Heredabilidad, interrelaciones y efectos genéticos de precocidad y otros caracteres agronómicos de cuatro variedades de trigo (Triticum vulgare L )
104
0
0
Texto completo
(2) ,. . •. TESIS APROBADA POR EL COMlTE CONSEJERO:. D'~''''':ZZ~====°=t8r:.-:::~==~='-''-:T--. MARIO ZAPATA B.. I.A., Ph.. JESUS ARIAS F.. l.A., Ph. D.. JOSE RrCAURTE GARCrA. l.A.,. Ph. D•. . .• •. ií.
(3) \. . •. " El Presidente de Tesis y el Consejo examinador de Grado, no serán responsables de las ideas emitidas por el Candidato" •. (Artículo 217 de los Estatutos de la Universidad Nac ional) •. • •. tU.
(4) • i. A mí Senora, Hija y mis Padres. iv.
(5) AGRADECIMIENTOS. El Autor expresa los agradecimientos a los Miembros del Comité Consejero. Doctores. Mario Zapata B, Jesús Arias F, José RicauE te Carcla, por las valiosas sugerencias en la redacción y presentación de la Tesis, y a la Oficina Central de Estadistica.. v.
(6) CONTENIDO Página. ;. .•. ,-. ¡. 1.. INTRODUCCION. 1. 2.. REVISION DE LITERATURA. 4. 2.1.. HETEROSIS. 4. 2"2,,. HEREDABIL IDAD. 8. 2.2,10 Conceptos Generales sobre Heredabi lidad. 8. 2 .. 202~ Usos de la Hereda.bilidad. 10. 2.2.3. Métodos para Calcular Heredabilidad. 11. 2.3.. CORRELACIONES FENOTIPICAS y GENOTIPICAS. 17. 2.4.. EFECTOS GENETICOS. 19. 2.5.. GANANCIA GENETICA. 21. 3.. MATERIALES Y 1-!ETODOS. 22. 3,1.. MATERIALES. 22. 3.2.. METODOS. 22. 3.2.1- Datos tomados sobre cada Planta en cada. 24. Generaci6n. ". • ,. 3.2.2- Promedio y Varianzas. 2.1. 3.2,3, Heredabilidad en Sentido Amplio. 25. 3.2.4. Heredabilidad en Sentido Estrecho. 25. 3.2.5. Correlaciones Fenotípicas. 26. 3.2.6. Correlaciones Genotipicas. 26. 3.2.7. Modelo de Hayman. 27. 3.2.8. Ganancia Genética. 28. vi.
(7) Página. ... .:;. ;.. RESULTADOS. 30. COMPORTAMIENTO DE LOS PADRES Y GENERACIONES. 30. 4.2.. HETEROSIS. 33. 4.3.. VARIANZAS DE GENERACIONES PARA CADA CARACTER. 35. 4.4.. ESTIMATIVOS DE HEREDABILIDAD. 38. 4.5.. CORRELACIONES FENOTIPICAS. f,l. 4.6,. EFECTOS GENETICOS. 45. 4.7.. GANANCIA GENETICA. 48. 5.. DISCUSION. 52. 5.1.. EPOCA DE ESPIGAMIENTO. 5.2.. RENDIMIENTO DE GRANO POR PLANTA. 56. 5.3,. PESO DE 50 GRANOS. 58. 5.4.. MACOLLAMIENTO POR PLANTA. 60. 5.5.. LONGITUD DE ESPIGA. 61. 5.6.. GANANCIA GENETICA. 64. 6.. CONCLUSIONES. 68. 7.. RESUMEN. 72. 8.. SUMMARY. 75. BIBLIOGRAFIA. 78. APENDICE. 86. • •. ·. •. , •. ,. y. y. GENOTIPICAS. MADUREZ FlSIOLOGICA. • vii. 52.
(8) LISTA DE TABLAS. ,. .. Página TABLA 1.. Número de planta utilizadas en cada. gene~. raci6n, nombre de las variedades para cada cruzamiento y lugar de obtenci6n de c,! da una ... TABLA 2.. Promedio de generaciones para época de pigamiento madurez fisiol6gica y. e~. rendimie~. to de grano por planta de cuatro cruzamiea tos de trigo, Tibaitatá TABLA 3.. 31. 1973.. Promedio de generaciones para peso de 50 grano~macollamient~. y longitud de espiga. de cuatro cruzamiento de trigo, Tibaitatá 32. 1973. TABLA 4.. Heterosis observada en la generación Fl d,! da en porcentaje en base al padre superior y al promedio de los padres para los. cará~. teres estudiados en cuatro cruzamientos de trigo, Tibaitatá TABLA 5.. 1973.. Varianza de generaciones para época de espigamiento, madurez fisiológica y. rendimie~. to de grano por planta de cuatro cruzamientps de trigo, Tibaitatá. 1973.. viii. 36.
(9) Página TABLA 6.. Varianza de generaciones para peso de 50. grano~. macollamiento y longitud de. espiga de cuatro cruzamientos de trigo, Tibaitatá TABLA 7.. 1973. 37. Estimativos de heredabilidad en sentido amplio y heredabilidad en sentido. estr~. cho en porcentaje para los carácteres estudiados en cuatro cruzamientos de tri go, Tibaitatá. 1973. TABLA 8.. 39. Correlaciones fenotípicas y genotípicas de seis carácteres agronómicos en todas. ". sus combinaciones posibles de los. cruz~. mientos Bonza 63 x Super Precoz e ICASamaaá 68 x Super Precoz, Tibaitata. 1913. TABLA 9.. 42. Correlaciones fenotípicas y genotípicas de seis carácteres agronómicos en todas sus combinaciones posibles de los. ... cruz~. mientos Icatá-l x Super Precoz y Coconuco x Super Precoz, Tibaitatá. 1973. ix. 43.
(10) Página TABLA 10. .'. Estimativos de efectos genéticos para. 108. carác teres ea tudiados en los cruzamientos de trigo Bonza 63 x Super Precoz y Cocon,!! co x Super Precox, ajuste del X2 &1 modelo de tres paré.metros y los tres parámetros genéticos, Tibaitatá TABLA 11.. 46. 191:3.. Estimativos de efectos genéticos para los carácteres estudiados en los cruzamientos de trigo Bouza 63 x Super Precoz y Cocon,!!. .. ca x Super Precoz, ajuste del x2 al modelo de tres parámetros y los tres parámetros. '<. genéticos, Tibaitatá TABLA 12.. 47. 1973.. Ganancia genética esperada en base a. here~. dabilidad en sentido amplio y a heredabil! dad en sentido estrecho. p~ra. los carácteres. estudiados en cuatro cruzamientos de trigo, Tibaitatá. 1973,. 49. ,>. •• x.
(11) LISTA DE TABLAS DEL APENDICE Página. •. TABLA l.. Epoca de espigamiento y madurez fisiológica del cruzamiento 4 (Coconuco x Super Precoz) en las respectivas generaciones.. TABLA 2.. 87. Rendimiento de grano por planta y pese de 50 granos del cruzamiento 4 (Coconueo x Super Precoz) en las respectivas gene88. raciones.. TABLA 3.. Epoca de espigamiente y madurez fisiol6gica del cruzamiento 1 (Bonza 63 x Super Precoz) en las respectivas generaciones.. TABLA 4.. 89. Rendimiento de grano por planta y peso de 50 granos del cruzamiento 1 (Bonza 63 x Super Precoz) en las respectivas gene90. raciones.. •. • xi.
(12) DEFINICION DE TERMINOS UTILIZADOS EN EL PRESENTE TRABAJO ••. •. Heterosis. =. Vigor híbrido tal que UD h1brido Fl cae fuera del intervalo de sus genitores con respecto a. •. uno o varios carActeres. Dominancia. =. Interacci6n·intraaUlica tal que uno de los alelos se manifiesta más o menos, en el homocigote, que su alelo alternante.. Epistasis. =. Describe todos los tipos de interacción intera1'lica. e~. donde cualquier manifestaci6n en un. locua está afectada por la fase gen'tica de cualquiera o de todos los demás loci. A los genes que no muestran dominancia se les llaman algunas veces "genes adi Uvos" o se dice que tienen efectos genéticos aditivos; cuan-. Efectos =. do se refiere a los genes de un locus significa ausencia de dominancia y cuando se refiera a ge, nes en diferentes loci significa ausencia de e-. aditivbS. pistasis.. ... Efectos genéticos dominantes. Cuando los genes en un 10cus muestran dominancia =. se les llaman "genes dominantes" o se dice que tienen efectos gen'ticos dominantes.. xii.
(13) Segregación •. transgresiva. Aparición en generaciones segregan tes de indi=. viduos que estAn fuera del intervalo de los nitores con respecto a. algú~. carácter.. Retrocruza-. Cruzamiento de un híbrido con cualquiera de. miento. sus padres. En. genétic~. g~. cruzamiento de Un. het~. rocigote con un recesivo homocigótico. Media de los cuadrados de las desviaciones de. V'arianza. .. una variable de su media. Cantidad numérica que da una característica de. Parámetro. "' una población.. Heredabilidad. Es la relación entre la varianza genética adi-. en aentido. tiva y la varianza fenotípica. En la revisión. estrecho. de literatura y en materiales y métodos se puede aclarar mAs este concepto.. Heredabilidad en sentido = amplio. Es la relación entre la varianza genética total y la varianza fenot1pica.. Es el avance genético esperado bajo selección, ". mide la diferencia entre el valor promedio genoGanancia tipico de las lLlIleas seleccionadas y el valor pr.Q Genética. =. medio genot1pico de lasliceal originales. xiii.
(14) Correlaciones ~. ••. fenotípicas. Mide el grado de '". o carActeres decir. ,. asociació~. agro~ómicos. te~iendo. entre variables. fenot1picamente, es. en cuenta la variáción. gen~tica. y la ambiental.. Correlaciones Genotípicas. Mide el grado de. '". agro~ómicos. asociació~. entre carácteres. genotip1csmente, es decir sin te-. ner en cuenta la variación ambiental. Es. una caracter1stica agronómica que mide la. Madurez. precocidad en dias de una variedad, en éste. Fisiológica. caso de trigo. En materiales y métodos se explica. CQJIIC. fué calculada •. •. Formación de individuos diferentes en el genotipo, o la presencia de individuos genct1pica-. Variabilidad Gen~tica. '". mettte diferentes, en contraste a las diferencias inducidas por el ambiente, el cual causa cambios temporales, no heredables de el fenotipo (variaci6n) •. •. • xiv.
(15) l.. INTRODUCCION. El trigo es uno de los alimentos básicos en la dieta alimenticia. ,• •. no sólo de los colombianos sino de los habitantes del mundo en general. Es de todos conocida la gran cantidad de divisas que gasta nuestro país anualmente en importaciones, para suplir la demanda nacional de éste cereal; en el ano de 1972 la demanda fué de 449.200 toneladas y se importaron 380.000 por un valor de 675 millones de pesos, habiendo sido su importación. en toneladas aún mayor en 1971 *.. Uno de los medios para satisfacer las necesidades de una población en constante crecimiento demográfico, es incrementar los rendimientas por unidad de superficie, 10 cual puede lograrse en parte, con la contribución del fitomejoramiento al proporcionar variedades de. p~. riada vegetativo corto y altos rendimientos. Es~c. estudio es un aporte al parcial cumplimiento de los objetl. vos del Programa Nacional de Trigo del Instituto Colombiano. Agropecu~. ri.o, Le.A, cuya meta primordial en los últimos anos ha sido la de. i~. tensificar la obtenéión de variedades precoces, de altos rendimientos, resistentes a enfermedades y de buena calidad panadera y molinera.. •. •. '>. * Colombia. Ministerio de Agricultura. Oficina de Planeamie~ to del Sector Agropecuario. Programas Agrícolas. (Bogotá, 1972) pp.81-89..
(16) 2. La obtención de variedades con las características antes anota-. •. das es importante porque algunas veces pueden escapar al ataque de enfermedades, heladas y factores climáticos adversos en los últimos. est~. dos del cultivo. Es importante también porque contribuyen a solucionar en parte los problemas de rotación de cultivos ya que los cultivadores pueden dar una mejor utilización a la tierra, especialmente en el. segu~. do semestre. Con variedades precoces se podrá obtener mayor rentabilidad en el cultivo de trigo sin aumentar las inversiones del agricultor. Es lógico que se necesita experiencia en Programas de. Mejoramie~. to para proporcionar evidencias sobre utilidad de selección de carac'teres, como precocidad, en relación con rendimiento. Los estimativos de la acción genética que contribuyen a la expresión de los caracteres, hetedabilidad, ganancia genética y correlaciones entre ellos son de importancia en Programas de Mejoramiento puesto que constituyen la base para establecer los criterios que se deben seguir en la selección del material segregante.. 1.1.. •. •. •. OBJETIVOS.. Los objetivos de éste estudio fueron determinar: a). Heterosis. b). Heredabilidad en sentido amplio y heredabilidad en sentido estrecho.
(17) 3. e). Ganancia $enética. d). Correlaciones fenotípicas y genotípicas entre caractéres. e). Efectos genéticos que contribuyen a la expresión de cada uno de los caractéres.. Los caractéres analizados en el presente trabajo fueron: Epoca de espigamiento, madurez fisiol6gica, rendimiento de grano por planta, peso del grano, macollamiento por planta y longitud de espiga. El trabajo se realizó durante los anos de 1972 y 1973, en el •. Centro Nacional de Investigaciones Agropecuarias "Tibaltatá" del Instituto Colombiano Agropecuario, LC.A.. •. •.
(18) 2.. 2.1.. REVISION DE LITERATURA. HETEROSlS. En el primer decenio del redescubrimiento de los trabajos de. Mende1, la hipótesis de la dominanc;l,a genf,tica para explicar el viger híbrido fue propuesta por Davenport en 1908, Bruce en 1910, y Keeble y Pellew en el mismo afta, citados por Allard (1967). Bajo esta hipótesis, el entrecruzamiento de líneas puras conducirá a la for mación de híbridos en lasque los genes perjudiciales recesivos procedentes de un padre están ocultos, como en el material original de polinización abierta, por alelas dominantes procedentes del otro padre. Otra hipótesis, propuesta independientemente por Shuell y East, en 1908, supuso que hay un estímulo fisiológico del desarrollo que aumenta con la diversidad de los gametos que se unen. Esta hipótesis Be conoce generalmente como hipótesis de la superdominancia de la he terosis (Allard 1967). Las observaciones de heterosis en trigo datan desde 1919 cuando Freeman citado por Briggle (1963), estudió la época de aparici6n de la primera espiga, altura y ancho de la hoja en cruzamientos in-. ,. cluyendo trigo Durum y 3 testigos comunes. Briggle (1963), hace una revisi6n de heterosia en trigo y concluye que todas las combinaciones parentales no resultan en expresi6n de vigor híbrido; algunos híbridos no se comportan tan bien como ambos padres incluidos. Mucha.
(19) 5. investigación falta por hacerse concerniente a la facilidad de •. produ~. cir trigo híbrido a escala comercial • McNeal. ~. al. (1965),al evaluar tres cruzamientos de trigo de. priGavera para heterosis encontraron que la F1 y F2 fueron intermedias entre los padres, y en ningún caso los caracteres agronómicos estudiados como rendimiento, componentes de rendimiento y calidad, excedieron al padre superior por una cantidad estadisticamente significante. El comportamiento de estos cruzamientos indicó que la e! trecha relación de los padres puede dsr poblaciones híbridas do poco a no heterosis. •. Heyne y Finney (1965), obtuvieron. m08tra~. 108. mis-. mos resultados al estudiar rendimiento, macollamiento, peso de grano, longitud de espiga y calidad en trigos rojos duros de invierno. Efectos heteróticos no fueron observados para componentes de rendimiento excepto peso de grano. El promedio de rendimiento de todos los Fl híbridos de trigo. --. en un experimento realizado por Gyawali et al. (1968), .. fu~. 24 por. ciento mayor que el mejor padre en cada cruzamiento. Híbridos rojo. .' •. •. blando x rojo blando y rojo blando x rojo duro dieron similares. v~. lores de heterosis, indicando que diversidsd de clases no son necesarias para heterosis. Híbridos con padres precoces y tardíos produjeron mAs heterosis para rendimiento de grano que híbridos con ambos padres precoces o ambos tardíos. En relación a peso de grano, el grano híbrido. fu~. más pesado que el promedio de padres en todos.
(20) 6. •. -. ,. los híbridos y más pesado que el padre mayor en 19 de los 21 híbridos, sin embargo, 2 de los híbridos solamente mostraron heterosis. e~. tadísticamente significativa. Heterosis para número de espigas sobre el promedio de padres fué evidente y la mayoría de los híbridos que presentaron heterosis significativa para rendimiento también presentaron heterosia positiva para número de espigas. Niveles de vigor híbrido fueron examinados por Fonseca y Patterson (1968 a) para precocidad, altura, tres componentes de rendimiento y rendimiento de grano en las generaciones Fl y F 2 de un. cr~. zamiento dialélico de siete variedades seleccionadas de trigos de invierno. Poca heterosis sobre el padre más precoz fue obtenida para precdcidad y, para número de espigas por surco fue significativa en solo tres de los 21 híbridos Fl;no fue significativa para. n~. mero de granos por espiga. La heterosis para el peso de grano varió de 101 a 115 por ciento obtenida en todos los 21 híbridos pero significante en 5 de ellos. La heterosia para rendimiento fue generalmente más alta que para los componentes individuales de rendimiento y solamente significativa en cuatro de los 21 híbridos. ".'. Wells y Lay (1970), en cruzamientos de trigos de primavera. .•. e~. contraron rendimientos que variaron de menos 14 por ciento a más 82 por ciento sobre el mejor padre. Walton (1971) , en 1968 observó. re!!;. dimient08 de 10 por ciento a 26 por ciento más que el padre más ren 1idor; en 1969, sin embargo, el rendimien to de los mismos h1bridos.
(21) 7. •. o. ••. ,. ,. fue de -11 por ciento a 4 por ciento sobre el padre más rendidor. Reterosis en avena fue reportada por Petr y Frey (1970), para rendimiento de grano en la Fl que varió de 1 a 30 por ciento sobre el mejor padre, con un promedio de 12 por ciento para todos los cr,l! ces. Positiva heterosis fue expresada para el número de panículas por planta, y para cada cruzamiento la época de espigamiento de la Fl , fue mas precoz que el promedio de los padres. Upadhyaya y Rasmusson (19ó7), en trabajos con cebada obtuvieron valores promedios de heterosis de las Fl basados en el promedio de los padres para rendimiento, peso de grano, granos por espiga, espigas por planta de 21,5; 5,9; 7,1 y 7,6 respectivamente. Cuando las Fl fueron comparadas con el padre mayor el promedio de heterosis para x>endimiento fué de 9,1 por ciento.. Gebrekidan y Rasmusson. (1970), al evaluar 27 híbridos de cebada en 4 ambientes seontraron heterosis sobre el padre superior, de 26 a 47 por>ciento con un. pr~. medio de 11 por ciento. Variedades de alto>rend1miento produjeron una gran proporción de híbridos de alto rendimiento en relación a variedades de bajo. ·,. rendimiento, mientras, máxima heterosis ocurri6 en cruces entre variedades de bajo rendimiento. Heterosis sobre el mejor padre no fue apreciable para caracteres comO números de espigas, granos por esp! ga, peso de grano y días de espigamiento..
(22) 8. Los estudios realizados por Pinthus (1963), sobre herencia del. •. espigamiento en trigo mostraron que la iniciaci6n temprana del espigamiento fue dominante en relaci6n a la iniciaci6n tardía, y un corto período de iniciaci6n a espigamiento fue dominante sobre un largo. p~. riodo. Petr y Frey (1966), en trabajos con avena observaron sobredominaneia para número de panículas por planta en la Fl , mientras estimativos para rendimiento variaron de parcial a sobredominancia. Para panículas cortas parcial dominancia, precocidad fue parcialmen te dominante y también pocas esiguillas por panícula.. 2.2.. •. HEREDABILIDAD .. 2.2.1.. Conceptos generales sobre heredabi1idad. La efectividad de la selecci6n para un carácter particular en. poblaciones de plantas es determinada en gran parte por la re1aci6n de variabilidad genética en relación a la variabilidad fenotípica entre plantas. La idea de separar la variabilidad. fenotípica de. una poblaci6n biológica en dos porciones, genética y ambiental, fue. .' •. ,. •. primero propuesta por Fisher, citado por Burton (1951). Fisher también demostr6 que la varianza genética total podría dividirse en: 1.. Varianza genética total, incluyendo: a. Varianza genética aditiva (Vg). b. Varianza debida a desviaciones dominantes del modelo aditivo..
(23) 9. .. .. .. ,-. c. Varianza debida a la interacción de genes no a1é1icos • 2.. Varianza debida a interacciones de los genotipos y el medio •. Heredabi1idad en sentido amplio es la relaci6n de la varianza genética total con la varianza fenotípica. Heredabi1idad en sentido es trecho es la relación de la varianza genética aditiva con la varianza fenotípica (Al1ard 1967). Para definir la heredabi1idad, se debe delinear un modelo que caracterice la variabilidad para un conjunto de observaciones. Hanaon (1963), estableció un valor para el estimativo de la expresión de un caracter dado (fenotipo). Este valor esté compuesto por la suma de dos componentes; el primero constituido por la porción genética aditiva (Ai), la porción genética dominante (Di) y la porción espistética (I i). El segundo componente comprende el efecto ambiental e incluye la interacción entre la porción genética y el ambiente (GE) 1j e incorpora además las variaciones causadas por fluctuaciones del ambiente que ocurren al azar N1jk.. Con 10 anterior el fenotipo re-. sultante se expresa en la siguiente forma:. ". • •. Yijk. = Ai. + Dí + Ii + (GE) 1j + Nijk. La heredabi1idad en sentido amplio estaria dada por la fórmula: Heredabilidad en Sentido amplio. Varianza aditiva + V.dominante + V. epitatica (V.aditiva+V.dominante+V. epitética)+Vg.xA+V. error).
(24) 10. •. Heredabilidad en Sentido estrecho. V. Aditiva. (v.aditiva+v.d~inante+V.espistática)+(v.g x a+V.error). , La varianza aditiva es el componente importante, puesto que es la causa principal de la semejanza entre padres y por lo tanto, la principal determinante de las propiedades genéticas observables de la poblaci6n y de la respuesta de ésta a la selección. Más aún es la única componente que puede ser estimada directamente a partir de las obseE :vaciones hechas en la población segregante. (Falconer, 1971). 2.2.2. •. Usos de la heredabilidad. Robinson (1963), afirma que cuando en una investigación se requi,!!.. re encontrar estimativos de heredabilidad es necesario conocer, con anterioridad que tipo de estimaciones son requeridas. Las razones que se dan para conocer el tipo de heredabilidad son: 1. Cuando la heredabilidad en sentido estrecho es alta se debe. usar como método de selección principalmente el masal; y cuando la heredabilidad empieza a ser baja se debe dar más énfasis a los métodos de pedigree. 2. Si la varianza debida a la epts tas is es re1a ti vamen.te al ta, se. • •. debe dar más importancia a la selección dentro de familias y al método de la linea pura. 3. Si la sobredominancia es alta, el método de mejoramiento a seguir es el de la endocr1a con el objeto de producir posteriormente h1-.
(25) 11. -bridos comerciales, principalmente en plantas alógamss. 4. Si la varianza debida a las interacciones dentro de la heredabilidad y el ambiente es relativamente grande, los. m~todos. de. mejoramiento deben tender mAs a producir variedades individuales para cada zona ecológica. La heredabilidad tiene algunas limitaciones en el mejoramiento de plantas, pues los caracteres mAs importantes como rendimiento y la cslidad están influenciadas por el medio. Además, esos carActeres son controlados por numerosos genes y su heredabilidad es baja y. •. por consiguiente el índice de selección no tiene mucha aplicación en las primeras generaciones. Precocidad se ha determinado como de alta heredabilidad en cereales como trigo y cebada, y los autores Pwers (1934), Pinthus (1963), Griffee (1925), Wexelsen (1933), Frey (1954), Fiuzat y Atkins (1953) encontraron que estaba determinada por menos de tres pares de factores. 2.2.3.. Métodos para calcular heredabilidad. Las. t~cnicas. para estimar el grado de heredabilidad en plantas. de cultivo econ6mico previamente reportadas por la literatura, caen. • • •. principalmente en cinco categorías: a) Regresión desendencia-padres. b) Componentes de varianza de un análisis de varianza..
(26) 12. ... c) Aproximación de la varianza no heredable de poblaciones ge' néticamente uniformes, para estimar la varianza genética. ,. t~. taL d) Unidades estandar. e). ~todos. presentado por Warner (1952) para determinar hereda-. bilidad en sentido estrecho. En el presente trabajo se utilizaron los métodos (e) y (e). a) Regresi6n descendencia-padres. El método consiste en hallar el coeficiente de regresión "b", usando como variable dependiente las características de la progenie y como independiente las de los padres. El valor da "bU eS igual al valor de la heredabilidad. Kronstad y Foote (1964), estimaron heredabiltdad en el sentido estecho por regresión en un cruzamiento dialé1ico de 10 padres en trigo de invierno, los estimativos para número de espigas por planta, espiguillas por espiga, granos por espiguillas, peso del grano fueron altamente significantes, excepto para rendimiento de grano. Fonseca y Patterson (1968 b), utilizaron regresión de Fl ó F2 sobre valores de los padres en trigo de invierno y obtuvieron los mismo resultados en here• •. •. dabllidad • Frey (1954 a), obtuvo heredabi1idad en cebada de la regresión del comportamiento de la generación F5 sobre la F4 para rendimiento, puntaje.
(27) 13. y espigamiento con 39, 96 Y 93 por ciento respectivamente, Rasmusson y Canne1 (1970), con el mismo procedimiento obtuvieron heredabi1idad de la relaci6n de respuesta en F a la selecci6n diferencial en F , 4 S y los valores obtenidos para número de espigas fue de 22 por ciento, granos por espiga de 8,6 por ciento, peso de grano 43,4 por ciento y rendimiento 18,2 por ciento. Frey y Borner (1957), y Hanson (1963), sostienen que la in teracci6n genotipo por ambiente asociada con una variaci6n mayor o menor de la escala fenotípica puede influir significativamente en la estlmaci6n de la heredabilidad y crear condiciones que hacen que esas estiID! ciones sean mayores de las que puedan obtenerse realmente, b) Componentes de varianza de un anAlisis de varianza. Davis (1961), obtuvo por este método heredabilidad en rendimiento de grano. •. de 40 por ciento en promedio de cuatro poblaciones diferen,tes de trigo. ~ebsock. y Amaya (1969), obtuvieron heredabilidades promedias de cuatro. cruzamientos de Triticum durum desf. Para peso de grano, puntaje y número de granos p'or espiga de 72, 67 y 69 por ciento respectivamente.. --. Busch et al. (1971), utilizaron trigos de primavera y obtuvieron para rendimiento de grano 29,6 por ciento en promedio de tres cruzamientos.. • • Estimativos de heredabilidad en avena basado sobre un afto,. •. calc~. lados en base a componentes de varianza fueron uniformemente altos para rendimiento, puntaje y espigamiento. Para altura y madurez fueron vari! bIes en las generaciones F2 y F3' Estimativos durante 2 aftos resultaron.
(28) 14. bajos para madurez; para espigamiento, rendimiento y altura permanecieron al tos. 'Al utilizar el método de regresión, la variación fue mayor en estos carácteres, (Pawlish y Shands, 1962). Frey y Horner (1955), al comparar los métodos 'de regresión y de componentes de varianza encontraron que el último, dá más exactitud entre predicción y gananciss actuales en experimentos de selección. Regresión tiende a subestimar ,el verdadero porcentaje de heredabilidad, Solamente espigamiento estuvo de acuerdo con la heredabilidad calculada por los dos métodos • •. Borthakur y Poehlman (1970), estimaron la heredabilidad de peso de grano en dos cruzamientos de cebada usando los padres y las generaciones Fl , F2 Y F3 , Los métodos usados fueron los de regresión de F3 - F , componentes de varianza y unidades estandar. Los estimativos 2 para esta caracteristica calculados por estos métodos fueron muy diferentes. Los obtenidos por componentes de varianza fueron los más altos alrededor del 45 por ciento. c) Aproximación de la varianza no heredable de poblaciones genéticamente uniformes para estimar la varianza genética total.. • •. Mahmud y Kramer (1951), para averiguar la heredabilidad en sentido amplio en soya, y Peter y Frey (1966), en avena, utilizaron como varianza no heredable a la raiz cuadrada del producto de las varianzas de los dos padres. Estos últimos obtuvieron valores de heredabilidad.
(29) 15. para longitud de pan1cula, espigamiento, panículas por planta y ren-. ·. dimiento de grano de 54, 87, 33, Y 53 por ciento respectivamente. E,!!. .. te procedimiento es el utilizado en el presente trabajo. Burt6n (1951), utiliz6 como varianza fenotípica la varianza de la F2 y como varianza ambiental la de la Fl en millo. Weber y Moorthy (1952), en estudios con s·oya utilizaron la raíz cubica del producto de la varianza ambiental. Johnson, Robinson y Comstock (1955), discuten la importancia de la interacci6n de genotipo x medio al estimar heredabilidad; en soya utilizan la varianza de la Fl y de los padres como varianza ambiental.. • d) Unidades estandar. Propuesto por Frey y Horner (1957), en avena de acuerdo a los autores, con este método se tiende a eliminar los efectos ambientales de diferentes aflos los cuales hacen disminuir los estimativos de heredabilidad. Lofgren et al. (1968), utilizaron los métodos de. regr~. sión y de unidades estandar y encontraron diferentes valores con los dos métodos para estimativos de contenido de proteína, calidad y. co~. ponentes de rendimiento estudiados en trigo. Concluyeron que la vari.!!. ci6n fenotípica fue apreciable de afio a afio. Reddi !! al. (1969), uti-. • •. lizaron los dos métodos anteriores y encontraron baja heredabilidad p.!!. ra peso de grano en dos cruzamientos de trigo. Una supuesta selecci6n y procedimiento de prueba fue designsda, en la cual rendimientos de 1, 2, 3, 4, y 5 aflos fueron combinados al azar para determinar el. efe~.
(30) 16. .,. to de número de años probados en relación a la heredabilidad obtenida con el promedio de 6 años. Ningún cambio considerable o consistente en los valores de unidades estandar fueron obtenidos cuando el número de años probados' fue incrementado. e) Método presentado por Warner (1952). Por éste método, el doble de la varianza de la F2 menos la suma de las varianzss de los dos retrocruzamientos fue usada para estimar la varianza aditiva. Johnson !! al. (1966). obtuvieron heredabilidad en druzamientos de trigo de invierno en sentido amplio con el empleo de la varianza de la Fl como, varianza no heredable y en el sentido estecho por el método de Warner. En la mayor1a de los casos heredabilidad en el sentido estrecho fue más pequeño que la correspondiente en el sentido amplio. Valores para longitud de espiga y peso de grano fueron los más consistentes entre el sentido amplio y el sentido estrecho. Muy bajo estimativo en el sentido estrecho fue obtenido para número de espigas por planta con un valor de 3,4 por ciento. Anwar y Chowdhry (1969), en cruzamientos de trigo de primavera emplearon la varianza de los padres como varianza ambiental para determinar la heredabilidad en sentido amplio y en sentido estrecho por el. • •. método de Warner, y obtuvieron similares resultados por los dos métodos. Los valores en el sentido amplio variaron de 50 a 70 por ciento y en el sentido estrecho de 12 a 40 por ciento; los valores bajo fueron para dimiento de grano y altos para espigamiento y altura.. re~.
(31) 17. 2.3. CORRELACIONES FENOTIPICAS y GENOTIPICAS. Johnson et al. (1966), obtuvieron en un estudio de 4 aftos con cuatro variedades de trigo de invierno diferentes en altura, que las variedades de tallo corto fueron más productivas en promedio. La variedad de alto rendimiento produjo consistentemente más granos por espiga, pero su peso de grano y número de espigas fue menor que las otras variedades. Shlehuber. ~. al. (1967), encontraron que no hay asociación ge-. nética entre espigamiento y características de calidad. Ellos obtuvieron correlaciones positivas genotípicas y fenotípicas entre espigamiento y altura, espigamiento y rendimiento. Baker et!l. (1969), observaron correlaciones significativas de 0,33 entre peso de 1.000 granos y rendimiento. La correlación fenotípica entre precocidad y rendimiento fue baja y negativa (-0,2883). La precocidad tuvo mayor influencia como efecto indirecto negativo sobre número de espigas (-0,3825). Los genotipos precoces mostraron una tendencia a tener pocos tallos y alto peso de grano pero pocos granos por espiga. Encontraron correlación. ,. .. alta y positiva entre número de espigas y rendimiento con un valor de 0,7082 e intermedia entre rendimiento y peso de grano con 0,4006 (Fonseca y Patterson, 1969 b). Lebsock y Amaya (1969), en Triticum durum Desf. Por medio de.
(32) 18. correlaciones fenotípicas y genotípicas concluyeron, que la selección para plantas precoces y de tallo corto podrían resultar en líneas con bajo peso de grano y puntaje. Correlaciones positivas y significativas se presentaron en algunos cruzamientos entre peso de grano y rendimiento, y entre rendimiento y puntaje. Reddi et al. (1969), observaron en trigo de primavera que las 11neas con tallo más corto tiende a tener espigas más largas y peso de grano menor. Mufioz y Zapata (1973), en un estudio con cuatro variedades de trigo y diferentes prácticas de cultivo durante cuatro semestres obtuvieron correlaciones fenotípicas positivas de rendimiento con maco11amiento y puntaje, y negativas con la altura. Macollamiento aumentó a medida que disminuy6 la densidad de siembra; se observ6 que a mayor longitud de espiga mayor rendimiento. El puntaje mostró correlaci6n positiva con macollamiento y altura negativa con el mismo carácter; resultados similares fueron observados por Paw1ish y Shands (1962), en avena. Ross (1955) en estudios con avena observó que las variedades precoces tenían menor número de hojas al tiempo de la iniciación de la pa-. • •. nícula, menor número entrenudos y la longitud del pedúnculo más corto que las variedades tardías. Murphy (1966), encontró correlaciones positivas. •. y significativas de rendimiento con panículas por planta y semillas por panícula, y negativas con peso de semilla. Las semillas por panícula.
(33) 19. con peso de semilla present6 correlaci6n negativa. Grafius (1956), con 40 variedades de avena encontr6 que el número de panículas por unidad de área y tamafto de panículas influyó en el rendimiento, pero que el tamafto de semilla no. Rasmusson y Cannel (1970), obtuvieron correlaciones fenotípicas en cebada entre rendimiento y cada uno de los componentes de rendimiento (granos por espiga, número de espigas y peso de grano) y no entre ellos.. 2.4. EFECTOS GENETICOS. Hayman (1958), propone un modelo de 3 y 6 parámetros. Cuando epístasis está ausente dd"d m, d y ti miden un valor constante, aditividad y dominancia. Cuando epístasis está presente y los datos Se ajus,. ,A,. '-'. ,Ao... ~. ~. ~. tan al modelo de 6 parámetros, m, d, ti, i, j Y 1 miden una constante, aditividad, dominancia y las tres clases de ep1stasis. En estas circunstancias las desviaciones de los promedios observados en las generaciones de sus promedios esperados, sobre el modelo de 3 parámetros, son epístaticos en naturaleza y revelan que clases de epístasis están modificando algún promedio dado. Este modelo es utilizado en este tra-. •. •. bajo. (El asterisco sirve para diferenciar entre modelo de tres y seis parámetros) . Brow. ~. al. (1966). en estudios sobre cruzamientos dial€licos. en trigo para determinar la acción de los genes no detectaron habilidad.
(34) 20. combinatoria específica; estos resultados con aquellos de Kronstad. ,. y Foote (1964), Wells y Lay (1970), Fonseca y Petterson (1968 b) Y Gyawa1i !! al. (1968), sugieren que una mayor parte de la variación genética para rendimiento en trigo puede ser explicada por habilidad combinatoria 'general y por lo tanto, en gran parte, de acción genética aditiva. Walton (1971), en dos-cruzamientos dia1é1icos encontró habilidad combinatoria general y en uno de ellos habilidad combinatoria específica altamente significativa para espigamiento en los dos cruzamientos. Al estudiar los efectos genéticos en la herencia de carActeres en trigo por medio del modelo de Haymsn, Chapman y McNea1 (1971), observaron en los dos. a~os. estudiados que la acción genética aditiva. obtuvo el mayor efecto sobre nÚmero de espigas y, tipos de acción genética aditiva y dominante influyeron en el peso de grano significativamente. Chapman y MeNes1 (1970), con iguales procedimientos encontraron efectos genéticos aditivos altamente significativos para proteína en el grano y solo dos de los cinco cruzamientos mostraron efectos do-. •. •. minantes significativos. Efectos epístAticos no fueron significativos en la herencia del número de espiguillas y peso de grano. Tipos de acción de genes aditivos y dominantes influyeron en el peso del grano. Los resultados del.
(35) 21. análisis genético indicaron que la selección de plantas individuales debería ser efectiva para época de espigamiento. Menores valores fueron obtenidos del análisis de tres cruzamientos para rendimiento (Bush et al. 1971). Upadhyaya y Rasmusson (1967), y Smith y Lambert (1968), en sus estudios encontraron significativa habilidad combinatoria general para rendimiento, peso de grano y madurez en cebada.. 2.5.. GANANCIA GENETICA. En experimentos con trigo Durum, Lebsock y Amana (1969), encon-. traron que las ganancias genéticas estimadas en base a heredabilidad en sentido amplio y las ganancias observádas, fueron similares para madurez y peso del grano. La ganancia esperada para madurez en cuatro cruzamientos, al seleccionar las plantas más precoces varió de -1,3 a -3,5 días y la observada varió de -0,8 a 3,2 días. La ganancia estimada para peso del grano varió de 0,17 a 0,40 gramos/lOO. Encontraron desacuerdo entre los valores estimados y observados para rendimiento y macollamiento.. •. •. Allard (1967), al utilizar la heredabi1idad en sentido estrecho encontró en trigo una ganancia genética estimada para espigamiento de. ,. 9,4 días..
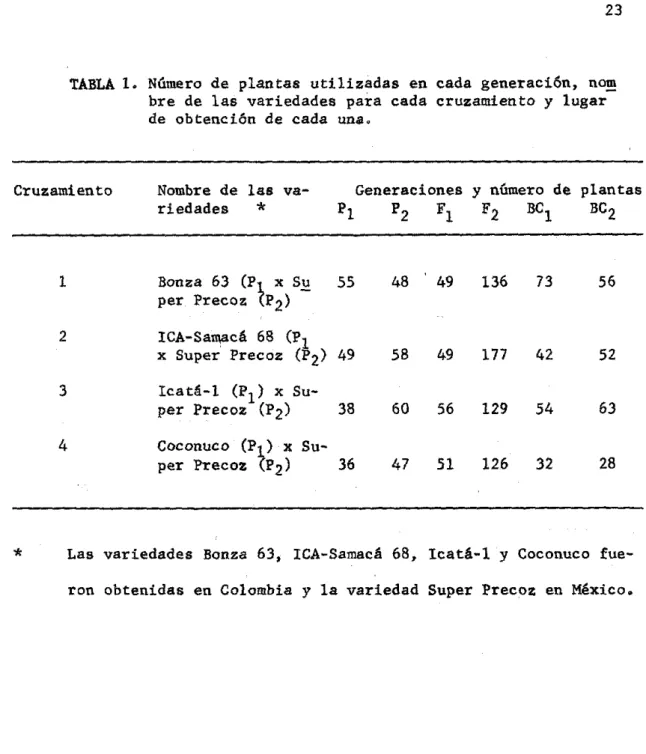
(36) 3.. 3. 1 •. MATERIALES Y METODOS. MATERIALES. El material. experimental usado en este estudio consistió de. poblaciones de cuatro cruzamientos en trigo; Triticum vu1gare L. En la tabla 1, aparece el número de plantas utilizadas en. c~. da generación nombre de las variedades para cada cruzamiento y lugar de obtención de cada una de ellas. Las variedades Bonza 63 e ICA-Samacá 68 por su periodo vegetativo largo se llamarán en esta investigación variedades tardías, las variedades Icatá-l y Coconuco de un periodo vegetativo intermedio se llamarán variedades intermedias en precodidad y la variedad Super Precoz de periodo vegetativo corto se llamará precoz; esta última se utilizó como padre común en todos los cruzamien tos. 3.2.. METODOS. En el primer semestre de 1972 se efectuaron en el campo del. Ce~. tro Nacional de Inves tigaciones Agropecuarias "Tibai ta tA" (C. N. l. A. "Tibaitatá) los cruzamientos para obtener la semilla híbrida o de pri". •. •. mera generación F1; parte de esta semilla se dejó en reserva y el res•. to se sembró en el invernadero el siguiente semestre (1972 b) para obtener las plantas F l , algunas de. ~stas. se cruzaron con sus respectivos. padres (F l x PI) y (F l x P2 ) para obtener la semilla de los retrocruz.! mientos, la semilla del resto de plantas no cruzadas también se·'cosechó..
(37) 23. TABLA l. Número de plantas utilizadas en cada generación,. n~. bre de las variedades para cada cruzamiento y lugar de obtención de cada una.. Cruzamiento. Nombre de las variedades *. Generaciones y número de plantas BC 2 F2 BC l PI P2 Fl. 1. Bonza 63 (P~ x S~ per Precoz P2). 55. 48. 49. 136. 73. 56. 2. ICA-Sa~cá 68 (PI x Super Precoz (P 2) 49. 58. 49. 177. 42. 52. Icatá-I (PI) x Super Precoz (P2). 38. 60. 56. 129. 54. 63. Coconuco (Pf x Su36 per Precoz P2). 47. 51. 126. 32. 28. 3 4. *. Las variedades Bonza 63, lCA-Samacá 68, lcatá-l y Coconuco fueron obtenidas en Colombia y la variedad Super Precoz en México.. • •. •.
(38) 24. En el primer semestre de 1973 las generaciones Pl , Pz' Fl' F2' BC l , y BC2 de los cuatro cruzamientos fueron sembradas en el campo del Centro Nacional de Investigaciones Agropecuariss "Tibaitatá", en un diseno de parcelas divididas donde las parcelas principales fueron los cruzamientos y las subparce1as las generaciones. Se rea1izaron tres replicaciones, cada una formada de cuatro cruzamientos y cada cruzamiento seis generaciones. Las generaciones P1 , P2, Fl ,. BC l , y BC2 tenian dos surcos cada una y la F 2 cinco surcos, los surcos separados 30 centimetros entre si contentan 10 plantas cada uno, espaciadas 20 centímetros. En los extremos de cada surco y de cada replicaci6n se sembraron plantas de la variedad Bonza 63, igualmente donde se perdía una planta, 3.2.1. Datos tomados sobre cada planta en cada generaci6n, Epoca de espigamiento.- Esta se obtuvo contando los días de la fecha de siembra, 20 de Febrero hasta que el 50% de las espigas habían salido fuera de la hoja de bandera. Madurez fisio16gica.- Días a partir de Julio 20 cuando la parte. ". del pedúnculo donde se inserta la espiga empez6 a tomar un color ama-. •. •. rillento en el 50% de los tallos, con base en sugerencias del Programa de Fisiología Vegetal. Se utiliz6 una fecha arbitraria ya que no alteraría los análisis estadísticos,.
(39) 25 • •. •. ,. Rendimiento de grano.- Peso en gramos de los granos obtenidos. Peso de 50. granos.~. Peso en gramos de 50 granos tomados al azar. de cada planta, pesados en una balanza marca Mettler con precisi6n de centécimas de gramo. Macollamiento.- Número de tallos por planta con espigas fértiles. Longitud de espiga.- Longitud en centímetros de la espiga obtenida en el tallo más alto. 3.2.2. Promedios y varianzas, Los datos de las tres replicaciones en conjunto fueron utilizados para computar promedios y varianzas. 3.2.3. Heredabilidad en sentido amplio.La heredabilidad en sentido amplio(relaci6n de varianza genética total a varianza fenotípica), fue calculada para cada carácter estudia". ... do por el método de Mahmud y Kramer (1951). con la siguiente f6rmula: H =. 6 -V6 F Z. 2P I •. 6. P. 2. 3.2.4. Heredabilidad en sentido estrecho.. x. 100. L.
(40) 26. La heredabilidad en sentido estrecho (relaci6n de varianza ge-. ••. nética aditiva a varianza fenotípica). fue computada de acuerdo al método propuesto por Warner (1952). por medio de la siguiente f6rmula:. x. H :. 100. 3.2.5. Correlaciones fenotípicas. Las correlaciones fenotípicas en todas las combinaciones posibles los carácteres fueron calculadas de acuerdo al método de Weber y Moorthy (1952), con la fórmula: Covarianza xy Fenotípica. F2. r. xy. :2. ($YF 2. 3.2.6. Correlaciones genotípicas. Las correlaciones genotípicas con el método de Burton (1951). donde: Cov. xy. Cov. xy F. Genotípica. r. :. F 2. 1. xy. ". .. Donde. ~2 es la varianza de la correspondiente generaci6n y. Cov. xy es la covarianza de dos carácteres..
(41) 27 3.2.7. Modelo de Hayman. efe~. El modelo de Hayman (1958), se utilizó para determinar los. tos genéticos de cada carácter agronómico en-los cruzamientos Bonza 63 x Super Precoz,. Coconuco x Super Precoz. Se seleccionaron estos. y. cruzamientos por que el primero podría representar al grupo tardío x Precoz y el segundo al intermedio en precodidad x Precoz. De acuerdo a este modelo, los valores promedios esperados para cada generación con la notación de CambIe, citado por Bush et al. (1971), son los siguientes: PI =. ro. + aa - ad + t dd. ~d. + a -. P2 = m - a - l¡ d + aa + ad + i; dd Fl = m + ~ d F. 2. +. i; dd. =m. Be l = m+ ~ d + t aa l¡ a +. Donde m. = mide. t. aa. una constante; a, d, aa, ad y dd representan. efectos genéticos aditivos, dominantes, aditivo x aditivo, aditivo x dominante y dominante x dominante, respectivamente. En el modelo de tres parámetros genéticos de Hayman, la cons-. ... tante m y los efectos genéticos principales a y d son estimados por:. v-. 2VF =. B. +. (P. 1. +. + 2F ) + (V- + 2V- ) (V- F. p. 2. 1. p. F. B. 1. 2 2. 2 E. 2. + B. (B. V-. I. F 2. 2. ».
(42) 28. 2VB. ~* =. (P. - P ) 2. 1. (VP. +. + 4. V- (B - B ) P 1 - 2. v-. ). B. .. v-. 2VF. (2F. - P. 1. B. +. - P ) 2. 1. v- vF. 2. (2F. - P. ) +. - P. 1. 2. B. 2. v-. 2VF. F. 1. 'd'* = (B. +. B. - P. 2. 1. 1. - P ) 2. +. v- vP. 2. 1. (F. B. - F ) 2 1. +. v- v-. - B - B ) 2 1 1. (2F. F. P. 2. 2 E. Donde: P , P , F • F , B y B 1 VP, VF. 1. 2. 1. 2. v-. y v-. F. B. 1. son promedios de generaciones. 2. corresponden a las varianzas del error de las. 2 2. respectivas generaciones y 2E. = (V-. P. + 2V. + v-) +. ) (2V. B. F. F. 1. 2. v-. 8V. F. B 2. Los promedios esperados de generaciones estimados por m, a y de (modelo de tres parámetros, en el cual epístasis es omitida) son comparados con los valores observados por medio del uso del ji cuadrado, con 3 grados libres. Cuando en el modelo anterior el ajuste es satisfactorio (no significativa la prueba de ji cuadrado), el efecto de epis-·. ". ... tasis no es significativo y el modelo de 6 parámetros no es necesario. 3.2.8. Ganancia genética..
(43) 29. La ganancia genética fue calculada de acuerdo al método utilizado por Allard (1967), con la siguiente f6rmula:. G. = K. ~A.. H.. s. Donde K a. = diferencial. de selección, al cinco por ciento igual. 2,06. = Desviación estandar fenotípica de la generaci6n F 2 H. • •. •. = Heredabi1idad. en. ~entido. amplio. y. en sentido estrecho..
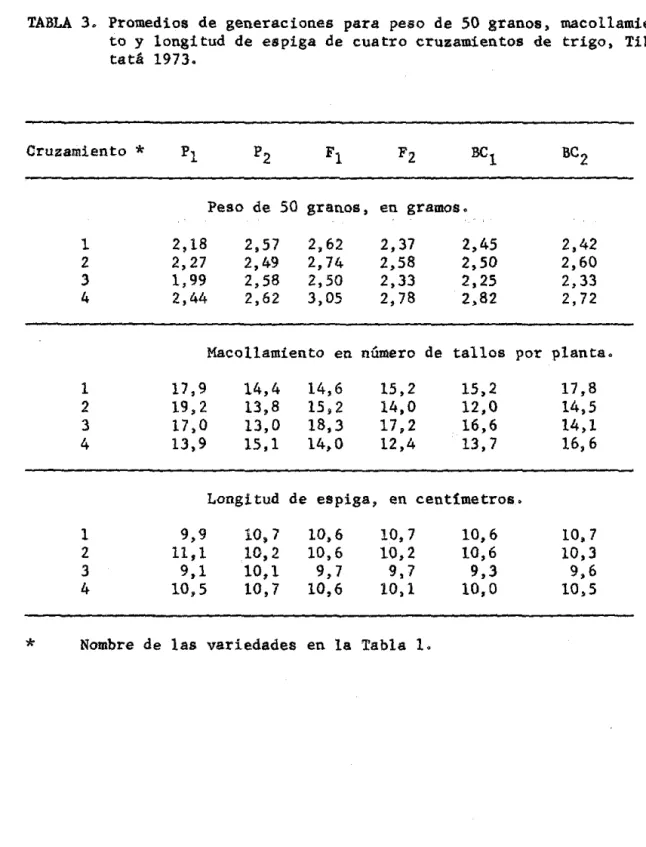
(44) 4.. RESULTADOS. • • •. 4.1.. COMPORTAMIENTO DE LOS PADRES Y GENERACIONES • En las tablas 2 y 3 se observa el promedio de generaciones para. cada uno de los carActeres época de espigamiento, madurez fisío16gica, rendimiento de grano, peso de 50 granos, maco11amiento por planta y longitud de espiga. Las variedades Bonza 63 e ICA-SamacA 68 fueron las más tardías· medidas en base a época de espigamiento y madurez fisio16gica, las variedades rcatá-l y Coconuco fueron intermedias, en precocidad y la más precoz fue la variedad Super Precoz. ICA-Samacá 68 fue la de mayor rendimiento de grano por planta, macollamiento y longitud de espiga. Bonza 63, lcatá-l y Coconuco tuvieron rendimientos similares pero inferiores a lca-SamacA 68, Super Precoz fue la de menor rendimiento. En Coconuco y Super Precoz de observ6 el mayor pe.so de 50 gra mos y menor maco1lamiento, en lcatA -1 el menor peso de 50 gramos y la menor longitud de espiga. Los valores de Fl para época de espigamiento y madurez fisio16gica fueron inferiores al promedio de los dos padres y los de F2. ... fueron superiores. Para rendimiento de grano por planta y peso. de. 50 gramos sucedi6 10 contrario, los valores de FI fueron superiores al promedio de padres y los de F inferiores y cercanos al promedio, 2.
(45) 3.1. ., ,. TABLA 2. Promedios de generaciones para épocas de espigamiento, madurez fisio16gica y rendimiento de granos por planta de cuatro cruzamientos de trigo, Tibaitatá 1973.. Cruzamiento 1-1. P. PI. 2. FI. F 2. BC. I. Be. Z. Epoca de espigamiento, en días por planta 1 2 3 4. 'o"~. 92,4 88,9 84,3 ' " 80,4. 75,4 75,5 75,3 72,8. 81,8 80,3 76,2 80,2. 90,7 87,2 85,8 80,8. 87,5 89,5 81,6 81,0. 90,3 83,1 82,4 77 ,4. Madurez fisiológica, en días por planta l 2 3. '~~. 'rJ. 21,8 14,6 8,6 8,4. 2,2 2,1 2,9 2,1 1". 8,6 6,7 2,4 6,5. 11,2 11,1 9,9 5,6. 10,4 13,5 6,7 5,2. *. **. 13,7 7,8 6,9 2,9. 1. Rendimiento de grano, en gramos por planta. 1 2. 3 4,. ... 32,85 39,66 32,24 32,87. 24,88 24,42 22,05 27,86. 31,02 36,11 36,77 34,55. 26,48 28,37 29,39 27,13. 26,12 22,41 29,81 28,91. *. Epoca de espigamiento a partir de Febrero 20.. **. Madurez fisiológica a partir de Julio 20.. 1-/. Nombre de las variedades en la Tabla 1.. 32,07 28,28 23,27 36,51.
(46) •• TABLA 3. Promedios de generaciones para peso de 50 granos, maco11amiea to y longitud de espiga de cuatro cruzamientos de trigo, Tibal tatá 1973.. Cruzamiento. *. P2. PI. FI. Be 1. BC 2. 2,45 2,50 2,25 2,82. 2,42 2,60 2,33 2,72. FZ. Peso de 50 granos, en gramos. 1 2 3. 4. 2,18 2,27 1,99 2,44. 2,57 2,49 2,58 2,62. 2,62 2,74 2,50 3,05. 2,37 2,58 2,33 2,78. Maco11amiento en número de tallos por planta. 1. 2 3. 4. 17,9 19,2 17,0 13,9. 14,4 13,8 13,0 15,1. 14,6 15,2 18,3 14,0. 15,2 14,0 17,2 12,4. 15,2 12,0 16,6 13,7. 17,8 14,5 14,1 16,6. Longitud de espiga, en centímetros. 1 2 3. 4. ;-.". • •. •. *. 9,9 11,1 9,1 10,5. 10,7 10,2 10,1 10,7. 10,6 10,6 9,7 10,6. 10,7 10,2 9,7 10,1. Nombre de las variedades en la Tabla l.. 10,6 10,6 9,3 10,0. 10,7 10,3 9,6 10,5.
(47) 33. respectivamente para rendimiento y peso del grano. Macollamiento por planta en las generaciones Fl y F2 fue inferior al promedio de los padres, con excepción del cruzamiento 3 (Icatá-l x Super Precoz) que. pr~. sentó un macollamiento superior en las dos generaciones. Longitud de espiga en las dos generaciones estuvo cerca al promedio de padres.. 4.2.. HETEROSIS. En la Tabla 4 se observa heterosis en porcentaje en base al pa-. dre superior y al promedio de los padres para cada uno de los. caráct~. res estudiados. Se encontró heterosis altamente significativa en base al padre más tardío y al promedio de los padres para época temprana de espigamiento y madurez fisiol6gica en tres cruzamientos con valores que variaron de -72 a -2,5 por ciento, y. heterosis en base al promedio de. los padres para época tardía de espigamiento en el cruzamiento 4 (Coconuco x Super Precoz) con un valor de 4,7 por ciento. Heterosis altamente significativa para rendimiento de grano por planta: solamente se encontr6 en el cruzamiento 3 sobre el promedio de los padres. Heterosis para peso de 50 granos altamente significativa sobre el promedio de padres 8e obtuvo en los custro cruzamientos y en base al padre superior en los cruzamientos 2 y 4 cuyos padres obtuvieron el mayor peso de 50 granos. En base al padre superior se observó heterosis para menor macollamiento en los cruzamientos 1 y 2; Y en el.
(48) •. ". TABLA 4. Heterosis observada en la generaci6n F1 dad~ en porcentaje en base al promedio del padf~ superior y al promedio de los padres para los carácteres estudiados en cuatro cruzamientos de trigo, Tibaitatá 1973.. Cruzamiento. Cruzamiento 1 _/ 1 Padre Superior. Promedio de padres. Padre Superior. Epoca de espigamiento. 88,S **. <)7~5. **. 90,3, **'. Madurez fisio16gica. 39,4 **. 71,7 **. 45,9 **. Rendimiento de grano. 94,4. Peso de 50 gr.! nos. 101,9. Mac011amiento. 81,6 **. Longitud de espiga. 99,1. * **. 1-/. = m ~. 107,5 110,3 ** 90,4 102,9. 91,0. .. Cruzamiento. 90,4 **. 95,S **. 99,8. 80,2 *. 27,9 **. 41,7 **. 77,4 * 123,8. 112,7. 79,2 **. 92,1. 95,5 *. 99,S. **. 4. Padre Promedio Superior de padres. 97,7. 115,1. Significativo al nivel del 5%. Significativo al nivel del 1%. Nombre de las variedades en la Tabla 1.. 3. Promedio Padre Promedio de padres Superior de padres. **. 110,0. Cruzamiento. 2. 104,7 **. 113,7. 135,1 **. 105,1. 96,9. 109,4 **. 116,4** 120,6 **. 107,6. 122,0 **. 92,7. 96,6. 101,0. 99,1. 100,0. 96,0 *. 113,8. '".!'-.
(49) 35. •. •. ,. •. cruzamiento 3 en base al promedio de padres, heterosis para mayor macE 11amiento a niveles altamente significativos. Longitud de espiga en los cruzamientos 2 y 3 en base al padre superior present6 heterosis signifi cativa para espiga corta.. 4.3.. VARIANZAS DE GENERACIONES PARA CADA CARACTER. Las Tablas 5 y 6 contienen las varianzas de generaciones para cada. carácter en cada uno de los cruzamientos. En general las varianzas de los padres Pl' de las poblaciones del padre P2 y de las Fl son. difere~. tes pero inferiores a las varianzas de las generaciones F2 en todos los carActeres estudiados. y BC para ~poca de es2 l pigamiento, madurez fisiológica y rendimiento de grano en la mayor1a de. Las varianzas de los retrocruzamientos BC. los cruzamientos fueron diferentes. Varianza de. ~poca. de espigamiento. varió de 21,50 a 79,98; en madurez fisio16gica de 10.09 a 42,20; en. re~. dimiento de grano de 110,19 a 181,44. Las varianzas para peso de 50 granos, macollam!ento y longitud de espiga fueron en general, similares en los dos retrocruzamientos; las variaciones para cada carácter respectivamente fueron: 0,069 a 0,094; 18,60 a 33,80 y 0,818 a 1,486.. •. •. Los cruzamientos 1 y 3 (Bonza 63 x Super Precoz e Icatá-l x Super Precoz) obtuvieron las mayores varianzas en la genersción F2 en compa-.
(50) '". TABLA 5. Varianzas de generaciones para época de espigamiento, madurez fisiológica y rendimiento de grano por planta de cuatro cruzamientos de trigo, Tibaitatá 1973.. Cruzamiento. *. PI. P. Fl. 2. F. 2. BC. BC. 1. 2. Epoca de espigamiento, varianzas. 1 2 3 4. 43,058 25,892 26,653 21,379. 34,548 30,079 24,496 26,013. 13,097 32,598 19,010 40,931. 71,788 36,148 28,486 49,920. 60,056 40,109 32,331 24,676. 79,988 30,323 21,507 44,765. Madurez fisiológica, varianzas. 1 2 3 4. 20,220 9,708 14,609 5,968. 10,022 17,958 2,642 18,413. 6,744 5,293 9,295 5,496. 32,424 29,909 32,714 20,370. 21,858 42,206 18,641 10,093. 41,569 14,734 22,080 10,883. Rendimiento de grano por planta, varianzas. 1 2 3 4. *. 127,880 200,882 142,163 86,256. 95,406 114,796 88,255 98,754. 104,418 192,920 147,263 163,475. Nombre de las variedades en la Tabla l.. 153,311 210,491 186,735 198,768. 110,197 134,838 172,859 181,449. 112,740 162,927 119,988 141,008. 1.<>. '".
(51) •. •. -,. TABLA 6. Varianzas de generaciones para peso de 50 granos, maco11amiento y longitud de espiga de cuatro cruzamientos de trigo, TibaitatA. 1973.. Cruzamiento. *. PI. P2. F1. BC 1. BC 2. 0,074 0,069 0,080 0,077. 0,074 0,062 0,093 0,094. F2. Peso de 50 granos, varianzas. 1 2 3 4. 0,051 0,026 0,026 0,026. 0,036 0,080 0,039 0,033. 0,023 0,057 0,036 0,068. 0,098 0,070 0,105 0,101. r r. I ,i. Maco11amiento, Varianzas. 1 2 3 4. 23,083 26,652 22,645 11,792. 22,970 20,308 17,642 15,575. 19,958 33,069 22,142 22,216. 37,524 37,725 46,807 25,848. 25,764 18,604 35,709 22,996. !. 33,809 29,113 32,091 22,543. Longitud de espiga, varianzas. 1 2 3 4. *. 0,970 1,093 0,654 0,700. 1,006 1,584 0,748 0,770. 0,951 1,156 0,618 1,003. 1,754 1,529 1,438 1,244. 1,163 1,486 0,920 1,128. 0,818 1,368 1,199 0,824. Nombre de las variedades en la Tabla 1.. .... U>.
(52) 38. ,. ración con las varianzas de los respectivos padres y de las Fl para los carActeres madurez fisiológica, rendimiento de grano por planta, peso de 50 granos, macollamiento y longitud de espiga. Los cruzamientos 1 y 4 obtuvieron los mayores valores en la F2 para época de espigamiento, mientras que las diferencias entre la generación F 2 y las respectivas generaciones homocigotas para este mismo carActer en los cruzamientos 2 y 3 fueron inferiores. En el cruzamiento 2 (ICA-SamacA 68 x Super Precoz) se observa que la diferencia al comparar la varianza de la generación F2 con la. varia~. zs de los respectivos padres y de las Fl es menor que la observada en los demAs cruzamientos para todos los caricteres excepto madurez fisio-. ,. lógica donde la diferencia es comparable a la que se observa en los cruzamientos 1 y 3. AdemAs en los carActeres peso de 50 granos y longitud de espiga el padre P 2 del cruzamiento 2 obtuvo una varianza ligeramente superior a la obtenida en la F2 •. 4.4.. ESTIMATIVOS DE HEREDABILIDAD. Estimativos de heredabilidad en sentido amplio y heredabilidad en. .. •. sentido estrecho en porcentaje, para los carActeres en cuatro. cruzamie~. tos de trigo se observan en la Tabla 7. Heredabilidad en sentido estrecho para época de espigamiento y madurez fisio16gica fue conSiderablemente baja en los cruzamientos 1 (Bonza 63 x Super Precoz) y 2 (ICA-Sa-.
(53) •. ,. •. ". TABLA 7. Estimativos de heredabi1idad en sentido amplio y heredabi1idad en sentido estrecho en porcentaje para los carActeres estudiados en cuatro cruzamientos de trigo, TibaitatA 1973.. Heredabi1idad en sentido amplio, en porcentaje Cruzamiento 1 2. 3 4. *. Epoca de espigamiento. Madurez Fisio16gica. Rendimiento por planta. 46,2 22,8 10,3 52,7. 63,9 76,0 64,3 71,8. 27,9 27,9 40,0 53,5. Peso de 50 granos 54,3 36,1 69,8 68,7. Macollamiento por planta. Longitud de espiga. 38,6 38,3 57,3 47,5. 43,6 13,9 51,3 40,9. Heredabilidad en sentido estrecho, en porcentaje 1 2 3 4. *. 4,9 5,2 11,0 60,9. 4,4 9,6 75,5 97,0. 54,5 58,5 43,0 37,7. 48,9 12,9 35,2 30,7. 41,2 73,5 55,1 23,8. 87,0 13,3 52,6 43,0. Nombre de las variedades en la Tabla l.. ..., >D.
(54) 40. ,. .. macA 68 x Super Precoz) alrededor del 6,0 por ciento en promedio de los dos cruzamientos; heredabilidad en sentido amplio fue mayor, 52 por ciento en promedio de los dos cruzamientos y los dos carActeres. Con excepción de época de espigamiento en el cruzamiento 3 que permaneció ligeramente constante, heredabilidad en sentido estrecho fue mayor que heredabilidad en sentido amplio en los cruzamientos 3 y. 4 para época de espigamiento y madurez fisiológica; los valores promedios de los dos cruzamientos y los dos carActeres fueron de 78 por. cie~. to para heredabilidad en sentido estrecho y de 63 por ciento para heredabilidad en sentido amplio. Rendimiento de grano por planta present6 una heredabilidad en seE tido estrecho que varió de 37,7 a 58,5 por ciento mayor que heredabilidad en sentido amplio que varió de 27,9 a 53,5 por ciento. La heredabilidad de peso de 50 granos en sentido estrecho vari6 de 12,9 a 48,9 por ciento y fue mayor en sentido amplio donde vari6 de 36,1 a 68,7 por ciento. Heredabilidad en sentido estrecho para macollamiento varió de 23,8 a 75,5 por ciento y en sentido amplio de 38,3 a 57,3 por ciento. Longi,. .. tud de espiga presentó heredabilidad similar en los dos sentidos, 43,2 por ciento en promedio, con excepción del cruzamiento 1 donde heredabi-. •. lidad en sentido estrecho fue de 87,0 por ciento y en sentido amplio fue de 43,6 por ciento..
(55) 41. En general, madurez fisio16gica presentó 108 mayores valores de heredabilidad al compararse con los demAs carActeres agron6micos.. 4.5.. CORRELACIONES FENOTIPICAS y GENOTIPICAS. Las Tablas 8 y 9 contienen las correlaciones fenotípicas y geno-. típicas de seis carActeres agronómicos en todas sus combinaciones posibles para cada uno de los cuatro cruzamientos. Epoca de espigamiento y madurez fisiológica presentaron correlaciones fenotípicas y genotípicas positivas y altamente significativas en los cuatro cruzamientos. Correlaciones genotípicas de época de e8pigamiento y madurez fisiológica con rendimiento fueron positivas, a excepción de época de espigamiento en el cruce 3 que fue negativa, es decir a mayor precocidad mayor rendimiento; estas correlaciones fueron altamente significativas a excepción de época de espigamiento en el cruzamiento 1 que fue significativa. Correlaciones genotípicas negativas y significantes se observaron también con peso de 50 granos en los cruzamientos 1 y 3 y positivas en los cruzamientos 2 y 4 con excepci6n de. .. •. época de espigamiento en el cruzamiento 2 que fue negativa. pero no significativa. Epoca de espigamiento y madurez fisio16gica estuvieron correlacionadas fenotipica y genotípicamente, y positiva y significativamente con macollamiento en los cruzamientos 1 y 3 a excepción de época de espiga-.
(56) 42. ,. .. TABLA 8. Correlaciones fenot1picas y geno ti picas de seis carácteres agr~ nómicos en todas sus combinaciones posibles, de los cruzamientos Bonza 63 x Super Precoz y ICA-Samacá 68 x Super Precoz, Tibaitatá 1973.. Bonza 63 x Super e. Carác teres Epoca de espigamento. ~recoz. Madurez Fisiol6gica. Rendimiento por planta. 0,721** (0,852)**. 0,081 (0,200)*. -0,096 0,312** (-0,200)* (0,565)**. 0,242** (0,337)**. 0,096 (0,703)**. 0,279** -0,109 (-0,328)**(0,969)**. 0,249* (0,606)**. Madurez fisiológica Rendimiento por planta. Peso de 50 granos. Macoll~. miento. Longitud de espiga. (),286** 0,787** (0,718)**(0,670)**. 0,100 (0,151). 0,096 (0,385)**. 0,051 (0,094). Peso de 50 granos Maco11amiento. 0,215* (0,152) lCA-Samacá 68 x Super Precoz. Carácteres Epoca de espigamiento Madurez fisiológica. .. ,. Madurez Fisiológica. Rendimiento Peso de por plsnta 50 granos. 0,635** (0,576)**. -0,158 -0,084 (1,528)** (-0,033) -0,126 (0,707)**. Rendimiento por planta Peso de 50 granos Ma'collamiento * **. Significativo al nivel del 5% Significativo al nivel del 1% Correlación genot1pica en parentesis.. Macoll~. miento. Longitu de espiga. -0,199** 0,292** (-0,785)**(10,751)**. 0,033 (0,823)**. -0,242** 0,251** (0,579)** (1,545)**. 0,310** (0,807 )**. 0,892 ** 0,405** (0,699)** (0,686)** 0,104 0,158 (-0,770)**(-0,905)** 0,303** (0,761)**.
(57) 43. .,. TABLA 9. Correlaciones fenotípicas y genotípicas de seis carActeres agron6micoB en todas sus combinaciones posibles, de los cruzamientos IcatA-l x Super Precoz y Coconuco x Super Precoz, TibaitatA 1973.. IcatA-l x Super Precoz CarActeres Epoca de espigamiento. Madurez fisi016gica 0,459** (0,662)**. Madurez fisic16gica. Rendimiento por planta. Peso de 50 granos. Macoll~. miento. Longitud de espiga. 0,016 (-0,386)**. -0,167 0,157 (-0,817)**(0,179)*. 0,073 (0,151). 0,150 (0,402)**. 0,229* -0,143 (-0,249)**(0,367)**. 0,112 (0,145). Rendimiento por planta. 0,280** 0,740** (0,025) (0,779)**. Peso de 50 granos. -0,155 (-0,488. Macollamiento. 0,474** (1,014)** 0,144 (0,097) 0,349** (0,400)**. Coconuco x Super Precoz CarActeres Epoca de espigamiento J!{adurez fisio16gica. .. •. Madurez fisio16gica 0,595** (0,597)**. Rendimiento por planta. ~ongitud. de. espiga. -0,058 (2,537)**. 0,130 -0,205* -0,129 (1,406)**(-1,509)** (-1,038)**. 0,012 (3.129)**. 0,259** -0,189* -0,100 (1,773)**(-1,636)** (-0,124). Rendimiento por planta Peso de 50 granos Macollamien to * **. Peso de Mac011~ 50 granos miento. Sign~ficativo al nivel del 5% Significativo al nivel del 1% Corre1aci6n genot1pica en paréntesis.. 0,473** (0,138). 0,760** (0,452)**. 0,182* (1,420)**. . 0,·349** (0,549)**. 0,147 (1,241)** 0,275** (0,477)**.
(58) •. •. 44. • miento en el cruzamiento 3 que fue fenotípicamente no significativa,. •. correlaciones negativas en los cruzamientos 2 y 4 a excepción de madu rez fisiológica que fue genéticamente positiva en el cruzamiento 2. Correlaciones altamente significativas y positivas de época de espigamiento y madurez fisiológica con longitud de espiga, se observaron en el cruzamiento 1 y 2, Y negativamente en el cruzamiento 4 con época de espigamiento. Rendimiento de grano por planta presentó correlaciones positivas fenotípicas y genotípicas altamente significativas con peso de 50. gr~. nos y macollamiento en el cruzamiento 1, y en los cruzamientos 2, 3,. ,. Y 4 con peso de 50 granos, macollamiento y longitud de espiga; la correlación genotipica con peso de 50 gramos en los cruzamientos 3 y 4 no fue signHicante. Peso de 50 granos presentó correlaciones positivas genotípicas y altamente significativas Con macollamiento en el cruzamiento 1 y,. macollamiento y longitud de espiga en el cruzamiento 4.. Correlacio-. nes negativas con macollamiento y longitud de espiga en el cruzamien. ... to 2 y con macollamiento en el cruzamiento 3. Macollamiento y longitud de espiga estuvieron correlacionados. •. fenotípica. y. genotípicamente a un nivel altamente significaÚvo en. los cruzamientos 2, 3 y 4. En general, los valores de correlación.
(59) 45. .' .. genotípica fueron mayoresqúif-los fenotípicos en todos los carAc teres •. • 4.6.. EFECTOS GENETICOS. En las Tablas 10 y 11 se obser"an los valores esperados de gener.!!. ciones para los carActeres época de espigamiento, madurez fisio16gica rendimiento de grano por planta, peso de 50 granos, macollamiento y. 10~. gitud de espiga de los cruzamientos de trigo Bonza 63 x Super Precoz (cruzamiento 1) y Coconuco x Super Precoz (cruzamiento 4), y el ajuste del ji cuadrado al modelo de tres parámetros y 108 tres parámetros genéticos, m constante, a efecto genético aditivo, d efecto genético. domina~. te.. ,. Al comparar los valores esperados de cada una de las generaciones para los carácteres en los cruzamientos 1. y 4 (Tablas 10 y 11) con los. valores observados (Tablas 2 y 3) se concluye que son muy similares y esto lo demuestra la prueba de Ji cuadrado <x2) no significante. El valor de X2 para una probabilidad del 0,01 y tres grados libres es de 11,34. En estas circunstancias las desviaciones de los promedios observados en las generaciones de sus promedios esperados sobre el modelo de tres pa-. ... rámetros podrían revelar que clases de epistasis estarían modificando algún promedio dado, pero todos los valores de X2 calculados son menores que los de las tablas y por lo tanto no existe efectos epistAticos •. • Todos los carActeres agronómicos estudiados en los cruzamientos 1.
(60) •. •. ,.. '\.. TABLA 10. Estimativos de efectos genéticos para los carActeres estudiados en los cruzamientos de trigo Bonza 63 x Super Precoz, ajuste del X2 al modelo de tres parámetros y los tres parámetros genéticos, Tibaitatá 1973.. Epoca de espigamiento. Madurez fisiológica. Rendimiento por planta. Crul;amiento 1. Cruzamiento 4. Cruzamiento 1. Cruzamiento 4. Crul!:smiento 1. Cruzamiento 4. P1. 93,67. 80,87. 20,41. 8,14. 29,62. 31,03. P2. 80,53. 73,32. 3,53. 1,99. 26,43. 26,92. F1. 84,05. 81,58. 9,31. 5,53. 29,39. 32,02. F2. 85,58. 79,34. 10,89. 5,30. 28,70. 30,75. Be 1. 88,86. 81,22. 14,83. 6,84. 29,50. 31,53. Be. 82,30. 77,45. 6,92. 3,76. 27,91. 29,97. 1,50. 0,05. 8,62. 0,78. 1,71. 2,41. m. 85,58. 79,34. 10,89. 5,30. 28,70. 30,75. a. 6,57. **. 3,78. **. 7,93. **. 3,07. **. 1,59. d. -3,04. **. 4,48. **. -3,15. **. 0,47. **. 1,36. Generación. 2. x2 Parámetros Genéticos. **. **. 1,55 2,55. Significativo al nivel del 1%. ""'". **.
(61) •. •. ~. '\,. TABLA 11. Estimativos de efectos genéticos para los carácteres estudiados en 10~ cruzamientos de trigo Bonza 63 x Super Precoz y Coconuco x Super Precoz, ajuste del X al modelo de tres parámetros y los tres parámetros genéticos, Tíbaitatá 1973.. Peso de 50 granos Generaci6n. Cruzamiento 1. Cruzamiento 4. Mac011amíento por planta Cruzamiento 1. Cruzamiento 4. Longitud de espiga Cruzamiento 1. Cruzamiento 4. PI. 2,18. 2,45. 16,29. 13,40. 10,71. 10,07. P2. 2,50. 2,60. 15,51. 15,52. 10,06. 10,73. F 1. 2,58. 3,04. 14,85. 13,32. 10,77. 10,30. F2. 2,46. 2,78. 15,62. 13,77. 10,58. 10,28. Be. 2,38. 2,74. 16,07. 13,35. 10,74. 10,12. Be Z. 2,54. 2,82. 15,18. 14,19. 10,41. 10,45. X2. 0,01. 0,01. 0,75. 0,60. 0,11. 0,03. m. 2,46. 2,78. 15,62. 13,78. 10,58. 10,29. a. -0,16. l. Parámetros Genéticos. d **. **. -0,08 **. 0,89 **. -0,83 **. 0,33 **. -0,33 **. 0,24 **. 0,51**. -1,55 **. -0,92 **. 0,38 **. -0,23 **. Significativo al nivel del 1%. .¡:.. '".
(62) 48. y 4 presentaron efectos. gen~ticos. aditivos y dominsntes altamente sig-. nificativos con excepción de rendimiento de grano por planta que solamente presentó efectos genéticos aditivos.. 4.7.. GANANCIA GENETICA. La tabla 12 contiene las ganancias genéticas esperadas en base a. heredabilidad en sentido amplio y a heredabilidad en sentido estrecho pars los carActeres estudiados en los cuatro cruzamientos. La ganancia genética para época de espigamiento en base a. ~reda. bilidad en sentido amplio vari6 de 1,1 a 8,1 días y para madurez fisiol6gica de 6,7 a 8,6 días. La ganancia en base a heredabilidad en sentido estrecho para espigamiento vari6 de 1,0 a 8,9 días y para madurez fi siol6gica de 0,5 a 9,0 días, a través de cuatro cruzamientos. En rendimiento de grano por planta se observ6 una ganancia genética en base a heredabilidad en sentido amplio menor que la observada en base a heredabi1idad en sentido estrecho en tres de los cruzamientos, mientras que en el cruzamiento 4 sucedió lo contrario; los estimativos a través de los cuatro cruzamientos variaron de 7,1 a 17,5 gramos por planta. Peso de. ... 50 granos presentó una ganancia en base a heredabilidad en sentido amplio mayor que en base a heredabilidad en sentido estrecho en todos los cruza-. •. mientos; los estimativos variaron de 0,10 a 0,46 gramos por 50 granos •.
(63) •. •. '. ",. .. -. •. '.. '.. TABLA 12. Ganancia genética esperada en base a heredabi1idad en sentido amplio y a heredabilidad en sentido estrecho para los carácteres estudiados en cuatro cruzamientos de trigo, Tibaitatá 1973.. Ganancia genética esperada (heredabilidad en sen.tido amplio) Cruzamiento. *. Epoca de espigamiento (días). Madurez fisiológica (días). Rendimiento por planta (gramos). Peso de 50 granos (gramos). Macollamiento por planta (# de tallos). Longitud de espiga (centímetros). 1. 8,1. 7,5. 7,1. 0,35. 4,8. 1,2. 2. 2,8. 8,6. 8,3. 0,19. 4,1f. 0,4. 3. 1,1. 7,6. 11,3. 0,46. 8,1. 1,3. 4. 7,7. 6,7. 15,5. 0,45. 5,0. 0,9. Ganancia genética esperada (heredabilidad en sentido estrecho). *. 1. 1,0. 0,5. 13,8. 0,32. 5,2. 2,4. 2. 1,0. 1,1. l7 ,5. 0,10. 9,3. 0,3. 3. 1,2. 8,9. 12,1. 0,23. 7,8. 1,3. 4. 8,9. 9,0. 10,9. 0,20. 2,5. 1,0. Nombre de las variedades en la Tabla l.. fe;.
(64) 50. .. .. La ganancia genética de macollamiento por planta en los cruzamientos 1 y 2 en base a heredabilidad en sentido amplio con un valor de 4,8 tallos por planta fue menor que en sentido estrecho con valores de 5,2 y 9,3 respectivamente para cada cruzamiento; lo contrario se observ6 en los cruzamientos 3 y 4. La ganancia de longitud de espiga fue similar en los dos sentidos en tres de los cruzamientos y en el cruzamiento 1 la ganancía fue mayor en el sentido estrecho; los valores estimados variaron de 0,3 a 2,4 centímetros en base a heredabilidad en sentido estrecho. El cruzamiento 3 (Icatá-l x Super Precoz), obtuvo los mayores valores de ganancia. gené.~~a. esperada en base a heredabilidad en sentido am-. plio y estrecho para la mayoría de los carácteres. Los cruzamientos 4 y 1 siguieron en orden de importancia. Más adelante se discutirán estos resultados ya que para un programa de selecci6n serían más ventajosos los cruzamientos 1 y 3 en su orden. Al considerar los cruzamientos. 1 y 2 como un grupo donde padrea ta! díos se cruzaron por un padre precoz y los cruzamientos 3 y 4 un grupo diferente, donde padres intermedios en precocidad •• cruzaron por un padre precoz, se observ6 que los promedios de época de espigamiento y madurez fisio16gica en las generaciones F , F , BC , y BC fueron mayores en los 2 l l 2 , .. cruzamientos 1 y 2 que en los cruzamientos 3 y 4;. las varianzas de los. retrocruzamientos y la F2 para los mismos carácteres en los cruzamientos. •. 1 y 2 en promedio, fueron mayores que en los cruzamientos 3 y 4, y la he-.
(65) 51. redabilidad en sentido estrecho de los cruzamientos 1 y 2 fue menor que. •. ,. ·. •. la obtenida en los cruzamientos 3 y 4 para los mismos carácteres de cocidado. pr~.
Figure



+7





Documento similar
dente: algunas decían que doña Leonor, "con muy grand rescelo e miedo que avía del rey don Pedro que nueva- mente regnaba, e de la reyna doña María, su madre del dicho rey,
Abstract: This paper reviews the dialogue and controversies between the paratexts of a corpus of collections of short novels –and romances– publi- shed from 1624 to 1637:
E Clamades andaua sienpre sobre el caua- 11o de madera, y en poco tienpo fue tan lexos, que el no sabia en donde estaña; pero el tomo muy gran esfuergo en si, y pensó yendo assi
6 Para la pervivencia de la tradición clásica y la mitología en la poesía machadiana, véase: Lasso de la Vega, José, “El mito clásico en la literatura española
d) que haya «identidad de órgano» (con identidad de Sala y Sección); e) que haya alteridad, es decir, que las sentencias aportadas sean de persona distinta a la recurrente, e) que
Ciaurriz quien, durante su primer arlo de estancia en Loyola 40 , catalogó sus fondos siguiendo la división previa a la que nos hemos referido; y si esta labor fue de
Las manifestaciones musicales y su organización institucional a lo largo de los siglos XVI al XVIII son aspectos poco conocidos de la cultura alicantina. Analizar el alcance y
Para recibir todos los números de referencia en un solo correo electrónico, es necesario que las solicitudes estén cumplimentadas y sean todos los datos válidos, incluido el
