Diversity of root nodule bacteria isolated from common bean (Phaseolus vulgaris L ) in Cuba: effect of Rhizobium – PGPR inoculation and IAA synthesis on development of bean genotypes
164
0
0
Texto completo
(2) Diversity of root nodule bacteria isolated from common bean (Phaseolus vulgaris L.) in Cuba: effect of Rhizobium – PGPR inoculation and IAA synthesis on development of bean genotypes. Ariany COLÁS SÁNCHEZ. Supervisors: Prof. Jos Vanderleyden, KU Leuven Prof. Jan Michiels, KU Leuven Dr. Roldán Torres Gutiérrez, UNL Members of the Examination Committee: Prof. Dirk De Waele, KU Leuven Dr. Maarten Fauvart, KU Leuven Dr. Yaquelín Nerey Orozco, UCLV Prof. Dirk Springael, KU Leuven. June 2014. Dissertation presented in partial fulfilment of the requirements for the degree of Doctor in Bioscience Engineering.
(3) Doctoraatsproefschrift nr. 1187 aan de faculteit Bio-ingenieurswetenschappen van de KU Leuven. © 2014 KU Leuven, Science, Engineering & Technology Uitgegeven in eigen beheer, Ariany Colás Sánchez, Cuba. Alle rechten voorbehouden. Niets uit deze uitgave mag worden vermenigvuldigd en/of openbaar gemaakt worden door middel van druk, fotokopie, microfilm, elektronisch of op welke andere wijze ook zonder voorafgaandelijke schriftelijke toestemming van de uitgever. All rights reserved. No part of the publication may be reproduced in any form by print, photoprint, microfilm, electronic or any other means without written permission from the publisher. ISBN 978-90-8826-363-7 D/2014/11.109/29.
(4) The research was supported by a doctoral research fellowship from IRO (Interfaculty Council for Development Co-operation of the KU Leuven).
(5)
(6) ACKNOWLEDGMENTS I would like to express my sincere and deepest gratitude to my supervisors, Professors Jos Vanderleyden and Jan Michiels, thank you very much for providing wonderful guidance, useful discussions and comments. Dear Jan certainly, without your help and support my dream would not have been possible. I extend my special gratitude to Dr. Roldán Torres Gutiérrez, who was part of the project from the beginning and one of the principal authors of this thesis. I would like to thank all members of the examination committee for their helpful suggestions and interesting comments to improve this document. I am grateful to Lara Ramaekers for facilitating the first stages in Belgium and giving me a warm welcome to this cold country. I also appreciate her, for supporting me in the search for grants and for all positive advices provided during the course of my experiments. To Steven, her husband, and their families, thank a lot, I hope that we can dance salsa in Cuba someday. My sincere appreciations go to Jos Desair, for providing productive and essential monitoring for HPLC analysis, thank you very much for being so patient. I also appreciate to Stijn Spapen for his support and appropriate advices in the conjugation tests. Many thanks go to my Cuban colleagues and friends: Amore (Cupull), Oralia, Joaquín, Cairo, Alianny, Bladimir, Sirley, Perico, Carrazana, Yordanys, Nayivis and Miki. To my thesis students Belky, Octavio and my college Esmildo, for providing professional and technical support during these years. Pero más que todo, muchas gracias, por hacerme reír y compartir mis penas y alegrías. Especiales son mis agradecimientos a la dirección de la Facultad de Ciencias Agropecuarias y el Departamento de Agronomía, por todo su apoyo institucional y personal. Thank you very much, to the doctoral research fellowship from IRO (Interfaculty Council for Development Co-operation of the KU Leuven) for funding all these years of study. Thank you very much to all my colleagues at CMPG and SPI group. Sandra, Maarten Fauvart, Maarten Ghequire and Hanne (I really enjoyed sharing the office with you). Maarten Ghequire (your comments and help were very useful, thank you very much for being there every time), Maarten Fauvart (thank you very much for your help). Cynthia (mujer, como voy a extrañar nuestros almuerzos, pero volveré a la boda ). To André from Olivia (my Belgium name) thank you very much for being so friendly. Jannick, Shanjun and Natalie, thanks a lot for hearing my presentations and helping me every time. Annita your help and organization were amazing. Casi en el final de esta página, pero siempre en el lugar más importante en mi vida, muchas gracias a mis padres Caridad Sánchez Moré y Rafael Colás Lazcano, quienes han dado todo para que cada día sea una mejor persona, mi hermano Adrián Colás Sánchez, mis tías Epe y Soty, mis primos Erick, Esima, Onel gracias por todo su apoyo y más que todo, GRACIAS POR EXISTIR!!!!!!.
(7) Tito el taxista, cuantas largas travesías desde Santa Clara a la Habana!!!!! No me queda más que agradecerte y espero que hagamos muchas más a lo largo de la vida. A mis fallecidos y siempre queridos tíos, Antolín Sánchez Moré y Dulce María Manzano Tápanes. Confío en que desde donde estén me estarán mirando con la satisfacción y el orgullo, de que todo el sacrificio que hicieron por mí, hoy se materializa, MUCHAS GRACIAS!!!!!!! Verónica y Giraldo, mi más caluroso y sincero agradecimiento, por hacerme sentir como en familia cada momento que disfruté con ustedes. Querida Margarita, muchas gracias por abrir las puertas de su corazón y su casa para mí. I would like to thank to my fiancé, Ahmed Chacón Iznaga (Gordis), it was because of your love and patience that made this possible. I thank you for all you did for me and for always being here. All this work is dedicated to you!!!!!! To all of you, thank you very much, Muchas gracias, Ariany.
(8) Summary. Summary. Common bean (Phaseolus vulgaris L.) is the most important food legume for the direct human consumption in the world and particularly in many parts of Latin America and Africa. Symbiotic nitrogen fixation (SNF) through association with root-nodulating rhizobia effectively contributes to an improved crop production and is an ecologically sound and low cost strategy to improve productivity in common bean cultivation. However, the amount of nitrogen fix by common bean in the field is often low compared to the SNF capacity of beans under optimal conditions and compared to other legumes. The success of SNF in bean cultivation is strongly influenced by prevailing environmental conditions. Further research to improve the efficiency and stability of SNF in bean cultivation is needed. In the rhizosphere and in their interaction with plants, rhizobia encounter many different plant compounds, including phytohormones like auxins, giberelinis and cytokinins. Several steps of rhizobia-legume interaction are monitored by these phytohormones, especially nodule formation. However, the exact role of bacterial IAA (indole-3-acetic acid) in nodule formation and functioning is still unknown. This study focuses on the characterization of the nodule inhabiting organisms by the application of molecular techniques and to investigate the agronomic implication of the legume-Rhizobium-Plant growth promoting rhizobacteria (PGPR), as well as to determine the possible role of bacterial IAA production in modulation of nodulation. In total, 20 gram negative strains were isolated from root nodules of common bean in the central region of Cuba. By partial sequencing of the 16S rRNA gene, six different genera,. i.e.. Arthrobacter,. Chryseobacterium,. Enterobacter,. Stenotrophomonas,. Pseudomonas and Rhizobium were recovered. Rhizobium was the only genus containing differences at the species level. In total four Rhizobium species were identified (Rhizobium sp., R. etli, R. pisi and R. radiobacter). Of which, only R. pisi and R. etli species nodulated the host plant. The phenotypic characterization showed the beneficial effect of the R. etli and R. pisi on bean genotypes, by increasing the nodulation, growth parameters and N fixation at early stage of common bean plants. Under field trial conditions, the nodulation, growth parameters and yield were stimulated significantly for BAT-477 as compared with the genotypes BAT-304, ICA Pijao and DOR-364, upon i.
(9) Summary. inoculation with the isolated Rhizobium strains. A survey among 60 local bean farmers showed that farmers use microbial inocula as well as other organic fertilizers when they have access to them. Resistance to pests, good taste which was always associated to black color and resistance to diseases, were included as preferential characteristic for choosing a new variety. Combinations of Rhizobium-PGPR were evaluated under different growth conditions, using two contrasting genotypes (BAT-477 and DOR-364). The results highlight the interaction between genotype, bacterial inoculant and environmental soil conditions. It was observed that native Rhizobium-Pseudomonas co-inoculation as compared to single Rhizobium inoculation increased the nodulation and growth parameters in greenhouse experiments as well as in a field trials. The combination Rhizobium-Pseudomonas showed the best results in yield in both genotypes and BAT-477 performed significantly better than DOR-364. In addition, all nodule endophytic bacterial strains showed plant growth promoting activities, the production of indole-3-acetic acid (IAA) resulted common to all of them, the synthesis of siderophores and phosphate solubilization was only produced for Pseudomonas strains. These results indicate a potential role of isolated bacterial in improving plant growth. Several lines of evidence suggest a role for phytohormones in root nodule development. In our study R. pisi R40983 (ipdC) transconjugant showed major nodule stimulation in common bean plants as well as major IAA production principally during the stationary growth phase. Based on the nodulation experiment, it was demonstrated that the introduction of these recombinant ipdC construct could improve nodulation. These findings contribute to the understanding of the interplay between Rhizobium, PGPR and the plant host under different soil conditions. Importantly, co-inoculation with R. pisi and P. monteilii could be an effective biofertilization strategy for common bean production. From this study, some important recommendations can be drawn on how to improve bean productivity in this region of Cuba.. ii.
(10) Samenvatting. Samenvatting. De boon (Phaseolus vulgaris L.) zijn het belangrijkste vlinderbloemige gewas voor menselijke voeding op wereldschaal en in het bijzonder in vele delen van Latijns Amerika en Afrika. Symbiotische stokstoffixatie (SSF) door de associatie van rhizobia met wortel nodulen draagt bij tot een verbeterde gewasproductie en is een goedkope en ecologische strategie om de opbrengst van boon cultivatie te verhogen. De hoeveelheid stikstoffixatie door de gewone bonenplant is echter zeer laag in vergelijking met de SFF onder optimale condities en vergeleken met andere peulvruchten. Het succes van SSF in de cultivatie van bonen wordt eveneens zeer sterk beïnvloed door omgevingscondities. Verder onderzoek naar manieren om de efficiëntie en de stabiliteit van SSF te verhogen is echter nog steeds noodzakelijk. Rhizobia treffen tijdens hun interactie met planten verschillende componenten aan, waaronder fytohormonen zoals auxinen , giberelinen en cytokininen. Verschillende stappen, waaronder de vorming van nodulen, in de interactie van rhizobia met vlinderbloemigen worden gereguleerd door deze fytohormonen. De precieze rol van bacterieel indol-3-azijnzuur in nodulevorming en werking is echter nog onbekend. Deze studie richt zich op de karakterisatie van de bezetting van nodulen door deze organismen en op de agronomische implicatie van de plant-rhizobium interacties. Hiernaast wordt ook de mogelijke rol van indol-3-azijnzuur in de modulatie van nodulen bestudeerd. In totaal werden 20 gram-negatieve stammen geïsoleerd van wortelnodules van de gewone boonplant in de centrale regio van Cuba. Door partiële sequenering van het 16S rRNA. gen. werden. zes. verschillende. genera. geïdentificeerd:. Arthrobacter,. Chryseobacterium, Enterobacter, Stenotrophomonas, Pseudomonas en Rhizobium. Alleen voor Rhizobium werden verschillen op species-niveau geobserveerd. In totaal werden vier Rhizobium species geïdentificeerd (Rhizobium sp., R. etli, R. pisi en R. radiobacter), waarvan alleen R. pisi en R. etli de gastheerplant konden noduleren. Fenotypische karakterisering toonde een stimulerend effect aan van R. etli en R. pisi op boongenotypes door een toename van nodulatie, groei en stikstoffixatie in een vroeg groeistadium. Onder veldcondities waren nodulatie, groei en opbrengst significant beter voor BAT-477 in vergelijking met genotypes BAT-304, ICA Pijao en DOR-364 na inoculatie met de iii.
(11) Samenvatting. geïsoleerde Rhizobium stammen. Een bevraging van 60 lokale boontelers wees uit dat ze zowel microbiële inocula als andere organische bemesting gebruiken als die beschikbaar zijn. Resistentie tegen plagen en een goede smaak, altijd gekoppeld aan zwarte kleur en ziekteresistentie, kwamen naar voren als voorkeurseigenschappen voor een nieuwe variëteit. Combinaties. van. Rhizobium-PGPR. werden. geëvalueerd. onder. verschillende. groeicondities waarbij voor Rhizobium gebruik werd gemaakt van 2 contrasterende genotypes (BAT-477 en DOR-364). De resultaten duiden op interactie tussen genotype, bacterieel inoculum en bodemcondities. Er werd geobserveerd dat inoculatie van de combinatie Rhizobium-Pseudomonas de nodulatie en groeiparameters verbeterden ten opzichte van enkel Rhizobium inoculatie, zowel in de serre als op het veld. De beste resultaten voor opbrengst werd bekomen wanneer Pseudomonas werd gecombineerd met één van beide Rhizobium genotypes. Echter, de combinatie met BAT-477 leidt tot significant betere resultaten in vergelijking met DOR-364. Bijkomend vertoonden alle nodule-geassocieerde. endophytische. bacteriën. plantgroeibevorderende. effecten. waarbij indole-3-acetic acid (IAA) werd geproduceerd door alle bacteriële stammen, sideroforen en componenten voor fosfaatsolubilisatie daarentegen werd enkel aangemaakt door Pseudomonas stammen. Deze. bevindingen. tonen. aan. dat. de. geïsoleerde. bacteriën. een. mogelijke. plantgroeibevorderende eigenschap hebben. Verschillende aanwijzingen werden gevonden voor een rol van fytohormonen in de vorming van wortelnodules. Vii een R. pisi R40983 (ipdC) transconjugant vertoonde in onze experimenten een verhoogde wortelnodulevorming in de gewone bonenplant evenals een verhoogde IAA productie gedurende stationaire fase. Nodulatie-experimenten toonden aan dat het recombinante ipdC construct nodulevorming verhoogde. Deze resultaten laten toe de samenhang van Rhizobium, PGPR en de gastheerplant onder verschillende bodemcondities beter te begrijpen. Belangrijk hierbij is dat co-inoculatie met R. pisi en P. monteilii een mogelijks effectieve bio-bemesting kan zijn voor het telen van de gewone bonenplant. Enkele belangrijke aanbevelingen uit dit onderzoek kunnen dan ook de productiviteit van de bonenteelt in deze Cubaanse regio verbeteren.. iv.
(12) Resumen. Resumen. El frijol común (Phaseolus vulgaris L.) es la leguminosa alimenticia más importante para el consumo humano directo en el mundo y particularmente en muchas partes de América Latina y África. La fijación simbiótica de nitrógeno (FSN) a través de la asociación con bacterias colonizadoras de la raíz “rhizobia”, contribuye eficazmente a una nutrición mejorada y es una estrategia ecológica y de bajo costo para mejorar la productividad en el cultivo de frijol común. Sin embargo, la cantidad de nitrógeno fijado por frijol común en condiciones de campo, es a menudo baja, en comparación a la fijada en condiciones óptimas comparado con otras leguminosas. El éxito de FSN en el cultivo de frijol, está fuertemente influenciada por las condiciones ambientales existentes. Por lo tanto, es necesario llevar a cabo investigaciones con vistas a mejorar la eficiencia y la estabilidad de FSN en el cultivo de frijol. En la rizosfera y su interacción con las plantas, rizobios interactúan con diferentes compuestos de las plantas, incluidas fitohormonas como las auxinas. Varios pasos de la interacción rizobias- leguminosas son monitoreados por estas fitohormonas, especialmente la formación de nódulos. Sin embargo, la función exacta del IAA (ácido indol-3-acético) en la formación y el funcionamiento de los nódulos es todavía desconocida. Este estudio se centra en caracterizar los microorganismo que habitan los nódulos de frijol común, mediante la aplicación de técnicas moleculares e investigar la implicación agronómica de la interacción leguminosa Rhizobium - PGPR. Así como determinar el posible papel de la producción de AIA por las bacterias en la modulación de la nodulación. En total, 20 bacterias gram negativas fueron aisladas de nódulos de las raíces del frijol común en la región central de Cuba. Por la secuenciación parcial de la región 16S rRNA, seis. géneros. diferentes:. Arthrobacter,. Chryseobacterium,. Enterobacter,. Stenotrophomonas, Pseudomonas y Rhizobium fueron recuperados. Rhizobium fue el único género que mostró diferencias a nivel de especies. En total fueron identificadas cuatro especies de Rhizobium (Rhizobium sp., R. etli, R. radiobacter y R. pisi). De las cuales, solo las especies R. pisi y R. etli nodularon la planta hospedera. La caracterización fenotípica mostró el efecto beneficioso de las especies R. etli y R. pisi en genotipos de frijol, mediante el aumento de los parámetros de nodulación y de crecimiento, así como v.
(13) Resumen. la fijación de N en la fase temprana de crecimiento de las plantas de frijol común. Bajo las condiciones de campo, la nodulación, los parámetros de crecimiento y rendimiento fueron estimulados significativamente en el genotipo BAT-477 en comparación con BAT- 304, ICA Pijao y DOR-364 después de la inoculación de las cepas de Rhizobium aisladas. La encuesta realizada a 60 agricultores locales de frijol, demostró que los agricultores utilizan inóculos microbianos, así como otras prácticas orgánicas solo cuando tienen acceso a las mismas. La resistencia a las plagas, el buen gusto que siempre se asocia con el color negro y la resistencia a las enfermedades, se incluyeron como preferencias para las características de una nueva variedad. Combinaciones de Rhizobium - PGPR fueron evaluadas bajo diferentes condiciones de crecimiento, utilizando dos genotipos contrastantes (BAT-477 y DOR-364). Los resultados ponen de manifiesto la interacción entre el genotipo, inóculo bacteriano y las condiciones de suelo. Se observó que la coinoculación de cepas nativas Rhizobium Pseudomonas aumentó los parámetros de nodulación y crecimiento, en comparación con la inoculación sencilla de Rhizobium, bajo condiciones de invernadero así como en el ensayo de campo. La combinación de Rhizobium - Pseudomonas mostró los mejores resultados en el rendimiento en los dos genotipos y el genotipo BAT-477 se desarrolló significativamente mejor que DOR-364. Además, todas las bacterianas endófitos mostraron actividades de promoción de crecimiento de las plantas, la producción de ácido indol-3-acético (IAA) fue común para todas. Por otra parte, la síntesis de sideróforos y solubilización de fosfato fue producida solo por las bacterias de Pseudomonas. Resultados que indican el posible papel de las bacterias aisladas en el fomento del crecimiento de las plantas. Varios estudios sugieren el papel de fitohormonas en el desarrollo de nódulos de la raíz. En nuestro estudio, R. pisi R40983 portadora de el gen ipdC mostró mayor estimulación del número de nódulos en plantas de frijol, así como mayor producción de IAA sobre todo durante la fase de crecimiento estacionario de la bacteria. Basado en el experimento nodulación fue demostrado que la introducción de ipdC podría mejorar la nodulación. Estos resultados contribuyen a la comprensión de la interacción entre Rhizobium, PGPR y la planta hospedera bajo diferentes condiciones de suelo. Es importante destacar que la co- inoculación con R. pisi y P. monteilii podría ser una estrategia de biofertilización eficaz para la producción de vi.
(14) Resumen. frijol común. A partir de este estudio algunas recomendaciones importantes se pueden extraer sobre la forma de mejorar la productividad del frijol en esta región de Cuba.. vii.
(15) List of abbreviations Al AMF ANOVA ARA BNF Br C Ca CAS CCS cfu CIAT CMPG CPA DAP DNA EMBL EPS GPP HPLC HR IAA ipdC Km LB Mn N N2 NDW NF(s) NFW NGP NH3 NN OM P PCR PDWp. Aluminum Arbuscular mycorrhizal fungi Analysis of variances Acetylene Reduction Assay/Acetylene Reduction Activity Biological nitrogen fixation Boron Carbon Calcium Chrome Azurol-S Cooperativa de Créditos y Servicios Colony forming units International Center for Tropical Agriculture Center of Microbial and Plant Genetic Cooperativas de Producción Agropecuaria Days after planting Deoxyribonucleic acid European molecular biology laboratory Exopolysaccharides Grains per plant High performance liquid chromatography Relative humidity Indole-3-acetic-acid Indole-3-pyruvate decarboxylase Kanamycin Luria–Bertani Manganese Nitrogen Di-nitrogen Nodule dry weight Nod factor(s) Nodule fresh weight Number of grain per plant Ammonia Nodule number Organic matter Phosphorus Polymerase chain reaction Plant dry weight viii.
(16) PGP PGPR PPP PVK PWP RDW RFW rRNA SDW SFW SNF SPAD SPAD ChlV SPE SPSS Tc Trp TSB UBPC UCLV YEM YMA. Plant growth promoting Plant-growth promoting rhizobacteria Pods per plant Pikovskaya’s agar medium Pod weight per plant Root dry weight Root fresh weight Ribosomal ribonucleic acid Shoot dry weight Shoot fresh weight Symbiotic nitrogen fixation Soil-Plant Analyses Development SPAD chlorophyll value Solid phase extraction Statistical Package for the Social Sciences Tetracycline Tryptophan Tryptic Soy Broth Unidad Básica de Producción Cooperativa Central University of Las Villas Yeast extract mannitol Yeast mannitol agar. ix.
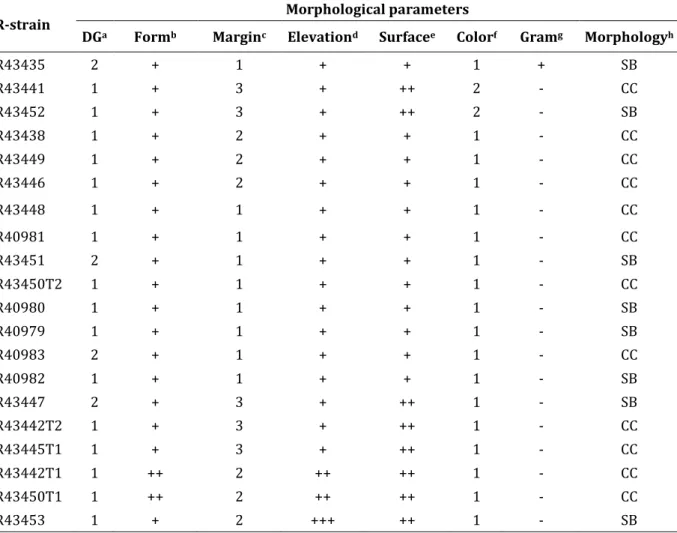
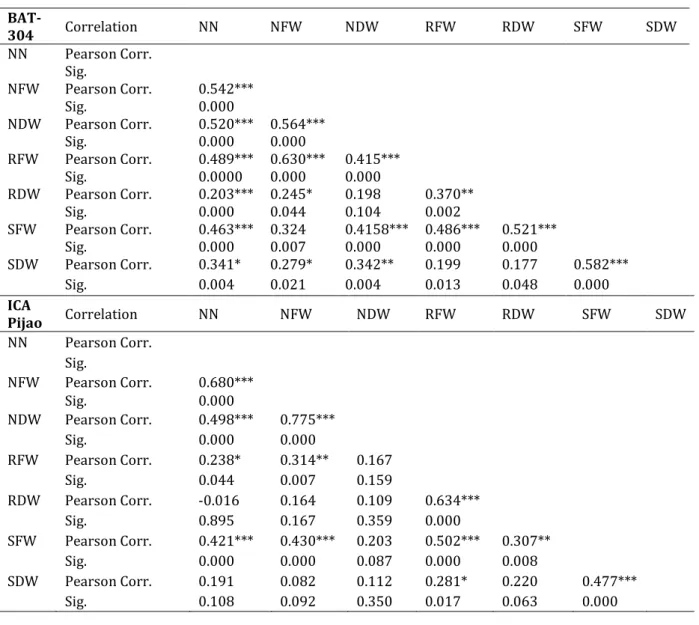
(17) Table of contents Summary ................................................................................................................................................................................. i Samenvatting ...................................................................................................................................................................... iii Resumen ................................................................................................................................................................................. v List of abbreviations ...................................................................................................................................................... viii Table of contents ................................................................................................................................................................. x Introduction ................................................................................................................................................................................................1 Positioning of the subject, objectives and thesis outline ...................................................................................................1 Chapter 1 ................................................................................................................................................................................ 7 Legumes - Rhizobium interaction: Stress factors in the soil nvironment that impact on nodulation. 7 1.1 Rhizobium-legume interaction ....................................................................................................................................................8 1.2 Rhizobia in the soil environment ...............................................................................................................................................9 1.2.1 Abiotic soil stress factors ................................................................................................................................................... 10 1.2.1.1 Soil acidity (pH) ............................................................................................................................................................ 10 1.2.1.2 Temperature .................................................................................................................................................................. 12 1.2.1.3 Salinity .............................................................................................................................................................................. 13 1.2.1.4 Soil nutrients (N and P) ............................................................................................................................................. 14 1.2.1.5 Drought stress (soil water status) ........................................................................................................................ 16 1.2.2 Biotic factors that impact on nodulation .................................................................................................................... 18 1.2.2.1 Mycorrhizal associations .......................................................................................................................................... 18 1.2.2.2 Rhizobium interaction with other soil microorganisms ............................................................................. 19 1.3 Role of auxin and cytokinin in nodule development ...................................................................................................... 20 1.4 Plant Growth Promoting Rhizobacteria (PGPR) ............................................................................................................... 22 1.5 Conclusion ......................................................................................................................................................................................... 24 Chapter 2 ............................................................................................................................................................................. 25 Morphological and genetic identification of rhizobia strains from Cuba and their behavior on different common bean (Phaseolus vulgaris L.) genotypes.............................................................................. 25 2.1 Introduction ...................................................................................................................................................................................... 26 2.2 Materials and methods ................................................................................................................................................................ 27 2.2.1 Sampling and sample preparation................................................................................................................................. 27 2.2.2 Isolation of bacteria ............................................................................................................................................................. 29 2.2.3 Morphological characterization...................................................................................................................................... 29 2.2.4 DNA extraction ....................................................................................................................................................................... 30 2.2.5 PCR amplification of the 16S rRNA gene .................................................................................................................... 30 2.2.6 Plant inoculation tests ........................................................................................................................................................ 31 2.2.6.1 Bacterial strains and inoculum preparation .................................................................................................... 31 2.2.6.2 Plant material and analysis of symbiotic traits in a climate controlled plant growth room ...... 31 2.2.6.3 Nitrogenase activity and nodulation test .......................................................................................................... 32 2.2.7 Greenhouse experiment ..................................................................................................................................................... 32 2.2.7.1 Bacterial strain and inoculum preparation ...................................................................................................... 32 2.2.7.2 Plant Material................................................................................................................................................................. 33 2.2.7.3 Measurement of plant parameters ....................................................................................................................... 33 2.2.7.4 Analysis of total nitrogen content in plant and biomass ............................................................................ 34 2.2.8 Statistical analysis................................................................................................................................................................. 34 2.3 Results ................................................................................................................................................................................................. 35 2.3.1 Morphological characterization...................................................................................................................................... 35 2.3.2 Genetic identification and diversity of bacteria associated with common bean nodules in Cuban soils ......................................................................................................................................................................................................... 36 2.3.3 Characterization of Rhizobium strains during symbiosis with bean genotypes (controlled conditions) .......................................................................................................................................................................................... 38 2.3.3.1 Nodulation test under controlled growth conditions .................................................................................. 38 x.
(18) 2.3.3.2 Effect of inoculation on local bean genotypes under control greenhouse condition in Cuba .... 40 2.3.3.3 Effect of inoculation on nitrogen content .......................................................................................................... 43 2.3.3.4 Correlation between SNF capacity and related growth traits in BAT-304 and ICA Pijao genotypes ....................................................................................................................................................................................... 44 2.4 Discussion .......................................................................................................................................................................................... 45 Chapter 3 ............................................................................................................................................................................. 49 Rhizobium inoculation effects on growth and yield of four common bean (Phaseolus vulgaris L.) genotypes grown under Cuban field conditions ................................................................................................... 49 3.1 Introduction ...................................................................................................................................................................................... 50 3.2 Materials and methods ................................................................................................................................................................ 51 3.2.1 Plant material, selected trial sites and experiment desing ................................................................................. 51 3.2.2 Bacterial strains and inoculum preparation ............................................................................................................. 52 3.2.3 Measurement of plant parameters ................................................................................................................................ 53 3.2.4 Survey ......................................................................................................................................................................................... 53 3.2.5 Statistical analysis................................................................................................................................................................. 54 3.3 Results ................................................................................................................................................................................................. 54 3.3.1 Symbiotic effectiveness of selected isolates under Cuban field conditions ................................................ 54 3.3.1.1 Effect of inoculation on nodulation and plant growth parameters ....................................................... 54 3.3.1.2 Evaluation of yield ....................................................................................................................................................... 58 3.3.4 A survey of bean farmers’ preference in bean varieties and ecological practices ................................... 60 3.4 Discussion .......................................................................................................................................................................................... 63 Chapter 4 ............................................................................................................................................................................. 67 Stimulatory effect of Pseudomonas monteilii on the Rhizobium-bean interaction under greenhouse conditions ........................................................................................................................................................................... 67 4.1 Introduction ...................................................................................................................................................................................... 68 4.2 Materials and methods ................................................................................................................................................................ 69 4.2.1 Bacterial strain and growth condition ......................................................................................................................... 69 4.2.2 Inoculum preparation ......................................................................................................................................................... 69 4.2.3 Plant material, growth condition and inoculation.................................................................................................. 69 4.2.4 Growth analysis and symbiotic traits ........................................................................................................................... 70 4.2.5 Statistical analysis................................................................................................................................................................. 70 4.3 Results ................................................................................................................................................................................................. 71 4.3.1 Effects of PGPR on nodulation and plant growth parameters of bean under greenhouse conditions .................................................................................................................................................................................................................. 71 4.3.2 Nitrogen fixation evaluation by measuring chlorophyll values ....................................................................... 73 4.3.3 Correlation between all evaluated parameters during the growth cycle .................................................... 74 4.4 Discussion .......................................................................................................................................................................................... 75 Chapter 5 ............................................................................................................................................................................. 79 Effects of co-inoculation of native Rhizobium and Pseudomonas strains on growth parameters and yield of two contrasting Phaseolus vulgaris L. genotypes under Cuban field conditions ..................... 79 5.1 Introduction ...................................................................................................................................................................................... 80 5.2 Materials and methods ................................................................................................................................................................ 81 5.2.1 Selected trial sites and plant material ......................................................................................................................... 81 5.2.2 Soil and climate characteristics of sites ...................................................................................................................... 83 5.2.3 Bacterial strains, growth condition and inoculum preparation in the trials .............................................. 83 5.2.4 Experimental design ............................................................................................................................................................ 84 5.2.5 Measurement of plant parameters ................................................................................................................................ 85 5.2.6 Statistical analysis................................................................................................................................................................. 85 5.3 Results ................................................................................................................................................................................................. 86 5.3.1 Effect of Rhizobium – Pseudomonas co-inoculation on nodulation and plant growth parameters .. 86 5.3.2 Effect of Rhizobium – Pseudomonas co-inoculation on yield and yield parameters ................................ 90 5.3.3 Correlations between yield and parameters measured during growth ....................................................... 92 5.4 Discussion .......................................................................................................................................................................................... 93 Chapter 6 ............................................................................................................................................................................. 97 Indole acetic acid production and other beneficial properties of Rhizobium and plant growthpromoting rhizobacteria: Role of ipdC in auxin synthesis and modulation of nodules ......................... 97 6.1 Introduction ...................................................................................................................................................................................... 98 xi.
(19) 6.2 Materials and methods ............................................................................................................................................................... 99 6.2.1 Bacterial strains and growth conditions ..................................................................................................................... 99 6.2.2 Determination of IAA production .................................................................................................................................. 99 6.2.2.1 Purification and analysis of purified IAA ........................................................................................................ 100 6.2.3 Phosphate solubilization ................................................................................................................................................ 101 6.2.4 Siderophore production .................................................................................................................................................. 102 6.2.5 Construction of R. pisi strains altered in IAA biosynthesis .............................................................................. 102 6.2.5.1 Bacterial strains and media .................................................................................................................................. 102 6.2.5.2 Bacterial matings ...................................................................................................................................................... 102 6.2.5.3 Analysis of IAA production by wild-type and genetically modified R. pisi strains ....................... 103 6.2.5.4 Seed inoculation and growth conditions ........................................................................................................ 103 6.2.6 Statistical analysis.............................................................................................................................................................. 104 6.3 Results .............................................................................................................................................................................................. 104 6.3.1 Indole-3-acetic production by PGPR associated to bean plants .................................................................... 104 6.3.2 Screening of phosphate-solubilizing and CAS bacteria ..................................................................................... 107 6.3.3 Characterization of ipdC genetically modified R. pisi ......................................................................................... 109 6.3.3.1 Differential nodulation response of R. pisi (pFAJ5002) and parental strains ................................ 110 6.4 Discussion ....................................................................................................................................................................................... 111 Chapter 7 ...........................................................................................................................................................................115 Major conclusions and perspectives .......................................................................................................................115 References ........................................................................................................................................................................121 Appendices .......................................................................................................................................................................138 List of Publications ........................................................................................................................................................143. xii.
(20)
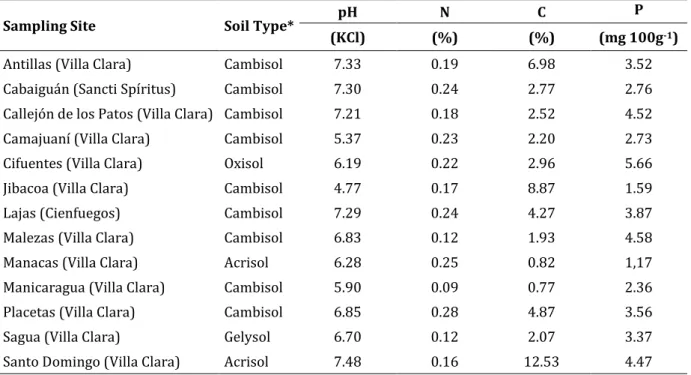
(21) Introduction. Introduction. Positioning of the subject, objectives and thesis outline Common bean (Phaseolus vulgaris L.) is economically the most important member of the genus Phaseolus and is cultivated in all parts of the world (Broughton et al. 2003; Drevon et al. 2001; Martínez-Romero 2003). It is particularly significant for Latin America. Cultivated common beans are very important sources of protein, vitamins and minerals for over 500 million people in Latin America and Africa (Broughton et al. 2003). The high content of protein, minerals and antioxidant compounds (Xu and Chang 2008) makes P. vulgaris a key crop for the food security and health in the developing countries and an excellent model food legume (Broughton et al. 2003). In Africa and Latin America, common bean production is often undertaken on small farms using marginal soils low in nitrogen (N) and phosphorus (P) content and with minimal technical inputs (Reviewed by Galeano 2012). Central America and parts of the Caribbean have high levels of production or consumption compared to their respective population size (Table 1). Especially in Cuba, dry beans are the core of the daily diet, either alone or in combination with rice. Consequently, beans are widely cultivated all over the country, with approximately 123 434 ha harvested in 2012 and a yield of 1.03 t ha-1 (ONE 2013). Table 1. Bean production in the Caribbean (taken from Beaver et al. 2012). Country Cuba Dominican Republic Haiti Jamaica Puerto Rico Caribbean. Area of production (ha). Production (Mt). Seed yield (kg ha-1). 110000. 130000. 1182. 40402. 32725. 810. 53000. 33000. 623. 220. 230. 1.045. 400. 1050. 2.500 (green shelled). 202906. 189941 1. Preferred seed types Black, red mottled and white Red mottled, black, cranberry, white Black, red mottled, yellow, white, pinto Pink striped, light red kidney White, pink, pinto, light red kidney.
(22) Introduction. P. vulgaris, like all the legumes plants, is capable to perform the process of symbiotic nitrogen fixation (SNF) in association with rhizobacteria collectively called rhizobia, grouping the genera Rhizobium, Sinorhizobium, Mesorhizobium, Bradyrhizobium, Allorhizobium and Azorhizobium (Weidner et al. 2003). This process between common bean and nitrogen-fixing bacteria is crucial in agricultural crop production. Major benefits of the legume-rhizobia symbiotic interaction are the reduction of nitrogen fertilizer requirements and the improvement of plant growth and health (Giller 2001). Symbiotic systems such as that of legumes and rhizobia can be a major source of N in most cropping systems with an average of 80% of the required nitrogen coming from biological N2 fixation (Graham and Vance 2000). However, the common bean is considered a poor nitrogen-fixer in comparison to other grain legumes, having values around 40% of the N2 derived of the association. This legume can fix more than 100 kg ha-1 of N in temperate regions or highland tropical environments, but the process of (SNF) in warmer tropical climates never reached this level (Hardarson et al. 1993). Understanding and improving SNF in common bean will be an economical and sustainable support for poor farmers. The formation of nodules on legume plant roots and specifically in common beans is one of the most important factors determining N2 fixation and consequently increase of growth and yield (Hungria et al. 2001). This complex process is governed by several intrinsic factors, physiologic characteristics of the macro (legume) and micro (rhizobia) symbionts (Michiels et al. 1998), and importantly by specific hormones, including auxins (indole-3-acetic acid, IAA), which are produced by both organisms during the interaction (Hirsch 1992; Remans et al. 2007; Spaepen et al. 2007). Root nodules have superior phytohormone content as compared with uninfected root tissues (Hirsch and Fang 1994). It was demonstrated that in two steps of nodules initiation (cell division and the establishment of the primordium) are required high auxin levels (deBilly et al. 2001). Some studies indicate that changes in auxin balance are essential for nodule initiation (Boot et al. 1999; Mathesius et al. 1998). Early Hirsh et al. (1989) demonstrated that incubation with synthetic auxin transport inhibitors stimulates the formation of nodule. 2.
(23) Introduction. Moreover, Mandal et al. (2009) showed that phenolic acids present in the nodule may play the role of stimulator for IAA production by the rhizobia. In addition, the sources of auxin in root nodules has not been clearly determined, Fedorova et al. (2005) attributed the high levels of IAA in young nodules to the polar transport. Strains collected from different soil types may offer the chance to select highly effective rhizobia for inoculants which will be an important tool for improving bean productivity in low input system in the central region of Cuba. HYPOTHESIS The identification and co-inoculation of native Rhizobium and plant growth promoting rhizobacteria (PGPR) auxin producer strains, can increase the nodulation, growth, N2 fixation and yield of common bean (Phaseolus vulgaris L.). GENERAL OBJECTIVE To study the unknown culturable endophytic bacterial diversity and plant growth promoting abilities of strains isolated from nodules of common bean plants, as well as to assess their effect on nodulation, growth parameters, N2 fixation and yield of common bean. SPECIFIC OBJECTIVES 1.. To isolate, identify and characterize bacterial strains from bean nodules in the central region of Cuba.. 2.. To determine the effect of isolated Rhizobium strains on plant growth and nitrogen content under different growth conditions.. 3.. To compare plant growth and symbiotic performance of selected bean genotypes under R. pisi-P. monteilii co-inoculation.. 4.. To investigate if co-inoculation of common bean seeds with combinations of specific rhizobia and plant growth-promoting Pseudomonas strains improves nodulation, plant growth and yield in different soil conditions in the Villa Clara province.. 5.. To evaluate the possibility of growth promotion and nodulation improvement in common bean through production of plant growth regulators by bacteria. 3.
(24) Introduction. 6.. To quantify and compare the IAA production of selected R. pisi and determine the role of ipdC plasmid on indole-acetic-acid production.. This thesis is presented in seven chapters describing, three large field experiments and two greenhouse experiments (Figure 1). A review of previously published literature, focusing on the nodulation process and the soil factors that impact nodulation is presented in Chapter 1. Isolation, identification and characterization of bacterial strains from bean nodules, nodulation studies under controlled conditions and the influence of Rhizobium inoculation and cultivar in greenhouse conditions are reported in Chapter 2. The influence of Rhizobium inoculation and genotype on growth, biomass accumulation of two local genotypes and yield under field conditions was studied in Chapter 3. Chapter 4 covers the effect of Rhizobium pisi – Pseudomonas monteilii inoculation on symbiotic traits and biomass stimulation of two contrasting bean genotypes. In Chapter 5 was studied, under field conditions the effect of co-inoculation of common bean seeds with combinations of specific rhizobia and plant growth-promoting Pseudomonas on nodulation, plant growth and yield in two different soil, in Cuba. Chapter 6 deals with the determination of IAA production by all nodulating Rhizobium and non-symbiotic strains and the possible role of ipdC overexpression on auxin levels Chapter 7 provides a general discussion and the conclusions from the work.. 4.
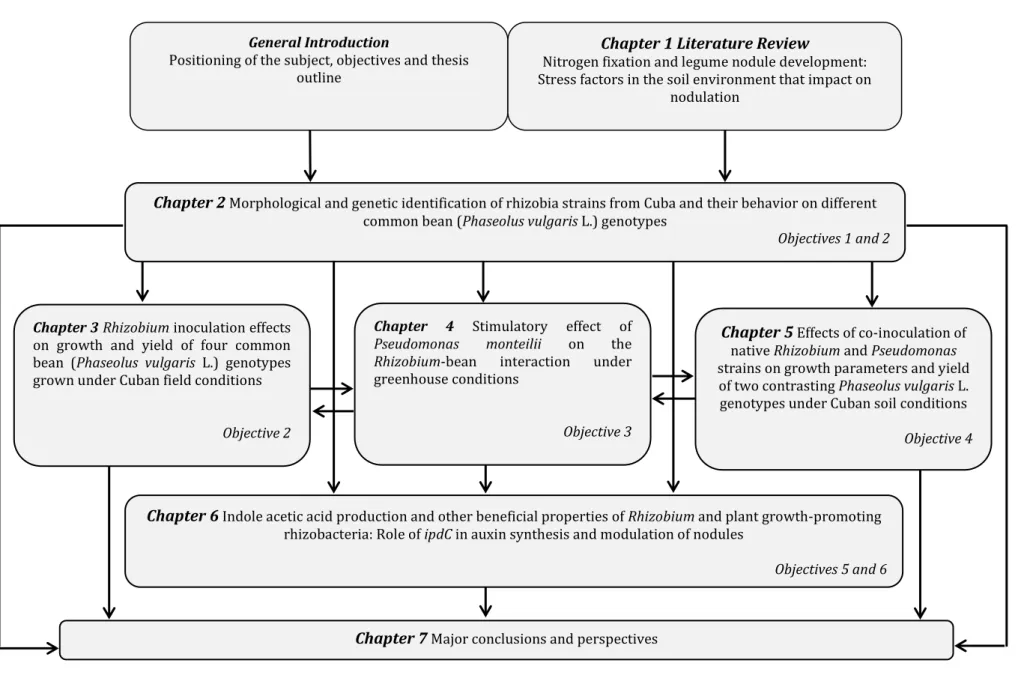
(25) Chapter 1 Literature Review. General Introduction Positioning of the subject, objectives and thesis outline. Nitrogen fixation and legume nodule development: Stress factors in the soil environment that impact on nodulation. Chapter 2 Morphological and genetic identification of rhizobia strains from Cuba and their behavior on different common bean (Phaseolus vulgaris L.) genotypes. Chapter 3 Rhizobium inoculation effects on growth and yield of four common bean (Phaseolus vulgaris L.) genotypes grown under Cuban field conditions. Chapter 4 Stimulatory effect of Pseudomonas monteilii on the Rhizobium-bean interaction under greenhouse conditions. Objective 2. Objective 3. Objectives 1 and 2. Chapter 5 Effects of co-inoculation of. native Rhizobium and Pseudomonas strains on growth parameters and yield of two contrasting Phaseolus vulgaris L. genotypes under Cuban soil conditions Objective 4. Chapter 6 Indole acetic acid production and other beneficial properties of Rhizobium and plant growth-promoting rhizobacteria: Role of ipdC in auxin synthesis and modulation of nodules. Objectives 5 and 6. Chapter 7 Major conclusions and perspectives Figure 1. Diagram representing thesis outline.. 1.
(26) 6.
(27) Chapter 1. Chapter 1 Legumes - Rhizobium interaction: Stress factors in the soil nvironment that impact on nodulation.. Abstract Legumes are a key food source and their capacity to fix nitrogen in symbiotic association with rhizobia contributes to improve crop yield and reduce negative impact of nitrogen fertilizers. However, the nitrogen fixing effectiveness of many legume–rhizobia combinations is poor. Variations in temperature, soil acidity and salinity, nutrient availability and humidity, among other factors, significantly impact the establisment and function of symbiosis between soil microorganisms and legumes, and consequently decreases their capacity to fix atmospheric nitrogen. In most cases, the microsymbiont is the more affected partner, with plants growing on mineral nitrogen usually less sensitive to these stresses. In this chapter, we discuss the stress factors that impact on rhizobia survival and their interaction with plants in the rhizosphere.. 7.
(28) Chapter 1. 1.1 Rhizobium-legume interaction Root nodule formation in legume plants is a complex process that results in nitrogen fixing symbiosis and requires a harmonized interaction between the host plant and the Rhizobium bacteria in the rhizosphere (Figure 1.1). Many rhizobia and host plants are extremely specific; however, some species of rhizobia interact with a high number of legumes (Pueppke and Broughton 1999). Nodulation takes place only when suitable species of legumes and Rhizobium come into contact (Wais et al. 2002). The excretion of specific compounds and induction of nod gene activity in the microsymbiont is the main role of the host plant in the process.. Figure 1.1. Schematic diagram of nodule development. Following the reception of flavonoid signals from root exudate the rhizobia attach to the surface of root hairs. Production of Nod factor by the rhizobia initiates a cascade of events (left to right), which at the morphological level include swelling, deformation and curling of the root hair, formation of the infection thread arising from the encapsulated bacterial cells, growth of the infection thread towards the nodule primordium with the release of the bacteria via infection droplets (Adapted from Kinkema et al. 2006).. The exchange of signals between both partners is the first step of nodule formation (Hayashi et al. 2014; Oldroyd 2011). The plant root releases root exudates (flavonoids and nutrients such as organic acids and amino acids) that attract rhizobia to particular sites of the legume roots and activate the Nod factor secretion by the bacteria (Russelle 2008). After Nod factor recognition by the plant, rhizobia attach to the root hair and 8.
(29) Chapter 1. promote its deformation, curling and cortical cell division in the root cortex (reviewed by Ferguson et al. 2010). The bacteria are then trapped in the curling root hair and enter the plant by forming an infection thread. Once inside, they multiply rapidly in the plant cells and are transformed into bacteroids, which become surrounded by plant cell membranes to form symbiosomes (reviewed by Hayashi et al. 2014). In this structure, rhizobia start the conversion of N2 to NH3 resulting in the nitrogen fixation process. 1.2 Rhizobia in the soil environment Prior to nodulation, rhizobia must function independently of the host plant as part of the global soil microbial population and as colonizers of the rhizosphere. Therefore, soil is a reservoir of Rhizobium strains and the intrinsic makeup of the soil can affect the outcome of competition. However, despite the numerous factors limiting their survival in the soil (Streeter 1994), these bacteria are common soil inhabitants in all climatic zones of the world (Andrade et al. 2002; Fagerli and Svenning 2005; Robertson et al. 1995). Rhizobium sp. survival in soil is influenced by a combination of factors, some of which are summarized in Table 1.3. Table 1.1 Factors influencing bacterial survival in soils (taken from van Veen et al. 1997). Origin. Factor. Effect. Biotic. Predation Competition Root growth. Population size decreases Population size decreases/antagonistic effect pathogens Release of organic compounds, enhancing survival. Clay minerals. Protection against predation. Water tension. High tension: water shortage, high osmolarity; low tension: anaerobism, increased nutrient availability by diffusion. Organic carbon. Selection for copiotrophic or oligotrophic species; limited organic carbon results in starvation and reduction in activity. Inorganic nutrients (N, P). Limitation results in starvation. pH. Selection for species; release of nutrients (e.g., P) or toxic compounds (e.g., Al3+).. Temperature. Metabolic activity as well as biotic (predatory) pressure affected.. Chemicals (toxic waste). Inhibition of sensitive organisms; selection of biodegradative, resistant, or tolerant forms. Abiotic. 9. on. plant.
(30) Chapter 1. Several abiotic soil factors play an important role in microorganism survival in the rhizosphere, e.g. textural type, pH, temperature, and humidity (Evans et al. 1993), nutrient availability and agricultural management regimes (Andrade et al. 2002; Laguerre et al. 2006; Palmer and Young 2000). In addition, several biotic factors can influence the number and diversity of indigenous rhizobia, particularly competition with other soil organisms, such as fungi and protozoa. In this chapter several biotic and abiotic soil factors that impact in nodulation are described. Understanding how rhizobial strains that inhabit the rhizosphere are affected by environmental conditions is a crucial target for improving nitrogen availability in sustainable agriculture, where legumes are a principal source of nitrogen. 1.2.1 Abiotic soil stress factors 1.2.1.1 Soil acidity (pH) The diversity and persistence of Rhizobium in the soil are significantly impacted by soil acidity (Andrade et al. 2002). In this context, acid soils have been associated with lower nodulation (Chemining’wa and Vessey, 2006). Several events in the symbiotic process are mostly sensitive to low pH levels (Duzan et al. 2004). Extreme soil acidity is usually linked with increased Al and Mn toxicity, which inhibit growth of the legume host, and reduced calcium (Ca) supply. These additional stresses impact the population density of rhizobia in the soil (Campo 1995). The microsymbiont is usually the more pH-sensitive partner (Hungria and Vargas, 2000). Acidity seems to exert a more severe influence on some root nodule bacteria, while others are more resistant. Within species, strains can vary hugely in their resistance to a low pH (Vlassak 1996; Brockwell et al. 1995; Hungria et al. 1997). Even when the optimum pH for Rhizobium growth is between 6.0 and 7.0 (Jordan 1984), some rhizobia strains are able to survive at pH ≤5.0 (Ademar et al. 2012). Some species have been reported to be extremely acid sensitive, among them Rhizobium tropici, Mesorhizobium loti, Bradyrhizobium sp. and Sinorhizobium meliloti (Brockwell et al. 1995). In this respect, Appunu et al. (2009) reported that Bradyrhizobia strains, isolated from indigenous cowpea plants, could withstand extreme acidic conditions, since they could survive and grow in the low pH, even at 3.5. Moreover, the minimum level of low 10.
(31) Chapter 1. pH for the majority of Rhizobium etli strains from Egypt was pH 4.7 (Shamseldin and Werner 2005). Acid tolerance reported in R. tropici type IIB, CIAT 899 (UMR 1899) from Colombia (Martínez et al. 1991; Graham et al. 1994; Shamseldin and Werner 2005) could explain the high frequency of this species in acid soils of Central America, Brazil, and Africa (Hungria and Vargas 2000). Bradyrhizobium japonicum could tolerate an acid environment more strongly than Sinorhizobium fredii. In extreme conditions of pH, the nodulation capacity of these species was principally influenced by their physiological characteristics (Yang et al. 2001). Declines in legume nodulation under acid conditions could be mitigated by supplementing. flavonoid. nod. gene. inducers. (Hungria. and. Stacey. 1997).. Supplementation of high levels of Nod factors had a positive effect on the overcoming low pH and temperature stress in B. japonicum (Duzan et al. 2004). Morón et al. (2005) showed that R. tropici (CIAT899) produced 52 different Nod factors (NFs) when exposed to acid pH compared with 29 at neutral pH. Among the different plant species, nodulated by different rhizobial species, the role of pH in signal interactions could differ (Indrasumunar et al. 2012). The underlying mechanisms that result in differences in acid tolerance in rhizobia strains have not been well defined (Correa and Barneix 1997). The capacity of acidtolerant rhizobia strains to conserve a consistent cytoplasm may explain this phenomenon (Chen et al. 1993a,b). The ability to grow in acidic conditions in Rhizobium sp. PR 389 involves intracellular and extracellular processes (Lei et al. 2011). Increased tolerance to low pH was observed in slow-growing bacteria as compared to fast growing (Graham et al. 1994). Only 37% of fast-growing rhizobial strains were able to grow in buffered medium at pH 4.0 and sixty percent grew at pH 9.5 (Hungria et al. 2001). Previously, it was determined that Rhizobium strains that produce greater amounts of exopolysaccharides (EPS) are more tolerant to acidic conditions when compared to strains that produce a lesser amount. Barberi et al. (2004) and Miguel and Moreira (2001) found similar results in strains of the genus Bradyrhizobium. They correlated the 11.
(32) Chapter 1. higher EPS produced by these bacteria with their better growth under acidic culture conditions. Efficient Rhizobium strains showed increased production of EPS at their optimal growing rates (Ademar et al. 2012). Differences among in vitro and acid soil tolerance by several Rhizobium strains have been observed. Higher competitiveness in both growth conditions at pH 5 was observed for several R. tropici strains (Ayango 1995). Recently, Indrasumunar et al. (2012) reported symbiotic effectiveness in acid soils for B. japonicum. Earlier, Asanuma and Ayanaba (1990) reported that Rhizobium strains isolated from acid soils did not grow on a nutrient medium with the same pH as the soil. Survival capacity of rhizobia was higher in soil than in nutrient medium at low pH (Appunu et al. 2009). Gemmell and Roughley (1993) observed higher tolerance to acidity in rhizobia isolated from acid soils as compared with those isolated from neutral soil. 1.2.1.2 Temperature Extreme soil temperatures and especially heat are known to affect rhizobial growth and survival and, consequently, the size of the rhizobial population. High soil temperatures in tropical regions are a main issue for biological nitrogen fixation by legume crops (Reviewed by Zhara 1999). In these soils, the upper limits for rhizobial survival are between 32-47 °C, although tolerance varies among species and strains (Hungria and Vargas 2000). The optimum growth temperature for most rhizobia is 25–30 °C (Zhang et al. 1995). However, during their life cycle, rhizobia are often exposed to higher temperatures. Effective nodulation in chickpea (Cicer arietinum L.) at low temperatures (9–15°C) was reported by Ogutcu et al. (2008). On the other hand, Hungria and Franco (1993) observed nodulation in common beans at high temperatures by Rhizobium leguminosarum bv. phaseoli, albeit with formation of ineffective nodules on roots and with poor N accumulation in shoot. Several studies have screened rhizobia for superior tolerance to high temperatures on culture medium and soils (Kulkarni and Nautiyal 1999; Marsh et al. 2006; Naeem et al. 2008). Some of them inferred that this behavior could be influenced by specific carbon sources in growth media Naeem et al. (2008). However, it should be noted that 12.
(33) Chapter 1. temperature tolerance of rhizobia may also depend on environmental conditions not easily reproduced in laboratory studies. Indeed, there are studies indicating that rhizobia endure high temperatures better in soil than in culture medium (Räsänen et al. 2001). Differences in the tolerance of high temperatures among species and strains of Rhizobium have been recognized, and the selection of strains for temperature tolerance has been suggested as a means of overcoming temperature stress (Surange et al. 1997). Some studies have confirmed that R. tropici endures higher temperatures than R. leguminosarum and R. etli (Hungria et al. 1993; Martínez-Romero et al. 1991; Mercante 1993; Sá et al. 1993). A 30°C/20°C temperature regime was optimum for Bradyrhizobium survival. Also, tolerance to elevated temperatures ≤ 35 °C was exhibited by B. japonicum species (Kluson 1986). However, R. leguminosarum bv. phaseoli was unable to fix N2 in symbiosis with bean at temperatures higher than 35°C (Hungria and Franco 1993). Some rhizobia isolated from environments with extreme temperatures in Africa showed a high percent of survival at 40°C (Eaglesham et al. 1991). Hungria and Stacey (1997) showed that a difference of 6 °C at 5 cm soil depth reduced the soybean Bradyrhizobium population by more the 10,000 cells g-1 of soil. At 39 °C, the excretion of nod gene inducers by soybean and common bean was decreased (Hungria and Vargas 2000). In conclusion, rhizosphere temperature impacts rhizobial survival in soil and exchange of molecular signals between host plants and rhizobia. 1.2.1.3 Salinity Agricultural production in several areas around the world is affected by soil salinity. Soils become infertile due to the high accumulation of salts in their horizons. Nearly 20% of the world’s cultivated land are affected by salinity, widely distributed throughout the world (Sumner 2000). Increases in the concentration of salts may negatively affect soil microbial populations, through direct toxicity and through osmotic stress (Tate 1995). Nodule formation decreases under salt stress conditions (Garg and Manchanda 2008).. 13.
(34) Chapter 1. Legume plants are better adapted to salinity than their rhizobial partners. Therefore an efficient Rhizobium–legume symbiosis under salt stress requires also the selection of salt-tolerant rhizobia (Zahran 1999). High salt concentration was reported to decrease nodulation and the amounts of N2 fixed (Bekki et al. 1987). Rhizobia show marked differences in salt tolerance. It is still unclear, however, whether those rhizobia able to grow under saline conditions are more efficient in nitrogen fixation than salt-sensitive rhizobia growing in saline environments (Tejera et al. 2004). Bradyrhizobium sp. RCM6 (R1), was highly efficient in improving growth and nutrition of Acacia cyanophylla under salinity stress (Hatimi 1999). Two R. etli strains isolated from Egyptian soils were highly tolerant to a salt concentration of up to 4% NaCl (Shamseldin and Werner 2005). In liquid culture, Rhizobium strains nodulating Phaseolus vulgaris, isolated from Morocco, were able to resist a sodium chloride concentration up to 4% NaCl (680 mM NaCl) (Priefer et al. 2001). In accordance with these results, Hashem et al. (1998) reported that several Rhizobium strains were tolerant to ˃3% NaCl and R. leguminosarum strains were shown to be tolerant to high NaCl concentrations in broth culture (Breedveld et al. 1991). Moreover, nodulation of R. leguminosarum 3841 was inhibited by salinity. This suppression was alleviated by adding high Ca2+ levels, which resulted in rhizobial survival and nodule stimulation (ElHamdaoui et al. 2003). Recently, Bibi et al. (2012) demonstrated that with increasing NaCl levels, the number of nodules per plant decreased in Vicia faba. On the other hand, Naher and Alam (2010) reported decreases in nodule number with the increase of salinity and showed a contrary effect on nodule size in several genotypes of Vigna radiata L.. Interestingly, some studies have shown a positive correlation between salt tolerance and adaptation to alkaline conditions (Kulkarni et al. 2000, Shamseldin and Werner 2005, Shamseldin 2005). 1.2.1.4 Soil nutrients (N and P) Nitrogen (N) and phosphorus (P) are equally very important nutrients for plant growth. However, their availability is generally low in arable soils, thus making them major limiting factors for crop growth and yield, especially in tropical and subtropical areas (Graham and Vance 2000; Hardarson and Atkins 2003). Today, only 30–50% of applied 14.
(35) Chapter 1. N fertilizer and 10–45% of P fertilizer are taken up by crops (Adesemoye and Kloeppe 2009; Garnett et al. 2009). In particular, nodulation and N2 fixation by numerous legumes are restricted by low availability of soil nutrients such as N, P, and micronutrients (Sanginga et al. 1995). For some researchers, low phosphorus (P) availability in the soil is the main limiting factor in symbiotic nitrogen fixation (Miklas et al. 2006). Even when the reserves of P in the soil are significant, the amount available to plants is just a tiny portion of the total (reviewed by Vessey 2003). Every aspect of the process of nitrogen fixation is limited by the availability of P (reviewed in Gyaneshwar et al. 2002). Nodules are considered an important sink for P since they demand high levels of ATP for nitrogenase functioning (Al-Niemi et al. 1998). The significance of P in nodules is reinforced by the higher P concentrations in nodules compared with other tissues (Vadez et al. 1997). Because of the high P requirements of nodules, efficient distribution and utilization of available P is essential for an optimal symbiotic interaction under P deficiency (Le Roux et al. 2006). Effects of P soil levels on the nodulation process have been described, especially on nodule number, mass or simple nodule scores. P-deficient plants are often limited in nodule initiation, development and growth, which results in low nitrogenase activity per plant and per unit nodule tissue (Olivera et al. 2004). In this regard, a decrease in nodule number and mass per plant was reported under P-deficiency with a significant genotype effect (Vadez et al. 1997). Moreover, phosphorus deficiency decreased nodule weight, number and functioning in soybean plants (Tsvetkova & Georgiev 2003). Application of P increased the number of nodules of soybean grown in heavily weathered acid soils Cassman et al. (1993) and an increased number of nodule primordia was observed with P application, thus it also has an important role in nodule initiation (Waluyo et al. 2004). In this respect, Remans (2007) showed that P deficiency strongly reduced growth and nodulation of two bean genotypes grown in different conditions. On the other hand, Sanginga et al. (1996), focusing on several African legumes grown at two P levels (0 and 7 kg P ha−1) in soils, observed that the application of P increased the weight of nodules but did not improve the proportion of N2 fixation. At a low nitrate concentration, increasing amounts of P promoted nodule formation and N2 fixation (Leidi and Rodríguez-Navarro 2000). 15.
(36) Chapter 1. Although application of nitrogen fertilizers in the soil affects the process of nodulation, it may be stimulated by relatively low levels of available nitrate or ammonia (see Zharan 1999). The endosymbiosis of legume plants and nitrogen-fixing bacteria resulted in the formation of root nodules, which is the principal mechanism of legumes plants of acquiring nitrogen (Jeuly et al. 2009). Successful nodule formation and function demand low mineral nitrogen supply and the impact of elevated rates of nitrate and ammonium sources resulted in inhibition of established nodules (Ferguson & Mathesius 2003). The amount of N2 symbiotically fixed by legume plants is strongly correlated with total nodule biomass, (Voisin et al. 2003). Transcriptome analysis in Medicago truncatula confirmed that long distance signals connecting the global plant N status, elicit a regulation on the nitrogen fixing mechanism and nodule development processes (Ruffel et al. 2008). In this study it was indicated that systemic signals of N status control N acquisition by the roots. Some experimental results demonstrated that nitrogen fertilizer application resulted in the reduction of nodule numbers in common bean (Otieno et al. 2009). Moreover, Namva et al. (2011) demonstrated that the application of high amounts of N fertilizer negatively impacted the nodulation of chickpea. Under greenhouse conditions, organic waste products significantly stimulated the productivity of legume plants as compared to plants treated with nitrogen fertilizer (Luqueño et al. 2010). 1.2.1.5 Drought stress (soil water status) Establishment and effectiveness of the legume-Rhizobium symbiosis is particularly sensitive to drought stress. Specifically, nitrogen fixation is more sensitive to water deficit than many other physiological processes (Serraj et al. 1999). Two stages of the symbiotic process are involved in decreases of nitrogenase activity under drought conditions: i. decline in nodule cortical permeability, limiting respiration, which results in a reduction in nitrogenase activity, ii. decreased activity of nodules in which nitrogenase activity can be inhibited by leghaemoglobin content (Reviewed by Hungria and Vargas 2000). The ability of Rhizobium sp. to survive at a low water potential in soil has been established by many studies. Several R. leguminosarum strains originating from saline 16.
(37) Chapter 1. areas exhibited greatest survival under low water potential. In two of them, the production of viable cells was more than 107 rhizobia g-1 of soil (Athar 1998). The response of rhizobia to low water potential results in morphological and physiological modifications, such as variations in nodule cortex structure and increase of carbohydrates in the dehydrated nodules (Reviewed by Vriezen et al. 2007). Recently Vicente et al. (2012) determined that in native rhizobia strains, nodule development and symbiotic nitrogen fixation declined with increasing levels of drought stress. Under conditions of elevated salt or other osmolytes, water stress is dominated by the movement of water in response to the water potential gradient (Reviewed by Sadowsky 2005). Many studies have focused on the response of rhizobial cells to variation in the osmotic potential induced by different osmolytes. Most of these studies evaluated the molecular mechanisms of osmoregulation induced by solutes (mainly NaCl), which are characteristic of the water stress condition in saline soils. A comparison of these responses to the responses induced by the non-permeating solute polyethylene glycol (PEG), typical for water stress situations in non-saline soils, has shown that the rhizobial mechanism of osmotolerance can differ depending on the nature of the water stress (Reviewed by Vlassak 1996). Several micro-organisms, including rhizobia, have developed a diversity of mechanisms for surviving to an increased osmolarity, mostly by increasing the concentration of a limited number of osmolytes (Mohammadi et al. 2012). By accumulating compatible solutes, R. meliloti overcomes osmotic stress-induced growth inhibition (Boscari et al. 2002). Moreover, in the presence of NaCl, sucrose and PEG, R. tropici imports potassium ions with the simultaneous synthesis and accumulation of glutamic acid in the cytoplasm (Castro and Martínez-Drets 1995). During desiccation, rhizobial populations are reduced in size (Reviewed by Vriezen et al. 2007). Therefore, their evolution and mobility is restricted under drought conditions, impacting on the bacterial community and consequently the initiation of symbiosis (Graham 1992). Both soil type and temperature are important factors influencing survival of rhizobia in desiccated soils (Boumahdi et al. 2001). Several studies showed that the decline in abundance of R. trifolii during drying on glass beads correlates with the extraction of water, which suggests that part of the decline in viable counts is caused 17.
Figure

+7




Documento similar
The objectives of this research were: (1) to assess the microbiological quality of water by counting mesophilic aerobic bacteria at different points of the WWTP; (2) to
All of these bacterial strains showed potential plant-growth promoting characteristics during the phenotypic test in selective media, suggesting that metal
This doctoral thesis aims to clarify the effect of habitual coffee consumption on the development of functional limitations, frailty and disability, as well as its effect on the
In this study, we evaluated the lipotoxic effect of PA on astrocyte cultures isolated from the hippocampus of male and female rats and our results indicate that PA reduces the
Tepary bean lectin showed dose dependent effects on the inhibition of viability as well as on colony formation in two human malignant cells lines (C33-A, Sw480); By contrast,
The tepary bean lectin showed lower hemagglutination activity than Phaseolus vulgaris hemagglutinin (PHA-E) toward trypsinized human A and O type erythrocytes.. The hemagglutination
The categories of GO terms for biological processes that are over-represented in each set of differentially expressed genes are the following: common up-regulated (A)
Plant growth promoting rhizobacteria (PGPR) can improve plant growth and soil fertility by different mechanisms such as increased mobilization of insoluble nutrients
