión de ecpA. ................. 52 Figura. 18. Los reguladores específicos de virulencia no tienen un papel en la regulación de ecp ............................................................................................................................................................................. 54 . Lista de tablas Tabla 1. Operones fimbriales identificados en C. rodentium. ................................................................... 18 Tabla 2. Cepas y plásmidos utilizados. ............................................................................................................. 28 . . . 4 . .
(7) 1. Resumen Citrobacter rodentium es una bacteria entérica causante de la hiperplasia colónica transmisible en ratones. Ha sido utilizada como modelo de estudio de los patógenos de humanos EPEC y EHEC debido a que, al igual que éstas, provoca una lesión conocida como de adherencia y destrucción (A/E). La capacidad para generar esta lesión está conferida por la isla de patogenicidad LEE, la cual fue adquirida de manera horizontal por estas tres bacterias. Además de los genes de virulencia codificados en la isla, estas bacterias utilizan adhesinas fimbriales y no fimbriales para colonizar al hospedero. Las fimbrias son filamentos proteicos que se prolongan a partir de la membrana bacteriana y permiten establecer contacto con otras células, tanto bacterianas como eucariontes, así como con superficies abióticas. Involucradas en procesos como inmunomodulación, conjugación, formación de biopelículas, “twitching motility” y adherencia, las fimbrias son, en ocasiones, factores de virulencia. ECP es una fimbria que se ensambla por la vía chaperona/usher y está altamente conservada en E. coli y otras enterobacterias, entre las que se encuentra C. rodentium. Se ha sugerido que ECP tiene un papel en procesos como formación de biopelículas, colonización de plantas (en el caso de EHEC) y colonización del epitelio intestinal (en EPEC y EHEC). Este trabajo tuvo como objetivo estudiar el mecanismo de regulación a nivel transcripcional del operón ecp en C. rodentium. Para cumplirlo, se determinó el sitio de inicio de la transcripción, el cual se localiza a 88 pb corriente arriba del ATG, y con base en el cual se identificaron las cajas -‐10 y -‐35. También se determinó la región reguladora mínima, la cual comprende hasta la posición -‐407 con respecto al sitio de inicio de la transcripción. A diferencia de lo que ocurre en EPEC y EHEC, en C. rodentium EcpR no parece estar involucrada en la regulación del operón, mientras que la proteína codificada por el gen ROD_29251, la cual posee un dominio EAL que podría estar involucrado en la modulación de la concentración del segundo mensajero c-‐di-‐GMP, actúa de manera positiva. Así mismo, se corroboró que la expresión de la fimbria está regulada por las condiciones de crecimiento. De acuerdo con los resultados obtenidos en este estudio, los reguladores específicos de virulencia (Ler, GrlR y GrlA) no juegan un papel en la regulación de ecp. En conjunto estos resultados sientan las bases para el estudio de la regulación de ECP en C. rodentium. . . 5 . .
(8) 2. Introducción 2.1 Citrobacter rodentium C. rodentium es una bacteria Gram-‐negativa causante de la enfermedad denominada hiperplasia colónica transmisible murina (TMCH, por sus siglas en inglés). Esta enfermedad puede ser mortal en ratones lactantes y ciertas cepas endogámicas, sin embargo, en ratones adultos la infección normalmente no es letal y C. rodentium se puede encontrar como parte de la microbiota comensal [33, 54]. El desarrollo de la infección por esta bacteria comienza pocas horas después de la inoculación oral, cuando ocurre la colonización en la superficie del tejido linfoide del ciego y, tres días después, las bacterias pueden ser encontradas también en el colon distal. La lesión principal, visible entre los días 5 y 14 post-‐inoculación (pi), es el engrosamiento del intestino comenzando en el colon distal y, en algunos casos, se extiende al resto del colon. El desarrollo de la lesión es acompañado por la producción de heces suaves. Entre los días 21-‐28 pi, C. rodentium deja de ser detectable, a las 7 semanas pi las lesiones desaparecen completamente y los ratones desarrollan inmunidad [32, 55]. C. rodentium es miembro de la familia Enterobacteriaceae, está relacionada genéticamente con Escherichia coli y conforma, junto con E. coli enteropatógena (EPEC) y E. coli enterohemorrágica (EHEC), un grupo de bacterias patógenas capaces de generar lesiones conocidas como de adherencia y destrucción (A/E) en el tracto gastrointestinal del hospedero [32, 55]. Aunque C. rodentium se encuentra menos relacionada a E. coli que a Salmonella (tomando como referencia sólo los genes de mantenimiento), existen un número significativo de genes, ausentes en E. coli K-‐12 pero conservados en C. rodentium, EHEC y EPEC, que se reconocen como factores de virulencia y que se encuentran principalmente en elementos genéticos móviles. Es la adquisición de estos factores la responsable de la convergencia en la estrategia de virulencia de estos tres patógenos [37]. 6 . .
(9) Las lesiones A/E son esenciales para la patogenicidad de estas bacterias y se caracterizan por la adherencia íntima a la membrana de las células epiteliales, la destrucción localizada de las microvellocidades y la formación de una estructura característica de copa o pedestal (Fig. 1)[32, 55]. La formación de esta lesión se da cuando la bacteria transloca hacia el citoplasma de la célula hospedera, a través de un sistema de secreción tipo 3 (SST3), una serie de proteínas efectoras entre las cuales se encuentra Tir. Esta proteína se integra en la membrana de la célula eucarionte donde actúa como receptor para una proteína de membrana externa de la bacteria llamada intimina y, por otro lado, interactúa con proteínas de la célula hospedera. El reclutamiento de proteínas mediado por Tir en el interior de la célula eucarionte, provoca una modificación del citoesqueleto de actina que induce la formación de la estructura en forma de pedestal debajo de la bacteria [32, 55]. Patógeno AE!. Intimina!. Polimerización localizada de actina.! ! !. Adherencia íntima!. Fig. 1. Formación de la estructura en forma de pedestal característica de las lesiones A/E. La bacteria transloca hacia el citoplasma de la célula hospedera varias proteínas efectoras, entre las cuales se encuentra el receptor translocado de intimina (Tir). Tir se inserta en la membrana de la célula hospedera y funciona como receptor para la proteína de membrana externa intimina. Por otro lado, Tir dispara la cascada de señalización que lleva al . 7 . .
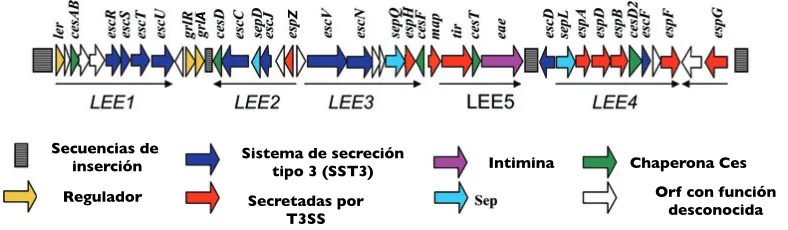
(10) reclutamiento de proteínas de la célula hospedera y la formación del pedestal. Modificado de Strynadka y Ness, 2002. . La habilidad para inducir las lesiones A/E es conferida por uno de los elementos móviles adquiridos por transferencia horizontal, la isla de patogenicidad LEE (“Locus of enterocyte effacement”). La isla LEE está formada por 5 operones policistrónicos que codifican para las proteínas del sistema de secreción tipo 3, las proteínas de adhesión Tir e intimina y las proteínas translocadoras y efectoras; así como un operón bicistrónico que codifica para los reguladores GrlR y GrlA (Fig. 2) [2, 55, 57]. La organización de los genes del LEE es casi idéntica en EPEC, EHEC y C. rodentium, sin embargo su localización dentro del cromosoma es diferente [33, 37, 47, 55]. . Secuencias de! inserción ! Regulador!. Z!. A!. 4 R. Mundy et al.. Sistema de secreción tipo 3 (SST3)!. Intimina!. Chaperona Ces! Orf con función desconocida!. Secretadas por T3SS!. Fig. 3. Gene organization, and assigned function, along the LEE region of C. rodentium.. Fig. 2. Representación esquemática de la isla de patogenicidad LEE de C. rodentium A C. rodentium espB mutant is avirulent (Newman et al., rodentium and EPEC. However, significant differences 1999). were observed betweengenes colonization dynamics and the mostrando algunos representativos. La isla LEE contiene 41 marcos de lectura abiertos MapEPEC was reported to be targeted to, and to interfere extent of colonic inflammation and hyperplasia of the wild(ORF’s), arreglados en 5 operones policistrónicos, denominados LEE1 a LEE5, y un operón type (wt) C. rodentium and DBS255 (eaeEPECα). In particwith function of, the mitochondria, to disrupt intestinal bicistrónico q ue c odifica p ara l os r eguladores G rlRA. Los operones LEE1 a LEE3 contienen los barrier functions and to induce formation of filopodia-like ular DBS255 (eaeEPECα) colonizes mice faster, with higher genes que codifican T3SS, operón LEE4 contiene la bacterial mayoría de during los genes extensions at the site of adhesion initial que colony-forming units (cfu) para isolatedel from stools el over the of infection (reviewed Garmendia et al., 2005). first 5 dayspara of infection (R. Mundyefectoras and G. Frankel, unpubl. codifican proteínas y el operón stages LEE5 codifica para inlas proteínas necesarias Infection of C3H/HeJ and C57Bl/6 mice with three indedata), induction of a stronger inflammatory response and para la adherencia íntima. Modificado de Mundy pendent et al., map 2005. ecp mutants has demonstrated that Map is not deeper penetration within the colonic crypts (Frankel. . et al., 1996). In contrast complementation of DBS255 with a plasmid encoding eaeEHECγ did not restore virulence (reviewed in Frankel et al., 2001). All A/E pathogens translocate their own intimin receptor, Tir, which integrates into the host cell plasma membrane. Tir is involved in a series of interactions with host cell proteins resulting in the formation of actin-rich pedestals beneath the adherent bacteria (reviewed in Frankel et al., 2001). A C. rodentium tir mutant was incapable of forming pedestals on HeLa cells; actin pedestal formation was restored by plasmids encoding TirCR or TirEPEC but not TirEHEC. These results are consistent with the fact that pedestal formation induced by EPEC and C. rodentium is dependent on phosphorylation of tyrosine at positions 474 and 471 (reviewed in Garmendia et al., 2005) and that TirCRY471F was unable to complement C. rodentium ∆tir in vitro (Deng et al., 2003). In vivo, ∆tirCR was unable to colonize mice, but colonization and virulence were restored by plasmid-encoded wt copies of TirCR, TirEPEC or TirEHEC. Interestingly, plasmid. . Modificado de Mundy et al 2005!. essential for colonization and disease (Deng et al., 2004; Mundy et al., 2004). However, map mutants have an intermediate phenotype in C3H/HeJ mice, being recovered in significantly lower numbers than the wt strain over the whole course of infection and persist for longer in the mouse colon (Mundy et al., 2004). In a mixed infection with 50% wt and 50% ∆map, the latter strain was extensively out-competed. Consistent with this finding, map mutants were found among the attenuated strains in both C. rodentium and EHEC STM screens (Mundy et al., 2003) suggesting that Map expression is advantageous in a competitive environment. EspF is also targeted to the mitochondria where it is involved in permeabilization of the mitochondrial membrane and induction of cell death. EspF has a role in disruption of intestinal barrier functions and in remodelling of the brush border microvilli (reviewed in Garmendia et al., 2005). Studies of EspF in the C. rodentium model have been carried out independently by three different groups. The espF mutants were made in strains: DBS100. 8 . .
(11) 2.2 C. rodentium como modelo animal Las enfermedades diarreicas constituyen un problema de salud importante en el mundo causando alrededor de 2 millones de muertes al año. Un número significativo de estos casos esta relacionado con cepas de E. coli diarreogénica, en particular EHEC y EPEC [6, 20]. EPEC es una de las principales causas de diarrea infantil en países en vías de desarrollo. Estudios realizados en Brasil, México, Sudáfrica y Bangladesh han demostrado que entre 30 y 40% de la diarrea infantil es causada por EPEC y, se estiman varios cientos de miles de muertes al año por esta causa [6]. La infección por EHEC causa colitis hemorrágica, diarrea sin sangre y el síndrome urémico hemolítico (este último como resultado de la producción de la toxina tipo Shiga). El principal reservorio de EHEC es el tracto intestinal bovino y la enfermedad se asocia con el consumo de alimento y agua contaminados. A diferencia de EPEC, los brotes de EHEC han sido detectados solamente en países desarrollados, incluyendo EUA, Japón y Reino Unido. La dosis infecciosa en EHEC es mucho menor a la de EPEC, estimada en menos de 100 bacterias [19]. Aunque se han detectado infecciones por patógenos A/E en diversos animales, entre los que se encuentran: reces, cerdos, cabras, borregos, corderos, caballos, gatos, perros, conejos y algunos primates, la única bacteria A/E conocida capaz de causar enfermedad en ratones es C. rodentium [55]. De los animales anteriormente mencionados, los ratones presentan varias ventajas como modelo animal para estudiar este tipo de enfermedades. La gran cantidad de información con la que se cuenta, el elevado numero de mutantes y cepas disponibles, la facilidad de manejo y el bajo costo (en comparación con el de otros animales) [33], hacen de la TMCH un modelo excelente para la investigación in vivo de las interacciones patógeno-‐ hospedero, brindando la posibilidad de manipular ambos para estudiar fenómenos como el reconocimiento y eliminación del patógeno, así como su transmisión [33]. Diversos estudios con C. rodentium han demostrado la eficacia de este modelo. Mutaciones de los genes de la isla LEE y de algunos efectores o factores de virulencia 9 . .
(12) fuera de ésta han permitido determinar la importancia que cada uno tiene para la virulencia [10, 32, 55], confirmando la información que se ha observado para EHEC y EPEC en ensayos con voluntarios, otros modelos animales y células en cultivo, así como aportando nueva información. Estudios con eliminaciones específicas en genes del sistema inmune han generado información acerca de la relación entre componentes particulares de éste, la inmunidad y la patología. De igual manera, se ha usado el modelo para estudiar el efecto de la microbiota en el desarrollo de la enfermedad y para estudios de transmisión. Además, la hiperplasia colónica se asocia con un aumento en la susceptibilidad a carcinógenos y algunas características de la infección por C. rodentium guardan parecido con los modelos animales para las enfermedades inflamatorias intestinales, por lo que la TMCH puede servir también como modelo para el estudio de estas enfermedades [33]. 2.3 Fimbrias. La adherencia es un evento importante en la colonización de superficies hospederas y la promoción de ésta por fimbrias es, por lo regular, un paso esencial en la infección [50]. Las estructuras de superficie más comúnmente involucradas en las primeras interacciones de las bacterias con la superficie de las células del hospedero son las fimbrias o pili, las cuales generalmente se unen a receptores específicos en el hospedero [35]. Las fimbrias son filamentos proteicos no flagelares que se prolongan a partir de la superficie bacteriana. Están presentes tanto en bacterias Gram-‐negativas como Gram-‐ positivas y se ha visto que están involucradas en procesos como conjugación, adherencia, formación de biopelículas e inmunomodulación [40]. A diferencia de los flagelos, las fimbrias no están involucradas en motilidad, excepto las fimbrias tipo IV, las cuales median una motilidad especializada en superficies semisólidas denominada 10 . .
(13) “twitching motility” [17]. Las bacterias Gram-‐negativas presentan una gran diversidad de fimbrias que se forman mediante polimerización no covalente de varias subunidades o pilinas [17]. Algunas fimbrias son factores de virulencia y son considerados como blancos para la elaboración de vacunas; además, las características adhesivas de éstas determinan el hospedero a infectar y el tropismo dentro de éste [35]. Entre los tipos de fimbrias más estudiados se encuentran aquellas que se ensamblan por la vía chaperona-‐usher. En esta vía, las pilinas son secretadas por medio de la vía general de secreción (SEC) al periplasma, donde se unen a chaperonas específicas que ayudan al plegamiento e impiden el ensamblaje prematuro de las subunidades. Posteriormente, el complejo pilina/chaperona es reconocido por la proteína “usher” que forma un poro en la membrana externa y sirve como plataforma para el ensamblaje del pilus (Fig. 3) [34]. Fimbria tipo I. =$)?* =$)<*. !12,'*3$)4#$&'#*. =$)=*. 5'#$&&'*6"&*%$&1(* !"##$%$&##$'()(*+,-,.'$ $,.$/+012$. =$)>*. =$)@*. .0*. !"#$%&'()'*. =$)+*. 7#'2(&-892*:"8*;0<* +$,-%&'()'*. ./*. . 11 . .
(14) Fig. 3. Representación esquemática de la biogénesis de la fimbria tipo 1 por la vía chaperona-‐ usher. ME-‐ membrana externa, MI-‐ membrana interna. En verde se muestra la proteína “usher” y en azul la chaperona. Modificado de Nishiyama et al., 2005. . Los genes que codifican para fimbrias ensambladas por la vía chaperona-‐usher regularmente están agrupados en operones, los cuales codifican para al menos 3 proteínas distintas: la subunidad principal, la chaperona periplásmica y la proteína “usher”. Además pueden codificar también para proteínas estructurales adicionales (ej. subunidades fimbriales menores), proteínas de ensamblaje (ej. chaperonas) o proteínas reguladoras [35]. Todos los operones fimbriales examinados hasta ahora codifican al menos para una proteína reguladora que activa o reprime la expresión de los genes del operón. La mayoría de estos operones comparten una organización genética similar: al principio se encuentran los genes involucrados en la regulación positiva o negativa, seguidos de la subunidad estructural principal. Los genes que codifican para la chaperona, la proteína “usher”, proteínas adaptadoras y la adhesina fimbrial constituyen el resto del operón (Fig. 4). (#)*+,-.%&'. /,0*.&,$.,'1#'#&2,34+,5#'1#+'".+*2'. 6*4*&.1,1#2'1#'+,'"*&7,'8.4$.+,$'1#+'".+*2''. !""""""#"""""""""""$"""""""""""""""%""""""""""""""""""""""""""""&""""""""""""""""""""""""""""'""""""""""""""""("""""""""""""""")""""""""""""""*""""""""""""""""+"""""""""""""""""," 6*4*&.1,1' 9&-+,'1#+' ".+*2' "$.&-.",+'. :2;#$'1#' 3#34$,&,'#<7#$&,'. =;,"#$>&,' "#$."+?23.-,'. 91,"7,1>$@'=>3">&#&7#' 91,"7,1>$@' 91;#2.&,'1#' .&.-.,1>$' "$.&-.",+'1#'+,' .&.-.,1>$' *&.%&' "*&7,'8.4$.+,$'. !"#$%&'!"!' (#)*+,-.%&'. /,0*.&,$.,'1#'#&2,34+,5#'1#+'".+*2' 6*4*&.1,1#2'1#'+,'"*&7,'8.4$.+,$'1#+'".+*2''. #""""""*"""""""""""$""""""""""""""""!"""""""""""""""&""""""""""""""""""""""""""""'"""""""""""""""""""""""""""""+"""""""""""""","""""""""""""""""""%" 6*4*&.1,1' "$.&-.",+'. =;,"#$>&,' :2;#$'1#' "#$."+?23.-,' 3#34$,&,'#<7#$&,'. 91,"7,1>$#2@' .&.-.,1>$#2@' 7#$3.&,1>$#2'. 91;#2.&,'1#' *&.%&'. !"#$%&'#$%'. . Fig. 4. Organización general de los operones que codifican para fimbrias ensambladas por la vía chaperona-‐usher: operón pap (panel superior) y operón fim (panel inferior). Al inicio de . 12 . .
(15) ambos operones se encuentran genes que codifican para proteínas reguladoras; inmediatamente después se encuentra el gen que codifica para la subunidad principal; hacia la mitad de los operones se encuentran los genes que codifican para la maquinaria de ensamblaje, compuesta por la chaperona y la proteína “usher”. Hacia el final de cada operón están los genes que codifican para las subunidades de la punta fibrilar del pilus, las cuales pueden variar en número, así como la adhesina. Modificado de Thanassi et al., 1998. . Ejemplos de operones que conservan esta organización son el operón fim, que codifica para las fimbrias tipo 1 (distribuidas a lo largo de la familia Enterobacteriaceae) y el operón pap, que codifica para las fimbrias P (descritas en E. coli uropatógena, UPEC). El operón fim está compuesto por los genes fimA-‐H (Fig. 4 panel inferior). FimA es la subunidad principal que forma la barra de entre 6 y 9 nm de ancho y de 1 a 2 !m de largo y se conecta, a través de FimF, a una punta formada por FimG y FimH, ésta última es una adhesina que reconoce receptores con manosa [40]. Los genes fimD y fimC codifican para una proteína de andamiaje y una chaperona, respectivamente [40]. Por su parte, el operón pap está compuesto por 11 genes denominados papIBAHCDJKEFG (Fig. 4 panel superior). Éstos codifican para las proteínas reguladoras PapI/B, la proteína “usher” (PapC), la chaperona (PapG) y la pilina principal (PapA). La punta fimbrilar está compuesta principalmente por PapE y PapK conectadas por la proteína adaptadora PapF a la adhesina PapG que reconoce glicolípidos con galactosa [17]. 2.3.1 Regulación. El hecho de que en el genoma de una bacteria se encuentren codificados varios operones fimbriales cuyo producto tiene características que le permiten, en muchos casos, cumplir funciones específicas, hace necesaria una regulación estricta de su expresión. Esta regulación debe estar acoplada a señales externas que le permitan identificar el momento adecuado para su producción. Por ejemplo, en el caso de la virulencia, ciertas fimbrias deben ser producidas durante la adherencia inicial pero es necesario que se dejen de producir en etapas posteriores de la infección para permitir . 13 . .
(16) su dispersión, tal es el caso de la fimbria tipo IV BFP (“bundle forming pilus”) de EPEC [22]. Algunos de los mecanismos de regulación transcripcional descritos hasta el momento para la producción de fimbrias ensambladas por la vía chaperona/usher son: elementos invertibles, metilación del DNA, señalización por c-‐di-‐GMP y unión de proteínas reguladoras a DNA. A continuación se describen brevemente algunos ejemplos de cada uno de estos mecanismos. La regulación mediada por un elemento invertible de la fimbria tipo I de E. coli ha sido ampliamente estudiada. La transcripción del operón fim es dependiente de la orientación de una región de 314 pb localizada corriente arriba de fimA denominada fimS. Esta región contiene al promotor y está flanqueada por dos secuencias repetidas invertidas de 9 pb. La transcripción del operón se da cuando fimS está orientado a favor de fimA (posición ON), mientras que en la orientación contraria (posición OFF) no hay transcripción. La inversión de fimS está mediada por la actividad de las recombinasas sitio-‐específicas FimB y E, las cuales actúan de manera independiente para prender o apagar la expresión [8]. FimE media la inversión de fimS principalmente de la posición ON a OFF, mientras que FimB puede mediar la inversión en ambas direcciones. Además de estas dos recombinasas, la expresión del operón fim también es modulada positivamente por los reguladores Lrp e IHF y negativamente por H-‐NS [8]. Un caso de regulación de fimbrias que involucra la metilación del DNA se observa en la fimbria P de UPEC. El control de la expresión de la subunidad principal PapA está determinado por el estado de metilación de dos sitios con secuencia GATC que se encuentran corriente arriba del promotor papAB. La transcripción de pap está influenciada por el regulador global Lrp y el específico PapI. A concentraciones altas de Lrp y relativamente bajas de PapI, Lrp se une a la región GATC más cercana al promotor inhibiendo la transcripción. Esta unión sólo se puede dar si los sitios no están metilados. Lrp compite con la metilasa de adeninas de DNA (Dam) por la unión a 14 . .
(17) este sitio. Después de la replicación del DNA y la división celular, las bajas concentraciones relativas de Lrp con respecto a las de PapI permiten que Lrp se una al sitio GATC distal impidiendo la metilación de éste pero permitiendo la del sitio proximal y así, la transcripción. Otras fimbrias en las que la metilación del DNA funciona como mecanismo de regulación son las K88, K99 y Sfa de E. coli. [8]. En Klebsiella pneumoniae la síntesis de la fimbria tipo 3, la cual está involucrada en virulencia y formación de biopelículas, se regula mediante la modulación de las concentraciones intracelulares de c-‐di-‐GMP. El incremento o disminución de este segundo mensajero depende de los cambios en la actividad relativa de las proteínas YfiN (con actividad de diguanilato ciclasa) y MrKJ (con actividad de fosfodiesterasa). El receptor de c-‐di-‐GMP, MrKH (que contiene un dominio PilZ de unión a este nucleótido), se une a la región reguladora del operón mrk en presencia del segundo mensajero, activando la transcripción [19, 56]. En esta misma bacteria, la proteína FimK, codificada por el último gen del operón fim (fimK), tiene un dominio EAL y se ha reportado que actúa como regulador negativo de la producción de esta fimbria [8, 42]. La represión y activación de la transcripción mediante la unión de proteínas a sitios en, o cercanos a, la región promotora, es otro de los mecanismos que intervienen en el control de la producción de fimbrias. Un ejemplo de un operón fimbrial regulado por este mecanismo es el de la fimbria Lpf (“long polar fimbria”), en cuya expresión intervienen el regulador global H-‐NS y el específico de virulencia Ler [8, 52]. En este caso, H-‐NS se une cerca de la región promotora, impidiendo la actividad de la RNA polimerasa, mientras que Ler compite con H-‐NS por la unión al DNA, desplazándola y permitiendo así la expresión del operón [8, 57]. 2.3.2 Dominio EAL El Bis-‐(3´-‐5´)-‐guanosín monofosfato cíclico dimérico (c-‐di-‐GMP) es un segundo mensajero global en bacterias. La señalización mediante este segundo mensajero se . 15 . .
(18) encuentra relacionada con la regulación de procesos como motilidad, virulencia, síntesis de fimbrias y formación de biopelículas, entre otros [43]. Las proteínas con dominios GGDEF (con actividad de diguanilato ciclasa, DGC) y EAL (con actividad de fosfodiesterasa, PDE), nombrados así por algunos de sus aminoácidos conservados, están involucradas en la síntesis y la degradación, respectivamente, del c-‐di-‐GMP [48]. Este tipo de dominios se encuentran distribuidos a lo largo de la mayoría de los genomas bacterianos y, en algunas especies, se encuentran ampliamente representados. Ejemplos de esto se observan en Pseudomonas aeruginosa con 53; E. coli con 36; Salmonella Typhimurium con 26 y Vibrio cholerae con 53 proteínas con uno o ambos dominios [13]. El dominio EAL cataliza la hidrólisis de c-‐di-‐GMP para formar el dinucleótido lineal 5´-‐fosfoguanilil-‐(3´-‐5´)-‐guanosina (pGpG), para lo que requiere de Mg2+ o Mn2+; mientras que el Zn+2 y el Ca+2 interfieren con su actividad. Aunque muchos dominios EAL conservan la actividad catalítica, se ha demostrado que algunos carecen de ésta y funcionan como sitios de unión a ligando o de unión a otras proteínas [53]. En algunos casos, el dominio EAL se puede encontrar como dominio funcional único, sin embargo, normalmente se encuentra en el extremo C-‐terminal de proteínas que contienen otros dominios funcionales en el extremo N-‐terminal, los cuales pueden funcionar como receptores de señal, moduladores de la actividad del dominio EAL o cumplir funciones de unión a DNA, entre otras. En la Fig. 5 se muestran algunos ejemplos de proteínas que además de tener el domino EAL presentan dominios como: GGDEF (DGC), HTH (unión a DNA) , BLUF (detección de luz azul), REC (receptor de respuesta), GAF (dominio de unión a cGMP) y PAS (detección de luz, O2 y potencial redox) [48]. . . . 16 . .
(19) Fig. 5. Organización de dominios en proteínas que contienen el dominio EAL caracterizadas estructural y/o funcionalmente. Los dominios GGDEF y EAL se muestran en verde y azul respectivamente. Los dominios reguladores se muestran en rojo y gris. El número total de residuos de aminoácidos se muestra en el extremo C-‐terminal. Los sitios importantes del dominio EAL incluyen: E188, N239, E272, D39 y D303 (que sirven para la coordinación del metal; mostrados en verde), E359 (que posiblemente actúa como base y en la coordinación del metal; mostrado en morado) y K323 (que es una lisina probablemente catalítica; mostrada en azul). Los residuos no conservados de estos sitios se muestran en gris. Modificado de Shirmer et al., 2009. . . 2.4 Operones fimbriales en C. rodentium C. rodentium tiene un total de 19 operones fimbriales (Tabla 1), de los cuales algunos se encuentran incompletos. De aquellos que se encuentran intactos, los productos de los operones kfc (“K99-‐like Factor Involved in Citrobacter Colonization”) y cfc (“Colonization Factor Citrobacter”) han sido relacionados con la colonización del tracto gastrointestinal, mientras que la fimbria polar larga, Lpf ,parece no tiene ningún papel en la virulencia [38]. La fimbria CFC está codificada por un probable operón de 12 genes y tiene homología con las fimbrias tipo IV CFA/III (“colonization factor antigen III”) de E. coli enterotoxigénica (ETEC), TCP (“toxin coregulatied pilus”) de Vibrio cholerae y BFP de EPEC. Las mutantes para dos de los genes del operón (cfcH y cfcI), son avirulentas y pierden la capacidad para colonizar el intestino [32]. Por su 17 . .
Figure



+7

![Fig.
7.
Modelo
de
la
regulación
transcripcional
del
operón
a
una
secuencia
que
sobrelapa
con
la
caja
TTCCT
distal
[25]](https://thumb-us.123doks.com/thumbv2/123dok_es/5390447.103284/27.612.111.500.73.236/modelo-regulacion-transcripcional-operon-secuencia-sobrelapa-ttcct-distal.webp)




Documento similar
"No porque las dos, que vinieron de Valencia, no merecieran ese favor, pues eran entrambas de tan grande espíritu […] La razón porque no vió Coronas para ellas, sería
que hasta que llegue el tiempo en que su regia planta ; | pise el hispano suelo... que hasta que el
Para ello, trabajaremos con una colección de cartas redactadas desde allí, impresa en Évora en 1598 y otros documentos jesuitas: el Sumario de las cosas de Japón (1583),
Habiendo organizado un movimiento revolucionario en Valencia a principios de 1929 y persistido en las reuniones conspirativo-constitucionalistas desde entonces —cierto que a aquellas
En cuarto lugar, se establecen unos medios para la actuación de re- fuerzo de la Cohesión (conducción y coordinación de las políticas eco- nómicas nacionales, políticas y acciones
Así, antes de adoptar una medida de salvaguardia, la Comisión tenía una reunión con los representantes del Estado cuyas productos iban a ser sometidos a la medida y ofrecía
b) El Tribunal Constitucional se encuadra dentro de una organiza- ción jurídico constitucional que asume la supremacía de los dere- chos fundamentales y que reconoce la separación
A partir de los resultados de este análisis en los que la entrevistadora es la protagonista frente a los entrevistados, la información política veraz, que se supone que
