Breeding ecology of brown booby (Sula leucogaster) in Gorgona Island, eastern tropical Pacific Ocean
9
0
0
Texto completo
(2) Brown booby on a Neotropical island. 11. Fig. 1. Map of Gorgona Natural National Park, with location of sampling sites in the main island of Gorgona and Gorgonilla Island.. Gorgona NNP supports 150 pairs of brown boobies, and is the most important known breeding site for the etesiaca subspecies (Naranjo et al. 2001; Ospina-Alvarez 2008). Gorgona NNP is a 49,200 ha reserve that includes the island of Gorgonilla, some islets and the surrounding marine area. A total of 9 nesting sites are located in the vicinity of Gorgonilla Island, while 2 islets in Juanchincho and another 2 islets in El Horno host nesting sites (Fig. 1). The 2 islets of Juanchincho are in the NW of Gorgona Island; in this zone brown pelicans (Pelecanus occidentalis), magnificent frigatebirds (Fregata magnificens) and blue-footed boobies (Sula nebouxii) are less frequent and no nests of these species have been found (Cadena-Lopez 2004; Ospina-Alvarez 2008). In Gorgona NNP, brown boobies breed asynchronously; in censuses carried out every 15 days, nests with eggs, chicks and fledged chicks were recorded simultaneously (Cadena-Lopez 2004; Ospina-Alvarez 2008). Despite this asynchrony, 2 reproductive peaks occur, with the period of higher activity between October and January. From March until July the number of nests was <27% of the preceding reproductive peak (Ospina-Alvarez 2008). This study describes a number of poorly known aspects of the reproductive biology of brown booby in Gorgona NNP. The objective was to provide better information on the breeding biology of brown boobies that can then be used to improve the. management of protected areas with this and other cohabiting marine bird species. METHODS Data were collected during surveys conducted between October 2002 and October 2003 in Gorgona and Gorgonilla Islands, in the Gorgona Natural National Park, Colombia (Fig. 1). Gorgona is the principal island, 27 km from the Colombian Pacific coast (2°56’ N, 78°12’ W). The mean annual temperature is 27°C, the relative humidity is 88%95%, and the mean annual precipitation is 6,694 mm. Surface temperature of the sea ranges between 27.0 and 28.5°C. Distributed around the main island of Gorgona and Gorgonilla Island are a number of small islets, cays and emergent rocks. From 9 to 16 October 2002 Gorgona NNP was surveyed and 9 nesting sites were found in Gorgonilla, 2 in El Horno and 2 in Juanchincho (Fig. 1). A 6 m boat was used to conduct visual surveys around Gorgona NNP to assess the presence of nesting sites. Nesting sites of more than 10 pairs were classified as colonies, less than 10 pairs as aggregations and trios were recorded separately. From October 2002 to October 2003 all nesting sites were surveyed to count pairs and eggs laid by each pair. In the Juanchincho II colony (JCH2 in Fig. 1), adults were gently captured at each nest. Morphometric parameters measured were body.
(3) 12. Ospina-Alvarez. Table 1. Brown booby variation in clutch sizes. Number of nests with 1, 2 or 3 eggs, percentages (in parentheses) and average clutch size in Gorgona NNP (this study) and other localities in the Pacific Ocean. Sources: [a] Nelson (2005); [b] Woodward (1972); [c] Heatwole et al. (1990). Number pairs or nests. 1 egg. 2 eggs. 3 eggs. Mean. Gorgona NNP (Pacific Ocean). 243. 74 (30.5). 165 (67.9). 4 (1.7). 1.71. Christmas Island (Indian Ocean) [a]. 72. 42 (56.9). 30 (41.7). 1 (1.4). 1.44. Christmas Island (Indian Ocean) [a]. 50. 34 (68.0). 16 (32.0). 0 (0.0). 1.32. Gulf of California (Pacific Ocean) [a]. 42. 7 (16.7). 34 (81.0). 1 (2.4). 1.86. Kure Island (Pacific Ocean) [b]. 93. 7 (7.5). 78 (83.9). 8 (8.6). 2.01. Cocos Island (Pacific Ocean) [a]. 24. 8 (33.3). 16 (66.7). 0 (0.0). 1.67. Bedout Island (Indian Ocean) [a]. 30. 16 (53.3). 14 (46.7). 0 (0.0). 1.47. Great Barrier Reef (Pacific Ocean) [c]. 39. 20 (51.0). 19 (49.0). 0 (0.0). 1.49. Locality. weight and length, wingspan, bill and tail length. Body weight was recorded early in the morning with a field scale (to the nearest 1 g). The wing length was measured with a flexible meter rule (to the nearest 1 mm) from the sternum to the tip of the longest primary, when the wing was outstretched from the body. Thus, wingspan was twice the wing length. Body length was measured with a flexible meter rule (to the nearest 1 mm) from the bill tip to the tip of the tail with the back on a flat plane and the neck extended. Bill and tail length were measured with calipers (to the nearest 0.1 mm). All metrics are indicated with standard deviations. The same colony was visited each day in the early morning to establish a date for the first laid egg. Following the beginning of laying, the nests were observed every 2 to 3 days to check for new eggs. On the day of laying, each egg was measured with calipers (length and width to the nearest 0.1 mm) and marked with indelible coloured markers according to laying sequence. Chicks were captured, weighed and gently massaged on the abdomen to facilitate regurgitation. Stomach contents were collected and preserved in formalin. In the laboratory, stomach contents were identified using Fishbase website (Froese & Pauly 2000) and Allen and Robertson (1994) as references. In Juanchincho I and II, nesting pairs were studied, and completely-built nests were measured for height, and internal and external diameters (to the nearest 1 cm). For each nest site several parameters were measured: height (m) from low tide, mean inclination (degrees), percentage of. herbaceous cover, number of bordering walls (0 3) and width of platform (m). Qualitative physical characters around the nest also were considered: type of land (top, cliff or platform), direction of inclination (null, towards the rock, towards the sea), roof presence or absence, presence or absence of cracks, and drainage type (good or dry, bad or floodable terrain). Comparisons between physical characteristics of each nest and randomly selected points in Juanchincho I and II were performed. Each of the variables and randomly selected points was tested using Pearson’s χ2 test for contingency tables and Fisher’s exact test where applicable. Statistical analyses were conducted using SPSS v.13 software and R 2.8 package. This study was carried out in accordance with the recommendations in the Ethics Code for Biologist, and was approved by the Professional Council of biologist at Gorgona NNP from the Ministry for the Environment of Colombia. RESULTS Although the brown booby is a species that breeds colonially, in Gorgona NNP it was also possible to find pairs nesting individually, in trios or in aggregations of up to 10 individuals. Of 132 pairs studied between October 2002 and December 2003, 77 (58.3%) nested in 6 different colonies, 46 (34.8%) nested in 7 aggregations of up to 10 pairs, 3 (2.3%) in a trio, 4 (3.0%) in groups of 2 pairs, and 2 (1.5%) nested alone. The majority of nesting pairs was found around Gorgonilla Island, but the.
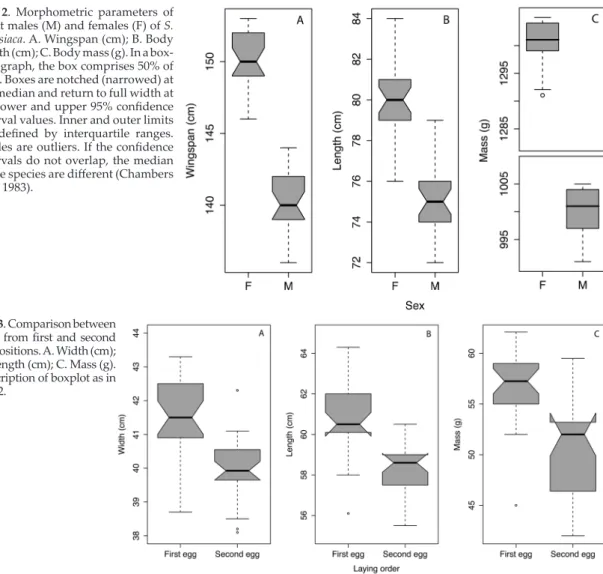
(4) Brown booby on a Neotropical island. 13. Fig. 2. Morphometric parameters of adult males (M) and females (F) of S. l. etesiaca. A. Wingspan (cm); B. Body length (cm); C. Body mass (g). In a boxplot graph, the box comprises 50% of data. Boxes are notched (narrowed) at the median and return to full width at the lower and upper 95% confidence interval values. Inner and outer limits are defined by interquartile ranges. Circles are outliers. If the confidence intervals do not overlap, the median of the species are different (Chambers et al. 1983).. Fig. 3. Comparison between eggs from first and second ovipositions. A. Width (cm); B. Length (cm); C. Mass (g). Description of boxplot as in Fig. 2.. bigger colonies were recorded in the Juanchincho I, Juanchincho II and El Horno Islets. In a sample of 50 randomly selected nesting sites, it was found that brown boobies built nests in 86% of the cases. Specifically, 31 (62%) were well-built nests and 12 (24%) were structureless masses of leaves, branches and guano. In the remaining 14% of the breeding attempts, brown boobies did not build nests, but instead deposited their eggs directly on the rock or in a small hole. As pairs breed more than once in a year, 243 eggs were recorded. The mean number of eggs per nest was 1.7 (Table 1). Around Gorgonilla, El Horno and Juanchincho Islands, brown boobies shared their breeding sites with blue-footed booby (Sula nebouxii) which used the same rocky cays and islets as resting sites even though they bred in other areas than Gorgona NNP. Juanchincho II was the only site where blue-footed boobies were not recorded during this study.. A total of 25 female and 27 male brown boobies were captured, weighed and measured (Fig. 2, Table 2). Mean body length of females was 80 ± 2.0 cm, their wingspan was 150 ± 2.0 cm and their body weight 1,300 ± 4.2 g. Males were smaller, with 75 ± 2.0 cm length; their wingspan was 140 ± 2.3 cm and their body weight was 1,000 ± 4.1 g. To minimise stress, only 9 female and 15 male brown boobies were choosen randomly and their tails and bills measured. The tails of males were longer (males: 17.89 ± 2.3 cm; females: 18.99 ± 2.84 cm) in relation with body length, and their bill was shorter (males: 9.17 cm ± 1.13; females: 9.86 cm ± 1.45). Statistics for bill and tail length were not performed due to the low sample size. Male/female body mass ratio was 0.77, indicating the presence of reversed sexual dimorphism. Significant differences between the sexes were found for all variables: body length (U-test: W = 600.5, P < 0.0001), wingspan length (U-test: W = 625.0, P < 0.0001) and.
(5) 14. Ospina-Alvarez. Table 2. Measurements of adult specimens of brown boobies in Gorgona NNP (this study) and comparison with subspecies from other localities around the world. For each characteristic the mean and number of individuals (between parentheses) is indicated. Sources: [a] Weimerskirch et al. (2009); [b] Hertel and Balance (1999); [c] Lindsey (1986); [d] Heller & Snodgrass (1901); [e] Nelson (2005); [f] Stager (1964). Note wingspan available for only first 4 populations. Wingspan (cm) Locality (subspecies). Body length (cm). Bill length (cm). Tail length (cm). Weight (g). Male. Female. Male. Female. Male. Female. Male. Female. Male. Female. Gorgona NNP (S. l. leucogaster). 140 (27). 150 (25). 75 (27). 80 (25). 9.2 (15). 9.9 (9). 17.9 (15). 19.0 (9). 1,000 (27). 1,300 (25). Gulf of California (S. l. leucogaster) [a]. 143 (4). 156 (5). -. -. -. -. 22.6 (6). 21.0 (3). 1,194 (19). 1,448 (20). -. -. -. -. -. -. Johnston Atoll (S. l. leucogaster) [b]. 152 (14). Northern Australia (S. l. plotus) [c]. 130 – 150 (?). 65 – 75 (?). -. 1,260 (14) 900 – 1,500 (?). -. Wing length (cm) 38.4 (1). 40.8 (3). 72.0 (1). 81.3 (3). -. -. 20.0 (1). 21.0 (3). -. -. -. -. -. -. 9.24 (?). 9.93 (?). -. -. -. -. Clipperton Island (S. l. brewsteri) [f]. 38.9 (4). 41.3 (2). -. -. -. 19.6 (4). 20.0 (2). -. -. Tres Marias Island (S. l. brewsteri) [f]. 37.2 (4). 39.7 (2). -. -. -. -. 17.6 (4). 18.2 (2). -. -. Various localities (S. l. leucogaster) [e]. 38.1 (13). 40.0 (10). -. -. 9.27 (13). 9.33 (10). 18.6 (13). 18.0 (10). -. -. Brazil (S. l. leucogaster) [e]. 39.5 (9). 40.2 (10). -. -. 9.35 (9). 9.42 (10). 19.4 (9). 19.1 (10). -. -. Cape Verde Islands (S. l. leucogaster) [e]. 39.2 (10). 40.4 (10). -. -. 9.47 (10). 10.0 (10). 19.0 (10). 19.1 (10). -. -. Ascencion Island (S. l. leucogaster) [e]. 39.0 (14). 40.5 (26). -. -. 8.90 (14). 9.50 (26). -. -. 1,025 (14). 1,325 (26). Various localities (S. l. etesiaca) [e]. 37.6 (6). 39.5 (11). -. -. 9.25 (6). 9.84 (11). 17.9 (5). 19.1 (11). -. -. Cocos Keeling & Christmas Islands (S. l. plotus) [e]. 39.5 (25). 41.6 (21). -. -. -. -. 20.8 (23). 20.6 (?). 962 (20). 1,260 (29). Clipperton Island (S. l. brewsteri) [d] Clipperton & Tres Marias Islands (S. l. brewsteri) [e]. body mass (U-test: W = 625.0, P < 0.0001) were all higher in females than in males. The length/mass ratio was higher in males (U-test: W = 0, P < 0.0001), suggesting that males were less heavy than females in relation to their length. In Juanchincho II, a total of 26 eggs were measured and weighed. Significant differences were found between the 2 eggs in a clutch (U-test: mass, W = 583.5; length, W = 612; width, W = 590; P < 0.0001 in all cases): first laid eggs were generally heavier, longer and wider than second-laid eggs (Fig. 3, Table 3). Brown booby chicks are naked at hatching, with transluscent purple skin. The first plumage. is downy, white and dense, which is then moulted into a brown plumage with fledging, at about 90 days, when individuals are normally beginning to fly. Growth of brown booby chicks showed a natural log increment, from hatching to an age of 90 days. This pattern fit a natural log equation where: body mass (g) = 0.8773 ln * days + 3.3895 (Fig. 4). Regurgitations of chicks were available for analysis in 27 occasions, involving individuals with ages between 45 and 90 days. The main items in the diet were longfin halfbeak (Hemiramphus saltator; 66.67%) and beautyfin flyingfish (Cypselurus callopterus; 25.93%), while the remaining part (7.4%) could not be identified due to its advanced stage of digestion..
(6) Brown booby on a Neotropical island The sites where aggregations and colonies were found had a mean height of 11.3 (± 3.2 cm) and a mean available surface of 196.1 (± 172.3 m2). The only difference between sites supporting colonies and sites supporting aggregations was the available area (U-test: W = 3.0, P = 0.01): the surface area of colony sites (326.1 ± 170.9 m2) was significantly greater than the surface of aggregation sites (84.7 ± 60.3 m2). A total of 31 well-built nests were measured; these had an external diameter of 31.5 ± 6.2 cm, an internal diameter of 17.4 ± 2.8 cm and a height of 5.1 cm ± 3.1 cm. The physical characteristics for 34 nests and the test for the selection of these characteristics by nesting pairs are summarised in Table 4. DISCUSSION Brown boobies in Gorgona NNP showed the reversed sexual dimorphism that is found in other populations of this species (Nelson 1978; 2005). Male/female body mass ratio was comparable with that calculated for other areas or subspecies, such as S. l. etesiaca in Johnston Atoll, S. l. plotus in Christmas Island and S. l. leucogaster in Ascension Island (Nelson 2005). This suggests that reversed sexual dimorphism in this species is widespread and no particular change in this pattern was evident in the subspecies of brown booby inhabiting Gorgona NNP. Even though a relatively large number of colonies was found in Gorgonilla, the number of individuals per colony was lower than in the other study sites, Juanchincho and El Horno. This difference may be due to both differences in predators and potential competitors. In Gorgona NNP, the breeding season of brown pelican occurs between January and April and the population is situated mainly in the Gorgonilla Island area. The reason for this may be the presence of the white-headed capuchin (Cebus capucinus) on the main island of Gorgona. Capuchins will eat bird eggs and this potential predator is not present in Gorgonilla (Alberico 1986). However, despite the spatial overlap, brown pelican may not be directly competing with brown boobies, because the pelican is a tree-nester while the booby is a rock-nester. On the other hand, Le Corre and Jouventin (1997) determined in a redfooted booby (Sula sula) colony that <1% of the population was losing food due to piracy by great frigatebird (Fregata minor) and lesser frigatebird (F. ariel). Diverse adopted strategies were identified to evade such pirating species, such as flying in large groups (>50 individuals) or returning to colonies during periods of low light. In Gorgona NNP, brown boobies form flocks with blue-footed booby, which can confer them a hypothetical advantage (OspinaAlvarez 2008). I can rule out kleptoparasitism by magnificent frigatebird on my study site but there may be a behavioural aversion encouraging brown. 15 A. B. Fig. 4. Growth parameters of S. l. etesiaca chicks. A. Weight increments (g) for 4 chicks from Juanchincho II colony, Gorgona NNP, Colombia in the reproductive season, Mar– Jul 2003. Single chicks are represented by lines; B. Logistic growth curve for the 4 chicks in Fig. 4A. The ideal growth model is the average of single growth curves for all chicks. The asymptotic mass was not different between groups.. boobies to identify a site with a high density of frigatebirds as less attractive. Blue-footed boobies have been reported to use Gorgona and Gorgonilla Islands and their surroundings as a roosting area, and this was observed also during this study. Although no occurrences of roosting were observed in Juanchincho, there were a large number of this species roosting in the other Gorgonilla Islands and El Horno. Competition with blue-footed boobies, if present, may therefore be less intense in Juanchincho leaving brown booby pairs with more energy resources to invest in reproduction. In the Juanchincho sites, breeding took place over the whole year in contrast with sites around Gorgonilla Island and El Horno (Ospina-Alvarez 2008), indicating that it may be the nesting site of.
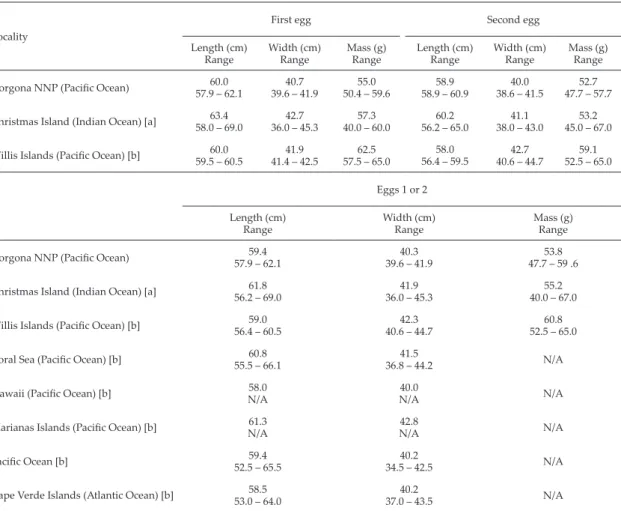
(7) 16. Ospina-Alvarez. Table 3. Brown booby egg dimensions in Gorgona NNP (this study) and other localities around the world. Sources: [a] Nelson (2005); [b] Serventy (1959). First egg Locality. Second egg. Length (cm) Range. Width (cm) Range. Mass (g) Range. Length (cm) Range. Width (cm) Range. Mass (g) Range. Gorgona NNP (Pacific Ocean). 60.0 57.9 – 62.1. 40.7 39.6 – 41.9. 55.0 50.4 – 59.6. 58.9 58.9 – 60.9. 40.0 38.6 – 41.5. 52.7 47.7 – 57.7. Christmas Island (Indian Ocean) [a]. 63.4 58.0 – 69.0. 42.7 36.0 – 45.3. 57.3 40.0 – 60.0. 60.2 56.2 – 65.0. 41.1 38.0 – 43.0. 53.2 45.0 – 67.0. Willis Islands (Pacific Ocean) [b]. 60.0 59.5 – 60.5. 41.9 41.4 – 42.5. 62.5 57.5 – 65.0. 58.0 56.4 – 59.5. 42.7 40.6 – 44.7. 59.1 52.5 – 65.0. Eggs 1 or 2 Length (cm) Range. Width (cm) Range. Mass (g) Range. Gorgona NNP (Pacific Ocean). 59.4 57.9 – 62.1. 40.3 39.6 – 41.9. 53.8 47.7 – 59 .6. Christmas Island (Indian Ocean) [a]. 61.8 56.2 – 69.0. 41.9 36.0 – 45.3. 55.2 40.0 – 67.0. Willis Islands (Pacific Ocean) [b]. 59.0 56.4 – 60.5. 42.3 40.6 – 44.7. 60.8 52.5 – 65.0. Coral Sea (Pacific Ocean) [b]. 60.8 55.5 – 66.1. 41.5 36.8 – 44.2. N/A. Hawaii (Pacific Ocean) [b]. 58.0 N/A. 40.0 N/A. N/A. Marianas Islands (Pacific Ocean) [b]. 61.3 N/A. 42.8 N/A. N/A. Pacific Ocean [b]. 59.4 52.5 – 65.5. 40.2 34.5 – 42.5. N/A. Cape Verde Islands (Atlantic Ocean) [b]. 58.5 53.0 – 64.0. 40.2 37.0 – 43.5. N/A. preference in Gorgona NNP for brown booby. Pairs which find an optimal site may be less inclined to abandon it and will likely remain to reproduce a second time. This may explain the extended breeding period in Juanchincho, and from this point of view, it is likely that this difference in site quality is maintained in subsequent years (Ospina-Alvarez 2008). My observations suggest that the “ideal” nest position for a brown booby would on an islet, preferably with the nest on a platform or on the highest point, and on a site that is relatively level and not too humid. The mean height of 7.9 m above sea level suggests nests are also built high enough to provide protection from waves. They also require sufficient space for landing (60 cm at least), and boobies appear to choose nest locations with 2 surrounding walls, grass and wind exposure (Table 4). An important characteristic of a protected area for the 21st century is that it helps maintain the diversity of ecosystems, species, genetic varieties. and ecological processes (McNeely 1994). Originally, Natural Parks were often viewed as delineated areas in which species could live in environments more optimal than in non-protected areas (IUCN 1969). However, fine-scale analyses of reproductive success between protected and non-protected areas to assess this assumption were rarely conducted (Schneider 2001). The structure, sizes and spacing of patches of habitat are each important in determining abundances of local populations and their rates of change (Underwood et al. 2000). Recent studies have emphasised the large variability in abundances of individual species at very small scales (Bell et al. 1993; Underwood et al. 2000). In some cases, the physical features of the habitat are sufficient to explain the patterns (Fairweather 1988; Archambault & Bourget 1996; Thompson et al. 1996) while in others, complex behavioural interactions among individuals are important in determining patterns of abundance at different spatial scales (e.g., Chapman 1994; Underwood et al..
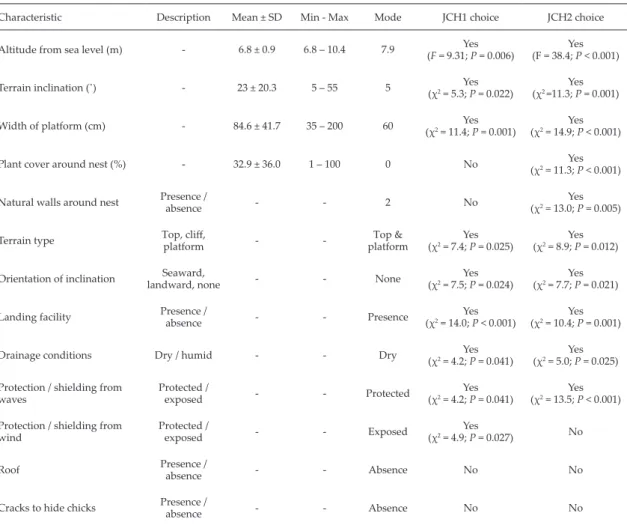
(8) Brown booby on a Neotropical island. 17. Table 4. Means, standard deviations, minimums and maximums for quantitative nest site variables and modes for quantitative and qualitative nest site variables for 34 brown boobies nests on the Juanchincho I and Juanchincho II colonies (Gorgona NNP). The overall means were calculated from 22 nest sites on Juanchincho II and 12 nest sites on Juanchincho I. Results of test for selection of each characteristic by nesting pairs is showed. Characteristic. Description. Mean ± SD. Min - Max. Mode. JCH1 choice. JCH2 choice. Altitude from sea level (m). -. 6.8 ± 0.9. 6.8 – 10.4. 7.9. Yes (F = 9.31; P = 0.006). Yes (F = 38.4; P < 0.001). Terrain inclination (�). -. 23 ± 20.3. 5 – 55. 5. Yes (χ2 = 5.3; P = 0.022). Yes (χ2 =11.3; P = 0.001). Width of platform (cm). -. 84.6 ± 41.7. 35 – 200. 60. Yes (χ2 = 11.4; P = 0.001). Yes (χ2 = 14.9; P < 0.001). Plant cover around nest (%). -. 32.9 ± 36.0. 1 – 100. 0. No. Yes (χ2 = 11.3; P < 0.001). Natural walls around nest. Presence / absence. -. -. 2. No. Yes (χ2 = 13.0; P = 0.005). Terrain type. Top, cliff, platform. -. -. Top & platform. Yes (χ2 = 7.4; P = 0.025). Yes (χ2 = 8.9; P = 0.012). Seaward, landward, none. -. -. None. Yes (χ2 = 7.5; P = 0.024). Yes (χ2 = 7.7; P = 0.021). Presence / absence. -. -. Presence. Yes (χ2 = 14.0; P < 0.001). Yes (χ2 = 10.4; P = 0.001). Dry / humid. -. -. Dry. Yes (χ2 = 4.2; P = 0.041). Yes (χ2 = 5.0; P = 0.025). Protection / shielding from waves. Protected / exposed. -. -. Protected. Yes (χ2 = 4.2; P = 0.041). Yes (χ2 = 13.5; P < 0.001). Protection / shielding from wind. Protected / exposed. -. -. Exposed. Yes (χ2 = 4.9; P = 0.027). No. Roof. Presence / absence. -. -. Absence. No. No. Cracks to hide chicks. Presence / absence. -. -. Absence. No. No. Orientation of inclination Landing facility Drainage conditions. 2000). Even protected areas host zones with highly variable quality, and this translates into differences in reserve effect between sites of the same NNP. These fine-grained characteristics should be taken into account when deciding the size of protected areas so as to improve the conservation of marine birds, especially when considering a colonial bird species with marked site-choice capabilities like S. l. etesiaca. Finally, I encourage research efforts to develop pattern models, aimed at explaining the interspecific competition, nest site and pair choice in S. l. etesiaca. My observations, even if just mensurative experiments (sensu Hurlbert 1984) provide a starting position to propose explanations or test theories. Another deeper approach is necessary to clarify the processes involved in the possible competition between S. l. etesiaca and the other species in. Gorgona NNP. If this competition exists, then a negative association between relevant variables in the species would have to be found. ACKNOWLEDGEMENTS The author wishes to acknowledge funding from Fondo para la Acción Ambiental de Colombia, Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales de Colombia, Gorgona Natural National Park, IDEA WILD, Prof. Luis German Naranjo, Samuele Tecchio and Jose Trancito Grueso. LITERATURE CITED Alberico, M. 1986. Los mamíferos. Pp. 193-207 In: Prahl, H.v.; Alberico, M. (eds.). Isla de Gorgona. Bogotá, Colombia: Textos Universitarios, Banco Popular. 252 pp. Allen, G.; Robertson, D. 1994. Fishes of the tropical eastern Pacific. Honolulu: University of Hawaii Press. 332 pp..
(9) 18. Ospina-Alvarez. Archambault, P.; Bourget, E. 1996. Scales of coastal heterogeneity and benthic intertidal species richness, diversity and abundance. Marine Ecology Progress Series 136: 111-121. Bell, G.; Lechowicz, M.J.; Appenzeller, A.; Chandler, M.; DeBlois, E.; Jackson, L.; Mackenzie, B.; Preziosi, R.; Schallenberg M.; Tinker, N. 1993. The spatial structure of the physical environment. Oecologia 96: 114-121. Cadena-Lopez, G. 2004. Distribución, abundancia y reproducción de las aves marinas (Pelecaniformes) en el Parque Nacional Natural Gorgona durante el año 2003. B. Sc. Thesis. Universidad del Valle, Cali, Colombia. Chambers J.M.; Cleveland, W.S.; Kleiner, B.; Tukey, P.A. 1983. Graphical methods for data analysis. Boston: Wadsworth & Brooks/Cole. 395 p. Chapman, M.G. 1994. Small- and broad-scale patterns of distribution of the upper-shore littorinid Nodilittorina pyramidalis in New South Wales. Australian Journal of Ecology 19: 83-95. Danchin, E.; Wagner, R.H. 1997. The evolution of coloniality: the emergence of new perspectives. Trends in Ecology & Evolution 12: 342-347. Fairweather, P.G. 1988. Predation creates haloes of bare space among prey on rocky seashores in New South Wales. Australian Journal of Ecology 13: 401-409. Froese, R.; Pauly, D. (eds). 2003. Fish Base. www.fishbase. org. Version 21/2003. Heatwole, H.; Abbott, P.; Jones, M. 1990. Egg-size and clutch size of the brown booby, Sula leucogaster, at Swain Reefs, Great Barrier Reef. Corella 14: 51–52. Heller E.; Snodgrass, R.E. 1901. Descriptions of two new species and three new subspecies of birds from the Eastern Pacific, collected by the Hopkins-Stanford expedition to the Galapagos Islands. Condor 3: 74–77. Hertel F.; Ballance, L.T. 1999. Wing ecomorphology of seabirds from Johnston Atoll. Condor 101: 549–556. Hurlbert, S.H. 1984. Pseudoreplication and the design of ecological field experiments. Ecological Monographs 54: 187-211. IUCN. 1969. Standards and nomenclature for protected areas. 10th General Assembly, New Delhi, India. Le Corre, M.; Jouventin, P. 1997. Kleptoparasitism in tropical seabirds: vulnerability and avoidance responses of a host species, the red-footed booby. Condor 99: 162-168. Lindsey T. 1986. The seabirds of Australia. North Ryde, Australia: Angus & Robertson Publishers. 518 p. McNeely, J. 1994. Protected areas for the 21st century: working to provide benefits to society. Biodiversity and Conservation 3: 390-405.. Murphy, R.C.; Jaques, F.L. 1936. Oceanic birds of South America: a study of species of the related coasts and seas, including the American quadrant of Antarctica, based upon the Brewster-Sanford Collection in the American Museum of Natural History. New York: American Museum of Natural History. 640 pp. Naranjo, L.G.; Aparicio, A,; Falk, P.E. 2001. Evaluación de áreas importantes para aves marinas y playeras en el Litoral Pacífico Colombiano. Cali, Colombia: Fondo para la protección del Medio Ambiente (FEN). 87 pp. Nelson, J.B. 1978. The Sulidae — gannets and boobies. Aberdeen University, Study Series 154. Oxford: Oxford University Press. 1024 pp. Nelson, J.B. 2005. Pelicans, cormorants, and their relatives: the Pelecaniformes. Oxford: Oxford University Press, 680 p. Ortiz-von Halle, B. 1990. Aspectos generales de la comunidad de aves. Pages 215-235 In: Aguirre, J.; Rangel, O. (eds.). Biota y ecosistemas de Gorgona. Bogotá, Colombia: Fondo para la protección del Medio Ambiente (FEN). 303 pp. Ospina-Alvarez, A. 2008.Coloniality of brown booby (Sula leucogaster) in Gorgona National Natural Park, eastern tropical Pacific. Ornitología Neotropical 19: 517 - 529. Schneider, D. 2001. The rise of the concept of scale in ecology. BioScience 51: 545-553. Serventy, D.L. 1959. Birds of Willis Island. Emu 59: 167 – 176. Stager K.E. 1964. The birds of Clipperton Island, eastern Pacific. Condor 66: 357–371. Thompson, R.C.; Wilson, B.J.; Tobin, M.L.; Hill, A.S.; Hawkins, S.J. 1996. Biologically generated habitat provision and diversity of rocky shore organisms at a hierarchy of spatial scales. Journal of Experimental Marine Biology and Ecology 202: 73-84. Underwood, A.; Chapman, M.; Connell, S. 2000. Observations in ecology: you can’t make progress on processes without understanding the patterns. Journal of Experimental Marine Biology and Ecology 250: 97-115. Weimerskirch, H.; Shaffer, S.A.; Tremblay, Y.; Costa, D.P.; Gadenne, H.; Kato, A.; Ropert-Coudert, Y.; Sato, K.; Aurioles, D. 2009. Species- and sex-specific differences in foraging behaviour and foraging zones in bluefooted and brown boobies in the Gulf of California. Marine Ecology Progress Series 391: 267–278. Woodward, P.W. 1972. The natural history of Kure Atoll, north-western Hawaiian Islands. Atoll Research Bulletin 164: 1-317..
(10)
Figure

Documento similar