The humoral immune response induced by two recombinant bacillus Calmette Guérin vaccines promotes viral clearance against the human respiratory sincytial virus and the human metapneumovirus in mice
200
0
0
Texto completo
(2) 2. PONTIFICIA UNIVERSIDAD CATÓLICA DE CHILE Facultad de Ciencias Biológicas Programa Doctorado en Ciencias Biológicas Mención Genética Molecular y Microbiología. TESIS DOCTORAL: THE HUMORAL IMMUNE RESPONSE INDUCED BY TWO RECOMBINANT BACILLUS CALMETTE-GUÉRIN VACCINES PROMOTES VIRAL CLEARANCE AGAINST THE HUMAN RESPIRATORY SYNCYTIAL VIRUS AND THE HUMAN METAPNEUMOVIRUS IN MICE. Tesis entregada a la Pontificia Universidad Católica de Chile en cumplimiento parcial de los requisitos para optar al Grado de Doctor en Ciencias con mención en Genética Molecular y Microbiología. Por. JORGE ANDRÉS SOTO RAMÍREZ. Tutor:. Dr. Alexis M. Kalergis Parra. Thesis Committee:. Dra. Susan Bueno Ramírez Dr. Marcelo López Lastra Dr. Rafael Medina Agosto, 2018.
(3) 3. PONTIFICIA UNIVERSIDAD CATÓLICA DE CHILE Facultad de Ciencias Biológicas Programa Doctorado en Ciencias Biológicas Mencion Genética Molecular y Microbiología La defensa final de la tesis Doctoral titulada: THE HUMORAL IMMUNE RESPONSE INDUCED BY TWO RECOMBINANT BACILLUS CALMETTE-GUÉRIN (rBCG) PROMOTES THE VIRAL CLEARANCE AGAINST THE HUMAN RESPIRATORY SYNCYTIAL VIRUS (HRSV) AND HUMAN METAPNEUMOVIRUS (HMPV) IN MURINE MODEL. Presentada con fecha de hoy por el candidato a Doctor JORGE ANDRÉS SOTO RAMÍREZ Ha sido aprobada por el Tribunal Examinador, constituido por los profesores abajo firmantes, calificándose el trabajo realizado, el manuscrito sometido y la defensa oral con nota _______. Dr. Alexis Kalergis Parra Director de tesis Facultad de Ciencias Biológicas PUC. Dra. Susan Bueno Ramírez Miembro Comisión de Tesis Facultad de Ciencias Biológicas PUC. Dr. Marcelo López Lastra Miembro Comisión de Tesis Facultad de Ciencias Biológicas PUC. Dr. Rafael Medina Miembro Comisión de Tesis Facultad de Ciencias Biológicas PUC Agosto, 2018.
(4) 4. ACKNOWLEDGMENT. First of all, I would like to thank my family for supporting me from the first day I made the decision to enter the doctoral program. In particular, I would like to thank them for always pushing me to improve myself and raising new challenges both academic and personal, being mainly my parents, Ignacio Soto, Ana María Ramírez and my brothers Elizabeth and Victor who were really fundamental in this process. In addition, I would like to thank the Diaz Medina family who were also a very important part of this process, being not only a second family for me, but an important part of mine. In particular, I would like to thank in special to Nelson Díaz and his parents. On the other hand, I would like to thank my closest friends who always supported me and encouraged me to keep going and to the new friends who have been integrated into my life and those who have left for heaven delivering a painful but important life teachings for our future. Especially, I would like to thank my colleagues and close friends of the Biomedical Molecular Immunology laboratory who were fundamental in the training process. In particular, I would like to thank Nicolás Galvez that was my partner of work, Gisela Canedo, Natalia Muñoz, Emma Rey-Jurado, Evelyn Jara and Janyra Espinoza who are not only colleagues but became great friends. In addition, I would like to thanks members of the Dr. Bueno’s laboratory in special to Catalina Pardo and Irenice Coronado. Additionally, I would like to thank my thesis director, Dr. Alexis Kalergis Parra, who allowed me to access his laboratory to work on a topic that was always of my liking. I would also like to thank him for trusting in my abilities and always supporting me during this formative stage, allowing me to exploit my abilities as a leader. I would like to thank CONICYT (grant number 21151028) for the funding given during the development of my doctorate, as a scholarship it allowed me to focus completely on being able to perform quality science and also allowed me to attend various international conferences that were very important for my development as a professional. Finally, I would like to thank to the Millennium Institute of Immunology and Immunotherapy for the funding and the support granted throughout the development of this thesis..
(5) 5. INDEX ACKNOLEDMENT…………………………………………………………………..……4 FIGURE INDEX……………………………………………………………………............8 TABLE INDEX……………………………………………………………………………..9 ABREVIATIONS…………………………………………….……………………………10 RESUMEN………………………………………………….……………………………...12 ABSTRACT………………………………………………….………………...………......13 1. INTRODUCTION………………………………………….……………………………...14 1.1 General aspects of hRSV and hMPV……………….……………………….…………14 1.2 Viral Structure of hRSV and hMPV…………………..……………………………….16 1.3 Viral Cycle of hRSV……………………………………..…………………………….19 1.4 Viral Cycle of hMPV………………………………………..…………………………19 1.5 Immune response against the hRSV infection…………………..……………………..20 1.6 Immune response against the hMPV infection……………………..………………….22 1.7 Host immune response evasion mechanisms of hRSV and hMPV…………….………24 1.8 Use of Bacillus Calmette-Guérin as a vector………………………………..…………25 1.9 Vaccines and Treatments…………………………………………………..…………..26 2. HYPHOTESES STAMENT………………………………………………..……………..33 2.1 Hyphotesis 1……………………………………………………………...……………33 2.2 Hyphotesis 2…………………………………………………………………...………33 3. OBJECTS……………………………………………………………………………….….34 3.1 General Aims………………………………………………………………………….34 3.2 Specific Aims for the Hypotesis 1……………………………………………………..34 3.3 Specific Aims for the Hypotesis 2………………………..………………………...…34 4. CHAPTER 1: RECOMBINANT BCG VACCINES REDUCE PNEUMOVIRUS- CAUSED AIRWAY PATHOLOGY BY INDUCING PROTECTIVE CELLULAR AND HUMORAL IMMUNITY………………………………………………………………………...…………36 4.1 ABSTRACT……………………………………………….…………………………37 4.2 INTRODUCCION …………………………………..………………………………38 4.3 MATERIALS AND METHODS………………………..…………………………..41 4.3.1 hRSV and hMPV propagation and titration……………...………………………41 4.3.2 Doses of BCG-WT, rBCG-N and rBCG-P for immunization……...…………….42 4.3.3 Dot-blot assays……………………………………………….………………….42 4.3.4 Mouse immunization and viral infection…………………...……………………43 4.3.5 Quantification of IgG isotypes……………………………..…………………….44 4.3.6 Determination of Linked Recognition mechanism…………..…………….…….45.
(6) 6. 4.3.7 Serum neutralization assays………...………………………………………..…..45 4.3.8 Passive transfer of immune IgG from rBCG-vaccinated mice to naive animal......46 4.3.9 Evaluation of hRSV- and hMPV-associated disease parameters……….………..47 4.3.10 Statistical analyses……………………………………………………….……..47 4.4 RESULTS……………………………………………………………….…...………48 4.4.1 rBCG-N and rBCG-P vaccines are able to reduce disease…….Associated parameters caused by hRSV or hMPV infection…...………..………48 4.4.2 Inmunization with rBCG strains enhances post-challenge antibody ……. responses for several viral proteins …………………………...…………..…….51 4.4.3 T and B cell linked antigen recognition leads to diversification of the ……..antibody response to viral proteins …………..……………..…………………..54 4.4.4 rBCG vaccines induce on efficient antibody class switching upon viral infection.58 4.4.5 rBCG-induced antibodies show virus neutralizing capacity ex vivo……………..59 4.4.6 Antibodies triggered by rBCG vaccination followed by cirus ………..challenge protect naïve mice from viral-induced lung pathology……….………61 4.5 DISCUSSION…………………….……………………………………….…………65 4.6 CONFLICT OF INTEREST …………………………………………..………...….72 4.7 AUTHOR CONTRIBUTIONS ………………………………………..……………72 4.8 FUNDING………………………………….………………………………….….….72 4.9 ACKNOWLEDGMENTS………………………………………………….……….72 4.10 SUPLEMENTARY FIGURES…………………………………………..………...73 5. CHAPTER 2: EVALUATION OF THE IMMUNE RESPONSE INDUCED BY A LOW DOSE OF RBCG-P AGAINST THE HUMAN METAPNEUMOVIRUS IN MURINE MODEL………………………………………………………………….……………………76 5.1 ABSTRACT………………………………………….……………………….………77 5.2 INTRODUCTION……………………………………….…………………………...78 5.3 MATERIALS AND METHODS…...…………………..………….………………...81 5.3.1 hMPV propagation and titration……………………...…………………………..81 5.3.2 Doses of BCG-WT and rBCG-P for immunization………………………………81 5.3.3 Mouse immunization and viral infection………………………..…………..……82 5.3.4 Quantification of IgG isotypes and IgA measurement………...……………….…83 5.3.5 Splenocytes culture, T cells transfer and total igG transfer to naïve mice……...…84 5.3.6 Evaluation of hMPV-associated disease parameters………………………..…....85 5.3.7 Statisticals analyses………………………………………………………………86 5.4. RESULTS…………………………………………………………….…………..…87 5.4.1 A unique low dose of rBCG-P is able to decrease the diseases …….parameters in early and late times after hMPV-infection……………..……….…87.
(7) 7. 5.4.2 A low dose of rBCG-P clone 12 is able to promote the activation …… of macrophages eliciting a decrease in the inflammation of the lungs ……. in early and late times after hMPV-infection……………………..……………90 5.4.3 The recombinant BCGs strains induce the activation of T and B cells ……. on late-hMPV infection…………………...…………………………….………92 5.4.4 Both rBCG-P clones promote the secretion of mucosal ……and serum immunoglobulins ………………………………..……...……………93 5.4.5 The passive transfer of T cells and serum from rBCG-P clone 12 ……..immunized mice promote a protective role against an hMPV-infection……..… 96 5.5 DISCUSSION……………………………………………………….…….………..101 5.6 CONFLICT OF INTEREST……………………………...…………..……...……106 5.7 AUTHOR CONTRIBUTIONS……………………………………….…...………106 5.8 FUNDING……………………………………………………………..……………106 5.9 ACKNOWLEDGMENTS………………………………………….…………...…106 6. DISCUSION…………………………………………………………….......………….…107 7. CONCLUSION……………………………………………………………..……….....…114 8. REFERENCES…………………………………………………………………………...116 9. APENDIX…………………………………………………………………………………129 9.1 Scientific meetings attended during this thesis………………………………………129 9.2 Publications generated in this thesis and PhD training………………………………130.
(8) 8. FIGURE INDEX Figure 1. Total number of viral-associated respiratory infection cases per …………..epidemiological week in Chile……………..…………...……….………..………16 Figure 2. Genome and Protein Structure of hRSV and hMPV ……………….………………18 Figure 3. Evaluation of infection parameters from animals infected with hRSV or hMPV….50 Figure 4. Evaluation of the induction of IgG specific-antibodies against viral ………….antigens after a viral infection………………………………………...…….….53-54 Figure 5. The rBCG-N vaccine promotes the secretion of antibodies against ………….several antigens through the Linked Recognition mechanism………………….…57 Figure 6. The rBCG strains induce a strong IgG2a isotype switching after a viral infection …59 Figure 7. Sera from immunized mice are able to neutralize both viruses in vitro……………60 Figure 8. Passive immunity transfer from sera of previously rBCG immunized ………… mice protects naïve mice from hRSV- and hMPV-associated pathology……...….. 63 Figure 9. Evaluation of infection parameters from hMPV-infected animals at …………day 7post infection………………………………………………………..………..88 Figure 10. Evaluation of infection parameters from hMPV-infected animals at …………...day 28post infection………………………..…………………………..…………89 Figure 11. Evaluation of myeloid population in lung from hMPV-infected …………...animals at day 7 and 28 post-infection………………….……………...…………91 Figure 12. Evaluation of lymphoid population in lung from hMPV-infected …………...animals at day 7 and 28 post-infection……………………………..……..………93 Figure 13. Evaluation of the induction of IgG and IgA specific-antibodies ……………against viral antigens after a viral infection……………………...……...………..96 Figure 14. Passive immunity transfer from cell of previously rBCG immunized mice …………..decrease the hMPV-associated pathology in naïve mice transferred ……………...98 Figure 15. Passive immunity transfer from sera of previously rBCG immunized mice …………...protects in naïve mice transferred from hMPV-associated pathology …………..100 Figure 16. Proposed Model of operation of both rBCG-N and rBCG-P …………………...109. Supplementary Figure 1. Characterization of recombinant BCGs strains ……...…...………73 Supplementary Figure2. SchemeLinked Recognition …………………………………..…..73 Supplementary Figure 3. The rBCG-N strain promotes the Linked Recognition in ……………………………animals transferred with specific T cells –N hRSV. ………...…...74 Supplementary Figure 4. CD4+ CD69+ T cell populations after 7 and 14 dpi ………………75.
(9) 9. TABLE INDEX Table 1. Clinical Trials vaccines and therapies against the hRSV……………….…………...29 Table 2. Clinical Trials vaccines against the hMPV…...……………………….…………….30.
(10) 10. ABREVIATIONS hRSV: Human Respiratory Syncytial Virus hMPV: Human Metapneumovirus ALRTI: acute lower respiratory tract infections NS1: Non-structural 1 NS2: Non-structural 2 ssRNA: Single-strand RNA BCG: Bacillus Calmette- Guérin rBCG: Recombinant Bacillus Calmette- Guérin rBCG-P: Recombinant Bacillus Calmette- Guérin expressing the protein P rBCG-N: Recombinant Bacillus Calmette- Guérin expressing the protein N FI-hRSV: Human Respiratory Syncytial Virus inactivated with formalin IFN: Interferon IL: Interleukin DPI: Days post infection AECs: Air cells epithelial DC: Dendritic cell IM: Interstitial macrophages AM: Alveolar macrophage VLP: Virus-like particle BAL: Bronchoalveolar lavage TLR: Toll-like receptor TCR: T cell receptor.
(11) 11. MHC: Major Histocompatibility Complex pMHC: Peptide-loaded major histocompatibility complex FDA: Food and Drug Administration TSLP: Thymic Stromal Lymphopoietin LRTI: Lower respiratory tract infection IRF3: Interferon regulatory factor 3 Th1: T helper cells 1 Th2: T helper cells 2 cGMP: Good Manufacturing Practice conditions.
(12) 12. RESUMEN. El virus respiratorio sincicial humano (VRSh) y metapneumovirus humano (MPVh) son dos de los principales agentes etiológicos con mayor incidencia asociada a enfermedades respiratorias a nivel mundial, la incidencia de estos virus se ve incrementada durante las estaciones de otoño e invierno siendo principalmente VRSh el mayor causante de hospitalizaciones. Los principales reservorios de infección son recién nacidos, niños menores a dos años, personas inmunocomprometidas y ancianos. Los cuadros clínicos característicos asociados a la infección por estos virus son bronquitis y pneumonía. Sin embargo, se ha determinado que ambos virus podrían ser causantes de la aparición temprana de asma en niños. Además, se ha descrito que MPVh podría influenciar la aparición de enfermedades pulmonares obstructivas crónicas (EPOC). La patología asociada a la infección de estos virus es principalmente desencadenada por una respuesta inflamatoria caracterizada por un incremento en la secreción de citoquinas IL-4, IL-5, IL-8 acompañado de la producción de mucus en el epitelio pulmonar y a una contracción de las vías aéreas. Sumado a esto, la infección de ambos virus promueve a que la respuesta antiviral generada por la secreción de IFN de tipo I sea inhibida, esto, junto con una mala activación de los Linfocitos T, promueve un estado de anergia de estas células por parte de la infección mediada por VRSh desencadenado por la acumulación de proteína N del virus en la zona inducción de la sinapsis inmunológica, mientras que MPVh promueve que exista una baja activación de estas células asociado posiblemente a factores solubles, sin embargo, esto aún no ha sido comprobado. En la actualidad, no existen vacunas licenciadas para ninguno de estos virus. Sin embargo, algunos prototipos están siendo estudiados ya sea, en uso con animales como en pruebas en humanos. En nuestro laboratorio se generaron cepas recombinantes de la Mycobacteria Bacillus Calmette-Guérin (BCG) que expresan la proteína N-VRSh (rBCG-N) o la proteína P-MPVh (rBCG-P). Ambas vacunas fueron evaluadas en modelo animal encontrándose una respuesta inmune protectora a nivel celular mediado principalmente por la actividad de Linfocitos T. En este trabajo de tesis, se evaluó la respuesta inmune humoral inducida por estas vacunas y se logró determinar que ambas vacunas fueron capaces de promover la eliminación del agente viral, disminuyendo la inflamación pulmonar y parámetros asociados a infección. Además, se determinó que los anticuerpos secretados en respuesta a las vacunas tenían un perfil neutralizante y protector en modelo in vivo. Por otro lado, basados en la buena respuesta obtenida se decidió evaluar dos clones de la vacuna rBCG-P en condiciones de proceso de manufactura GMP y se evaluó el rol protector utilizando una dosis de inmunización menor a la usada anteriormente. Los resultados obtenidos permitieron concluir que la cepa rBCG-P clone 12 fue capaz de disminuir la inflamación pulmonar acompañada de una eficiente respuesta inmune humoral y celular vinculada a la activación de células T CD4+, CD8+, linfocitos B y una activa secreción de anticuerpos IgA e IgG. Finalmente, se determinó que la transferencia de anticuerpos y/o células provenientes de animales vacunados con esta cepa recombinante mostraron una eficiente eliminación del virus promoviéndose así una respuesta inmune protectora..
(13) 13. ABSTRACT. The human respiratory syncytial virus (hRSV) and human metapneumovirus (hMPV) are two of the main etiologic agents with the highest incidence associated with respiratory diseases worldwide. The incidence of these viruses is increased during the autumn and winter seasons, being mainly hRSV associated with causes of hospitalizations. The main infection reservoir of both viruses are newborns, children under two years old, immunocompromised people and the elderly. The clinical symptoms associated with infection by these viruses are bronchiolitis and pneumonia. However, it has been determined that both viruses could be responsible for the early onset of asthma in children. In addition, it has been described that hMPV could promote the appearance of chronic obstructive pulmonary diseases (COPD). The pathology associated with the infection of these viruses is mainly triggered by an inflammatory response characterized by an increase in the secretion of cytokines IL-4, IL-5, IL-8 accompanied by the mucus production in the pulmonary epithelium and a contraction of the airways. Added to this, the infection of both viruses, promotes an inefficient antiviral response triggering by the inhibition of type I IFN, as well as a bad activation of the T lymphocytes. hRSV promotes an anergic state in T cell triggered by the accumulation of N-hRSV protein in the zone when occurs the immunological synapse, while hMPV promotes a low activation of these cells possibly associated with soluble factors, however, it is unknown. At present, there are no licensed vaccines for any of these viruses. However, some prototypes have been studied in animals or in clinical trials. In our laboratory, a recombinant Bacillus Calmette-Guérin (rBCG) expressing NhRSV protein (rBCG-N) or the P-hMPV protein (rBCG-P) was generated. Both vaccines were evaluated in an animal model, finding a protective immune response at the cellular level mediated mainly by the activity of T lymphocytes. In this doctoral thesis, the humoral immune response induced by these vaccines was evaluated and it was determined that both vaccines were able to promote the elimination of the viral agent, decreasing pulmonary inflammation and parameters associated with infection. Furthermore, it was determined that the antibodies secreted in response to the vaccines had a neutralizing capacity and a protective role in vivo. On the other hand, based on the good response obtained with the vaccines previously mentioned, it was decided to evaluate two clones of the rBCG-P vaccine under conditions of the GMP manufacturing process and the protective role was evaluated using a lower immunization dose than previously used. The results obtained allowed us to conclude that the strain rBCG-P clone 12 was able to decrease the lung inflammation triggering an efficient humoral and cellular immune response linked to the activation of CD4+, CD8+ T cells, B lymphocytes and an active secretion of IgA and IgG antibodies. Finally, we determinate that the transfer of antibodies or T cells from rBCG-P clone 12 immunized mice to naïve receptor mice showed an efficient elimination of the virus with a protective immune response..
(14) 14. 1. INTRODUCTION. 1.1. General aspects of hRSV and hMPV. Human Respiratory Syncytial Virus (hRSV) and human Metapneumovirus (hMPV) are two respiratory viruses that infect at several growth stages in humans. In addition, both viruses are the main causal agents of acute lower respiratory tract infections (ALRTI) and hospitalizations during the winter months (Kahn, 2006; Nair et al., 2010). Risk factors to develop these viral infections include newborns, premature children, elderly and immunocompromised people (Garcia et al., 2010; Kahn, 2006; Lüsebrink et al., 2010; Nair et al., 2010; Schildgen et al., 2011; Van Drunen Littel-Van Den Hurk & Watkiss, 2012; Zeng et al., 2011). The main hRSV and hMPV-associated symptoms include cough, wheezing, fever, airways obstruction and bronchiolitis (Feuillet, Lina, Rosa-Calatrava, & Boivin, 2012; Hacking & Hull, 2002; Hall, Simőes, & Anderson, 2013). However, despite both viruses have similar symptomatology it is known that hRSV affects mainly the lower respiratory airways while hMPV affects the upper and lower respiratory airway, respectively (Boivin et al., 2002; Domachowske & Rosenberg, 1999; John V Williams et al., 2006). HRSV was first isolated and identified in 1956 from a colony of chimpanzees, however, in 1957 an identical pathogenic agent was isolated from children that presented strong respiratory damage, confirming that this viral agent was the same as that previously found in chimpanzees (Chanock, Roizman, & Myers, 1957). Alternatively, hMPV was first identified in 2001 from young Dutch children with respiratory symptoms and illness (B G van den Hoogen et al., 2001), but some retrospective reports confirm that many cases without identifying the.
(15) 15. pathogen agent involved in the illness was mainly associated with hMPV-infection (van den Hoogen et al., 2001; Crowe, 2004). In the case of hRSV, a retrospective reports suggest that hRSV-infection was the responsible for almost 22% of the ALTRI associated with 33.8 million of children under five years with this clinical diagnostic in 2005 worldwide (Nair et al., 2010). In addition, the hRSV infection has been associated with a rate of hospitalization from 66,000 to 199,000 with high mortality in children under 1 year mainly associated with developing countries (Iwane, Farnon, & Gerber, 2013; Nair et al., 2010). On the other hand, it has been estimated that around 6% of the respiratory illness in children younger than 3 years are associated to hMPV, being considered one of the most important viruses among all respiratory illness-associated viruses causing young children hospitalizations (Boivin et al., 2003). It has also been reported hMPV-associated hospitalizations at a rate of 1 per 1000 children minor 5 years and 3 per 1000 infants minor of 6 months. In addition, the prevalence of hMPV infection has been reported to be similar to influenza virus and types 1 to 3 parainfluenza virus (Edwards et al., 2013a). In Chile, the viral infections are reported mainly in the autumn and winter season. Furthermore, the main viral agent associated with the collapse of the public health system is hRSV being responsible of the 70% to 80% of hospitalizations in the winter months (Palomino, Larrañaga, Villagra, Camacho, & Avendaño, 2004). The Figure 1 shows the incidence of viral infections in the different weeks of the year in a period of 4 years where hRSV is associated with the greatest number of infection cases while hMPV shows a minor number of infection cases. Up to date, out of the 800 positives cases in 2018, the 50.5% has been associated with hRSV while the 6.6% were positive for hMPV (Respiratory virus surveillance report 2018. Institute of Public Health of Chile)..
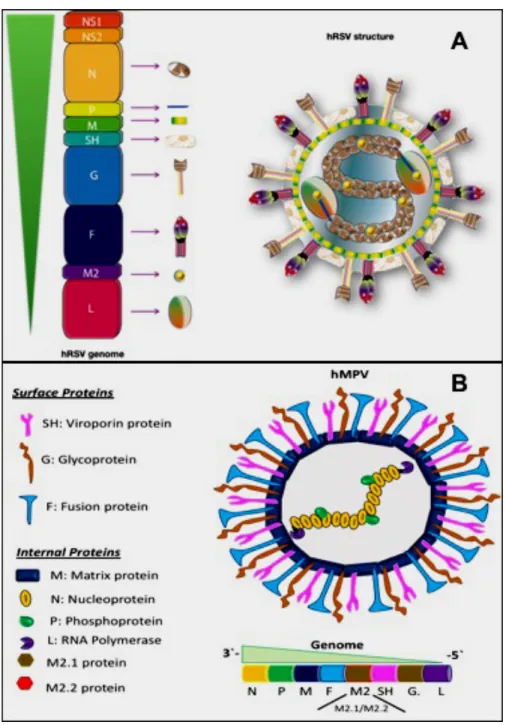
(16) 16. Figure 1. Total number of viral-associated respiratory infection cases per epidemiological week in Chile. The graph shows the number of cases associated with different viral infections in Chile in the different weeks of the year. The analysis was made from the 2015 to at the present week of 2018. The viruses analyzed are the following: Metapneumovirus (yellow), Influenza B (calypso), Influenza A (purple), Parainfluenza (green), Adenovirus (red) and respiratory syncytial virus (blue). Also, the percentage of positivity of the total samples analyzed is shown in the orange line (Source: Institute of Public Health of Chile). http://www.ispch.cl/sites/default/files/documento/2018/08/Informe%20circulaci%C3%B3n%2 0virus%20respiratorios%20SE31%2007-08-2018.pdf).. 1.2. Viral Structure of hRSV and hMPV. Both viruses are RNA negative-stranded enveloped viruses, belonging to the Pneumoviridae family, being hRSV and hMPV part of the Orthopneumovirus and Metapneumovirus genus, respectively. HRSV has a genome of 15.2 Kb that contain 10 genes and codified to 11 proteins including two non-structural proteins (NS) and nine structural proteins transcribed in the following order 3`- NS1-NS2-N-P-M-SH-F-G-M2.1-M2.2-L- 5`.
(17) 17. (Figure 2A) (Gálvez, Soto, & Kalergis, 2017; Hacking & Hull, 2002). Both NS1 and NS2 proteins are the main predominant proteins found in the first stage of the viral replication, being these proteins considered as relevant virulence factors of hRSV (Ling, Tran, & Teng, 2009; Lo, Brazas, & Holtzman, 2005; Pretel, Camporeale, & de Prat-Gay, 2013). Importantly, NS1 is associated with the inhibition of the Interferon (IFN) response mediated by the impairment of the cellular apoptosis of the infected cells while the NS2 is associated with the cell death promoting an accumulation of these cells that can obstruct the airways (Liesman et al., 2014). On the other hand, the hMPV has a genome of 13 Kb consisting of 8 genes that codify to nine structural proteins in the following order 3`-N-P-M-F-M2.1-M2.2-SH-G-L- 5` as compared to the hRSV (Bernadette G van den Hoogen, Bestebroer, Osterhaus, & Fouchier, 2002)(Figure 2B). However, despite hMPV do not present the non-structural protein, its effect in the host cell is similar to the triggered by hRSV, avoiding the elimination of the virus for the immune system through different little-known or unknown immune evasion mechanisms that possibly are mediated by different factors as compared with hRSV (Huck et al., 2007)..
(18) 18. Figure 2. Genome and Protein Structure of hRSV and hMPV. The panel A showed the hRSV genomic organization and viral structure where is identified the 10 genes that codify to 11 proteins. The hRSV proteins are divide two groups: the non-structural proteins (NS1 and NS2), and structural protein where the N, P, M, M2.1, M2.2 and L are considered as internal proteins while SH, G, F are surface proteins. In the panel B showed the hMPV genomic organization and viral structure where is identified the 8 genes that codify to 9 structural proteins divided in internal proteins (N, P, M, M2.1, M2.2 and L) and surface proteins (SH, G, F). (Adapted from Galvez et al. 2017 and Soto J., et al 2018 In revision on Frontiers in Immunology)..
(19) 19. 1.3. Viral Cycle of hRSV. The viral cycle of hRSV begins with the anchor of the G glycoprotein to the plasmatic membrane. Then, the F protein promotes the fusion between the viral envelope and the plasmatic membrane of the host cell and allows the entry of the genetic material. Afterwards, the viral RNA covered by the N nucleoprotein is released into the cytoplasm, where the process of replication and transcription are carried out (Hacking and Hull 2002, Ogra 2004, Collins and Melero 2011). In this step, the actin is required to activate the viral polymerase (L protein) function as a transcription factor (Harpen et al. 2009) after another protein like the actinmodulator protein profile that participates as a cofactor promotes the transcription process on the hRSV genes (Harpen et al. 2009). Finally, the new ssRNA (-) genome obtained is used like a template to synthesize a messenger RNA that will be used to synthetize new proteins by the host ribosomes. In addition, the ssRNA (-) is also used as a template for the synthesis of ssRNA (+) antigenome that later will be used to generate new ssRNA (-) genome (Hacking and Hull 2002, Ogra 2004, Collins and Melero 2011). Finally, it is possible to identify new viral particles early within 10 to 12 hours post-infection (Collins and Crowe 2007).. 1.4. Viral Cycle of hMPV. The viral replicative cycle of hMPV begins with the attachment of the virus to the host cell through the G glycoprotein. Later, the F protein localized in the viral envelope interacts with the host plasmatic membrane, promoting the fusion of them (Terrier OCG et al., 2008). Such process allows the entry of the nucleocapsid into the cellular cytoplasm where the proteins N, P and L release the RNA and are matched to create an active polymerase complex triggering the replication and transcription of the genetic material (Easton AJ et al.,2004). The messenger.
(20) 20. RNA is used to the translation process generating new protein that will be used for assembly of new viral particles next to the new strands of genetic material synthesized. The production of new viral particles occurs during the budding at the surface of the host cell membrane (Mackay IM et al.,2003). Additionally, it has been described that hMPV is able to persist in respiratory infected cells of mice after the replication viral cycle is completed (Alvarez, Harrod, Shieh, Zaki, & Tripp, 2004a; Alvarez & Tripp, 2005). In these studies Plates Forming Unit (PFUs) in the lungs of infected mice from day 28 to 60 post-infection were found (Alvarez et al., 2004a). Also, viral RNA was detected even at day 180 post infection by Reverse- Transcription quantitative Polymerase Chain Reaction (RT-qPCR) suggesting that hMPV have mechanisms to persist inside of the host cell, avoiding the viral clearance until at least 2 months after the infection (Alvarez et al., 2004a; Alvarez & Tripp, 2005). Other possible mechanism associated with such persistence is associated with an increase in the Bcl-2 cellular protein expression in the surface of hMPV-infected airways epithelial cell (AECs) promoting the inhibition of the apoptosis machinery (Marsico et al., 2018).. 1.5. Immune response against the hRSV infection. The hRSV-induced immune response in the lung promotes an inefficient Th-2 profile associated with the IL-4, IL-5 and IL-13 cytokines production (Roman M et al., 1997), herein generating a proinflammatory environment with an exacerbation of mucus and a flow reduction in the airways with a low viral clearance the virus (Becker Y et al., 2006; Lee H et al., 2012). Importantly, such hRSV-induced cytokine profile has been associated with a risk factor to induce asthma (Pullan C et al., 1982; Kumar R et al., 2014). Additionally, the high secretion of different cytokines and chemokines such as IL-6, IL-8, TNF-α, CXCL2, and CXCL10, increase.
(21) 21. the recruitment of different innate cell including neutrophils, macrophages and dendritic cells, thereby raising the inflammation in the tissue (Roman M et al., 1997; Bendelja K et al., 2000; Wolterink K et al., 2013; Bohmwald K et al., 2015). Furthermore, the weak T cell immune response induced by the hRSV infection could be associated with the deficient immunological synapse between the infected DCs and the naïve T cell (Gonzalez P et al., 2008; Cespedes P et al., 2014). The inefficient immunological synapse triggers the lack of an antiviral response because do not exist a proper activation of CD4+ and CD8+ T IFNg+ and due to the low secretion of granzyme B and perforin factors (Janyra A. Espinoza, Bueno, Riedel, & Kalergis, 2014; Vallbracht, Unsöld, & Ehl, 2006; Varga, Wang, Welsh, & Braciale, 2001). On the other hand, the hRSV-induced humoral response is known to be not effective. Such humoral response consists in the secretion of IgG1 and IgG3 isotype antibodies without a neutralizing capacity, herein causing a poor viral clearance in the damage tissue (Magro et al., 2012). In the case of children under 6 month of age whose mother were exposed to hRSV during the pregnancy, present neutralizing antibodies able to control the viral infection (Chu et al., 2014; Jans et al., 2017; Ogilvie, Vathenen, Radford, Codd, & Key, 1981). However, it has been observed that long period after the 6 month of year are involved with a decrease in this antibody transferred by the mother to child promoting an increase in the infection (Jounai et al., 2017). Furthermore, it is reported that the main problem associated with the hRSV-infection in infants is because children under 4 month of year presents low somatic mutation, process that is very important to promote the maturation and the diversity of the B cells (Sande, Cane, & Nokes, 2014)..
(22) 22. Instead, infants with lower respiratory tract infections (LRTI) that trigger a lethal bronchitis have shown to present high levels of IgM, IgG and IgA in the postmortem lungs, which is associated with an increase of the mature B cell (Raes et al., 1997; ROMÁN et al., 1997). These increase of the antibodies classes could be associated with an over expression of B-cell stimulating factors such as a proliferation-inducing ligand (APRIL) and B-cell-activating factor (BAFF) that are stronger expressed by an special type of plasma cell in the presence of the LRTI-associated hRSV-infection (A. et al., 2011; McNamara et al., 2013). 1.6. Immune response against the hMPV infection. The immune response generated by hMPV infection is mainly associated with a Th-17 profile that is characterized by the IL-6 and TNF-α secretion in the lung (Huck B et al., 2007). Such immune response is also accompanied by an inadequate Th-2 profile, which is described by the early secretion of interleukin (IL)-4, IL-5, IL-8 and other pro-inflammatory cytokines (Romagnani S et al., 2003; Fietta P et al., 2009; Ito T et al., 2004). On the other hand, the thymic stromal lymphopoietin (TSLP) cytokine has been described to impair the T cell activation, generating a delay in the Th-1 immune response and promoting the secretion of Th-2- like cytokines, followed by neutrophil recruitment to the lungs (Lay M et al., 2015). In addition, an increase of the mucus secretion in the goblet cells associated with an exacerbation in the inflammatory response and a collapse in the airways are found (Boivin G et al., 2003; Hamelin et al., 2006). It has also been reported that in some critical-health care cases the hMPV-infection promotes a chronic pulmonary obstructive disease and an exacerbated response such as asthma in humans (Hamelin et al., 2006; Peiris J et al., 2003). Additionally, the infection of hMPV in mice has been described to promote an early Th1-like profile that promote the initial viral control, but this response switches towards a Th2-like profile at early time of 7 and 14 days.
(23) 23. post-infection (Alvarez R et al., 2004; Alvarez R et al., 2005). On the other hand, the effect of the hMPV infection in dendritic cells (DCs) promotes a decrease in the activation of CD4+ T cells associated with low levels of surface markers such as CD25, CD69 and CD71 as compared to T cells co-cultured with uninfected DCs (Cespedes PF et al., 2013). Importantly, the hMPV infection in human peripheral blood mononuclear also induces an inefficient CD4+ T IFNg+ cells promoting a delay in the antiviral immune response and triggering a low viral clearance of the infection (Douville R et al., 2006). On the other hand, similar to the previously reported in hRSV, the inefficient host immune response found in infants is mainly associated to the absence of somatic mutation in the B cell populations, process that it is necessary to promote the maturation and increase the diversity of these cells (John V. Williams, Weitkamp, Blum, LaFleur, & Crowe, 2009). Also, a high detection of antibodies in children of different age groups, suggesting a minor humoral role in the viral clearance (Lüsebrink et al., 2010) as well as a possible interference in the B cell activation induced by the limited helper T cells (Talaat et al., 2013). In contrast, the B cell population and the antibody secretion have been evaluated in murine model observing a delay in the B activation and the antibody secretion (Alvarez & Tripp, 2005). However, the passive transfer of the sera from infected mice have shown a partial protective role (Alvarez & Tripp, 2005). Two candidates used to promote the antibody secretion have been the M- and F-hMPV proteins. Both proteins were evaluated in the production of virus -like particle (VLP) promoting the activation of B cell populations and the secretion of neutralizing antibodies whose effect increase only in presence of the TiterMax Gold or αgalactosylceramide adjuvant (Cox et al., 2014)..
(24) 24. 1.7. Host immune response evasion mechanisms of hRSV and hMPV. Both hRSV and hMPV viruses inhibit the immune system through the inhibition of the type I IFN (Bao, Liu, et al., 2008; Lo et al., 2005) and the impairment of the activation of T cells population. hRSV induces the inhibition of the type I IFN through the NS1 and NS2 proteins. These proteins impair the phosphorylation and translocation of the interferon regulatory factor 3 (IRF3). (Spann, Tran, & Collins, 2005). Other mechanism used by hRSV is the inhibition of the NF-kB pathway (Tian et al., 2002). Similarly, hMPV is also able to impair the type I IFN through the inhibition of IRF3 and NF-kB, which has been involved to the G, SH and M2.2 proteins in different processes (Bao, Kolli, et al., 2008; Bao, Liu, et al., 2008; Cheemarla & Guerrero-Plata, 2017; Chen et al., 2016; Ren et al., 2012; Seth, Sun, Ea, & Chen, 2005). Another pathway described in the type I IFN inhibition by hMPV is associated with the activation of mainly the Toll like receptors (TLR)3 and TLR7. (Goutagny et al., 2010; Ren et al., 2012). On the other hand, hRSV has been found to be able to impair of T cell population through the inhibition of the immunological synapses between the dendritic cell (DC) with the T naive lymphocyte mediated by the nucleoprotein (N) (Cespedes et al., 2014). Such inhibition occurs because the N protein promotes the cluster formation near the contact zone between the major histocompatibility complex (MHC) and the T cell receptor (TCR) triggering a poor communication that affect the polarization of the T naïve cell in a Th-1 profile (Cespedes et al., 2014). Contrary to hRSV, the induction of the Golgi polarization occurs in hMPV infection, but such impairment is done in the signalization of the TCR in the T naïve cell, a process that could be associated with a soluble factor, however, such hMPV immune evasion remains unknown. Moreover, it has been described that the deletion of both viral G and SH genes is related to an.
(25) 25. increase of the number of immunological synapses between DCs stimulated by hMPV and memory CD4+ T cells, therefore promoting the activation of these memory T cells. Altogether, it has been shown that G and SH proteins might reduce the ability of hMPV-infected DCs to activate CD4+ T cells, since both proteins are associated with a decrease in the internalization of the virus into DCs (Le Nouen et al., 2014). Finally, recently it has been described other mechanisms of hMPV evasion from immune system such as the effect of the SH glycoprotein on the inhibition of the NF-kB pathway in human lung epithelial cells (Bao, Kolli, et al., 2008).. 1.8. Use of Bacillus Calmette-Guérin as a vector. The Bacillus Calmette-Guérin (BCG) is an attenuated Mycobacterium bovis used as a vaccine against the Mycobacterium tuberculosis, being widely used worldwide and considered as the first vaccine used in humans (Calmette et al., 1928; Barletta B, 1990). In addition, BCG has been used as a vector to generate vaccines against some pathogens like Borrelia burgdorferi (Langermann S et al., 1994), Human Immunodeficiency Virus (HIV) (Fuerst T et al., 1991; Kim BJ et al., 2018; Power C et al., 2010), Malaria (Matsumoto S et al., 1998), Measles virus (Zhu Y et al., 1997), Hepatitis C virus (Uno-Furuta S et al., 2003), Influenza virus (Power C et al., 2010) hRSV (Bueno S et al., 2008; Cautivo C et al., 2009) and hMPV (Palavecino C et al., 2014). The main feature of BCG is the ability to increase the Th-1 immune response, herein promoting a better MHC-I presentation for the Antigen Presenting Cell (APCs) towards the activation of CD8+ T naïve cell (Grode L et al., 2005; Sun R et al., 2009). Also, BCG exerts production of high levels of IFN-g and other cytokines such as IL-17 and IL-22, thereby helping with the efficient immune response against different intracellular bacteria and viruses through.
(26) 26. driving the balance of increase the ratio Th-1/Th-17 immune response, which has been identified in prolonged times after vaccination with BCG (Kleinnijenhuis J et al., 2014). Additionally, it has been reported that BCG immunization induces an increase of the IgG1, IgG2, IgG3 antibody titer, triggering a strong humoral immune response against Mycobacterium tuberculosis (Hoft et al., 1999) or inducing high IgA antibody titer associated with the detection of lipoarabinomannan (LAM) that is an important component of the mycobacterial cell wall (Brown et al., 2003). Also, different reports that use BCG strains administrated in different pathways as oral, intranasal and intradermal have been associated with an increase in the mucosal or sera antibody secretion (Hamasur et al., 2003; A. Williams et al., 2004). Also, the BCG vaccination promotes the raise of the long-lived memory B cells next to the increase in the phagocytosis mediated between both IgA and IgG synergy. Such synergy triggers an enhanced of the cell mediated cytotoxicity and IL-17-associated neutrophils recruitment to the lungs. Other function associated with the BCG vaccination is related to the selection and survival of the B cell and its maturation towards plasma cell and memory B cell by Follicular B-helper T cell (TFH) (Moliva, Turner, & Torrelles, 2017).. 1.9. Vaccines and Treatments. Nowadays, no licensed vaccine against hRSV or hMPV is available despite the efforts made by different research groups. Currently, according to the PATH, almost 6 different candidates are under development (Table 1). In 1960, it was developed the first vaccine against the hRSV, the Formalin-Inactivated hRSV (FI-hRSV). Importantly, the effects of such FIHRSV vaccine in children were disastrous since children vaccinated with this vaccine after being exposed to a natural hRSV infection triggers an exacerbation of the hRSV-associated.
(27) 27. immunopathology with a pro-inflammatory environmental, a high mucus secretion and collapse of the airways (Kim et al., 1969; Murphy & Walsh, 1988). Such FI-hRSV vaccine-caused effects generated an increase in the number of hospitalized children and the death of two children vaccinated with the FI-hRSV. Therefore, such vaccine was not authorized its approval in humans and was eliminated of the market proposal. Later, it has been proposed that the FIHRSV-induced exacerbation might be associated with an allergic-like response and a possible change in the epitope of viral protein, herein promoting the recruitment of neutrophils, eosinophils and pro-inflammatory cytokines to the lungs (Prince, Curtis, Yim, & Porter, 2001). Nevertheless, other vaccine prototypes using mutant virus with depletion in different glycoproteins like the G, F, SH (Garg et al., 2014; Nguyen et al., 2013; Russell et al., 2015); subunit vaccine(Cayatte et al., 2017), Virus Like-Particles (VLPs) (Ghildyal, Mills, Murray, Vardaxis, & Meanger, 2002; Kurt-Jones et al., 2000) or chimeric proteins (Lambert et al., 2015) have been studied and evaluated in different animal models such as mice, cotton rat, syrian hamster, obtaining promising results (Krarup et al., 2015; Lambert et al., 2015). However, despite that some of these vaccine prototypes was able to promote a safe and security effects in early clinical trial, the presence of reactogenicity response in some subjects reported in the phase II or III clinical trials were enough to be discarded as a possible commercial vaccine. Nowadays, the only licensed and available prophylactic treatment against hRSV consists in the use of a humanized monoclonal antibody against the F-hRSV protein named Palivizumab (Synagys®) whose effect also is enhanced with the use of artificial ventilation and clinical measures that help children breathe and eliminate lung obstruction (Huang, Incognito, Cheng, Ulbrandt, & Wu, 2010). Such prophylactic treatment has been approved by the USA Food and Drug Administration (FDA) and is administrated mainly in high-risk infants, such as premature.
(28) 28. children or with respiratory insufficiencies, being the best opportunity today to decrease the hRSV-associated symptomology. Furthermore, the complete treatment consists in five doses and may not necessarily avoid the strongest respiratory symptoms of the ALRTI induced by the hRSV infection, being effective only in the 50% of the cases (Huang et al., 2010). Furthermore, the absence of an immunological memory induced by this treatment accompanied by the high cost of each dose decreases its commercial appeal. Nowadays, other similar monoclonal anti-F antibodies like the Motivizumab, MEDI-524, and MEDI 8897 have been developed while new approaches against the G glycoprotein has also been made such as the 131-2G mab that have shown an effective decrease in the hRSV-associated pulmonary damage in murine model infection (Abarca et al., 2009; Fernández et al., 2010; Gill & Welliver, 2009; Lagos et al., 2009; Ramilo et al., 2014; Robbie et al., 2013; Weisman, 2009)..
(29) 29. Table 1. Clinical Trials vaccines and therapies against the hRSV.. * Corresponding to the name of the vaccine prototype evaluated. (Adapted from path.org) https://www.path.org/resources/rsv-vaccine-and-mab-snapshot/. With regards to hMPV, it does not exist any commercial vaccine or antibody approved as a treatment against the virus-infection. Such lack of vaccines in development could be associated with the limited knowledge with respect to the evasion mechanisms of host immune system induced during the hMPV-infection. Nevertheless, the use of attenuated live vaccines, recombinant virus, VLP, subunits vaccines have been explored in preclinical studies in rodents.
(30) 30. as mice and cotton rat and non-human primates (Biacchesi et al., 2005; Cox et al., 2014; DiazDinamarca et al., 2018; Karron, San Mateo, Wanionek, Collins, & Buchholz, 2017; Lévy et al., 2013; MacPhail et al., 2004; Schickli et al., 2008). In addition, similar strategies applied in other respiratory viruses such as hRSV, influenza has been explored. Surprisingly, from the year 2010 to date it has been reported three different vaccine prototypes in clinical phases (clinicaltrials.gov, Table 1). Moreover, some different strategies used to develop prophylaxis therapies against the hMPV-infection have been described. Some of these strategies involved the use of ribavirin and polyclonal antibody IVIg (Kitanovski et al., 2013) that promote the neutralizing antibody secretion. Importantly, the use of monoclonal antibodies promotes the protective role against the hMPV-infection in hamster model (M.-E. Hamelin et al., 2010; Ulbrandt et al., 2006) and mice and cellular cultures (Schuster et al., 2015). Table 2. Clinical Trials vaccines against the hMPV.. * Corresponding to the name of the vaccine prototype evaluated. (Adapted from clinicaltrials.gov)https://clinicaltrials.gov/ct2/results?cond=hMPV&term=&cntry= &state=&city=&dist= On the other hand, a new vaccine prototype against both hRSV and hMPV has been developed in our laboratory in Chile (Bueno et al., 2008; Cautivo et al., 2010; Céspedes et al., 2017; Palavecino, Cespedes, Gomez, Kalergis, & Bueno, 2014). These vaccines consist in a.
(31) 31. recombinant Bacillus Calmette-Guérin (rBCG) expressing the N nucleoprotein of hRSV (rBCGN) or the P phosphoprotein of hMPV (rBCG-P). Previous report using a high dose of 1x108 CFU of both vaccines demonstrated that animals immunized with these prototypes were able to decrease virus-associated immunopathology including the reduction of neutrophils infiltrated into the lungs, decreased of inflammation of the tissue and the viral load (Bueno et al., 2008; Cautivo et al., 2010; Céspedes et al., 2017; Palavecino et al., 2014). Additionally, it was possible to detect an early recruitment of IFN-g+ CD4+ and CD8+ T cells into the lungs. Also, the transference of T cells-induced from the immunized mice to naïve mice showed that these T cells were able to protect against the viral infection of both viruses (Bueno et al., 2008; Cautivo et al., 2010; Céspedes et al., 2017; Palavecino et al., 2014). Moreover, the promising results obtained from the cellular immune response induced by the recombinant strain allowing generate the rBCG-N vaccine in Good Manufacture Practice conditions (cGMP) showed similar results using a unique dose of 1 x105 CFU (Céspedes et al., 2017). Such dose was validated in the 90’s for the use in human as a protective role of a n unique and low dose of 1 x105 CFU (Aggarwal & Dutta, 1995). Such dose is currently used worldwide and approved by the USA Food and Drug Administration (FDA) and the World Health Organization (WHO). Given the positive results obtained using a low dose of the rBCG-N in our laboratory, this experimental vaccine is currently being evaluated in a clinical trial led by the Pontificia Universidad Católica de Chile (Table 1). However, the effect of the humoral immune response induced by rBCG-N with respect to the natural hRSV-infection has not been evaluated. The Good Manufacturing Practices conditions (cGMP) is a process strictly necessary to generate products approved for human use. This process is characterized by following different regulatory processes that control the use of factors including the culture media, supplement.
(32) 32. enrichment absence, scaling of culture and restriction of antibiotics, among others, might affect the yield of the initial product before being subjected to the manufacturing process. Based on the promising results obtained from the rBCG-P vaccine. We propose that to carry out a manufacturing process of this vaccine we could have a first step for the development of a vaccine against hMPV for human use. Therefore, if we use the previously background mentioned that both recombinant vaccines promote a protective cellular immune response associated with the activation of CD4+ and CD8+ secretors of IFN-g , we propose that both vaccines could also promote a protective humoral immune response characterized by a switching isotype and neutralizing effect capable of controlling the viral infection as compared with the immune response induced by the natural infection of hRSV- or hMPV..
(33) 33. 2. HYPHOTESES STAMENT. 2.1. HYPHOTESIS 1 TWO RECOMBINANT BACILLUS CALMETTE-GUÉRIN EXPRESSING THE N NUCLEOPROTEIN OF HRSV (rBCG-N) OR THE P PHOSPHOPROTEIN OF HMPV (rBCG-P) INDUCE A PROTECTIVE HUMORAL IMMUNE RESPONSE IN MICE PROMOTING A CHANGE IN THE ANTIBODY SECRETION WITH RESPECT TO THE NATURAL VIRAL INFECTIONS.. 2.2. HYPHOTESIS 2 THE UNIQUE DOSE OF A RECOMBINANT BACILLUS CALMETTE-GUÉRIN EXPRESSING THE P PHOSPHOPROTEIN OF HMPV (rBCG-P) UNDER GOOD MANUFACTURING PRACTICES CONDITION (cGMP) APPROVED FOR HUMAN USE PROMOTES A PROTECTIVE CELLULAR AND HUMORAL IMMUNE RESPONSE AS COMPARED WITH THE NATURAL VIRAL INFECTION GENERATED BY HMPV IN MICE..
(34) 34. 3. OBJECTS 3.1. GENERAL AIMS •. To evaluate the humoral immune response induced by two recombinant BCG strains against the hRSV or hMPV-infection in a murine model.. •. To evaluate the cellular and humoral immune response induced by a unique dose of a cGMP recombinant Bacillus Calmette-Guérin expressing the P phosphoprotein of hMPV in a murine model after the viral infection.. 3.2 SPECIFIC AIMS FOR THE HYPOTESIS 1 •. To evaluate whether both rBCG-N and rBCG-P vaccines are able to induce an increase in the hRSV- and hMPV-specific antibody levels in sera with respect to the hRSV- or hMPV- natural infection.. •. To evaluate whether both rBCG-N and rBCG-P vaccines promote an isotype switch and induce a neutralizing effect of the antibodies secreted in immunized and infected animals as compared with the natural viral infection.. •. To evaluate the protective role of the antibodies secreted in the immunized animals into a naïve mice receptor against hRSV or hMPV infections.. 3.3. SPECIFIC AIMS FOR THE HYPOTESIS 2 •. To determine whether a unique dose of 1x105 CFU of cGMP rBCG-P is able to decrease the hMPV- associated pathology in mice.. •. To evaluate the innate and adaptive immune cells promoted by the cGMP rBCG-P immunization in the lungs from hMPV-infected mice..
(35) 35. •. To evaluate the humoral immune response induced by the cGMP rBCG-P immunization in hMPV-infected mice.. •. To evaluate in vivo the protective role of the sera and cell induced by cGMP rBCGP immunization with a unique dose of 1x105 CFU of vaccine transferred into naive mice..
(36) 36. 4. CHAPTER 1. Recombinant BCG vaccines reduce pneumovirus-caused airway pathology by inducing protective cellular and humoral immunity rBCG induces protection against pneumoviruses Jorge A. Soto+1, Nicolás M. S. Galvez+1, Claudia A. Rivera1, Christian E. Palavecino1, Pablo F. Céspedes1, Emma Rey-Jurado1, Susan M. Bueno1, Alexis M. Kalergis*1,2. 1. Millennium Institute of Immunology and Immunotherapy, Departamento de Genética Molecular y Microbiología, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, Santiago, Chile. 2 Departamento de Endocrinología, Facultad de Medicina, Pontificia Universidad Católica de Chile, Santiago, Chile.. *Correspondence: Dr.Alexis Kalergis Parra [email protected] Keywords: hRSV, Orthopneumovirus, hMPV, antibodies, humoral immune response, vaccine,.
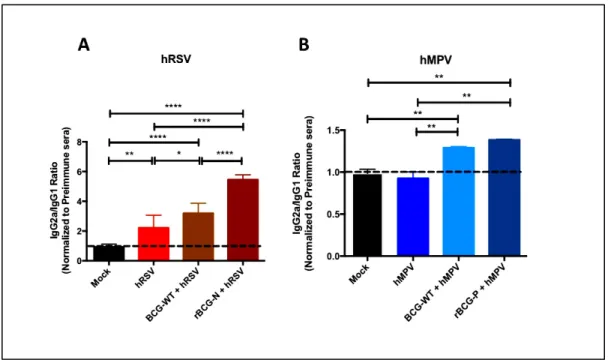
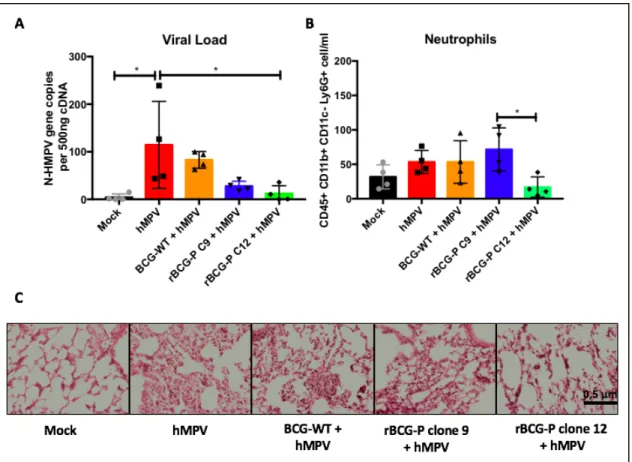
(37) 37. 4.1. ABSTRACT The Human Respiratory Syncytial Virus (hRSV) and the Human Metapneumovirus (hMPV) are two pneunoviruses and lead as causative agents of acute lower respiratory tract infections (ALRTIs) affecting young infants, the elderly and immunocompromised patients globally. Since these pathogens were first discovered, many approaches for the licensing of safe and effective vaccines have been explored being unsuccessful to date. We have previously described that immunization with recombinant strains of Mycobacterium bovis Bacillus Calmette-Guérin (rBCG) expressing the hRSV nucleoprotein (rBCG-N) or the hMPV phosphoprotein (rBCG-P) induced immune protection against each respective virus. These vaccines efficiently promoted viral clearance without significant lung damage, mainly through the induction of a T helper 1 cellular immunity. Here we show that upon viral challenge, rBCGimmunized developed protective humoral immunity, characterized by production of antibodies specific for most hRSV and hMPV proteins. Further, isotype switching from IgG1 to IgG2a was observed in mice immunized with rBCG vaccines and correlated with an increased viral clearance, as compared to unimmunized animals. Finally, sera obtained from animals immunized with rBCG vaccines and infected with their respective viruses exhibited virus neutralizing capacity and protected naïve mice from viral replication and pulmonary disease. These results support the notion that the use of rBCG strains could be considered as an effective vaccination approach against other respiratory viruses with similar biology as hRSV and hMPV..
(38) 38. 4.2. INTRODUCTION For almost a century, Mycobacterium bovis Bacillus Calmette-Guérin (BCG) has been widely used to prevent Tuberculosis and has also been characterized as an effective T helper type 1 (Th1) inducer (Zhang et al., 2013). Further, BCG has been shown to be safe in adults, infants and newborns. The approach of using BCG as a vector for recombinant expression of heterologous antigens has been previously tested for several pathogens, such as measles virus, rotavirus, hepatitis B virus, Plasmodium yoelii, Bordetella pertussis and Toxoplasma gondii, exhibiting promising results in mouse models for those diseases (Dennehy, Bourn, Steele, & Williamson, 2007; Fennelly, Flynn, Ter Meulen, Liebert, & Bloom, 1995; Matsumoto et al., 2000; Medeiros, Armôa, Dellagostin, & McIntosh, 2005; Rezende, De Moraes, Matos, Mcintoch, & Armoa, 2005; Wang et al., 2007). Worldwide, human Respiratory Syncytial Virus (hRSV) is the leading cause of acute lower respiratory tract infections (ALRTIs). HRSV was first identified in 1956 and mainly afflicts infants, young children, elderly and immunocompromised patients, causing about 34 millions of ALRTIs and approximately 200,000 deaths per year (Nair et al., 2010). Next to hRSV, human Metapneumovirus (hMPV) is the second cause of ALRTIs (Lefebvre et al., 2016) and was first identified in 2001 (Berry, Gamieldien, & Fielding, 2015). Although the overall burden in hospitalization remains poorly characterized for hMPV, as it was recently identified, it has been estimated that about a 7%-19% of children hospitalization can be due solely to this virus(Edwards et al., 2013b; John V Williams et al., 2004). Furthermore, clinical studies have shown that by the age of 5, virtually every child has been infected with both of these viruses, exhibiting classical ALRTI manifestations, which include fever, cough, wheezing and some clinical manifestation like bronchiolitis, laryngotracheitis, acute bronchitis and pneumonia.
(39) 39. (Berry et al., 2015; Edwards et al., 2013b; Hahn et al., 2013; Nair et al., 2010; Tregoning & Schwarze, 2010; John V Williams et al., 2004) with an increase in the mucus production, obstruction of bronchoalveolar spaces, exacerbated inflammatory response and the generation of airway hyper-responsiveness (Afonso et al., 2016; P. L. Collins & Melero, 2011; Mukherjee & Lukacs, 2013). In addition, prospective surveillance studies have suggested that children affected by severe hMPV infections usually require longer recovery periods at intensive care units than do children infected with hRSV(Edwards et al., 2013b; Hahn et al., 2013; Tregoning & Schwarze, 2010; John V Williams et al., 2004). Both hRSV and hMPV are RNA singlestranded, negative sense enveloped viruses, belonging to the Pneumoviridae family, particularly the Orthopneumovirus genus and the Metapneumovirus genus, respectively (Afonso et al., 2016). Furthermore, hRSV has been recently renamed as human Orthopneumovirus (Afonso et al., 2016). Some reports have suggested that the host immune system is unable to generate an effective and protective immunological memory against either of these viruses, which after disease resolution, prompts the acquisition of repeated infections throughout life (Janyra A. Espinoza et al., 2014; Gomez, Guisle-Marsollier, Bohmwald, Bueno, & Kalergis, 2014). Accordingly, it has been described that the nucleoprotein of hRSV (N-hRSV) is able to inhibit the assembly of an effective immunological synapse, apparently by clustering with the pMHCTCR complex (Céspedes et al., 2014). Also, N-hRSV blunts the interferon response by interacting with MDA5 and MAVS, pivotal elements in the main pathways associated with the viral clearance (Caly, Ghildyal, & Jans, 2015). On the other hand, the phosphoprotein of hMPV (P-hMPV) has been described as a crucial component for the assembly of the virus replication core (Derdowski et al., 2008). It has been reported that P of hMPV-B1 serotype could interfere.
(40) 40. with the RIG-I pathway, prompting the inhibition of the interferon I pathway (Goutagny et al., 2010). Considering this, both proteins have been previously suggested as possible candidate antigens for the induction of a strong and protective cellular immune response against either hRSV or hMPV infections when used for immunization, respectively (Bueno et al., 2008; Palavecino et al., 2014). Our group has previously reported that recombinant BCG strains (rBCG) expressing either N-hRSV (rBCG-N) or P-hMPV (rBCG-P) as heterologous antigens, can protect against infection by hRSV or hMPV, respectively (Bueno et al., 2008; Palavecino et al., 2014). In this work, we evaluated the previously unexplored humoral immune response induced in mice immunized with either rBCG-N and rBCG-P. We observed that the post-challenge antibody response is enhanced by the established immunity elicited by both rBCG vaccines. This concerted response was able to significantly decrease viral replication and disease by promoting the secretion of neutralizing antibodies specific against the attachment and the fusion glycoproteins of both paramyxoviruses. These results suggest that rBCG strains are good vaccine candidates able to induce a cellular immune response capable of boosting the humoral immune response against unrelated antigens and to prevent the disease cause by both pneumoviruses..
(41) 41. 4.3. MATERIALS AND METHODS 4.3.1. hRSV and hMPV propagation and titration HEp-2 cells (American Type Culture Collection, CCL-23TM) and LLC-MK2 (American Type Culture Collection, CCL-7TM) were used to propagate hRSV serogroup A2, strain 13018– 8 (clinical isolate obtained from the Instituto de Salud Pública de Chile) and hMPV serogroup A, strain CZ0107 (clinical isolate obtained from the Laboratorio de Infectología y Virología of the Hospital Clínico, Pontificia Universidad Católica de Chile) (J A Espinoza et al., 2013; Reina et al., 2007). Briefly, cell monolayers were grown in T75 flasks with DMEM (Life Technologies Invitrogen, Carlsbad, CA) supplemented with 10% FBS (Gibco Invitrogen Corp, Carlsbad) for HEp-2 cells and Opti-MEM supplemented with 5% FBS for LLC-MK2 cells, until 80-90% confluence. Flasks containing 5 ml of infection medium (DMEM 1% FBS for hRSV and OptiMEM 5% FBS medium, supplemented with CaCl2 (100 µg/ml) for hMPV) were inoculated with 2 x 105 Plaque formation units (PFU) of the respective virus and incubated at 37 °C. After viral adsorption (2 h), supernatants were replaced with fresh medium (DMEM 1%FBS and OptiMEM) and incubated for 48 h for hRSV and 72 h for hMPV, until visible cytopathic effect was observed. For harvesting, cells were scraped, and the flask content was pooled and centrifuged first at 300 x g for 10 min and then at 500 x g for 10 min in order to remove cell debris. In parallel, supernatants of non-infected cells monolayers (HEp-2 and LLC-MK2) were collected as previously described and used as non-infectious control (Mock). Viral titers of supernatants were determined by immunocytochemistry in 96-well plates with HEp-2 and LLC-MK2 cells monolayers, as previously described (Céspedes, Gonzalez, & Kalergis, 2013a; J A Espinoza et al., 2013; Reina et al., 2007; Tollefson, Cox, & Williams, 2010). hRSV and hMPV inoculums were routinely evaluated for lipopolysaccharide and Mycoplasma contamination..
Figure



+7




Documento similar
