The emerging role of matrix metalloproteases of the ADAM family in male germ cell apoptosis
14
0
0
Texto completo
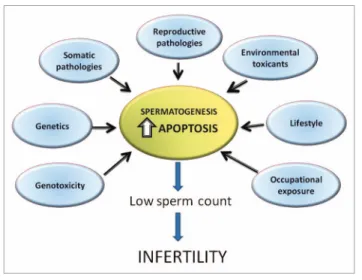
(2) Figure 1. Factors affecting germ cell apoptosis in spermatogenesis.. important to differentiate between apoptosis induction and apoptosis execution. Apoptosis initiation or induction can be defined as the molecular signaling events which trigger the extrinsic or intrinsic apoptotic pathways. In this way, events such as protein kinase activation, calcium signaling or oxidative stress could be regard at the initiation step of apoptosis. In this way, the activation of signaling cascades and their cross-talk produce activation of apoptotic and anti-apoptotic genes (as a defence mechanism). However, a cell with competence to undergo apoptosis is able to reverse this process thanks to the induction of anti-apoptotic genes until the cell passes the “point of no return” and the execution program is activated.31 Apoptosis execution involves the activation of caspases, a family of serine proteases with around 14 members, which are synthesized as inactive zymogens and become active upon specific death stimuli.32 The common strategy adopted to achieve apical caspase activation is the formation of complexes containing several caspase zymogens.33,34 In this way, apical caspase activation can be induced by the intrinsic (mitochondrial) pathway or by the extrinsic (cell death receptor) pathway. Several cellular mechanisms can be recognized in order to prevent and/or regulate caspase activation. Among these, we could mention: (1) cellular caspase inhibitor expression; (2) cell death receptor activation; (3) mitochondrial membrane permeability regulation by Bcl-2 proteins and (4) regulation of multiprotein caspase-containing protein complexes.29 Therefore, the final balance between pro-apoptotic and anti-apoptotic signals will define whether or not the cell will activate the executioner program of apoptosis. Evidence supporting the extrinsic pathway in germ apoptosis. The extrinsic pathway is characterized by the activation of death receptors such as Fas (CD95/Apo-1) or the tumor necrosis factor receptor 1 (TNFR1, also called p55 or CD120a), which have been shown to be expressed by germ cells in the mouse, rat and human testis.35,36 In mouse testes, additional death receptors such as TRAIL-R1 (DR4) are present in elongating spermatids and in Leydig cells, while TRAIL-R2 (also known as DR5, APO-2 and KILLER) is expressed predominantly by round and. 196. elongating spermatids and Leydig cells.37 In the human, the DR4 receptor is faintly detected in some myoid cells and intense immunoexpression has been detected in Sertoli cells, whereas strong expression is detected in spermatocytes, round and elongated spermatids. The DR5 receptor is highly expressed by spermatocytes, round and elongated spermatids, but not detected in Leydig or Sertoli cells. In the mammalian testis, death receptor ligands such as Trail (DR4 and DR5 ligand) have been shown to be expressed by all germ cells, as well as Sertoli and Leydig cells. On the other hand, Fas ligand (FasL) is expressed by Sertoli cells,38 although other reports have detected it in germ cells.39 Thus, ligand-induced germ cell apoptosis could be triggered in a juxta/paracrine or autocrine fashion upon ligand binding to its cognate receptor. Activation (trimerization) of death receptors in response to ligand binding induces the formation of a multimeric complex named the death-induced complex (DISC), which allows for the activation of procaspase-8 in mice along with caspase-10 in humans.35,40,41 Caspase-8 and/or 10 initiate apoptosis by inducing proteolytic processing of caspases-3, -6 and -7.42,43 Among these, caspase-3 is the main executioner caspase involved in fragmentation of the Golgi apparatus, nuclear lamina and DNA (due in part to proteolysis of a DNAse inhibitor) and the decrease in mitochondrial membrane potential.41,44,45 Evidence supporting the extrinsic pathway in physiological germ cell apoptosis during the first round of spermatogenesis comes from studies showing an increase in levels of the Fas receptor and activation of caspases -8, -3, -2 and -6 in apoptotic germ cells.46,47 In addition, pharmacological inhibition of caspase-8 reduces by more than 50% the number of apoptotic germ cells, suggesting an important role of the extrinsic pathway in this process. Nevertheless, these data do not rule out the involvement of other death receptors which are found in testes.36,37 However, male mice harbouring a spontaneous loss of function mutation in the Fas gene (lpr mice) or FasL (gld mice) are fertile with normal spermatogenesis, suggesting that the Fas/FasL pathway may be dispensable in germ cell apoptosis.48 Despite this, FasL mutant gld (generalized lymphoproliferative disease) mice that lack a functional Fas signaling pathway have a small but significant increase in testis weight and numbers of spermatid heads per testis, as compared with wild type mice. In addition, gld mice show a slightly higher spontaneous incidence of germ cell apoptosis than wild type mice, suggesting that under normal conditions, the Fas/FasL system could have a role in physiological (constitutive) germ cell apoptosis.49 In humans, altered meiotic and post-meiotic germ cell maturation might be associated with an upregulation of Fas gene expression and apoptosis.50,51 Besides physiological apoptosis, evidence suggests the involvement of the extrinsic apoptosis pathway after exposure to etoposide, ethanol,52 methoxychlor,53 GNRH antagonists,54 maternal and dietary selenium,55 ionising radiation,56,57 zearalenone,58 and testicular pathologies such as autoimmune orchitis,59 maturation arrest, Sertoli cell-only syndrome60 and cryptorchidism.61 However, contradictory evidence has been found with the environmental xenoestrogen mono-(2-ethylhexyl)phthalate (MEHP). Spermatogenesis. Volume 1 Issue 3.
(3) regarding the role of Fas/FasL.62,63 Therefore, the extrinsic apoptotic pathway is activated during germ cell apoptosis. Evidence supporting the intrinsic pathway in germ cell apoptosis. The intrinsic pathway is characterized by a decrease in mitochondrial membrane potential and the release of cytochrome c, which along with dATP, the cytosolic protein Apaf-1 and procaspase-9, assembles into a complex called the apoptosome. Within this complex, procaspase-9 becomes active and then cleaves caspase-3, connecting the intrinsic and the extrinsic pathways.33,34 Mitochondrial membrane permeabilization is carried out by proapoptotic proteins belonging to the B-cell lymphoma-2 (Bcl-2) family, which encompasses proteins that promote apoptosis, whereas other members prevent apoptosis.64 BCL-2, BCL-x and BCL-w are three anti-apoptotic proteins that interact with and repress the activity of pro-apoptotic proteins. Two general classes of proapoptotic family proteins exist: (1) those that share three homology regions (BH1, BH2 and BH3), which are termed multidomain proteins; and (2) those that share little sequence homology except for the conserved BH3 domain, also termed “BH3-only” proteins.64-67 These multidomain proteins are also known as the Bcl-2/Bax-family in honour of Bax (Bcl-2 antagonist X), the first Bcl-2 antagonist discovered by Korsmeyer. Other members of this group are Bcl-2 antagonist/killer (Bak) and Bcl-2 ovarian killer (BOK).68,69 Among the “BH3-only” group, we can find Bcl2 antagonist of cell death (BAD) and p53 upstream modulator (PUMA).70 It appears that the pro-apoptotic family members Bax and Bak are crucial for the induction of permeabilization of the outer mitochondrial membrane (OMM) and the subsequent release of apoptogenic molecules, such as cytochrome c and DIABLO (also known as SMAC), which leads to caspase-9 activation.32,71 Evidence suggesting the involvement of the intrinsic pathway during physiological apoptosis comes from studies showing upregulation of Bcl-2 proapoptotic proteins and activation of caspase-9. These data are reinforced by findings showing that ablation of apoptotic genes (Bax, Bim or Bik) or overexpression of anti-apoptotic genes (such as Bcl-2) are associated with infertility due to the arrest of spermatogenesis at the onset of meiosis.5,32,71-76 In addition to physiological apoptosis, evidence suggesting the involvement of the intrinsic apoptosis pathway has been shown after testicular heat stress,61 exposure to environmental toxicants such as bisphenol A77 or diethyl maleate,78 autoimmune orchitis79 and streptozotocin-induced diabetes.80 Therefore, there is clear evidence that apoptosis and not other cell death forms (i.e., necrosis or autophagy) play a major role in germ cell demise. ADAM Metalloproteases The family of metalloproteinases known as metalloproteases and disintegrins (ADAMs) has a central role in juxta/paracrine and autocrine signaling by controlling the ectodomain shedding of different ligands and receptors.81 A typical ADAM consists of an N-terminal signal peptide, followed by a prodomain, a metalloprotease domain, a disintegrin domain, a cysteine-rich region, an EGF-like domain, a transmembrane region and a cytoplasmic domain. Interestingly, only about 60% of ADAMs contain. www.landesbioscience.com. a functional catalytic domain (HExGHxxGXXHD).82-84 So far, about 20 ADAM genes have been discovered in the human and 37 in the mouse; to date, eight of the latter have been found only as predictions from the genomic sequence.84 ADAM metalloproteases are widely distributed throughout different tissues and at different stages of development. However, the expression pattern and function of specific isoforms are tightly regulated during development, cell differentiation, insults and pathological conditions. Proteolytically active ADAMs can shed or cleave a large array of substrates.85 The ADAM metalloprotease domain is preceded by a prodomain which is thought to work as an intramolecular chaperone controlling the folding of the catalytic domain. The prodomain (usually around 20 kDa) maintains the protease domain in an inactive state, and may also influence its intracellular trafficking.83,86,87 ADAM17, ADAM10 and ADAM9 prodomains are removed by a protein convertase (e.g., furin) in the secretory pathway. Curiously, ADAM8 and ADAM28 have been shown to undergo autocatalytic prodomain removal.88,89 In this way, ADAM proteins involved in fertilization are expressed during spermatogenesis as a full length form, and are not processed until the spermatozoa has reached the epididymis. There, they undergo removal of the prodomain and the metalloprotease domain, which suggests that they do not function as sheddases during spermatogenesis, sperm maturation and/or fertilization.90-92 This is in striking contrast to ADAM17 or ADAM10, which show prodomain processing during spermatogenesis and their activity is important for inducing germ cell apoptosis.93 On the other hand, the disintegrin and cysteine-rich domains exhibit adhesive properties in different tissues, and the important role of proteolytically inactive ADAMs in reproductive organs (such as ADAM2, 3, 12 and 23) suggest a relevant role of these adhesive properties in biological process such as fertilization. Studies from ADAM knockout mice have indicated that ADAM 8, 9, 12 and 15 are dispensable for normal development.84,94,95 However, these results do not counteract the idea that they may have a function in cell responses against external insults or pathological conditions. In particular, ablation of ADAM8, 9, 12 and 15 genes do not have any overt consequences in testis development and male fertility. On the contrary, ablation of ADAM10, 17 and 19 shows that they have essential roles during development, particularly during heart differentiation.96,97 In the next section, we will summarize the data concerning the localization and putative function (in some cases) of ADAM proteins in the male reproductive tract. ADAM metalloproteases in fertilization. ADAM metalloproteases deserve special attention in mammalian fertilization. At the cellular level, at least three steps are required to accomplish fertilization: (1) sperm adhesion to the egg zona pellucida; (2) sperm adhesion to the egg plasma membrane; and (3) spermegg fusion.98-103 Three ADAMs has been described to have an important role in fertilization: fertilin, a heterodimer formed by the subunits fertilin-α (ADAM1) and fertilin-β (ADAM2), and cyritestin (ADAM3). However, these are not unique molecules that act to promote sperm and egg membrane fusion, because mice with sperm lacking ADAM1 and ADAM2 display normal in vitro. Spermatogenesis. 197.
(4) fertilization.104 ADAM1, ADAM2 and ADAM3 proteins are expressed in the spermatozoa of different mammals.105 In particular, ADAM1 and ADAM2 are located in the posterior head of spermatozoa when they pass through the epididymis,106 and ADAM3 is located in the acrosomal domain of spermatids and mature sperm.92,107 ADAMs 1–3 are synthesized as precursors and then the pro- and metalloprotease domains are proteolytically removed during epididymal maturation, so mature ADAMs have their disintegrin domain at the N-terminal.106,107 Previous knockout mouse studies of ADAMs 1A, 1B, 2 and 3 showed that male mice with deletions in the genes for Adam1a, Adam2 or Adam3 are infertile due to sperm which are defective during the fertilization process.104,108 When ADAM1a, which is ER specific and not found in mature sperm, was knocked out, ADAM2 remained intact, but the sperm lost surface expression of ADAM3 and ZP-binding ability.109 This result clearly indicates that ADAM1b/ADAM2 (i.e., fertilin) is not essential for fertilization, at least in mice. ADAM24 may also play a role in fertilization because spermatozoa deficient in this protein show reduced fertility and many polyspermic embryos.110 The available evidence suggests that ADAM3 is a good candidate for spermegg interactions, at least in mice, since in humans it has been identified only as a pseudogene, suggesting that genes other than ADAM3 must be involved in this process.111 Therefore, the available data suggest that a complex interaction between ADAM1a, ADAM1b and ADAM3 (and probably other proteins), rather than unique molecules, is required to achieve proper fertilization. Knockout studies in mice have provided evidence of the complicated relationship among the ADAM1A, ADAM1B, ADAM2 and ADAM3 proteins. ADAM1 is absent and ADAM2 is dramatically reduced in Adam3-null mice.105 ADAM4 and ADAM6 are dramatically reduced in Adam2 and Adam3 knockout sperm, and premature processing of ADAM4 has been detected in ADAM3-null mice.90 In addition, it has been shown the presence of different macromolecular complexes between ADAM1A/ ADAM2, ADAM1B/ADAM2 and ADAM2/ADAM3, as well as ADAM6/ADAM2 and ADAM6/ADAM3 in testes and mature sperm. Moreover, during epididymal transit, ADAM7 is associated with membranous vesicles (epididymosomes). This protein is transferred to the sperm surface during epididymal transit in mouse and human, and is subsequently expressed on the entire head of acrosome-intact sperm and acrosome-reacted spermatozoa.112,113 In this way, ADAM7 becomes an integral plasma membrane protein in sperm. Moreover, ADAM7 forms a complex with calnexin, heat shock protein 5 and integral membrane protein 2B in lipid rafts during sperm capacitation.114 Thus, there is a complex interaction between the different ADAM proteins in mammalian spermatozoa and, so far, their precise roles in sperm maturation, capacitation and fertilization are still not fully understood. ADAM24 is located as a monomer in the equatorial region of the sperm surface in mouse.110 When the sperm pass through the epididymis, ADAM24 located on the sperm membrane is processed by removing the pre-domain.110 Thus, it is clear that ADAM proteins play a major role in fertilization, but this is not the only activity performed by these proteases in reproduction. In fact, ADAM proteins are also present in other reproductive. 198. organs, such as the prostate, the vas deferens and the seminal vesicle, suggesting they may also play a role in the development and/ or physiology of those tissues. In addition, the mammalian testis expresses other isoforms (i.e., ADAM10 or ADAM17) which have some important roles during spermatogenesis. Localization and distribution of ADAM proteins in the male reproductive tract. Localization of ADAM proteins in the testis. So far, many ADAM proteins have been described as being expressed in the testis, and a major group of them (ADAMs 2, 3, 5, 6, 21, 24, 25, 26, 29 and 30) are exclusively detected in this organ, as evaluated by RT-PCR (Figs. 2, 3 and Table 1). In this regard, ADAM1 and ADAM2 (also called fertilin-α and fertilin-β, respectively) were first discovered as a heterodimer on the sperm surface.115,116 They are found only in germ cells and their expression is developmentally regulated during spermatogenesis.117,118 ADAM3 is found in mammalian sperm and its mRNA is expressed in primary spermatocytes (pachytene and diplotene), secondary spermatocytes and in spermatids,92,109,117,119,120 but not in a Sertoli cell line (15P-1), which suggests that ADAM3 is only expressed in germ cells. ADAM5 mRNA is expressed in mouse testes in spermatocytes and spermatids (Fig. 3). In the testis, ADAM5 protein localizes at the sperm surface as a precursor which is processed during epididymal maturation.121 ADAM5 protein levels, similar to ADAM7 levels, are reduced in ADAM2 and ADAM3 null sperm. So, ADAM5 and ADAM7 expression depend upon ADAM2 and ADAM3 expression.121 mRNAs of ADAMs genes have been detected in testes of different species6, 20, 27, 29–31, 33 either in germ or Sertoli cells, and in some cases the expression pattern is available (ADAM24, 25, 26 and 34), but there is a lack of studies showing their function in spermatogenesis and/or fertilization (Fig. 3 and Table 1). ADAM19 is transiently expressed in the seminiferous cords of mouse testes as shown by immunohistochemistry and RT-PCR from 15.5–19.5 days post-coitus to two-day old animals.122 Interestingly, it seems that ADAM19 mRNA levels are reduced by follicle stimulating hormone (FSH) in a time-dependent manner.122 ADAM32 is principally expressed in mouse testes and epididymis. Its expression has been found during the meiotic prophase and in pachytene spermatocytes as a precursor protein. In the epididymis, ADAM32 is processed and localized at the sperm surface.112,117,121 Since ADAM32 is highly homologous to ADAM2 and ADAM3, it has a possible role in spermatogenesis and/or fertilization. Evidence for ADAM4, 6, 21, 24–27, 29, and 30 expression in Sertoli cells has been found in studies where these proteins have been detected in the Sertoli cell-derived cell line 15P-1.117 According to the experimental biochemical data, most of the ADAMs expressed in the testis are proteolytically inactive, since the prodomain and the metalloprotease domain are removed during epididymal transit. However, ADAM10, ADAM15 and ADAM17 are the only three ADAMs detected as precursors and in their catalytically active forms without the prodomain.93,123 We have proposed that catalytically active ADAM10 and ADAM17 could play a role in germ cell apoptosis (see below), but the role of active ADAM15 remains unknown.. Spermatogenesis. Volume 1 Issue 3.
(5) Figure 2. Schematic representation of ADAM metalloprotease distribution in the male reproductive tract. The figure shows a representation of the male reproductive tract; the boxes indicate the ADAM metalloproteases expressed in the respective organs.. Epididymis and vas deferens. Six ADAMs (4, 11, 15, 17, 23 and 31) have been detected in the vas deferens of mouse, rat, and human by RT-PCR (Table 1 and Fig. 2). The specific tissue localization and regulation of these ADAM proteins during development or pathological conditions have not yet been explored. The Adam7 gene is expressed in human, rat and monkey epididymis, with higher levels in the proximal caput than in the distal regions.112,113,124-127 At least in mouse and rat, its mRNA is found exclusively in the epididymis, and its expression strongly relies upon androgen levels.113,124 As stated above, ADAM7 is detected in the principal cells of the epididymis and is transferred to the spermatozoa. Biochemical studies have shown that no processing or modification of ADAM7 occurs in the epididymis, and this protein is found at the cell surface in mature spermatozoa. ADAM28 is expressed in mouse and rat almost exclusively in the caput of the epididymis.88,112 Testicular factors rather than androgens are required for ADAM28 gene expression, since in gonadectomized mice, ADAM28 expression disappeared; after castration, ADAM28 levels did not fully recover after treatment with dihydrotestosterone.112,124 Other ADAM proteins such as ADAM15, ADAM23, ADAM31 and ADAM32 have been found to be expressed, but not exclusively, in mouse epididymis, but their function and subcellular localization are unknown.. www.landesbioscience.com. These observations suggest that the transfer of ADAM7 to epididymal spermatozoa, and the expression dependence of ADAM5 and ADAM7 on the expression of ADAM2, ADAM3 and probably the other ADAMs expressed in the epididymis, could be related to the acquisition of the fertilization capability of sperm during maturation (Fig. 2). Prostate. ADAM9 has been detected in human prostate cancer, and its mRNA and protein levels are significantly upregulated under these conditions compared to normal tissue.128,129 Moreover, mouse prostate cancer in the context of an ADAM9 deficiency shows a well-differentiated tumour, whereas littermate controls expressing wild type ADAM9 had predominantly poorly differentiated, and in some cases significantly larger, tumours. Gain-of-function experiments have shown that ADAM9 overexpression induces epithelial hyperplasia, and later leads to prostatic intraepithelial neoplasia lesions.130,131 Interestingly ADAM9 has been found to bind integrin αvβ5 which suggest that this protein could have functions related to cell adhesion and migration during prostate development and/or cancer.132 ADAM15 at the mRNA and protein levels is significantly overexpressed in multiple types of adenocarcinoma, specifically during the metastatic progression of human prostate and breast cancer.133,134 Knockdown of ADAM15 decreases cell migration and adhesion of PC3 cells (an androgen-independent prostate cancer cell line) to specific extracellular matrix proteins, as well as. Spermatogenesis. 199.
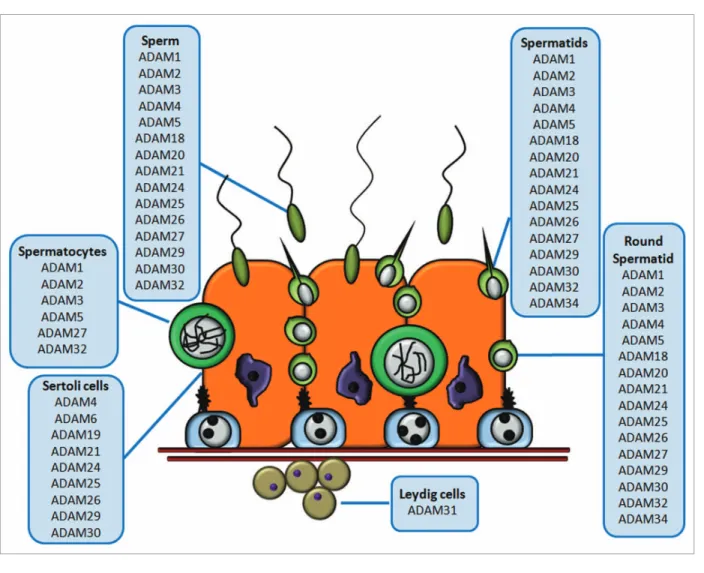
(6) Figure 3. Schematic representation of ADAM metalloprotease distribution in the seminiferous epithelium. The cartoon depicts a representation of the seminiferous epithelium, and the boxes indicate the ADAM metalloproteases expressed in different cell types. The figure shows ADAM metalloproteases with a specific cell type distribution; other ADAMs distributed throughout all cell types (e.g., ADAM10 or ADAM17) are not depicted in this scheme.. the cleavage of N-cadherin.134 This experimental evidence is consistent with the ability of ADAM15 to bind α5β1 and αvβ3 integrins.135,136 This experimental evidence suggests that ADAM15 may play a role in prostate tumour cell interactions and in the metastatic progression of human prostate cancer.134 ADAM10 and ADAM17 as Inducers of Germ Cell Apoptosis: A Working Model ADAM10 and ADAM17 are two widely expressed metalloproteases linked to the ectodomain release of not only ligands, but also receptors such as Notch, epidermal growth factor (EGFR), the p75 neurotrophin receptor (p75NTR), and c-kit.81,84,85 Despite the fact that ADAM1 was the first member of the ADAM family discovered in the testis, it was not until five years later that TACE (tumor necrosis factor-α converting enzyme) was recognized as a member of the metalloprotease family.137 Since then, TACE/ ADAM17 has been established as a key protease in mammalian development. Several studies have shown that the receptor tyrosine kinase c-kit is a substrate of ADAM17.138 The c-kit receptor. 200. is a 145 kDa glycosylated transmembrane protein, which upon binding to its ligand (SCF), initiates a signaling cascade leading to proliferation, differentiation, migration and survival.139-141 In studies with cultured stem cells derived from mastocytes and umbilical cord endothelial cells, release of the c-kit ectodomain has been found to be dependent on TACE/ADAM17 activity.140 Moreover, embryonic cells have shown a significant increase in apoptosis after being incubated with PMA, a phorbol ester known to activate PKC, but not in cells expressing a catalytically inactive mutant of TACE/ADAM17. This result suggests a strong link between TACE/ADAM17, processing of the c-kit ectodomain and activation of apoptosis. Our own data shows that in vivo PMA induces apoptosis in germ cells from 21-day old rats, which was prevented when a pharmacological inhibitor of ADAM17 was present at the time of PMA administration.93 ADAM17, but not ADAM10, is upregulated in germ cells undergoing physiological apoptosis and pharmacological inhibitors of ADAM17 are able to significantly prevent physiological apoptosis. Overall, these results uncover a new a role for ADAM17 as a key physiological regulator of germ cell apoptosis during. Spermatogenesis. Volume 1 Issue 3.
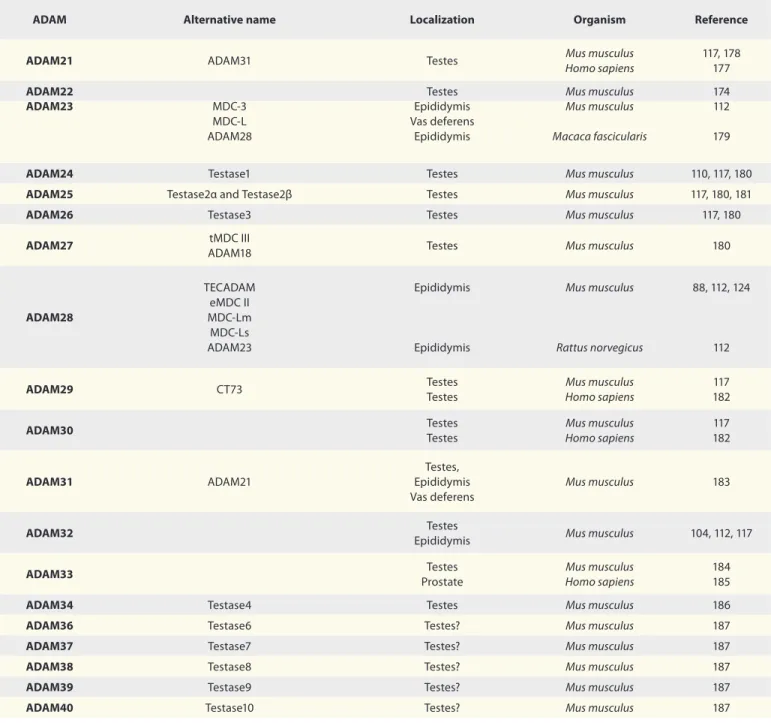
(7) Table 1. Distribution of ADAM metalloproteases in reproductive male tract ADAM. Alternative name. Localization. Organism. Reference. ADAM1. Fertilinα PH-30α. Testes Prostate. Mus musculus Rattus norvegicus. 120 125. ADAM2. Fertilinβ PH-30β. Testes. Mus musculus. 120. ADAM3. Cyritestin tMDC I. Testes. Mus musculus. 108, 117, 120. Mus musculus. ADAM4. tMDC V. Testes Vas deferens Testes Prostate. ADAM5. tMDC II. Testes Testes Testes. Mus musculus Macaca fascicularis Rattus norvegicus. 120 170 125. ADAM6. tMDC IV. Testes Testes Prostate Testes. Mus musculus Macaca fascicularis. 117 170. Rattus norvegicus. 125. ADAM7. Sperm maturation-related glycoprotein GP-83 EAP I. Epididymis Epididymis Epididymis Epididymis Epididymal Sperm. Homo sapiens Rattus norvegicus Macaca fascicularis Mus musculus. 126 112, 127 127 112, 124 113. ADAM8. MS2 CD156a Macrophage cysteine-rich glycoprotein. Prostate. Homo sapiens. 129. ADAM9. Cellular disintegrin-related protein Meltrin-gamma Myeloma cell metalloproteinase MDC9. Testes Testes Prostate Testes. Mus musculus Homo sapiens Rattus norvegicus. 171 171 128, 171, 172 157. Testes. Rattus norvegicus. 157. Prostate. Homo sapiens. 172, 173. CDw156 Kuzbanian protein homolog CD156c Myelin-associated metalloproteinase MADM. ADAM10. Rattus norvegicus. 117, 120 112 125. ADAM11. MDC. Vas deferens Testes. Mus musculus. 112 174. ADAM12. Meltrin-alpha. Prostate cancer. Mus musculus. 175. Prostate Testes Epididymis Vas deferens. Homo sapiens Mus musculus. ADAM15. Metalloprotease RGD disintegrin protein MDC-15 Metargidin AD56 CRII-7. 133, 134 123 112. TNFalpha-converting enzyme (TACE) CD156b. Testes Vas deferens Prostate. Rattus norvegicus. ADAM17. Homo sapiens. 93, 157 112 172. ADAM18. tMDC III. Testes Testes Testes. Rattus norvegicus Macaca fascicularis Homo sapiens. 125 170 176. ADAM19. Meltrin-beta MADDAM FKSG34 protein. Testes. Mus musculus. 122. Testes. Homo sapiens. 177. ADAM20. www.landesbioscience.com. Spermatogenesis. 201.
(8) Table 1. Distribution of ADAM metalloproteases in reproductive male tract (continued) ADAM. Alternative name. Localization. Organism. Reference. ADAM21. ADAM31. Testes. Mus musculus Homo sapiens. 117, 178 177. MDC-3 MDC-L ADAM28. Testes Epididymis Vas deferens Epididymis. Mus musculus Mus musculus. 174 112. Macaca fascicularis. 179. ADAM24. Testase1. Testes. Mus musculus. 110, 117, 180. ADAM25. Testase2α and Testase2β. Testes. Mus musculus. 117, 180, 181. ADAM26. Testase3. Testes. Mus musculus. 117, 180. ADAM27. tMDC III ADAM18. Testes. Mus musculus. 180. Epididymis. Mus musculus. 88, 112, 124. ADAM28. TECADAM eMDC II MDC-Lm MDC-Ls ADAM23. Epididymis. Rattus norvegicus. 112. Testes Testes. Mus musculus Homo sapiens. 117 182. Testes Testes. Mus musculus Homo sapiens. 117 182. Testes, Epididymis Vas deferens. Mus musculus. 183. ADAM32. Testes Epididymis. Mus musculus. 104, 112, 117. ADAM33. Testes Prostate. Mus musculus Homo sapiens. 184 185. ADAM22 ADAM23. ADAM29. CT73. ADAM30. ADAM31. ADAM21. ADAM34. Testase4. Testes. Mus musculus. 186. ADAM36. Testase6. Testes?. Mus musculus. 187. ADAM37. Testase7. Testes?. Mus musculus. 187. ADAM38. Testase8. Testes?. Mus musculus. 187. ADAM39. Testase9. Testes?. Mus musculus. 187. ADAM40. Testase10. Testes?. Mus musculus. 187. mammalian spermatogenesis. These data raise several questions such as: Which is/are the physiological substrate(s) shed by ADAM17 during germ cell apoptosis? How is ADAM17 activated under physiological conditions? Does ADAM17 participate in non-physiological germ cell apoptosis (induced by heat stress or genotoxins)? Is ADAM17 involved in germ cell apoptosis under pathological conditions? Here we would like to propose a model of germ cell apoptosis where shedding of the extracellular domain of c-kit by ADAM17 leads to germ cell apoptosis (Fig. 4). This proposal is based on our experimental evidence that germ cells undergoing physiological apoptosis lack the extracellular domain of c-kit.93 We propose that this event will turn off survival signaling, such as the PI3K/Akt pathway, which in turn will trigger activation. 202. of proapoptotic signals such as p53 and/or p73. The central role of c-kit in germ cell apoptosis comes from studies showing that blocking the binding of c-kit to its ligand (SCF) in Sertoli cells induces apoptosis.142 In addition, an inactivating point mutation at the intracellular domain of c-kit, which blocks binding of phosphoinositide-3-phosphate kinase (PI3K), induces apoptosis of germ cells and infertility.75,142,143 Moreover, genetic evidence in the mouse testis suggests that inactivation of c-kit signaling is linked to apoptosis mediated by the expression of Fas.144 In humans, associations between polymorphisms in the KIT gene and idiopathic infertility have also been found.145 Thus, inactivation of c-kit seems to promote apoptosis of mammalian germ cells during spermatogenesis, and probably leads to male infertility. Even though c-kit is a good candidate as the physiological. Spermatogenesis. Volume 1 Issue 3.
(9) Figure 4. A schematic drawing illustrating an emerging mechanism of action of ADAM metalloproteases in germ cell apoptosis. Active ADAMs metalloproteases cleave specific substrates at the cell surface of germ cells (ADAM17) or Sertoli cells (ADAM10). In this model, the activation of ADAM10 at the surface of the Sertoli cell would have a pro-survival effect by decreasing the half-life of FasL. On the other hand, activation of ADAM17 in the germ cell (spermatocytes and/or spermatogonia) would be by oxidative damage and/or DNA damage. Both insults would converge, activating the tyrosine kinase c-Abl, which in turn would activate p38 MAPK and/or PKC, promoting activation of ADAM17 and probably its transit to the cell surface. Thus, apoptosis induction is due to the inhibition of survival signals (extracellular c-kit domain shedding) and the induction of apoptotic signals (Fas receptor activation).. substrate of ADAM17, we cannot rule out that other survival receptors expressed by germ cells could also be processed by ADAM17 during germ cell apoptosis. Following c-kit extracellular domain shedding by ADAM17, we propose the stabilization/activation of p53 and/or p73, two well-known proteins involved in the transcriptional and posttranscriptional activation of many pro-apoptotic molecules.146-148 The role of p53 has been extensively studied in germ cells and in other apoptosis models. Even though experimental evidence has shown a role for p53 in physiological and genotoxin-induced apoptosis, mice lacking p53 show no major defects in spermatogenesis, which casts doubts on its central role during this process.24 Thus, there is a need to look for other related proteins which may fulfil the key role of p53 in apoptosis. p73 is a member of the p53 family which has been shown to have, like p53, the ability to induce apoptosis in different cell types. p73 is a transcription factor which shares amino acid homology and structural similarity with p53, especially within its DNA binding domain. Because of this, p73 share several target genes with p53 such as Bax, Puma and Fas.149,150 p73 is expressed in several isoforms, by alternative use of its promoter, generating TAp73, which contains the transactivation domain, or ΔNp73, which does not contain the transactivation domain.146,151 It has been show that, while the. www.landesbioscience.com. TAp73 isoforms can induce apoptosis, ΔNp73 isoforms act as anti-apoptotic factors. p73 transcriptional activity is modulated by the tyrosine kinase c-Abl and pharmacological inhibition of this enzyme with STI571 (STI) leads to the abolition of p73 activity.152-154 p53, p73 and c-Abl are expressed in germ cells, and we have shown that in vivo pharmacological inhibition of c-Abl prevents germ cell apoptosis induced by etoposide exposure.152,155 Our unpublished results show that STI prevents physiological apoptosis during the first wave of spermatogenesis, positioning p73 as a key transcriptional regulator of proapoptotic genes in germ cells (Codelia V, unpublished results). Moreover, p53 and p73 are highly expressed in apoptotic germ cells, which places these two master genes in a central position in the apoptotic initiation cascade (Fig. 4).47 As a complementary pathway, Fas could be activated by binding to membrane-bound FasL, which is expressed by Sertoli cells (Fig. 4). Experimental evidence has shown that FasL could be processed by ADAM10, but not by ADAM17.156 In this way, activation of ADAM10 could release the extracellular domain of FasL and prevent its binding to Fas in germ cells. This proposal came from experiments showing that only membrane-bound but not soluble FasL can induce apoptosis. Thus, caspase activation in germ cells will be the consequence of the combined action of the intrinsic and extrinsic pathways.. Spermatogenesis. 203.
(10) Figure 5. Pharmacological inhibition of p38MAPK prevents physiological and etoposide-induced germ cell apoptosis. (A) During the first round of spermatogenesis a maximum rate of germ cell apoptosis is observed in 25-day old rats (Control). In order to evaluate the role of p38 MAPK, different concentration of a pharmacological inhibitor (PD169316) were injected in the testes of 24-day-old rats and evaluated apoptosis 24 h after, when rats were 25 days old. PD169316 decreases the rate of physiological germ cell apoptosis in a concentration dependent manner. A high concentration of this inhibitor is toxic to germ cells. (B) Etoposide (100 μM) induces a significant increase in germ cell apoptosis when it is administered intratesticularly in 21-day old rats. However, when etoposide is administered along with 5 μM PD169316 (PD), germ cell apoptosis decreases significantly as compared to etoposide alone. *p < 0.05 n = 3.. Prodomain processing and cell surface exposure are two important events in ADAM activation. We have shown that, upon exposure to etoposide (an inhibitor of toposisomerase-II), ADAM10 and ADAM17 are upregulated at the cell surface, which correlates with germ cell apoptosis in 21-day old rats.157 Interestingly, neither ADAM10 nor ADAM17 were upregulated in testes subjected to heat stress (41°C for 15 min).15,157 Moreover, ADAM9, another metalloprotease involved in apoptosis after oxidative stress, is not upregulated by etoposide or heat stress, suggesting that upregulation of ADAM10 and ADAM17 is not a general process for all cell surface proteins. It remains controversial as to whether or not the shedding activity of ADAM17 and/or ADAM10 is regulated exclusively by prodomain release. In this way, p38 MAPK and protein kinase C (PKC) (via PMA) phosphorylates the intracellular domain of ADAM17.158,159 We have shown that after heat stress, p38 MAPK is activated in apoptotic germ cells,15 and our unpublished data indicates that a pharmacological inhibitor of p38 MAPK (PD169316) prevents physiological germ cell apoptosis in a concentration-dependent manner (Fig. 5). Additionally, PD169316 prevents germ cell apoptosis induced by etoposide, suggesting that p38 MAPK is involved both in physiological and genotoxicity-induced germ cell apoptosis. Interestingly, PKC and p38 MAPK can be phosphorylated (activated) by c-Abl, the same tyrosine kinase that targets p73. This kinase is present in germ cells and it has been shown to be highly expressed in primary spermatocytes; c-Abl-null mice are infertile, showing a disruption in spermatogenesis.160-162 Thus, we propose that, in germ cells, c-Abl could activate p38 MAPK and PKC, which in turn will phosphorylate ADAM17 and trigger its activation, probably by promoting its translocation to the cell surface and/or prodomain release (Fig. 4). At the cell surface, or during its transit to its final localization, ADAM17 could induce shedding of the extracellular domain of survival receptors (e.g., c-kit), which in turn will induce apoptosis.. 204. It has been shown that c-Abl as well as p73 and p53 are activated by oxidative stress and DNA damage.163 In germ cells, it has been proposed that abnormal recombination in spermatocytes might be a major source of DNA damage, which could activate c-Abl/p73 and/or p53. On the other hand, oxidative stress has not been assessed directly in apoptotic germ cells, but its relevance is known in several pathological conditions and after environmental toxicant (e.g., xenoestrogens) exposure.164 Although merely speculative, we would like to propose that oxidative stress and/or DNA damage can be sensed in germ cells by ataxia-telangiectasia mutated (ATM) and DNA-PK, two tyrosine kinases which phosphorylate and activate c-Abl in somatic cells.165,166 In somatic cells, oxidative stress induces posttranslational modifications and assembly of the DNA-PK complex on DNA along with activation of ATM.167 Therefore, it is not farfetched to propose that a similar mechanism may work in stressed germ cells (spermatocytes and spermatogonia), thereby inducing apoptosis. Our model may also apply to pathological (e.g., varicocele) and environmental situations (e.g., occupational toxin exposure) where oxidative damage has been shown to be an important contributor to inducing germ cell apoptosis.168,169 Interestingly, the environmental toxicant mono-(2-ethylhexyl) phthalate (MEHP) induces upregulation of matrix metalloproteases, including ADAM17.62 Therefore, the model of germ cell apoptosis proposed in this work may apply to many pathological conditions and environmental conditions experienced by the mammalian testis (Fig. 1). Concluding Remarks It is well known the importance of some ADAMs in fertilization (e.g., ADAM1-3), and it seems that they do not have a major role during spermatogenesis since gene ablation studies have shown no overt effects in testis histology. ADAMs have a complex gene expression pattern throughout the male reproductive tract and. Spermatogenesis. Volume 1 Issue 3.
(11) some of them are expressed in specific organs (e.g., only in testis), while others are found in different compartments (e.g., ADAM15 in testis and prostate), suggesting that their function may change according to the cellular context. In this review we have highlight recent work showing that ADAM17 and ADAM10 are expressed in mammalian testes and they participate regulating physiological and induced germ cell apoptosis. Our molecular model states that both cell autonomous and para/juxtacrine signaling events between germ-Sertoli cell are important in order to regulate References 1.. 2.. 3.. 4.. 5.. 6.. 7.. 8.. 9.. 10.. 11. 12.. 13.. Moreno RD, Lizama C, Urzua N, Vergara SP, Reyes JG. Caspase activation throughout the first wave of spermatogenesis in the rat. Cell Tissue Res 2006; 325:533-40; PMID:16598500; DOI:10.1007/s00441006-0186-4. Oakberg EF. A description of spermiogenesis in the mouse and its use in analysis of the cycle of the seminiferous epithelium and germ cell renewal. Am J Anat 1956; 99:391-413; PMID:13402725; DOI:10.1002/ aja.1000990303. Rodriguez I, Ody C, Araki K, Garcia I, Vassalli P. An early and massive wave of germinal cell apoptosis is required for the development of functional spermatogenesis. EMBO J 1997; 16:2262-70; PMID:9171341; DOI:10.1093/emboj/16.9.2262. Jahnukainen K, Chrysis D, Hou M, Parvinen M, Eksborg S, Soder O. Increased apoptosis occurring during the first wave of spermatogenesis is stage-specific and primarily affects midpachytene spermatocytes in the rat testis. Biol Reprod 2004; 70:290-6; PMID:14522836; DOI:10.1095/biolreprod.103.018390. Feng HL, Sandlow JI, Sparks AE, Sandra A, Zheng LJ. Decreased expression of the c-kit receptor is associated with increased apoptosis in subfertile human testes. Fertil Steril 1999; 71:85-9; PMID:9935121; DOI:10.1016/S0015-0282(98)00401-4. Huynh T, Mollard R, Trounson A. Selected genetic factors associated with male infertility. Hum Reprod Update 2002; 8:183-98; PMID:12099633; DOI:10.1093/humupd/8.2.183. Ji G, Gu A, Hu F, Wang S, Liang J, Xia Y, et al. Polymorphisms in cell death pathway genes are associated with altered sperm apoptosis and poor semen quality. Hum Reprod 2009; 24:2439-46; PMID:19542541; DOI:10.1093/humrep/dep223. Lee JY, Dada R, Sabanegh E, Carpi A, Agarwal A. Role of genetics in azoospermia. Urology 2011; 77:598-601; PMID:21195467; DOI:10.1016/j.urology.2010.10.001. Stouffs K, Lissens W, Verheyen G, Van Landuyt L, Goossens A, Tournaye H, et al. Expression pattern of the Y-linked PRY gene suggests a function in apoptosis but not in spermatogenesis. Mol Hum Reprod 2004; 10:15-21; PMID:14665702; DOI:10.1093/molehr/ gah010. Weikert S, Schrader M, Muller M, Krause H, Miller K. Expression of the apoptosis inhibitor survivin in testicular tissue of infertile patients. Int J Androl 2004; 27:161-5; PMID:15139971; DOI:10.1111/j.13652605.2004.00467.x. Sinawat S. The environmental impact on male fertility. J Med Assoc Thai 2000; 83:880-5; PMID:10998841. Jung A, Schuppe HC. Influence of genital heat stress on semen quality in humans. Andrologia 2007; 39:203-15; PMID:18076419; DOI:10.1111/j.14390272.2007.00794.x. Paul C, Melton DW, Saunders PT. Do heat stress and deficits in DNA repair pathways have a negative impact on male fertility? Mol Hum Reprod 2008; 14:1-8; PMID:18175790; DOI:10.1093/molehr/gam089.. www.landesbioscience.com. apoptosis. This model may stimulate new ideas and research avenues in order to elucidate the role of ADAMs in germ cell apoptosis, and probably the participation of other components in germ cell apoptosis. Acknowledgments. Part of this work was financed by a FONDECYT grant (1110778) to R.D.M.. 14. Bieber AM, Marcon L, Hales BF, Robaire B. Effects of chemotherapeutic agents for testicular cancer on the male rat reproductive system, spermatozoa and fertility. J Androl 2006; 27:189-200; PMID:16278370; DOI:10.2164/jandrol.05103. 15. Lizama C, Lagos CF, Lagos-Cabre R, Cantuarias L, Rivera F, Huenchunir P, et al. Calpain inhibitors prevent p38 MAPK activation and germ cell apoptosis after heat stress in pubertal rat testes. J Cell Physiol 2009; 221:296-305; PMID:19598167; DOI:10.1002/ jcp.21868. 16. Meachem SJ, Ruwanpura SM, Ziolkowski J, Ague JM, Skinner MK, Loveland KL. Developmentally distinct in vivo effects of FSH on proliferation and apoptosis during testis maturation. J Endocrinol 2005; 186:42946; PMID:16135663; DOI:10.1677/joe.1.06121. 17. Ortiz RJ, Lizama C, Codelia VA, Moreno RD. A molecular evaluation of germ cell death induced by etoposide in pubertal rat testes. Mol Hum Reprod 2009; 15:363-71; PMID:19346530; DOI:10.1093/molehr/ gap024. 18. Russell LB. Effects of male germ-cell stage on the frequency, nature and spectrum of induced specific-locus mutations in the mouse. Genetica 2004; 122:25-36; PMID:15619958; DOI:10.1007/s10709-004-1443-7. 19. Allemand I, Anglo A, Jeantet AY, Cerutti I, May E. Testicular wild-type p53 expression in transgenic mice induces spermiogenesis alterations ranging from differentiation defects to apoptosis. Oncogene 1999; 18:6521-30; PMID:10597255; DOI:10.1038/ sj.onc.1203052. 20. Beumer TL, Roepers-Gajadien HL, Gademan IS, van Buul PP, Gil-Gomez G, Rutgers DH, et al. The role of the tumor suppressor p53 in spermatogenesis. Cell Death Differ 1998; 5:669-77; PMID:10200522; DOI:10.1038/sj.cdd.4400396. 21. Elliott MR, Ravichandran KS. ELMO1 signaling in apoptotic germ cell clearance and spermatogenesis. Ann NY Acad Sci 2010; 1209:30-6; PMID:20958313; DOI:10.1111/j.1749-6632.2010.05764.x. 22. Honarpour N, Du C, Richardson JA, Hammer RE, Wang X, Herz J. Adult Apaf-1-deficient mice exhibit male infertility. Dev Biol 2000; 218:248-58; PMID:10656767; DOI:10.1006/dbio.1999.9585. 23. Knudson CM, Tung KS, Tourtellotte WG, Brown GA, Korsmeyer SJ. Bax-deficient mice with lymphoid hyperplasia and male germ cell death. Science 1995; 270:96-9; PMID:7569956; DOI:10.1126/science.270.5233.96. 24. Yin Y, Stahl BC, DeWolf WC, Morgentaler A. p53-mediated germ cell quality control in spermatogenesis. Dev Biol 1998; 204:165-71; PMID:9851850; DOI:10.1006/dbio.1998.9074. 25. Blottner S, Schon J, Roelants H. Apoptosis is not the cause of seasonal testicular involution in roe deer. Cell Tissue Res 2007; 327:615-24; PMID:17093922; DOI:10.1007/s00441-006-0328-8. 26. Comitato R, Esposito T, Cerbo G, Angelini F, Varriale B, Cardone A. Impairment of spermatogenesis and enhancement of testicular germ cell apoptosis induced by exogenous all-trans-retinoic acid in adult lizard Podarcis sicula. J Exp Zool A Comp Exp Biol 2006; 305:288-98; PMID:16432891.. Spermatogenesis. 27. Zhang L, Han XK, Qi YY, Liu Y, Chen QS. Seasonal effects on apoptosis and proliferation of germ cells in the testes of the Chinese soft-shelled turtle, Pelodiscus sinensis. Theriogenology 2008; 69:114858; PMID:18377973; DOI:10.1016/j.theriogenology.2008.01.028. 28. Kua CY, Kuida K. Cell death in mammalian development. In: Yin XM, Dong Z, Eds. Essential of Apoptosis: A guide for basic and Clinical Research. Totowa: Humana Press 2003. 29. Degterev A, Yuan J. Expansion and evolution of cell death programmes. Nat Rev Mol Cell Biol 2008; 9:378-90; PMID:18414491; DOI:10.1038/nrm2393. 30. Fadeel B, Ottosson A, Pervaiz S. Big wheel keeps on turning: apoptosome regulation and its role in chemoresistance. Cell Death Differ 2008; 15:443-52; PMID:17975549; DOI:10.1038/sj.cdd.4402265. 31. Tanuma SI. Molecular Mechanismis of Apoptosis. In: Sluyser M, Ed. Apoptosis in Normal Development and Cancer. Bristol: Taylor & Francis, Ltd. 1999; 39-61. 32. Riedl SJ, Shi Y. Molecular mechanisms of caspase regulation during apoptosis. Nat Rev Mol Cell Biol 2004; 5:897-907; PMID:15520809; DOI:10.1038/ nrm1496. 33. Shi Y. Mechanical aspects of apoptosome assembly. Curr Opin Cell Biol 2006; 18:677-84; PMID:17046227; DOI:10.1016/j.ceb.2006.09.006. 34. Shi Y. Apoptosome: the cellular engine for the activation of caspase-9. Structure 2002; 10:2858; PMID:12005427; DOI:10.1016/S09692126(02)00732-3. 35. Park HH. Structural analyses of death domains and their interactions. Apoptosis 2011; 16:209-20; PMID:21207148; DOI:10.1007/s10495-010-0571-z. 36. Pentikäinen V, Erkkila K, Suomalainen L, Otala M, Pentikainen MO, Parvinen M, et al. TNFalpha downregulates the Fas ligand and inhibits germ cell apoptosis in the human testis. J Clin Endocrinol Metab 2001; 86:4480-8; PMID:11549697; DOI:10.1210/ jc.86.9.4480. 37. Grataroli R, Vindrieux D, Gougeon A, Benahmed M. Expression of tumor necrosis factor-alpha-related apoptosis-inducing ligand and its receptors in rat testis during development. Biol Reprod 2002; 66:1707-15; PMID:12021051; DOI:10.1095/biolreprod66.6.1707. 38. Lee J, Richburg JH, Shipp EB, Meistrich ML, Boekelheide K. The Fas system, a regulator of testicular germ cell apoptosis, is differentially upregulated in Sertoli cell versus germ cell injury of the testis. Endocrinology 1999; 140:852-8; PMID:9927315; DOI:10.1210/en.140.2.852. 39. D’Alessio A, Riccioli A, Lauretti P, Padula F, Muciaccia B, De Cesaris P, et al. Testicular FasL is expressed by sperm cells. Proc Natl Acad Sci USA 2001; 98:331621; PMID:11248076; DOI:10.1073/pnas.051566098. 40. Henkler F, Behrle E, Dennehy KM, Wicovsky A, Peters N, Warnke C, et al. The extracellular domains of FasL and Fas are sufficient for the formation of supramolecular FasL-Fas clusters of high stability. J Cell Biol 2005; 168:1087-98; PMID:15795317; DOI:10.1083/ jcb.200501048. 41. Salvesen GS. Caspases: opening the boxes and interpreting the arrows. Cell Death Differ 2002; 9:3-5; PMID:11803369; DOI:10.1038/sj.cdd.4400963.. 205.
(12) 42. Scaffidi C, Fulda S, Srinivasan A, Friesen C, Li F, Tomaselli KJ, et al. Two CD95 (APO-1/Fas) signaling pathways. EMBO J 1998; 17:1675-87; PMID:9501089; DOI:10.1093/emboj/17.6.1675. 43. Scaffidi C, Schmitz I, Zha J, Korsmeyer SJ, Krammer PH, Peter ME. Differential modulation of apoptosis sensitivity in CD95 type I and type II cells. J Biol Chem 1999; 274:22532-8; PMID:10428830; DOI:10.1074/jbc.274.32.22532. 44. Kuribayashi K, Mayes PA, El-Deiry WS. What are caspases 3 and 7 doing upstream of the mitochondria? Cancer Biol Ther 2006; 5:763-5; PMID:16921264; DOI:10.4161/cbt.5.7.3228. 45. Sato T, Machida T, Takahashi S, Iyama S, Sato Y, Kuribayashi K, et al. Fas-mediated apoptosome formation is dependent on reactive oxygen species derived from mitochondrial permeability transition in Jurkat cells. J Immunol 2004; 173:285-96; PMID:15210786. 46. Codelia VA, Cisternas P, Moreno RD. Relevance of caspase activity during apoptosis in pubertal rat spermatogenesis. Mol Reprod Dev 2008; 75:881-9; PMID:17926353; DOI:10.1002/mrd.20822. 47. Lizama C, Alfaro I, Reyes JG, Moreno RD. Upregulation of CD95 (Apo-1/Fas) is associated with spermatocyte apoptosis during the first round of spermatogenesis in the rat. Apoptosis 2007; 12:499-512; PMID:17195944; DOI:10.1007/s10495-006-0012-1. 48. Lee J, Richburg JH, Younkin SC, Boekelheide K. The Fas system is a key regulator of germ cell apoptosis in the testis. Endocrinology 1997; 138:2081-8; PMID:9112408; DOI:10.1210/en.138.5.2081. 49. Richburg JH, Nanez A. Fas- or FasL-deficient mice display an increased sensitivity to nitrobenzene-induced testicular germ cell apoptosis. Toxicol Lett 2003; 139:1-10; PMID:12595153; DOI:10.1016/S03784274(02)00419-8. 50. Francavilla S, D’Abrizio P, Cordeschi G, Pelliccione F, Necozione S, Ulisse S, et al. Fas expression correlates with human germ cell degeneration in meiotic and post-meiotic arrest of spermatogenesis. Mol Hum Reprod 2002; 8:213-20; PMID:11870228; DOI:10.1093/molehr/8.3.213. 51. Francavilla S, D’Abrizio P, Rucci N, Silvano G, Properzi G, Straface E, et al. Fas and Fas ligand expression in fetal and adult human testis with normal or deranged spermatogenesis. J Clin Endocrinol Metab 2000; 85:2692-700; PMID:10946867; DOI:10.1210/ jc.85.8.2692. 52. Jana K, Jana N, De DK, Guha SK. Ethanol induces mouse spermatogenic cell apoptosis in vivo through overexpression of Fas/Fas-L, p53 and caspase-3 along with cytochrome c translocation and glutathione depletion. Mol Reprod Dev 2010; 77:820-33; PMID:20803734; DOI:10.1002/mrd.21227. 53. Vaithinathan S, Saradha B, Mathur PP. Methoxychlor induces apoptosis via mitochondria- and FasL-mediated pathways in adult rat testis. Chem Biol Interact 2010; 185:110-8; PMID:20227399; DOI:10.1016/j. cbi.2010.03.014. 54. Pareek TK, Joshi AR, Sanyal A, Dighe RR. Insights into male germ cell apoptosis due to depletion of gonadotropins caused by GnRH antagonists. Apoptosis 2007; 12:1085-100; PMID:17268770; DOI:10.1007/ s10495-006-0039-3. 55. Shi L, Yue W, Zhang C, Ren Y, Zhu X, Wang Q, et al. Effects of maternal and dietary selenium (Se-enriched yeast) on oxidative status in testis and apoptosis of germ cells during spermatogenesis of their offspring in goats. Anim Reprod Sci 2010; 119:212-8; PMID:20226605; DOI:10.1016/j.anireprosci.2010.02.012. 56. Coureuil M, Ugolin N, Tavernier M, Chevillard S, Barroca V, Fouchet P, et al. Puma and Trail/Dr5 pathways control radiation-induced apoptosis in distinct populations of testicular progenitors. PLoS ONE 2010; 5:12134; PMID:20711434; DOI:10.1371/journal. pone.0012134.. 206. 57. Forand A, Bernardino-Sgherri J. A critical role of PUMA in maintenance of genomic integrity of murine spermatogonial stem cell precursors after genotoxic stress. Cell Res 2009; 19:1018-30; PMID:19417777; DOI:10.1038/cr.2009.50. 58. Jee Y, Noh EM, Cho ES, Son HY. Involvement of the Fas and Fas ligand in testicular germ cell apoptosis by zearalenone in rat. J Vet Sci 2010; 11:115-9; PMID:20458151; DOI:10.4142/jvs.2010.11.2.115. 59. Suescun MO, Rival C, Theas MS, Calandra RS, Lustig L. Involvement of tumor necrosis factor-alpha in the pathogenesis of autoimmune orchitis in rats. Biol Reprod 2003; 68:2114-21; PMID:12606341; DOI:10.1095/biolreprod.102.011189. 60. Kim SK, Yoon YD, Park YS, Seo JT, Kim JH. Involvement of the Fas-Fas ligand system and active caspase-3 in abnormal apoptosis in human testes with maturation arrest and Sertoli cell-only syndrome. Fertil Steril 2007; 87:547-53; PMID:17123522; DOI:10.1016/j.fertnstert.2006.07.1524. 61. Yin Y, Stahl BC, DeWolf WC, Morgentaler A. p53 and Fas are sequential mechanisms of testicular germ cell apoptosis. J Androl 2002; 23:64-70; PMID:11780924. 62. Yao PL, Lin YC, Richburg JH. TNFalpha-mediated disruption of spermatogenesis in response to Sertoli cell injury in rodents is partially regulated by MMP2. Biol Reprod 2009; 80:581-9; PMID:19038859; DOI:10.1095/biolreprod.108.073122. 63. Lin YC, Yao PL, Richburg JH. FasL gene-deficient mice display a limited disruption in spermatogenesis and inhibition of mono-(2-ethylhexyl) phthalate-induced germ cell apoptosis. Toxicol Sci 2010; 114:335-45; PMID:20100735; DOI:10.1093/toxsci/kfq015. 64. Youle RJ, Strasser A. The BCL-2 protein family: opposing activities that mediate cell death. Nat Rev Mol Cell Biol 2008; 9:47-59; PMID:18097445; DOI:10.1038/ nrm2308. 65. Chen L, Willis SN, Wei A, Smith BJ, Fletcher JI, Hinds MG, et al. Differential targeting of prosurvival Bcl-2 proteins by their BH3-only ligands allows complementary apoptotic function. Mol Cell 2005; 17:393-403; PMID:15694340; DOI:10.1016/j.molcel.2004.12.030. 66. Ku B, Liang C, Jung JU, Oh BH. Evidence that inhibition of BAX activation by BCL-2 involves its tight and preferential interaction with the BH3 domain of BAX. Cell Res 2011; 21:627-41; PMID:21060336; DOI:10.1038/cr.2010.149. 67. Zhai D, Jin C, Huang Z, Satterthwait AC, Reed JC. Differential regulation of Bax and Bak by anti-apoptotic Bcl-2 family proteins Bcl-B and Mcl-1. J Biol Chem 2008; 283:9580-6; PMID:18178565; DOI:10.1074/ jbc.M708426200. 68. Hsu SY, Hsueh AJ. A splicing variant of the Bcl-2 member Bok with a truncated BH3 domain induces apoptosis but does not dimerize with antiapoptotic Bcl-2 proteins in vitro. J Biol Chem 1998; 273:3013946; PMID:9804769; DOI:10.1074/jbc.273.46.30139. 69. Hsu SY, Lin P, Hsueh AJ. BOD (Bcl-2-related ovarian death gene) is an ovarian BH3 domain-containing proapoptotic Bcl-2 protein capable of dimerization with diverse antiapoptotic Bcl-2 members. Mol Endocrinol 1998; 12:1432-40; DOI:10.1210/me.12.9.1432. 70. Villunger A, Michalak EM, Coultas L, Mullauer F, Bock G, Ausserlechner MJ, et al. p53- and druginduced apoptotic responses mediated by BH3-only proteins puma and noxa. Science 2003; 302:1036-8; PMID:14500851; DOI:10.1126/science.1090072. 71. Westphal D, Dewson G, Czabotar PE, Kluck RM. Molecular biology of Bax and Bak activation and action. Biochim Biophys Acta 2011; 1813:52131; PMID:21195116; DOI:10.1016/j.bbamcr.2010.12.019. 72. Coultas L, Bouillet P, Loveland KL, Meachem S, Perlman H, Adams JM, et al. Concomitant loss of proapoptotic BH3-only Bcl-2 antagonists Bik and Bim arrests spermatogenesis. EMBO J 2005; 24:3963-73; PMID:16270031; DOI:10.1038/sj.emboj.7600857.. Spermatogenesis. 73. Russell LD, Chiarini-Garcia H, Korsmeyer SJ, Knudson CM. Bax-dependent spermatogonia apoptosis is required for testicular development and spermatogenesis. Biol Reprod 2002; 66:950-8; PMID:11906913; DOI:10.1095/biolreprod66.4.950. 74. Yamamoto CM, Hikim AP, Lue Y, Portugal AM, Guo TB, Hsu SY, et al. Impairment of spermatogenesis in transgenic mice with selective overexpression of Bcl-2 in the somatic cells of the testis. J Androl 2001; 22:98191; PMID:11700863. 75. Yan W, Huang JX, Lax AS, Pelliniemi L, Salminen E, Poutanen M, et al. Overexpression of Bcl-W in the testis disrupts spermatogenesis: revelation of a role of BCL-W in male germ cell cycle control. Mol Endocrinol 2003; 17:1868-79; DOI:10.1210/ me.2002-0389. 76. Yan W, Suominen J, Samson M, Jegou B, Toppari J. Involvement of Bcl-2 family proteins in germ cell apoptosis during testicular development in the rat and pro-survival effect of stem cell factor on germ cells in vitro. Mol Cell Endocrinol 2000; 165:115-29; PMID:10940490; DOI:10.1016/S03037207(00)00257-4. 77. Li YJ, Song TB, Cai YY, Zhou JS, Song X, Zhao X, et al. Bisphenol A exposure induces apoptosis and upregulation of Fas/FasL and caspase-3 expression in the testes of mice. Toxicol Sci 2009; 108:427-36; PMID:19193734; DOI:10.1093/toxsci/kfp024. 78. Kalia S, Bansal MP. Diethyl maleate-induced oxidative stress leads to testicular germ cell apoptosis involving Bax and Bcl-2. J Biochem Mol Toxicol 2008; 22:37181; PMID:19110998; DOI:10.1002/jbt.20252. 79. Theas MS, Rival C, Jarazo-Dietrich S, Jacobo P, Guazzone VA, Lustig L. Tumour necrosis factor-alpha released by testicular macrophages induces apoptosis of germ cells in autoimmune orchitis. Hum Reprod 2008; 23:1865-72; PMID:18579514; DOI:10.1093/ humrep/den240. 80. Koh PO. Streptozotocin-induced diabetes increases apoptosis through JNK phosphorylation and Bax activation in rat testes. J Vet Med Sci 2007; 69:969-71; PMID:17917385; DOI:10.1292/jvms.69.969. 81. Blobel CP. ADAMs: key components in EGFR signaling and development. Nat Rev Mol Cell Biol 2005; 6:32-43; PMID:15688065; DOI:10.1038/nrm1548. 82. Seals DF, Courtneidge SA. The ADAMs family of metalloproteases: multidomain proteins with multiple functions. Genes Dev 2003; 17:7-30; PMID:12514095; DOI:10.1101/gad.1039703. 83. Takeda S. Three-dimensional domain architecture of the ADAM family proteinases. Semin Cell Dev Biol 2009; 20:146-52; PMID:18706512; DOI:10.1016/j. semcdb.2008.07.009. 84. White JM, Bridges L, DeSimone D, Tomezuk M, Wolfsberg TG. Introduction to the ADAM Family. In: Hooper NM, Lendeckel U, Eds. The ADASM family of Proteases. Dordrecht: Springer 2005; 1-29. 85. Huovila AP, Turner AJ, Pelto-Huikko M, Karkkainen I, Ortiz RM. Shedding light on ADAM metalloproteinases. Trends Biochem Sci 2005; 30:413-22; PMID:15949939; DOI:10.1016/j.tibs.2005.05.006. 86. Pruessmeyer J, Ludwig A. The good, the bad and the ugly substrates for ADAM10 and ADAM17 in brain pathology, inflammation and cancer. Semin Cell Dev Biol 2009; 20:164-74; PMID:18951988; DOI:10.1016/j.semcdb.2008.09.005. 87. Tousseyn T, Jorissen E, Reiss K, Hartmann D. (Make) stick and cut loose—disintegrin metalloproteases in development and disease. Birth Defects Res C Embryo Today 2006; 78:24-46; PMID:16622847; DOI:10.1002/bdrc.20066. 88. Howard L, Maciewicz RA, Blobel CP. Cloning and characterization of ADAM28: evidence for autocatalytic pro-domain removal and for cell surface localization of mature ADAM28. Biochem J 2000; 348:21-7; PMID:10794709; DOI:10.1042/02646021:3480021.. Volume 1 Issue 3.
(13) 89. Schlomann U, Wildeboer D, Webster A, Antropova O, Zeuschner D, Knight CG, et al. The metalloprotease disintegrin ADAM8. Processing by autocatalysis is required for proteolytic activity and cell adhesion. J Biol Chem 2002; 277:48210-9; PMID:12372841; DOI:10.1074/jbc.M203355200. 90. Han C, Choi E, Park I, Lee B, Jin S, Kim do H, Nishimura H, Cho C. Comprehensive analysis of reproductive ADAMs: relationship of ADAM4 and ADAM6 with an ADAM complex required for fertilization in mice. Biol Reprod 2009; 80:1001-8; PMID:19129510; DOI:10.1095/biolreprod.108.073700. 91. Blobel CP. Functional processing of fertilin: evidence for a critical role of proteolysis in sperm maturation and activation. Rev Reprod 2000; 5:75-83; PMID:10864851; DOI:10.1530/ror.0.0050075. 92. Linder B, Bammer S, Heinlein UA. Delayed translation and posttranslational processing of cyritestin, an integral transmembrane protein of the mouse acrosome. Exp Cell Res 1995; 221:66-72; PMID:7589257; DOI:10.1006/excr.1995.1353. 93. Lizama C, Rojas-Benitez D, Antonelli M, Ludwig A, Bustamante-Marin X, Brouwer-Visser J, et al. TACE/ ADAM17 is involved in germ cell apoptosis during rat spermatogenesis. Reproduction 2010; 140:305-17; PMID:20501791; DOI:10.1530/REP-10-0104. 94. Jacobsen J, Wewer UM. Targeting ADAM12 in human disease: head, body or tail? Curr Pharm Des 2009; 15:2300-10; PMID:19601832; DOI:10.2174/138161209788682389. 95. Kelly K, Hutchinson G, Nebenius-Oosthuizen D, Smith AJ, Bartsch JW, Horiuchi K, et al. Metalloproteasedisintegrin ADAM8: expression analysis and targeted deletion in mice. Dev Dyn 2005; 232:221-31; PMID:15580619; DOI:10.1002/dvdy.20221. 96. Deuss M, Reiss K, Hartmann D. Part-time alpha-secretases: the functional biology of ADAM 9, 10 and 17. Curr Alzheimer Res 2008; 5:187-201; PMID:18393804; DOI:10.2174/156720508783954686. 97. Sahin U, Weskamp G, Kelly K, Zhou HM, Higashiyama S, Peschon J, et al. Distinct roles for ADAM10 and ADAM17 in ectodomain shedding of six EGFR ligands. J Cell Biol 2004; 164:769-79; PMID:14993236; DOI:10.1083/jcb.200307137. 98. Evans JP. Sperm disintegrins, egg integrins and other cell adhesion molecules of mammalian gamete plasma membrane interactions. Front Biosci 1999; 4:114-31; PMID:9889181; DOI:10.2741/Evans. 99. Evans JP. Fertilin beta and other ADAMs as integrin ligands: insights into cell adhesion and fertilization. Bioessays 2001; 23:628-39; PMID:11462216; DOI:10.1002/bies.1088. 100. Miller DJ, Shi X, Burkin H. Molecular basis of mammalian gamete binding. Recent Prog Horm Res 2002; 57:37-73; PMID:12017553; DOI:10.1210/ rp.57.1.37. 101. Primakoff P, Myles DG. The ADAM gene family: surface proteins with adhesion and protease activity. Trends Genet 2000; 16:83-7; PMID:10652535; DOI:10.1016/S0168-9525(99)01926-5. 102. Rubinstein E, Ziyyat A, Wolf JP, Le Naour F, Boucheix C. The molecular players of sperm-egg fusion in mammals. Semin Cell Dev Biol 2006; 17:254-63; PMID:16574441; DOI:10.1016/j. semcdb.2006.02.012. 103. Talbot P, Shur BD, Myles DG. Cell adhesion and fertilization: steps in oocyte transport, sperm-zona pellucida interactions and sperm-egg fusion. Biol Reprod 2003; 68:1-9; PMID:12493688; DOI:10.1095/biolreprod.102.007856. 104. Kim E, Yamashita M, Nakanishi T, Park KE, Kimura M, Kashiwabara S, et al. Mouse sperm lacking ADAM1b/ADAM2 fertilin can fuse with the egg plasma membrane and effect fertilization. J Biol Chem 2006; 281:5634-9; PMID:16407235; DOI:10.1074/ jbc.M510558200. 105. Ikawa M, Inoue N, Benham AM, Okabe M. Fertilization: a sperm’s journey to and interaction with the oocyte. J Clin Invest 2010; 120:984-94; PMID:20364096; DOI:10.1172/JCI41585.. www.landesbioscience.com. 106. Hunnicutt GR, Koppel DE, Myles DG. Analysis of the process of localization of fertilin to the sperm posterior head plasma membrane domain during sperm maturation in the epididymis. Dev Biol 1997; 191:146-59; PMID:9356178; DOI:10.1006/dbio.1997.8700. 107. Kim E, Nishimura H, Iwase S, Yamagata K, Kashiwabara S, Baba T. Synthesis, processing and subcellular localization of mouse ADAM3 during spermatogenesis and epididymal sperm transport. J Reprod Dev 2004; 50:571-8; PMID:15514464; DOI:10.1262/jrd.50.571. 108. Nishimura H, Cho C, Branciforte DR, Myles DG, Primakoff P. Analysis of loss of adhesive function in sperm lacking cyritestin or fertilin beta. Dev Biol 2001; 233:204-13; PMID:11319869; DOI:10.1006/ dbio.2001.0166. 109. Nishimura H, Kim E, Nakanishi T, Baba T. Possible function of the ADAM1a/ADAM2 Fertilin complex in the appearance of ADAM3 on the sperm surface. J Biol Chem 2004; 279:34957-62; PMID:15194697; DOI:10.1074/jbc.M314249200. 110. Zhu GZ, Myles DG, Primakoff P. Testase 1 (ADAM 24) a plasma membrane-anchored sperm protease implicated in sperm function during epididymal maturation or fertilization. J Cell Sci 2001; 114:1787-94; PMID:11309208. 111. Grzmil P, Kim Y, Shamsadin R, Neesen J, Adham IM, Heinlein UA, et al. Human cyritestin genes (CYRN1 and CYRN2) are non-functional. Biochem J 2001; 357:551-6; PMID:11439107; DOI:10.1042/02646021:3570551. 112. Oh J, Woo JM, Choi E, Kim T, Cho BN, Park ZY, et al. Molecular, biochemical and cellular characterization of epididymal ADAMs, ADAM7 and ADAM28. Biochem Biophys Res Commun 2005; 331:1374-83; PMID:15883027; DOI:10.1016/j.bbrc.2005.04.067. 113. Oh JS, Han C, Cho C. ADAM7 is associated with epididymosomes and integrated into sperm plasma membrane. Mol Cells 2009; 28:441-6; PMID:19855936; DOI:10.1007/s10059-009-0140-x. 114. Han C, Park I, Lee B, Jin S, Choi H, Kwon JT, et al. Identification of heat shock protein 5, calnexin and integral membrane protein 2B as Adam7-interacting membrane proteins in mouse sperm. J Cell Physiol 2011; 226:1186-95; PMID:20945367; DOI:10.1002/ jcp.22444. 115. Blobel CP, Wolfsberg TG, Turck CW, Myles DG, Primakoff P, White JM. A potential fusion peptide and an integrin ligand domain in a protein active in sperm-egg fusion. Nature 1992; 356:248-52; PMID:1552944; DOI:10.1038/356248a0. 116. Wolfsberg TG, Bazan JF, Blobel CP, Myles DG, Primakoff P, White JM. The precursor region of a protein active in sperm-egg fusion contains a metalloprotease and a disintegrin domain: structural, functional and evolutionary implications. Proc Natl Acad Sci USA 1993; 90:10783-7; PMID:8248170; DOI:10.1073/ pnas.90.22.10783. 117. Choi I, Oh J, Cho BN, Ahnn J, Jung YK, Han Kim D, et al. Characterization and comparative genomic analysis of intronless Adams with testicular gene expression. Genomics 2004; 83:636-46; PMID:15028286; DOI:10.1016/j.ygeno.2003.10.001. 118. Wolfsberg TG, Primakoff P, Myles DG, White JM. ADAM, a novel family of membrane proteins containing A Disintegrin And Metalloprotease domain: multipotential functions in cell-cell and cell-matrix interactions. J Cell Biol 1995; 131:275-8; PMID:7593158; DOI:10.1083/jcb.131.2.275. 119. Muro Y, Okabe M. Mechanisms of fertilization—a view from the study of gene-manipulated mice. J Androl 2011; 32:218-25; PMID:20966425; DOI:10.2164/ jandrol.110.010900. 120. Wolfsberg TG, Straight PD, Gerena RL, Huovila AP, Primakoff P, Myles DG, et al. ADAM, a widely distributed and developmentally regulated gene family encoding membrane proteins with a disintegrin and metalloprotease domain. Dev Biol 1995; 169:378-83; PMID:7750654; DOI:10.1006/dbio.1995.1152.. Spermatogenesis. 121. Kim T, Oh J, Woo JM, Choi E, Im SH, Yoo YJ, et al. Expression and relationship of male reproductive ADAMs in mouse. Biol Reprod 2006; 74:744-50; PMID:16407499; DOI:10.1095/biolreprod.105.048892. 122. Bu SM, Yang YJ, Miao CL, Li HJ, Newcomer RG, Sang QX, et al. Developmental and hormonal regulation of meltrin beta (ADAM19) expression in mouse testes during embryonic and postnatal life. Life Sci 2006; 79:2112-8; PMID:16884740; DOI:10.1016/j. lfs.2006.07.002. 123. Pastén-Hidalgo K, Hernandez-Rivas R, Roa-Espitia AL, Sanchez-Gutierrez M, Martinez-Perez F, Monrroy AO, et al. Presence, processing and localization of mouse ADAM15 during sperm maturation and the role of its disintegrin domain during sperm-egg binding. Reproduction 2008; 136:41-51; PMID:18390692; DOI:10.1530/REP-07-0300. 124. Cornwall GA, Hsia N. ADAM7, a member of the ADAM (a disintegrin and metalloprotease) gene family is specifically expressed in the mouse anterior pituitary and epididymis. Endocrinology 1997; 138:4262-72; PMID:9322939; DOI:10.1210/en.138.10.4262. 125. Frayne J, Jury JA, Barker HL, Hall L. Rat MDC family of proteins: sequence analysis, tissue distribution and expression in prepubertal and adult rat testis. Mol Reprod Dev 1997; 48:15967; PMID:9291465; DOI:10.1002/(SICI)10982795(199710)48:2<159::AID-MRD3>3.0.CO;2-R. 126. Lin YC, Sun GH, Lee YM, Guo YW, Liu HW. Cloning and characterization of a complementary DNA encoding a human epididymis-associated disintegrin and metalloprotease 7 protein. Biol Reprod 2001; 65:944-50; PMID:11514362; DOI:10.1095/ biolreprod65.3.944. 127. Perry AC, Jones R, Barker PJ, Hall L. A mammalian epididymal protein with remarkable sequence similarity to snake venom haemorrhagic peptides. Biochem J 1992; 286:671-5; PMID:1417724. 128. Fritzsche FR, Jung M, Tolle A, Wild P, Hartmann A, Wassermann K, et al. ADAM9 expression is a significant and independent prognostic marker of PSA relapse in prostate cancer. Eur Urol 2008; 54:1097-106; PMID:18061337; DOI:10.1016/j.eururo.2007.11.034. 129. Fritzsche FR, Jung M, Xu C, Rabien A, Schicktanz H, Stephan C, et al. ADAM8 expression in prostate cancer is associated with parameters of unfavorable prognosis. Virchows Arch 2006; 449:628-36; PMID:17106710; DOI:10.1007/s00428-006-0315-1. 130. Peduto L. ADAM9 as a potential target molecule in cancer. Curr Pharm Des 2009; 15:2282-7; PMID:19601830; DOI:10.2174/138161209788682415. 131. Peduto L, Reuter VE, Shaffer DR, Scher HI, Blobel CP. Critical function for ADAM9 in mouse prostate cancer. Cancer Res 2005; 65:9312-9; PMID:16230393; DOI:10.1158/0008-5472.CAN-05-1063. 132. Mahimkar RM, Visaya O, Pollock AS, Lovett DH. The disintegrin domain of ADAM9: a ligand for multiple beta1 renal integrins. Biochem J 2005; 385:461-8; PMID:15361064; DOI:10.1042/BJ20041133. 133. Kuefer R, Day KC, Kleer CG, Sabel MS, Hofer MD, Varambally S, et al. ADAM15 disintegrin is associated with aggressive prostate and breast cancer disease. Neoplasia 2006; 8:319-29; PMID:16756724; DOI:10.1593/neo.05682. 134. Najy AJ, Day KC, Day ML. ADAM15 supports prostate cancer metastasis by modulating tumor cellendothelial cell interaction. Cancer Res 2008; 68:10929; PMID:18281484; DOI:10.1158/0008-5472.CAN07-2432. 135. Chen Q, Meng LH, Zhu CH, Lin LP, Lu H, Ding J. ADAM15 suppresses cell motility by driving integrin alpha5beta1 cell surface expression via Erk inactivation. Int J Biochem Cell Biol 2008; 40:2164-73; PMID:18387333; DOI:10.1016/j.biocel.2008.02.021. 136. Lu D, Chung KF, Xia M, Lu X, Scully M, Kakkar V. Integrin binding characteristics of the disintegrinlike domain of ADAM-15. Thromb Haemost 2006; 96:642-51; PMID:17080222.. 207.
Figure

+3


Documento similar