Análisis in silico de la Competencia y Coexistencia entre Streptococcus sanguinis y Streptococcus mutans en el biofilm dental
98
0
0
Texto completo
(2) UNIVERSIDAD NACIONAL “PEDRO RUIZ GALLO” FACULTAD DE CIENCIAS BIOLÓGICAS DEPARTAMENTO ACADÉMICO DE BIOLOGÍA. TÍTULO: Análisis in silico de la Competencia y Coexistencia entre. Streptococcus sanguinis y Streptococcus mutans en el biofilm dental.. TESIS PARA OPTAR EL TÍTULO PROFESIONAL DE: LICENCIADO EN BIOLOGÍA PRESENTADO POR: Br. Percy Omar Túllume Vergara. APROBADO POR:. …………………………………………. Mario Cecilio Moreno Mantilla JURADO, Presidente. …………………..………………………… Martha Vergara Espinoza JURADO, Secretaria. ……………….…………………………. Félix Gonzales Flores JURADO, Vocal. ..…………………….……………… Pedro Jorge Chimoy Effio PATROCINADOR. Lambayeque – Perú 2014 II.
(3) DEDICATORIA: Este trabajo de investigación está dedicado a la memoria de las personas que me apoyaron en toda mi etapa estudiantil, mi madre Carmen Túllume Mechán y por sus consejos a mi abuelita Rosaría Mechán Gonzales (Q.P.D.). III.
(4) AGRADECIMIENTOS Al culminar el presente documento siento una especial satisfacción porque representa el final a un largo y arduo trabajo de mañana, tarde y a veces de noche para entregar este documento. Mis sinceros agradecimientos al Dr. Pedro Chimoy Effio, patrocinador del presente trabajo, por su orientación y siempre bien dirigidos consejos a lo largo del desarrollo de mi desarrollo como profesional. Gracias amigo. Agradecimientos a mis Co-asesores Dra. Wendy Gonzales D. por su orientación y siempre bien dirigidos consejos a lo largo del trabajo de investigación, al Dr. (c) Juan Pablo Cárdenas quienes me apoyaron y fueron los pilares de este humilde pero sacrificado trabajo de memoria de pregrado, me brindaron ayuda en la redacción de los resultados y discusión de los mismos ambas personas del hermano país de Chile. Y al artífice de la idea Dr. Rodrigo Giacaman, Odontólogo con sus comentarios acertados acerca de la investigación. A mi madre, Carmen por brindarme todo el apoyo necesario para seguir adelante, y sobre todo por inculcar valores y consejos en mi formación. A la Dra. Blga. Delia Sánchez de CERPER S.A. por su constante apoyo moral para terminar la presente tesis. Gracias a esos grandes amigos del CEGENP-UNPRG, personas tan especial, que compartimos carpetas en los cursos de Bioinformática que organizamos, Sinceramente gracias. Y en especial gracias a ti cariño Maribel Z. por tu paciencia, comprensión y por ayudarme en los momentos más estresantes del trabajo. Mi más sincera y profunda gratitud a todos. Espero no olvidarme de nadie.. IV.
(5) CONTENIDO ABSTRACT I. II. III.. INTRODUCCIÓN ANTECEDENTES BIBLIOGRÁFICOS MATERIALES Y MÉTODOS 3.1 Material 3.1.1 Muestras 3.2 Métodos 3.2.1 Tipo de estudio 3.2.2 Metodología 3.2.2.1 Caracterización de la información genómica mediante categorización en la Base de datos de COGs (Clusters of Orthologous Groups) y análisis comparativo mediante Mauve (Primera Fase). 3.2.2.2 Identificacion de rutas metabolicas especificas mediante KEEG entre S. mutans UA159 y S. sanguinis SK36 (Segunda Fase). 3.2.2.3 Descripción de un modelo de via se señalización celular entre S. mutans UA159 y S. sanguinis SK36 (Tercera Fase). 3.2.2.4 Realización de comparaciones de proteínas importantes en la competencia (Cuarta Fase). 3.2.3 Análisis de los datos.. IV.. RESULTADOS 4.1 Comparación genómica de las categorías funcionales COGs, análisis comparativo y identificacion de rutas metabolicas especificas mediante KEEG. 4.2 Identificación de rutas metabolicas especificas mediante KEEG entre S. mutans UA159 y S. sanguinis SK36. 4.3 Descripción y reconstrucción del sistema de Quórum sénsing implicado en la transformación genética. 4.4 Comparación de proteínas importantes en la competencia entre S. mutans UA159 y S. sanguinis SK36.. V. VI. VII. VIII. IX. X. XI.. DISCUSIÓN CONCLUSIONES RECOMENDACIONES GLOSARIO RESUMEN REFERENCIAS BIBLIOGRÁFICAS ANEXOS V.
(6) Abstract. Tooth decay is one of the most common diseases in humans and affects the quality of life in people of all ages. Moreover, it demands high resources of health services. In order to reduce tooth decay, a better understanding of the pathogenic role of microorganisms in dental diseases is needed. Studies have revealed the organisms who colonize the oral cavity and how they coexist. S. sanguinis is a primary colonizer in the oral cavity and produce quantities of hydrogen peroxide (H2O2) which inhibit the growth of S. mutans. In the other hand, S. mutans produces antimicrobial peptides (mutacins) which appear to play a role in the competition against S. sanguinis. The aim of this research was to predict the metabolic model of competition and coexistence between S.sanguinis SK36 and S. mutans UA159 in the dental biofilm. Two genomes were analyzed by bioinformatics tools. Our results demonstrate that the competition and the coexistence is not only for the production of H2O2 in S. sanguinis, but their genome encodes several proteins as glutathione peroxidase, V-ATPase enzyme complex, proteins relates in the quorum sensing system and PrtS (serine protease), which could give them several advantages. However, the S. mutans genome has a complex group of bacteriocins (mutacins). We founded seven, previously unreported, possible mutacins. As we described, that could be the unusuals ecological advantages in each specie to survive in the stressful and fluctuating dental biofilm environment. The model “in silico” about competition and coexistence between S. sanguinis SK36 and S. mutans UA159 has intended to create an useful framework to start an experimental research in the validation process.. VI.
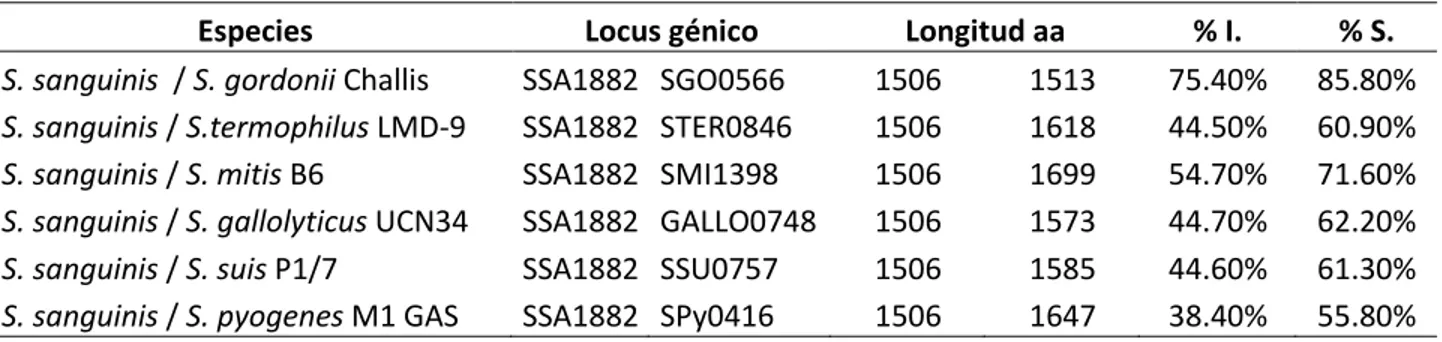
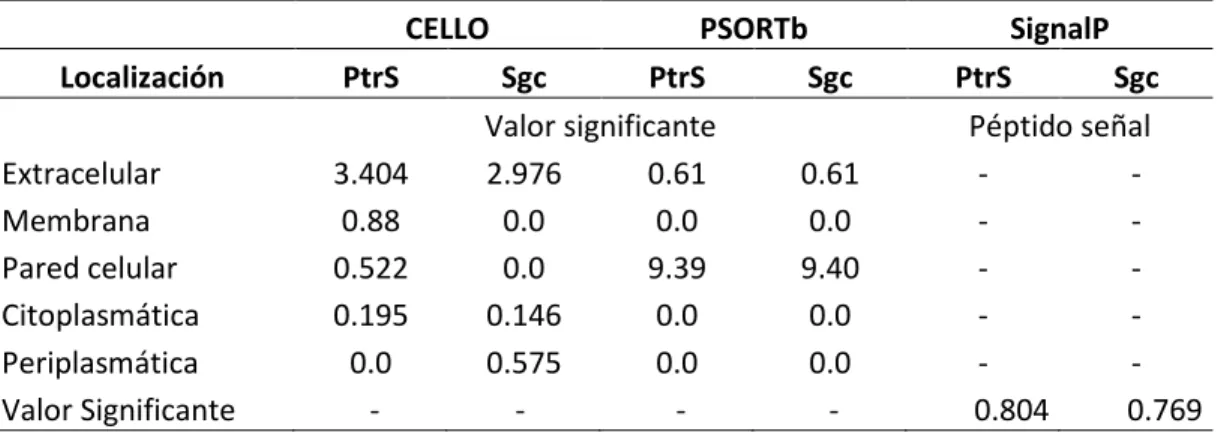
(7) LISTA DE TABLAS. Tabla 1. Comparación de las secuencias de proteínas Com involucradas en la formación del sistema de Quórum sensing de S. sanguinis y S. mutans, caracterizados molecularmente en S. pneumoniae D39. Tabla 2. Comparación de la similitud del sistema de ATPasas-F presentes en ambas especies. Tabla 3. Sistema de ATPasas F y V presentes en ambos genomas. Tabla 4. Número de genes regulados por la proteína CcpA entre S. sanguinis y S. mutans, según Regprecise. Tabla 5. Alineamiento de las secuencias de proteínas serin proteasas en el género Streptococcus sp. mediante Matcher v.6.3.1. Tabla 6. Análisis de localización celular de sgc y PrtS mediante CELLO, PSORTb y SignalP. Tabla 7. Posibles mutacinas con motivo -GG- ubicadas mediante software Artemis en el genoma de S. mutans UA159.. VII.
(8) LISTA DE FIGURAS Figura 01. Ensayo in vitro de la competencia y coexistencia de S. sanguinis y S. mutans en el biofilm (biopelícula) dental. Figura 02. Bloques colineares locales (LCBs) entre dos secuencias cromosómicas de S. mutans y S. sanguinis. Figura 03. Las categorías funcionales COGs entre S. mutans y S. sanguinis. Figura 04. Predicción del mapa del metabolismo global que comparten S. sanguinis y S. mutans. Figura 05. Reconstrucción del mapa del metabolismo del piruvato entre S. mutans y S. sanguinis, mediante análisis de sus genomas. Figura 06. Comparación de la vía de utilización del glutatión entre S. sanguinis y S. mutans. Figura 07. Alineamiento múltiple y alineamiento local de glutatión peroxidasa EC 1.11.1.9. Figura 08. Análisis filogenético de la enzima glutatión peroxidasa de S. sanguinis con respecto a otros organismos. Figura 09. Histograma del análisis comparativo de las secuencias de proteínas Com involucradas en el sistema de Quórum sensing entre S. sanguinis y S. mutans. Figura 10. Modelo propuesto para la transformación genética mediada por el sistema de Quórum sensing en S. sanguinis y S. mutans. Figura 11. Organización genómica propuesta para los operón unc y atp de S. sanguinis y S. mutans. Figura 12. Organización genómica propuesta para el posible operón ntp de S. sanguinis SK36. Figura 13. El alineamiento múltiple de secuencia (MSA) de la proteína CcpA en especies del género Streptococcus. Figura 14. Comparación de las vías metabólicas reguladas por CcpA en S. mutans y S. sanguinis. Figura 15. Alineamiento múltiple de secuencia (MSA) incompleto de posibles bacteriocinas (mutacinas) de S. mutans UA159. Figura 16. Esquema del predictivo Modelo de coexistencia en S. mutans y S. sanguinis.. VIII.
(9) LISTA DE ANEXOS Anexo 01: Tabla 08. Tabla general de proteínas analizadas en el presente estudio, con significancia estadística a través del parámetro e-value y % de identidad. Tabla 09. Numero y porcentaje de genes para las categorías funcionales COGs de S. sanguinis SK36 y S. mutans UA159. Anexo 02: Figura 16. Diagrama de Veen. Comparación total de genes (ORFs) entre S. sanguinis SK36 que presenta 1960 ORFs y S. mutans UA159 presenta 2253 ORFs Anexo 03: Figura 17. Flujograma del trabajo bioinformático (análisis in silico) el cual comprende cuatro fases detalladas en la metodología del trabajo. Anexo 04: Lista de web de Base de datos (BD) y softwares utilizados en el estudio bioinformático.. IX.
(10) I.. INTRODUCCIÓN. La caries dental es una enfermedad infecciosa, transmisible e inducida por la placa bacteriana, asociada a un huésped susceptible portador de una flora odontopatógena que produce ácidos, a través de la fermentación de carbohidratos que provocan la disolución y destrucción localizada de los tejidos duros del diente (Loesche, 1982). Además la caries es una de las enfermedades más comunes en el ser humano, aunque rara vez amenaza la vida, es un problema que afecta la calidad de vida de las personas (Sathyanarayanan et al., 2002) y demanda altos recursos de parte de los proveedores de servicios de salud. Esta enfermedad se asocia con la presencia de un biofilm bacteriano dependiente de azúcares fermentables de la dieta (Vadiakas, 2008). Con el fin de disminuir la prevalencia de caries, una mejor comprensión del papel patogénico de los microorganismos en las enfermedades dentales es necesario (Marsh, 2003). Se plantea en estudios que la caries dental la padecen aproximadamente el 96% en el mundo y un 99% de la población de América Latina (Rodríguez et al., 2006). La condición de Salud Bucal en el Perú, atraviesa una situación crítica debido a la alta prevalencia de enfermedades odontoestomatológicas, así tenemos que la prevalencia de caries dental es 95%, enfermedad periodontal 85%, constituyendo un problema de salud pública. Además en lo que se refiere a caries dental el índice de dientes cariados, perdidos y obturados (CPOd), a los 12 años es de aproximadamente 6 CPOd, ubicándose según la OPS en un país en estado de emergencia (MINSA, 2012). Asimismo, las interacciones cooperativas entre especies probablemente juegan un rol importante en el desarrollo del biofilm dental, las interacciones antagonistas entre diferentes especies pueden ser equitativamente importantes dadas las condiciones de la cavidad oral (Kreth et al., 2005).. 1.
(11) Estudios previos realizados permiten conocer los organismos involucrados en la coexistencia y el antagonismo dentro de los cuales están los estreptococos orales, como S. sanguinis, que produce cantidades de peróxido de hidrogeno (H2O2) que inhiben el crecimiento de S. mutans (Kreth et al., 2008). Sin embargo, la evidencia molecular es aun débil y requiere de mayor investigación. Aunque S. sanguinis también produce péptidos antimicrobianos (sanguicina) (Deng et al., 2004), estos péptidos no podrian jugar un papel en la competencia contra S. mutans. Desde el punto de vista clínico, el predominio de S. sanguinis sobre S. mutans en el biofilm dental podría asociarse con una menor prevalencia de caries en niños (Ge et al., 2008). De otro modo, la Bioinformática es impulsada por avances en tecnologías de alto rendimiento biológico y cada vez mayores números de genomas secuenciados. La construcción de modelos in silico a escala genómica proporciona poderosas herramientas para investigar una amplia gama de sistemas biológicos. Esta permite el análisis masivo y riguroso de procesos celulares realizados por un microorganismo, generando predicciones sobre posibles eventos moleculares con un mayor grado de confiabilidad que lo proveniente por la intuición. Incluso, permiten predecir genes esenciales en todo el genoma de un organismo favoreciendo la selección de aquellas moléculas que puedan ser blancos de la acción de fármacos (Roberts et al., 2007), para posteriormente ser evaluadas experimentalmente in vitro e in vivo (Fatumo et al., 2009). El campo bioinformático se está tornando más amplio: de una docena de genomas secuenciados en el año 1997 aumentaban a noventa genomas seuenciados cinco años mas tarde y se siguen aún secuenciando, aumentando esta cifra exponencialmente. Es por eso que la comparación de múltiples genomas (genómica comparativa) se ha convertido en un tema frontera de investigación (Chain et al., 2003). Como consecuencia de los avances tecnológicos destinados al campo de la genómica se ha logrado proyectar a la comunidad científica, la secuenciación de genomas de cualquier organismo de interés científico o económico. Hoy en día existen más de 4 122 genomas bacterianos completamente secuenciados y 16 255 proyectos más en curso (GOLD: http://.genomesonline.org, actualizado a Agosto del 2013). 2.
(12) Estrategias basadas en genómica comparativa de múltiples cromosomas de especies filogenéticamente distintas pueden proporcionar huellas acerca de las presión selectiva que regulan a determinado grupos de genes que pueden ofrecer respuestas sobre los mecanismos de la evolución, variación fenotípica y del material genético externo a través de la transferencia genética horizontal. Esto ha dado buenos resultados en comparaciones entre especies estrechamente relacionadas, como el ser humano y el chimpancé (Nobrega & Pennacchio, 2003). Hasta la fecha, no se presentan estudios a nivel bioinformático para entender las rutas metabólicas y vías de señalización que involucran las interacciones moleculares de coexistencia y antagonismo en el biofilm. Por lo tanto, es necesario analizar y dilucidar la genómica de la bacteria comensal del biofilm dental S. sanguinis SK36 y de la bacteria sindicada como la más cariogénica S. mutans UA159. Según lo expuesto, se formuló el siguiente problema ¿Cuál es modelo metabólico general de la competencia y coexistencia entre S. sanguinis SK36 y S. mutans UA159 en el biofilm dental? Para solucionar este problema se plantearon los siguientes objetivos: Caracterizar la información obtenida del genoma de Streptococcus sanguinis SK36 y Streptococcus mutans UA159 para los genes expresados según categorización existente, mediante la base de datos bioinformáticos COGs. Identificar las rutas metabólicas específicas de Streptococcus sanguinis SK36 y Streptococcus mutans UA159, mediante KEGG. Describir el modelo de vía de señalización entre Streptococcus sanguinis SK36 y Streptococcus mutans UA159 en base a información bibliográfica y experimental con el servidor Pubmed con microorganismos relacionados filogenéticamente. Comparar proteínas significativas involucradas en la competencia y coexistencia en el genoma de Streptococcus sanguinis SK36 y Streptococcus mutans UA159. En el presente estudio se usó el área de genómica comparativa que presenta alto impacto biotecnológico y provee de nuevos datos para incrementar conocimientos acerca de las interacciones moleculares dilucidando las rutas metabólicas y vías de señalización que involucran las interacciones moleculares de antagonismo de S. sanguinis SK36 y S. mutans UA159. 3.
(13) II.. 2.1. ANTECEDENTES BIBLIOGRÁFICOS. Microorganismos Orales. La cavidad bucal es uno de los hábitats microbianos más heterogéneos y. complejos del cuerpo (Madigan et al., 2003). La microbiota oral funciona como una parte de la defensa del huésped, actuando como una barrera al promover competencia por los nutrientes esenciales y crear condiciones desfavorables para los organismos exógenos que pueden ser patógenos para el huésped. Se han encontrado más de 700 taxones en la cavidad oral, sin embargo no están presentes en la misma boca (Aas et al., 2005). La composición también varía en los diferentes sitios de la cavidad oral, por ejemplo una gran carga bacteriana mucho más diversa esta en el dorso de la lengua. La mayoría de estos microbios son inofensivos y están en equilibrio, pero bajo ciertas condiciones algunos pueden causar infecciones bucales como la caries o enfermedad periodontal (Sakamoto et al., 2005). Se han realizado estudios de análisis molecular asociado a la microflora bacteriana que causa la caries por el método ARNr 16S. Se demostró que la dentina cariada está dominada por bacterias grampositivas, en particular los géneros Actinomyces, Lactobacillus, Propionibacterium y Streptococcus. Además se identificaron 95 taxones, de los cuales los predominantes en el análisis molecular fueron S. mutans (16%), Lactobacillus gasseri/jhonsonii (13%) y Lactobacillus rhamnosus (8%). (Munson et al., 2004). Streptococcus es un género dominante en la cavidad oral humana, representando aproximadamente el 20% de las más de 700 especies de bacterias que han sido identificadas y alrededor del 80% de los colonizadores iniciales del biofilm (Rosan y Lamont, 2000).. 4.
(14) Los estreptococos están presentes en todas las placas dentales, se incrementan en número cuando se desarrollan procesos cariosos y se encuentran más frecuentemente sobre superficies cariadas que sobre superficies libres de caries (MacDonald, 1962). El género Streptococcus reagrupa especies muy diferentes por sus características quimiotaxonómicas y por su poder patógeno: Grupo S. mutans: S. mutans, S. sobrinus, S. cricetus, S. rattus, S. ferus, S. macacae y S. downei; Grupo S. anginosus: S. anginosus, S. constellatus y S. intermediun; Grupo S. mitis: S. sanguinis, S. parasanguis, S. gordonii, S. mitis, S. oralis, y S. cirsta; Grupo S. salivarius: S. salivarius y S. vestibularis (Mouton et al., 1995). Así mismo tenemos estreptococos orales que son compatibles con la salud (por ejemplo, las especies Streptococcus gordonii y Streptococcus sanguinis) y patógenos (por ejemplo, el cariogénico del grupo, Streptococcus mutans). Debido a que los estreptococos tienen similitud en los requerimientos metabólicos, han desarrollado estrategias de defensa que conducen al antagonismo entre especies (conocido como interferencia bacteriana) (Kreth et al., 2008). 2.2. Streptococcus mutans. El nombre de Streptococcus mutans fue dado en 1924 por J. K. Clarke a los. estreptococos que se habían aislado en lesiones cariosas, pero no fue hasta los años sesenta del siglo XX que se le prestó mayor atención. Se realizaron estudios, sobre la transmisión de la caries en animales donde, se observó la analogía entre los estreptococos cariogénicos y el S. mutans identificado de J.K. Clarke. En la actualidad de las 7 especies del grupo de Streptococcus mutans se distinguen 2 humanas: S. mutans y S. sobrinus y 5 animales: S. cricetus, S. rattus, S. ferus, S. macacae y S. downei (Fitzgerald et al., 1960). S. mutans da nombre a un grupo de siete especies estrechamente relacionadas referidas colectivamente como Streptococcus mutans.. 5.
(15) Los hábitats principales de S. mutans son la boca, la faringe y el intestino (Loesche, 1986), S. mutans es el principal agente patógeno asociado a la caries dental, vive casi exclusivamente en los biofilm densamente poblados que se forman en la superficie del diente. Grandes cantidades de ácido orgánico son producidas por las células embebidas en el biofilm, creando un medio ácido que erosiona la superficie del diente y por lo tanto provoca la caries. Por lo general, la aparición de S. mutans en las cavidades de los dientes es seguido por la caries después de 6-24 meses (Mayooran et al., 2000). S. mutans presenta habilidades como aciduricidad/acidogenicidad, la formación de biofilms, la competencia natural y la producción de bacteriocinas (mutacinas) son cruciales para su virulencia (Banas, 2004; Kuramitsu, 1993). En consecuencia, la virulencia de S. mutans es causado por su capacidad para formar biofilm, producir ácidos orgánicos y tolerar el estrés ambiental, particularmente bajo condiciones de bajo pH (Ajdic et al., 2002). En el año 2002 fue secuenciado el genoma completo de S. mutans cepa UA159 el cual está organizado en un cromosoma circular de 2, 030 936 pares de bases. El promedio de contenido GC es de 36.82 % similar a otros organismos con bajos índices de GC. Además posee 62 tRNAs y cinco operones rRNA. Las secuencias codificantes de proteínas representan el 85,66 % del genoma y está organizado en 1963 ORFs, con promedio de longitud de genes de 885 bases. La clave de su estrategia de supervivencia radica en el metabolismo de carbohidratos, el cual es capaz de metabolizar una amplia gama de azúcares. Presenta genes para el transporte y metabolismo de glucosa, sacarosa, lactosa, galactosa, manosa, celobiosa, trehalosa, rafinosa, ribulosa, maltosa/maltodextrina, β-glucosida, melobiosa, isomaltosácaridos y posiblemente sorbosa (Ajdic et al., 2002).. 6.
(16) 2.3. Streptococcus sanguinis. S. sanguinis es un colonizador primario de la cavidad bucal humana y uno de los. microorganismos que colonizan los dientes (Kuramitsu et al., 2007). Es una bacteria grampositiva perteneciente al grupo de los Estreptococos viridans (Kreth et al., 2008). S. sanguinis sirve como una cuerda donde pueden fijarse otros microorganismos orales que colonizan la superficie del diente formando la placa dental (Yamaguchi et al., 2006). La patogenicidad de este organismo no se limita a las infecciones orales, sino que se extiende y provoca el peligro de la vida, presentándose en una enfermedad endovascular llamada endocarditis. La endocarditis es una infección grave de las válvulas o el revestimiento del corazón (Turner et al., 2009). Al igual que los otros estreptococos orales, esta bacteria produce α-hemólisis en agar sangre, una característica vinculada a la capacidad de los Estreptococos viridians para oxidar la hemoglobina de los eritrocitos por la secreción de H2O2 (Barnard et al., 1996). Los avances en el campo de la bioinformática han acelerado el progreso de la investigación médica en la lucha contra enfermedades. Es así que hace unos años el genoma de S. sanguinis fue secuenciado (Xu et al., 2007). El genoma de S. sanguinis presenta las siguientes características generales: está organizado en un cromosoma circular de 2, 388 435 pares de bases, el promedio de G+C es de 43,40% el cual es alto comparado con otros genomas de especies de estreptococos (35,62 a 39,72 %). Además el genoma contiene genes que codifican 2 274 proteínas predichas, de las cuales el 89% exhiben similitud significativa a proteínas de otros organismos y el 22% son proteínas hipotéticamente conservadas.. 7.
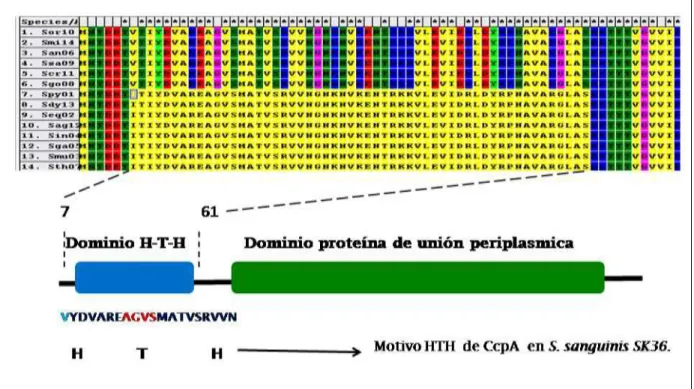
(17) 2.4 Elementos involucrados en la competencia dentro de la biopelícula dental. La placa dental, donde se encuentran cientos de especies bacterianas, es el mejor ejemplo de un biofilm natural en el cuerpo humano. Pese a que se han realizado muchos estudios en la formación de biofilm monoespecificos, estos no representan verdaderamente la dinámica del biofilm oral encontrado en la boca. El biofilm oral muestra una intensa competencia así como sinergismo metabólico entre varias las especies microbianas constituyentes. Nobbs et al., (2009) indica que S. mutans, S. gordonii y otros estreptococos orales forman fácilmente el. biofilm. de una especie en el. laboratorio y en una variedad de superficies. La formación de biofilm depende de actividades de loci genéticos múltiples. Es así, que se han estudiado mutaciones en genes que codifican la formación de biofilm y los componentes del sistema quórum sensing, Por ejemplo, mutaciones en los genes comCDE (Li et al., 2002); TCSS, vicRK (Senadheera et al., 2005) y genes que codifican para permeasas de iones metálicos (Rolerson et al., 2006) pueden afectar a S. mutans en la formación del biofilm. Asimismo, la formación optima del biofilm por parte de S. gordonii y S. sanguinis también requiere de múltiples productos génicos incluyendo sortasas (Nobbs et al., 2007) además de las proteínas involucrados con el metabolismo de βglucósido y el control de catabolitos (Kilic et al., 2004; Loo et al., 2003), biosíntesis de nucleótidos (Ge et al., 2008), el estrés oxidativo (Kuboniwa, et al., 2006), y detección ambiental y señalización (Gilmore et al., 2003). Uehara et al, (2006) estableció sustancias que regulan interacciones entre especies. Es así que la producción de H2O2 está relacionada con la competencia y la convivencia dentro de las comunidades microbianas como los biofilms orales. Los estreptococos son capaces de producir sustancias inhibidoras tales como H 2O2 para reducir el crecimiento de microorganismos co-residentes por ejemplo, S. sanguinis puede producir H2O2, que inhibe el crecimiento de Staphylococcus aureus. Díaz et al., (2006) y después Kreth et al, (2008) indican que los competidores como los estreptococos del. 8.
(18) grupo Mitis colonizan en un principio el biofilm dental, crecen más rápido, y puede generar H2O2, que es un potente inhibidor del crecimiento del grupo S. mutans. Becker et al., (2002) realizó estudios in vivo enfocados en S. sanguinis donde se considera inofensivo en la cavidad oral. En individuos sanos se presenta un número significativamente elevado en la placa dental, respecto a los individuos con caries que prácticamente no tienen niveles detectables de S. sanguinis. En efecto, S. sanguinis y el cariogénico S. mutans son mutuamente antagónicas, el crecimiento de S. mutans se retrasa cuando S. sanguinis está presente en gran abundancia en la cavidad bucal en seres humanos (Caufield et al., 2000). Por otro lado Woodruff et al., (1998) indican que las células bacterianas competentes tratan de generar sustancias para inhibir el crecimiento de otras y colonizar ambientes con fluctuaciones como el biofilm oral. S. mutans, junto con muchas otras bacterias. grampositivas. producen. pequeños. péptidos antibacterianos. llamadas. bacteriocinas. Las bacteriocinas producidas por S. mutans, llamadas mutacinas, pueden proporcionar la fuerza selectiva necesaria para la colonización inicial o sostenimiento en la placa dental de ese patógeno. En condiciones donde las células tienen la energía suficiente para competir, pero no suficiente alimento para un crecimiento óptimo se activa la producción de mutacina (por parte de S. mutans) o H2O2 (por parte de S. sanguinis). Desde el punto de vista económico de la célula, la síntesis de mutacina y H2O2 se relaciona con el equilibrio entre el costo y beneficio. La biosíntesis de mutacinas I y IV es un proceso energéticamente costoso. Para mutacina I, por lo menos 11 productos genéticos son necesarios de sintetizar para producir una molécula de mutacina funcional (Qi et al., 2001), y para mutacina IV, se requieren por lo menos cinco productos génicos. Estas estrategias bien reguladas son necesarias para la supervivencia y perpetuación de la especie en una comunidad multiespecifica en condiciones naturales como es la cavidad oral donde los ciclos de abundancia, hambruna y fluctuaciones de pH son habituales y ocurren repetidamente durante el día (Kuramitsu et al, 2007). 9.
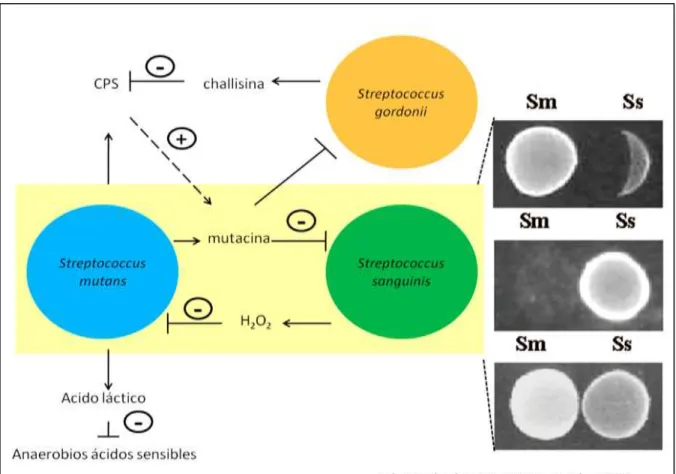
(19) Figura 01. Ensayo in vitro de la competencia y coexistencia de S. sanguinis y S. mutans en el biofilm (biopelícula) dental. Estudios realizado por Kuramitsu et al., (2007). Donde indica el antagonismo de ambas especies por H2O2 o por mutacinas. Además S. gordonii produce una proteína challisina que inactiva los péptidos estimulantes de la competencia (CPS). S. mutans produce altas cantidades de ácido producto de su metabolismo fermentativo inhibiendo el crecimiento de especies sensibles al ácido. En la parte de la derecha se muestran las inhibiciones: superior (S. mutans sobre S. sanguinis), medio (S. sanguinis sobre S. mutans) y inferior (coexistencia entre S. mutans y S. sanguinis).. 10.
(20) Kreth et al, (2008) demostró en ensayos in vitro que la disponibilidad de oxígeno y de glucosa sustancialmente influyen en la competición entre especies de estreptococos. S. mutans por ejemplo, es capaz de producir mutacinas inhibidoras del crecimiento, independientemente de la disponibilidad de oxígeno (Kreth et al., 2005). Socransky et al., (2002) indican que la tensión de oxígeno puede disminuir durante la formación del biofilm, debido a la limitación de la difusión y al rápido consumo de este. Con relación al H2O2 como un componente antimicrobiano, no se produce cuando las células crecen en condiciones anaeróbicas. Pero la producción de H2O2 bajo condiciones aeróbicas podría ser suficiente para funcionar en la competencia entre especies. El H2O2 está involucrado en importantes rutas metabólicas, en la potencial virulencia patogénica y la competencia interespecifica. Se han estudiado algunos genes como el gen spxB involucrado en la producción de H2O2 (Ramos-Montañez et al., 2008), pero su regulación y su relación con otros genes supuestamente implicados en la misma vía aún no están claras. La proteína anotada como spxB es una descarboxilasa que cataliza la conversión de piruvato, fosfato inorgánico (Pi) y oxígeno molecular (O2) a H2O2, dióxido de carbono (CO2) y acetil fosfato, que actúa como un dador de grupos fosforilo de alta energía (Ramos-Montañez et al., 2008) Piruvato+O2+Pi. CO2+H2O2+Acetil~P.. Zheng et al., (2011) demostró la importancia del regulador de CcpA (Proteína A de control del catabolito) en el control de la producción de H2O2, en la muerte celular dependiente de H2O2 y la liberación de ADN extracelular. Además indica que CcpA reprime la expresión de spx. En mutantes (∆CcpA) de S. sanguinis se observó mayor expresión de la proteína Spx, por lo tanto elevada producción de H2O2. La ventaja ecológica de esta inusual regulación dependiente de CcpA podría haber conducido a incrementar el número de S. sanguinis en comparación con S. gordonii en el biofilm dental y podría ser una de las razones por la que S. sanguinis es capaz de competir exitosamente con los patógenos antagonistas tales como S. mutans en el biofilm oral.. 11.
(21) Kreth et al., (2009) demuestran que el H2O2 contribuye en la liberación de ADN en S. sanguinis y S. gordonii, esto posiblemente apoye la formación de biofilms orales y podría facilitar el intercambio de material genético entre las cepas transformantes. La captación de ADN extracelular es otro mecanismo de competencia que está presente en los estreptococos. Podría ser un estado fisiológico transitorio durante el cual el ADN que ha sido tomado del medio ambiente, conduce a la transferencia horizontal de genes, a la plasticidad genómica y a la adquisición de nuevos los rasgos genéticos (Morrison, 1997). 2.5 Sistema de Quórum sénsing. La comunicación entre célula-célula es una actividad fundamental realizada por muchos tipos de células. Anteriormente, las bacterias fueron consideradas como células individuales que buscaban nutrientes y se multiplican independientemente. Sin embargo estudios nos muestran que las bacterias son capaces de tener actividades coordinadas como el Quórum sénsing (QS). Muchas bacterias, incluyendo los estreptococos son conocidos por regular diversos procesos fisiológicos a través de este mecanismo. En bacterias grampositivas, el sistema de QS consiste de tres componentes, un péptido señal y un sistema regulador de dos componentes, el cual presenta un sensor histidina quinasa anclado a la membrana y una proteína reguladora de respuesta (Kleerebezem et al., 1997). El QS regula la virulencia en muchos patógenos humanos (Bassler, 1999). Ejemplos de funciones del QS en bacterias grampositivas incluyen la formación de biofilms (Li et al., 2002a), el desarrollo de competencias genéticas (Havarstein et al., 1995), respuesta de tolerancia al ácido y producción de péptidos antimicrobianos (Kleerebezem & Quadri., 2001). Los detalles moleculares del desarrollo de competencias, incluyendo la identificación de genes que regulan las competencias, se analizaron inicialmente en Streptococcus pneumoniae (Johnsborg & Havarstein, 2009).. 12.
(22) Se sabe que el sistema QS, regula numerosas actividades fisiológicas en S. gordonii, S. pneumoniae, S. mutans (Cvitkovitch, 2001) y la inducción de factores de virulencia en Staphylococcus aureus (Kleerebezem & Quadri., 2001). El sistema QS puede influenciar en la formación de la estructura del biofilm en bacterias grampositivas (Loo et al., 2000). Un mutante defectivo del biofilm en S. gordonii, presenta un transposón insertado en el gen comD, que codifica una proteína sensor histidina quinasa requerida para la competencia genética. Esto podría dar a entender que la formación del biofilm por S. gordonii involucra la comunicación célula- célula a través del Sistema QS. La inactivación de algunos genes comCDE, que codifican el sistema QS resulta en la formación de un biofilm anormal (Li et al., 2002b). Se realizaron estudios en S. mutans demostrando que la formación de biofilm, tolerancia a ácido y la transformación genética son mediados por el sistema de QS (Cvitkovitch et al., 2003). El sistema QS, para poder activarse depende de péptidos estimulantes de la competencia (CSP). Los CSP extracelulares (ambientales) son detectados por el sistema de dos componentes (TCSS), y los CSP a concentración umbral activan la respuesta celular (Pestova et al., 1996). En S. pneumoniae, el desarrollo de la competencia requiere de la producción de CSP, estos son codificados por genes comC cuyos productos son secretados al medio extracelular por proteínas de transporte ComAB (Hui & Morrison, 1991). Se ha reconocido que la señal de QS en S. pneumoniae es iniciada, a través de la actividad de un modulador global de la transcripción, el gen comX el cual es un factor sigma alternativo durante el desarrollo de la competencia genética (Lee y Morrison, 1999). La transcripción del gen comX, es regulado por comE regulador de respuesta del sistema de QS (Ween et al., 1999). El factor sigma ComX, inicia la transcripción de operones específicos de la competencia involucrados en la recombinación y captación de ADN por reconocimiento de la caja com, que presenta la secuencia consenso (TACGAATA) en estas regiones promotoras (Lee & Morrison, 1999).. 13.
(23) III. 3.1.. MATERIALES Y MÉTODOS.. MATERIAL. 3.1.1 Muestras El estudio se realizó mediante el análisis del genoma de la cepa Streptococcus mutans UA159, aislado clínico del serotipo C, de la Universidad de Alabama, Birgmingham, EE. UU. con Número de accesión: AE014133 en GenBank y el genoma de la cepa Streptococcus sanguinis SK36, de Virginia Commonwealth University con RefSeq NC_009009.1. Sus secuencias genómicas se encuentran depositadas en GenBank y son de. libre acceso en NCBI (National Center for Biotechnology Information). Disponible en: (http://www.ncbi.nlm.nih.gov/).. 3.2.. MÉTODO. 3.2.1 Tipo de estudio. El presente informe es una investigación descriptiva, según el método de reconstrucción metabólica propuesto por Valdés et al., 2003 con modificaciones por el autor. 3.2.2. Metodología La capacitación para realizar el presente trabajo se logró mediante el manejo de bases de datos y software en bioinformática específicamente en genómica y del análisis de los datos arrojados con ayuda de libros de la especialidad. El trabajo consistió en ir de los detalles genómicos (generales) a los detalles específicos con proteínas significativas indicadoras de competencia.. 14.
(24) 3.2.2.1 Caracterización de la información genómica mediante categorización en la Base de datos de COGs (Clusters of Orthologous Groups) y análisis comparativo mediante Mauve. La primera estrategia (primera fase) consistió en realizar un análisis comparativo de los dos genomas basados en la secuencias genómicas depositadas en GenBank para lo cual se obtuvo la versión más reciente. En este paso se utilizó los software MAUVE v. 2.3.1 y Artemis v.13.2.0, los cuales presentan. una. excelente interfase gráfica para buscar regiones conservadas (sintenia) y no conservadas. (especificas). presentes. en. ambos. microorganismos.. En. la. caracterización del contexto genómico, se encontraron ortólogos en otros microorganismos cercanos filogenéticamente, esto se obtuvó a través de Microbes Online (Dehal et al., 2009). Caracterizacion mediante la Base de datos (BD) COGs (Clusters of Orthologous Groups o conjuntos de grupos de ortólogos). Cada COGs se define como el grupo de tres o más proteínas, estos han sido desarrollados mediante la comparación de todos contra todos de secuencias de proteínas codificadas en genomas completos secuenciados, usando el programa BLAST. Los COGs se obtuvieron de la BD MicroScope, MaGe (2006). 3.2.2.2 Identificación de rutas metabólicas especificas mediante KEEG entre S. mutans UA159 y S. sanguinis SK36. La segunda estrategia (segunda fase) consistió en hacer un borrador de la reconstrucción metabólica de las rutas más relevantes y el mapa global de las vías para el desarrollo de la competencia en Streptococcus mutans UA159 y Streptococcus sanguinis SK36, basado en la anotación genómica de ambas especies. Para esto, se necesitó la versión más reciente disponible de la anotación de ambos organismos en GenBank, ya que la calidad y fiabilidad de la anotación genómica es crucial para la calidad de la reconstrucción metabólica in silico.. 15.
(25) A partir de la anotación se identificaron las reacciones metabólicas catalizadas por los productos de genes de S. mutans y S. sanguinis. Estas se conectaron usando los números de la Enzyme Commission (EC) y bases de datos de reacciones bioquímicas como las bases de datos: KEGG (Kanehisa et al., 2006), MetaCyc (Karp et al., 2002) y BRENDA (Barthelmes et al., 2007) obteniéndose una lista de metabolismos candidatos. Además se utilizó el servidor MicroScope (2006). Para el refinamiento y validación del modelo metabólico general se usó el servidor web: PubMed (http://www.ncbi.nlm.nih.gov/PubMed) el cual provee de información experimental en S. sanguinis y S. mutans, además de otros microorganismos cercanos filogenéticamente.. 3.2.2.3 Descripción de un modelo de vía se señalización celular entre S. mutans UA159 y S. sanguinis SK36. En este paso se utilizó el software Artemis v.13.2.0, Microbes Online, BlastP y Pubmed. Se procedió analizar en el genoma mediante Artemis todas las posibles proteínas anotadas que participen en procesos de señalización con ayuda de Pubmed, estas fueron contrastadas mediante BlastP (Altschul et al., 1997), para obtener los mejores alineamientos con altas puntuación y se escogió el alineamiento con el mejor E-value. Además se verificó el contexto genético mediante Microbes online. Las secuencias fueron analizadas en Microscope (https://genoscope.cns.fr/agc/microscope/home/index). (Tercera fase). 16.
(26) 3.2.2.4 Realización de comparación de proteínas importantes en la competencia y coexistencia. Se encontró proteínas de interés en competencia con posible función conocida que presentaron altos porcentajes de identidad y score. Estas fueron de interés en la investigación y se caracterizaron usando herramientas disponibles en Base de datos en la Web: (cuarta fase). a.- Relaciones de similitud de la estructura primaria Matcher v.6.3.1 (www.ebi.ac.uk/Tools/psa/), predicciones de estructura secundaria PSI-PRED (Jones, 1999), predicción de proteínas transmembrana TMHMM v.2.0 (Krogh et al., 2001), predicción de motivos Prosite (Hofmann et al., 1999), predicción de dominios Pfam (Bateman et al., 2002), MEME (Bailey & Elkan 1994).y sitios de localización de proteínas PSORT (Nakai et al., 1999) predicción de la localización subcelular CELLO v. 2.5 (Yu et al., 2006) péptido señal SignalP 4.0 (Petersen et al., 2011), además TblasN y BlastP (Altschul et al., 1997). b.- Se encontraron posibles proteínas ancestrales. Para ello se utilizó análisis filogenético: Las secuencias de aminoácidos de las proteínas se alinearon con MSA usando el programa Clustal X v. 2.0 (Thompson et al., 1997), cuya información permitió el diseño del árbol filogenético. Para este propósito se usó el método de máxima parsimonia aplicando el modelo de matrix JTT (Jones et al., 1992), considerando 1000 bootstrap como rèplicas y omisión de gaps. Este análisis se realizó usando el programa MEGA v. 5.05. (Tamura et al., 2011). El análisis filogenético se realizó específicamente de la enzima glutatión peroxidasa de distintas bacterias. Los códigos de GenBank de las glutatión peroxidasa descargadas de NCBI son: A001 B002 C003 D004 E005. E. coli str. K-12 substr. MG1655 Streptococcus sanguinis SK36 S. pneumoniae TIGR4 Desulfotalea psychrophila LSv54 Bdellovibrio bacteriovorus HD100. (NP_416225.1) (YP_001035467.1) (NP_344850.1) (YP_064268.1) (NP_967328.1) 17.
(27) F006 G007 H008 I009 J010. Bacillus subtilis subsp. subtilis str. 168 Shewanella oneidensis MR-1 Streptococcus mitis B6 Streptococcus gordonii str. Challis substr. CH1 Saccharomyces cerevisiae rm11-1a1.. (NP_390073.1) (NP_717176.1) (YP_003446873.1) (YP_001450057.1) (NP_012303.1). En la parte de los anexos se encuentra una figura que muestra las fases del trabajo realizado (Flujograma: Anexo 04, Figura 17).. 3.2.3. Análisis de los datos. Después de la búsqueda de secuencia de proteínas mediante BlastP, y secuencia de nucleótidos Blastn en la base de datos no redundante (NCBI), cada alineamiento fue analizado para medir su significancia estadística. De los alineamientos resultantes llamados pares de alta puntuación (High Score Pairs o HSPs). Se reportaron sólo los pares de alineamientos que obtuvieron una probabilidad menor al parámetro E- value. El e-value fue usado para la obtención de genes o proteínas homólogas (alta identidad) en organismos cercanos evolutivamente. E-value llamado valor esperado de corte, el cual permite definir qué pares alineamientos buenos queremos obtener de acuerdo a su significación estadística. Cuanto menor es el valor del E- value, más significativo es un alineamiento. El análisis fue riguroso y se utilizó un e- value < 10-5 o 0.00005. Además las comparaciones de secuencias mediante alineamiento local fueron medidas con el parámetro porcentaje de identidad (P.I.). Para que ser significantes este valor debe ser >30%. Un valor de E= 0.0 significa 1e-179. En anexos se deja una lista de los principales sitios web utilizados en el presente trabajo de investigación. Los resultados arrojados por las BDs y software fueron incorporados a una base de datos en Microsoft Office Excel 2007 y el diseño de figuras en Microsoft Power Point 2007, para su correspondiente descripción. Se trabajó con un equipo procesador Core I5.. 18.
(28) VI. RESULTADOS Los resultados del presente estudio fueron divididos en 4 secciones para el mejor detalle: 4.1 Comparación genómica de las categorías funcionales COGs, análisis comparativo 4.2 Identificación de rutas metabolicas especificas mediante KEEG entre S. mutans UA159 y S. sanguinis SK36. 4.3 Descripción y reconstrucción del sistema de Quórum sénsing implicado en la transformación genética. 4.4 Realización de comparación de proteínas importantes en la competencia y coexistencia.. 4.1 COMPARACIÓN GENÓMICA Y LAS CATEGORIAS FUNCIONALES COGs.. Como primera estrategia se necesitaba conocer cómo es la conservación general de los genes en los genomas de S. mutans UA159 (en adelante S. mutans) y S. sanguinis SK36 (en adelante S. sanguinis). Se analizó mediante MAUVE (Figura 02) las dos secuencias genómicas, se encontró 137 Bloques Colineares Locales (LCBs) con un peso mínimo de 1495pb. Los (LCB) representan puntos de referencia (homología/ regiones conservadas compartidas) en los cromosomas analizados. Los LCBs que se encuentran en la misma ubicación pero en posiciones superior e inferior unidos por líneas verticales y oblicuas representan el complemento inverso de la secuencia de ADN de referencia. Los 137 LCBs con líneas de conexión (sintenia) entre ambos genomas identifican la ubicación de cada LCB ortólogo en los dos genomas. Regiones con zonas blancas o sin unión en el LCB indican la presencia de secuencias (ORFs) específicos de cada especie o con escasa similitud en la secuencias. Además se observó que la cadena cromosómica comparada entre ambas especies muestran reordenamientos de amplias extensiones de grupos de ORFs, excepto en las posiciones de las 100 000 pb que coiciden con el origen de replicación del ADN en ambos genomas se muestran grupos de secuencias con ortólogos conservados.. 19.
(29) Figura 02. Bloques colineares locales (LCBs) entre dos secuencias cromosómicas de S. mutans UA159 y S. sanguinis SK36. Representación de 137 LCBs entre las secuencias cromosómicas de ADN de las dos especies, el análisis fue realizado mediante el software MAUVE 2.3.1 con un peso mínimo de 20 1495pb. Se muestra alto reordemaniento, inserciones entre ambos genomas..
(30) Se analizó las zonas marcadas (flechas) en S. sanguinis y se encontró un grupo de ORFs que codifican para enzimas de biosíntesis de la vitamina B12, transportadores ABC y proteínas hipotéticas. (Dato no mostrado). En el lugar marcado con una flecha en el genoma de S. mutans se encontró ORFs que le dan resistencia a bacitracina, transposasas, permeasas y proteínas hipotéticas. El análisis genómico entre ambas especies, consistió en clasificar los genes anotados que codifican marcos abiertos de lectura (ORFs) de S. mutans y S. sanguinis, a través de las categorías COGs (Clusters of Orthologous Groups). Los COGs están divididos en 22 categorías, se elaboró la tabla 09 (Anexo 01) con los números de ORFs de cada categoría funcional y se realizó un histograma (Figura 03) para comparar las categorías COGs entre S. sanguinis y S. mutans. Para S. sanguinis el 78,03% de los ORFs se clasifican al menos en un COG (1857 ORFs/2380) y en S. mutans un 80.39 % de los ORFs se clasifican al menos en un COG (1652 ORFs/2055) (https://genoscope.cns.fr/agc/microscope/home/index), actualizado a agosto, 2013. Los ORFs de cada categoría de ambos microorganismos se muestran en porcentajes para cada una de las 22 categorías funcionales. Se detallan (Figura 03) cada una de las 22 categorías funcionales de la célula microbiana. De acuerdo con el análisis realizado de las categorías COGs (Clusters of Orthologous Groups). Se analizaron las categorías más relevantes con respecto al número de genes involucrados. S. sanguinis y S. mutans presentan proteínas con relación a la categoría [C], de producción y conversión de energía, por encima del 3.00% de sus ORFs.Con respecto la categoría [G] Metabolismo y transporte de carbohidratos, presentan un porcentaje similar cercano al 9.00% de sus ORFs. Con respecto a la categoría [E] metabolismo y transporte de aminoácidos ambas especies presentan por encima del 14.30% de su ORFs que codifican para proteínas asociadas a esta categoría y respecto a la categoría [P] Metabolismo y transporte de iones inorgánicos presentan ORFs por encima del 9.00%.. 21.
(31) 17,00% 16,00% 15,00% 14,00% 13,00% 12,00% 11,00% 10,00% 9,00% 8,00% 7,00% 6,00% 5,00% 4,00% 3,00% 2,00% 1,00% 0,00%. S. sanguinis SK36 S. mutans UA159. [B] [C] [D] [E] [F] [G] [H] [I] [J] [K] [L] [M] [N] [O] [P] [Q] [R] [S] [T] [U] [V] [W]. PORCENTAJES DE ORFs (Genes). Categorias COGs. CATEGORIAS FUNCIONALES COGs. Figura 03. Las categorías funcionales COGs entre S. mutans UA159 y S. sanguinis SK36. Fueron analizadas 22 categorías con respecto al porcentaje en el número de marcos abiertos de lectura (ORFs) de los genes en cada categoría, que son las siguientes: [B] Estructura y dinámica de la cromatina; [C] Producción y conversión de energía; [D] Control del ciclo celular, división celular; [E] Metabolismo y transporte de aminoácidos; [F] Metabolismo y transporte de nucleótidos; [G] Metabolismo y transporte de Carbohidratos; [H] Metabolismo y transporte de Coenzimas; [I] Metabolismo y transporte de lípidos; [J] Traducción, biogénesis y estructura ribosomal; [K] Transcripción; [L] Replicación, recombinación y reparación; [M] Biogénesis de la envoltura, membrana y pared; [N] Motilidad celular; [O] Modificación postraduccional y chaperonas; [P] Metabolismo y transporte de iones inorgánicos; [Q] Biosíntesis de metabolitos secundarios; [R] Predicción solo de función general; [S] Función desconocida [T] Mecanismos de Transducción de señales; [U]Tráfico intracelular, secreción y transporte vesicular; [V] Mecanismos de defensa; [W] Estructura extracelulares. El análisis fue realizado en el servidor COGs y Microscope (https://genoscope.cns.fr/agc/microscope/home/index).. 22.
(32) 4.2 COMPARACIÓN DE LOS MAPAS DE LAS VÍAS METABOLICAS ENTRE S. mutans UA159 y S. sanguinis SK36, según KEEG y MetaCyc. La segunda estrategia realizó un mapa modelo del metabolismo global en ambos microorganismos en base a sus posibles proteínas. Se analizaron 183 vías que están presentes en el Dominio Bacteria, como las vías metabólicas y vías de señalización a partir de los mapas de referencia en KEGG y MetaCyc. Además con el soporte de revisión bibliográfica en PubMed (http://ncbi.nlm.nih.gov/pubmed), se obtuvieron diferencias entre el metabolismo de S. mutans UA159 y S. sanguinis SK36, para luego realizar el análisis de las vías más relevantes. Las diferencias encontradas a partir del análisis de los mapas referenciales (Figura 04) se describen para S. sanguinis, el cual presenta vías únicas. En lo que corresponde a metabolismo de aminoácidos, se encontró que S. sanguinis presenta genes candidatos para degradación de histidina. Otra vía hallada única en S. sanguinis, fue la biosíntesis de esfingolípidos, junto con la síntesis de glicerolípidos. Se encontraron además las rutas del metabolismo de nucleótidos y metabolismo de porfirinas. En S. mutans, el análisis (Figura 04), no encontró rutas específicas ya sean vías completas o incompletas con sus grupos de genes. Sin embargo se encontraron enzimas exclusivas de esta especie en color rojo, posiblemente confiriéndole alguna ventaja frente a su competidor S. sanguinis. Se describen los genes que codifican las enzimas exclusivas de S. mutans son: Ipk, 4-difosfocitidil-2-C-metil-D-eritritol quinasa (EC 2.7.1.148), dxs, 1deoxi-D-xilulosa-5-fosfato sintasa (EC 2.2.1.7) presente en la biosíntesis de terpenoides; SacB, beta-D-fructosiltransferasa (EC 2.4.1.10), que forma parte del metabolismo de la sacarosa y el almidón; pycB, oxaloacetato descarboxilasa, subunidad alfa (EC 4.1.1.3); Smu264, agmatina deiminasa (EC 3.5.3.12), que pertenecen al metabolismo de prolina y arginina;. Smu2127,. succinato. semialdehído. deshidrogenasa,. perteneciente. al. metabolismo de glutamato, aspartato y alanina; pckA, fosfoenolpiruvato carboxiquinasa (ATP) (EC 4.1.1.49), presente en la glucólisis/gluconeogénesis, ciclo de Krebs, metabolismo del piruvato; Smu139, oxalato decarboxilasa (EC 4.1.1.2) presente en el metabolismo de glioxilato. Glutamato sintasa smu.365, smu.366 degradación de la glutamina. 23.
(33) Figura 04. Predicción del mapa del metabolismo global que comparten S. sanguinis SK36 y S. mutans UA159. Las líneas de color azul son las vías metabólicas que comparten ambos microorganismos, las líneas verde son exclusivas de S. sanguinis y las líneas rojas fueron exclusivas de S. mutans en todo el esquema están representadas las enzimas y productos intermedios de las reacciones, el trabajo se realizó mediante el servidor KEGG (http://genome.jp/kegg/).. 24.
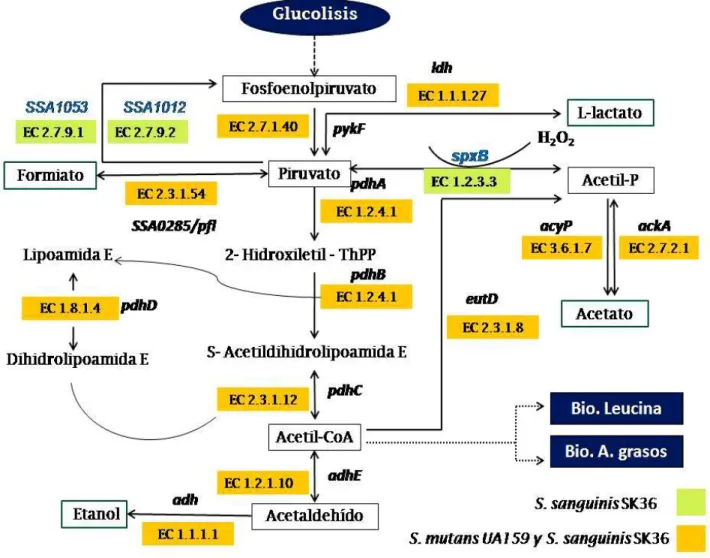
(34) 4.2.1 Reconstrucción y comparación de la vía del metabolismo del piruvato. La via del piruvato es una via central para la fermentación de azucares en los estreptococos. La vía fue reconstruida en base a sus genomas, mediante Pubmed, KEGG y MetaCyc se realizó la reconstrucción del metabolismo del piruvato para ambas especies (Figura 05). El metabolismo del piruvato es conservado en S. sanguinis y S. mutans, estos presentan ortólogos con altos porcentajes de identidad, representado una de sus principales vías para la degradación y formación de energía metabólica (ATP). Sin embargo se presentan enzimas adicionales en S. sanguinis que le podrían conferir una ventaja en la competencia con respecto a S. mutans en la biopelicula dental. En ambas especies se presentan combinaciones similares de genes que codifican enzimas involucradas en esta vía. Hay diferencias en tres enzimas únicas en S. sanguinis SK36, como piruvato fosfato diquinasa (EC 2.7.9.1; SSA1053), fosfoenolpiruvato sintasa (EC 2.7.9.2; SSA1012) y la piruvato oxidasa (EC 1.2.3.3; SSA0391), las cuales no están presentes en S. mutans. El piruvato proveniente de la vía glucolítica puede ser degradado, por medio de la fermentación acidoláctica como se ha descrito para Streptococcus spp. (Madigan et al., 2003). La fermentación acidoláctica, se divide en a) fermentación homoláctica a lactato y b) fermentación heteroláctica produciendo etanol, formiato, lactato y acetato. Estos procesos ocurren a través de una serie de reacciones en ambas especies cuyo punto de partida es la transferencia del grupo fosfato del fosfoenolpiruvato al ADP, formándose ATP y piruvato a través de la enzima piruvato quinasa (EC 2.7.1.40). Se encontró diferencias con respecto a algunos productos génicos en esta vía del metabolismo de piruvato. Radican en que sólo en S. sanguinis el piruvato puede transformase nuevamente en fosfoenolpiruvato a través de dos enzimas piruvato fosfato diquinasa (EC 2.7.9.1; SSA1053) y fosfoenolpiruvato sintasa (EC 2.7.9.2; SSA1012). Además S. sanguinis posee otra reacción alternativa catalizada por la piruvato oxidasa (EC 1.2.3.3, SSA0391) que acelera la transformación de piruvato en acetilfosfato, liberándose H2O2 (Figura 05). 25.
(35) Figura 05. Reconstrucción del mapa del metabolismo del piruvato entre S. mutans UA159 y S. sanguinis SK36, mediante análisis de sus genomas. Se muestra los productos intermediarios y enzimas codificados por los genes presentes en el genoma de S. sanguinis (verde) y S. mutans (naranja).La reacción catalizada por la piruvato oxidasa (EC 1.2.3.3, (SSA0391)) la cual produce H2O2, le proporcionaría una ventaja competitiva a S. sanguinis contra S. mutans, en la biopelícula dental en condiciones aeróbicas (O2). Ldh (Lactato deshidrogenasa), pyKF (Piruvato quinasa), spxB (Piruvato oxidasa), pdhA (piruvato dehidrogenasa E1 subunidad alfa), pdhC (piruvato dehidrogenasa E2), pdhD (Dihidrolipoamida deshidrogenasa), adhE (acetaldehído deshidrogenasa), adh (alcohol deshidrogenasa), acyP (acilfosfatasa), ackA (acetato quinasa), pfl (piruvato formato liasa), eutD (fosfato acetil transferasa). Para este análisis se usaron servidores KEGG, BRENDA y MetaCyc.. 26.
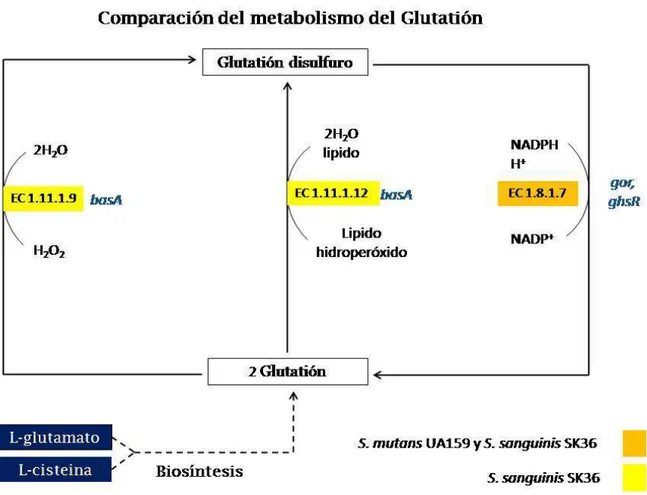
(36) 4.2.2 Predicción de la vía de utilización del glutatión en ambas especies. Se elaboró un mapa comparativo de la vía de utilización del glutatión (Figura 06), para S. mutans y S. sanguinis. La biosíntesis del glutatión proviene del metabolismo del glutamato y cisteína. Ambos organismos comparten una enzima anotada como glutatión reductasa (EC 1.8.1.7) codificada por el gen gor en S. sanguinis y ghsR en S. mutans. En S. sanguinis también se encontró una posible enzima glutatión peroxidasa (EC 1.11.1.9; EC 1.11.1.12; COG0386), codificada por el gen basA (SSA1523). Esta enzima se encuentra en la posición 1 532 207 – 1 532 680 del genoma de S. sanguinis, el producto del gen basA consta de 157 residuos de aa.. Figura 06. Comparación de la vía de utilización del glutatión entre S sanguinis y S. mutans. Se muestra la presencia del gen basA (SSA1523) que codifica una posible glutatión peroxidasa (EC 1.11.1.9; EC 1.11.1.12) en S. sanguinis y la ausencia de esta enzima en S. mutans, sin embargo se presenta en ambas especies una posible enzima glutatión reductasa (EC 1.8.1.7). El análisis se realizó mediante los servidores MetaCyc y KEGG. 27.
(37) 4.2.2.1 Análisis de la posible enzima anotada como glutatión peroxidasa: Se pretendió dilucidar si la secuencia de animoácidos era conservada en los sitios catalíticos de la enzima. Por lo tanto se realizó alineamiento múltiple de secuencia (MSA) con microorganismos cercanos filogenéticamente que presentan la enzima glutatión peroxidasa y se haya caracterizado. Para realizar el análisis se extrajó 10 secuencias de glutatión peroxidasas de especies diferentes depositadas en GenBank, (ver Métodos, tercera fase. b). Se encontró (Figura 7A) aminoácidos altamente conservados (cisteínas y glicinas) en posiciones del sitio activo de las enzimas del género Streptococcus. Además presenta el dominio GSHPx (PF00255) característico de glutatión peroxidasa. Para analizar la conservación de la enzima glutatión peroxidasa en organismos procariontes, se realizó un alineamiento local de las secuencias de E. coli str. K-12 substr. MG1655 y S. sanguinis SK36, arrojando identidad de 41% y similitud de 57.1%. E. coli fue tomado debido que esta caracterizada su enzima y nos serviría como sustento experimental (Figura 7B). El mismo procedimiento se ejecutó entre las secuencias de glutatión peroxidasa (GPXs) del eucariota Saccharomyces cerevisiae versus S. sanguinis, arrojando identidad de 50% y similitud de 66.5%. Ambos resultados muestran altos grados de conservación de la secuencia de esta enzima en organismos procariontes y eucariontes arrojando que los motivos estructurales son importantes para su función biológica y enzimática.. 28.
(38) Figura 07. Alineamiento múltiple y alineamiento local de glutatión peroxidasa EC 1.11.1.9. (A) Alineamiento múltiple de glutatión peroxidasa de diferentes especies filogenéticamente cercanas, las especies de este análisis están codificadas (Ver Metodología), realizado mediante el software ClustalX v. 2. (B) Alineamiento local, de dos glutatión peroxidasas de E. coli str. K-12 substr. MG1655 código A001 y S. sanguinis SK36 B002.. Se usó Matcher v. 6.3.1. (http:/ ebi.ac.uk/Tools/psa/emboss_matcher/).. 29.
(39) 4.2.2.2 Análisis filogenético de las secuencias de glutatión peroxidasa: Se realizó en análisis de cómo estaban relacionadas evolutivamente las secuencias de aminoácidos de enzimas glutatión peroxidasa de microorganismos cercanos filogenéticamente con respecto a S. sanguinis (B002), se usó como grupo externo el eucariota S. cerevisae var.rm11-1a 1. El resultado obtenido fue un árbol filogenético (Figura 08), basado en el método de máxima verosimilitud, usando en el modelo matrix JTT. Se utilizaron 1000 bootstrap como réplicas, agrupando las secuencias en 6 grupos.. Los resultados (Figura 08) indican que las enzimas glutatión peroxidasa de S. sanguinis y S. gordonii (Challis) se ubican en una rama en la cual posiblemente comparten un ancestro común y además que divergieron hace poco tiempo. Se podría predecir que esta enzima es homóloga y posiblemente realicen la misma función con respecto a S. gordoni. En una rama aparecen las cuatro especies del género Streptococcus donde las ramas indican relación de la enzima con respecto a su carácter evolutivo dentro del grupo de Estreptococos mostrándose como un bloque aparte y formando un linaje que posiblemente divergió de un ancestro común con Bacillus subtilis subsp. Subtilis. Algo interesante es que Shewanella oneidensis se ubica en una rama independiente, significa que la enzima de este organismo presenta alto grado de divergencia y posiblemente apareció antes que los demás organismos analizados durante el proceso evolutivo.. 30.
(40) Figura 08. Análisis filogenético de la enzima glutatión peroxidasa de S. sanguinis con respecto a otros organismos que presentan la enzima. Se observó a S. sanguinis, dentro del grupo de los Streptococcus, alejado del grupo externo el eucariota S. cerevisae -rm11-1a 1. Usamos 10 taxones para el análisis. El árbol fue generado usando MEGA versión 5.05 (Tamura et al., 2011). La escala representa el número de sustituciones de aminoácidos por sitio en la cadena peptidica.. 31.
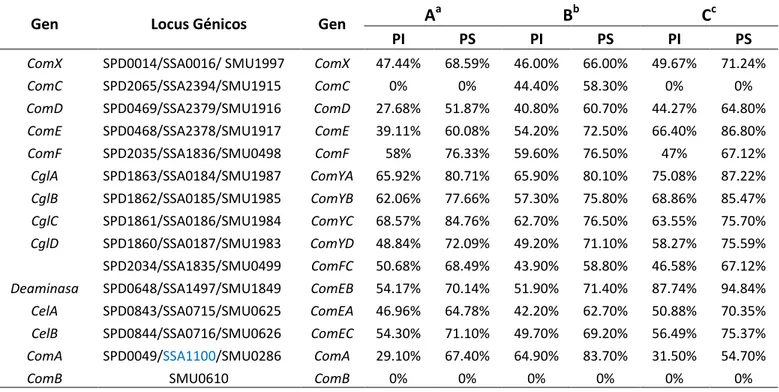
(41) 4.3 DESCRIPCIÓN Y RECONSTRUCCIÓN DEL SISTEMA DE QUÓRUM SENSING: Implicado en la transformación genética en S. sanguinis y S. mutans. 4.3.1 Análisis y reconstrucción de las proteínas de competencia Com componentes del Quórum sensing. Se análizó las proteínas Com involucradas en el sistema de (QS) entre S. sanguinis y S. mutans mediante alineamiento local con Matcher v. 6.3.1. Se usó los % de identidad y % de similitud para obtener datos. Los genes anotados fueron contrastados con sus ortólogos con la especie mas cercana filogenéticamente, Streptococcus pneumoniae cuyos productos génicos han sido corroborados experimentalmente. Los resultados (Tabla 01 y Figura 09) indican que las proteínas codificadas por el operón comCDE, muestran mediano grado de conservación. La secuencia aa de la proteína comC es completamente diferente entre ambos microorganismos. ComX, presenta notable grado de conservación, esta proteína tiene como función unirse al ARN polimerasa. Los genes del operón comAB, codifican proteínas con una alta conservación entre S. mutans, sin embargo S. sanguinis no presenta estos transportadores tipo ABC, esta función podría ser llevada a cabo por otro tipo de transportadores del tipo ABC. No se reconoció ninguna proteína anotada ComA y ComB en S. sanguinis que presente un ortólogo en S. pneumoniae. En S. mutans presenta la proteína ComB pero no presenta homólogos. Respecto a los genes del operón comYABCD, codificantes de proteínas del locus Y en ambas especies, presentan un alto grado de conservación en las secuencias aminoacídicas. Además el operón comEABC, codifica proteínas con alta identidad, posiblemente presente dominios estructurales y funcionales conservados que son importantes para el sistema de transformación genética en ambos microorganismos.. 32.
(42) Tabla 01. Comparación de las secuencias de proteínas de la competencia Com involucradas en la formación del sistema de Quórum sensing de S. sanguinis y S. mutans, caracterizados molecularmente en S. pneumoniae D39. Gen. Locus Génicos. Gen. Aa. Bb. Cc. PI. PS. PI. PS. PI. PS. ComX. SPD0014/SSA0016/ SMU1997. ComX. 47.44%. 68.59%. 46.00%. 66.00%. 49.67%. 71.24%. ComC. SPD2065/SSA2394/SMU1915. ComC. 0%. 0%. 44.40%. 58.30%. 0%. 0%. ComD. SPD0469/SSA2379/SMU1916. ComD. 27.68%. 51.87%. 40.80%. 60.70%. 44.27%. 64.80%. ComE. SPD0468/SSA2378/SMU1917. ComE. 39.11%. 60.08%. 54.20%. 72.50%. 66.40%. 86.80%. ComF. SPD2035/SSA1836/SMU0498. ComF. 58%. 76.33%. 59.60%. 76.50%. 47%. 67.12%. CglA. SPD1863/SSA0184/SMU1987. ComYA. 65.92%. 80.71%. 65.90%. 80.10%. 75.08%. 87.22%. CglB. SPD1862/SSA0185/SMU1985. ComYB. 62.06%. 77.66%. 57.30%. 75.80%. 68.86%. 85.47%. CglC. SPD1861/SSA0186/SMU1984. ComYC. 68.57%. 84.76%. 62.70%. 76.50%. 63.55%. 75.70%. CglD. SPD1860/SSA0187/SMU1983. ComYD. 48.84%. 72.09%. 49.20%. 71.10%. 58.27%. 75.59%. SPD2034/SSA1835/SMU0499. ComFC. 50.68%. 68.49%. 43.90%. 58.80%. 46.58%. 67.12%. Deaminasa. SPD0648/SSA1497/SMU1849. ComEB. 54.17%. 70.14%. 51.90%. 71.40%. 87.74%. 94.84%. CelA. SPD0843/SSA0715/SMU0625. ComEA. 46.96%. 64.78%. 42.20%. 62.70%. 50.88%. 70.35%. CelB. SPD0844/SSA0716/SMU0626. ComEC. 54.30%. 71.10%. 49.70%. 69.20%. 56.49%. 75.37%. ComA. SPD0049/SSA1100/SMU0286. ComA. 29.10%. 67.40%. 64.90%. 83.70%. 31.50%. 54.70%. ComB. 0%. 0%. 0%. 0%. 0%. 0%. ComB. SMU0610. a. A : Alineamiento de S. sanguinis /S. mutans b B : Alineamiento de S. pneumoniae/S. mutans c C :Alineamiento de S.pneumoniae/S. sanguinis Alineamiento local realizado en Macther, PI: porcentaje de Identidad ; PS: porcentaje de Similitud.. comparación de proteinas Com 90,00% 80,00% 70,00% 60,00% 50,00% 40,00% 30,00% 20,00% 10,00% 0,00%. % Identidad % Similitud. Figura 09. Histograma del análisis comparativo de las secuencias de proteínas Com involucradas en el Sistema de Quórum sensing entre S. sanguinis y S. mutans. BlastP (http://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins) mediante Matcher v.6.3.1. 33.
(43) 4.3.2. Descripción del modelo del sistema de Quórum sensing (QS) en S. sanguinis y S. mutans en la biopelícula dental. Realizado el análisis de las posibles proteínas involucradas en la competencia se procedió a describir el sistema de QS (Figura 10), en ambos microorganismos basados en el modelo de la bacteria S. pneumoniae de las proteínas validadas experimentalmente en esta especie. El desarrollo de la competencia genética natural es mediado a través del sistema de Quórum sensing (Cvitkovitch et al., 2003). La vía regulatoria se inicia con señales causadas por péptidos estimulantes de la competencia (CSP) en el medio extracelular, este deriva desde un pre-péptido (CSP) codificado por el gen comC (SSA2394/SMU1915). Luego es exportado al exterior de la célula a través de un transportador tipo ABC con unión a ATP, codificado por los genes comA (SSA1100/SMU0286) y comB (SMU0287). El CSP en el medio extracelular es detectado a través del Sistema regulador de dos componentes (TCS), compuesto por una proteína histidina quinasa que se autofosforila en un residuo de histidina, la cual es codificada por el gen comD (SSA2379/SMU1916). La proteína reguladora de respuesta, la cual recibe un grupo fosfato en un residuo de aspartato es codificada por el gen comE (SSA2378/SMU1917). La proteína ComE regula la expresión del gen comX (SSA0016/SMU1997), cuyo producto es un factor sigma el cual se une a la ARN polimerasa. La ARN polimerasa junto al factor sigma ComX se adhiere a la caja com, este complejo reconoce específicamente la secuencia TACGAATA, caja presente en genes tardíos, los cuales son requeridos para sintetizar proteínas de competencia para la internalización del ADN extracelular que se encuentra en el biofilm (biopelícula) dental.. 34.
(44) En ambos genomas se encontró el locus Y donde se encuentra la disposición un grupo de genes comYA (SSA0184/SMU1987), comYB (SSA0185/SMU1985), comYC (SSA0186/SMU1984), comYD (SSA0187/SMU1983). Estos genes codifican proteínas que presentan regiones con hélices transmembrana posiblemente se ubiquen en la pared y membrana celular. Sus ortologós a estos genes en S. pneumoniae serian CglABCD. Los genes comEA (SSA0715/SMU0625) y comEC (SSA0716/SMU0626), también codifican proteínas que muestran regiones con hélices transmembranales y estarían ubicadas en la membrana citoplasmática. El gen comEB (SSA1497/SMU1849) codifica una posible proteína deaminasa. Los ortólogos de los genes comEAC en S. pneumoniae son los genes CelAB, ComF. Se predice que éstos forman un complejo proteico en forma de un poro o tubo que atraviesa la pared y la membrana celular para la captación e internalización del ADN. En el biofilm se da la competencia entre especies interespecificas que internalizan el ADN que se encuentran en el medio debido posiblemente a procesos de lisis celular causados por H2O2 o de mutacinas.. 35.
(45) Figura 10. Modelo propuesto para la transformación genética mediada por el sistema de Quórum sensing en S. sanguinis SK36 y S. mutans UA159. Se usó como soporte y modelo secuencias de proteínas caracterizadas y validadas molecularmente en S. pneumoniae involucradas en la competencia. Los detalles son descritos en el texto 4.3.2. Análisis realizado mediante BlastP y MicroScope (https://genoscope.cns.fr/agc/microscope/home/index).. 36.
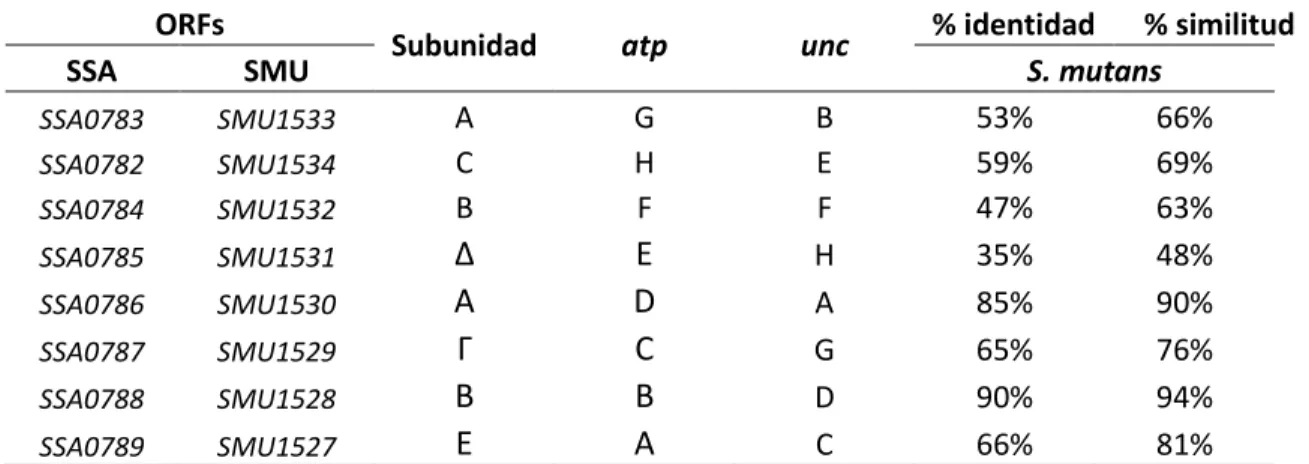
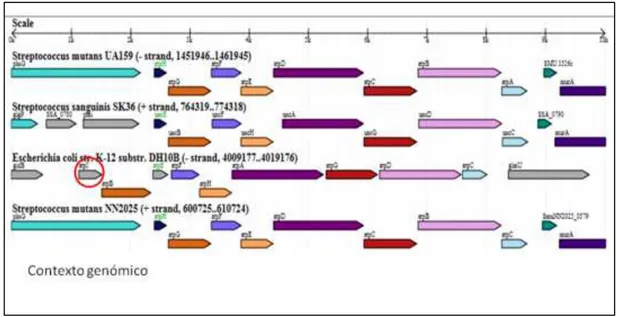
(46) 4.4 COMPARACIÓN DE PROTEINAS IMPORTANTES EN LA COMPETENCIA Y COEXISTENCIA ENTRE S. sanguinis y S. mutans. 4.4.1 Análisis de los complejos enzimáticos ATPasas (tipo F – V) en S. sanguinis SK36 y S. mutans UA159. La fosforilación oxidativa de ambas especies presenta el sistema de ATPasas tipo F, que es común en bacterias. Los operones unc y atp fueron caracterizados (Tabla 03) mediante MicrobesOnline (http://microbesonline.org/) la disposición de los genes en el genoma de S. sanguinis y de S. mutans y su relación con organismos cercanos filogenéticamente que presentan este sistema de ATPasa-F. Se encontró en S. sanguinis que este sistema es codificado por el operón unc el cual contiene 8 genes SSA0782 - 0789. En S. mutans se presenta el operón atp que contiene también 8 genes SMU 1527- 1534. Todos los genes presentan dominios conservados del sistema de ATPasas-F. También se comparó la identidad de los 8 ORFs contenido en los respectivos operones (Tabla 02) y su organización (Figura 11) del contexto genómico relacionados con E. coli. E.coli presenta 9 genes para codificar el sistema de ATPasas-F entre ellos el gen atpI, que no se encontró en el genoma de S. mutans ni de S. sanguinis. Además se encontró el operón ntp, que presenta 8 genes anotados como ntpA, ntpB, ntpC, ntpD, ntpE, ntpG, ntpI, ntpK (SSA0085-0093) presentes en el genoma de S. sanguinis que podrían codificar posibles proteínas estructurales de la enzima ATPasa vacuolar, sin embargo estos posibles genes están ausentes en S. mutans (Tabla 03). Se detallan las características de la enzima ATPasa-V de S. sanguinis, con sus códigos EC, números de aminoácidos por proteínas y códigos Pfam encontrados.. 37.
Figure
+7
Documento similar
Sin embargo, con el consumo crónico o excesivo de etanol se favorece el metabolismo del retinol, ya que el etanol induce enzimas que degradan ambas sustancias, como el citocromo
(82) En ese sentido, incluso alguien tan poco "sospechoso" de querer favorecer la investigación y experimentación de preembriones como Albin ESER ha prevenido del uso a
El nuevo Decreto reforzaba el poder militar al asumir el Comandante General del Reino Tserclaes de Tilly todos los poderes –militar, político, económico y gubernativo–; ampliaba
De acuerdo con Harold Bloom en The Anxiety of Influence (1973), el Libro de buen amor reescribe (y modifica) el Pamphihis, pero el Pamphilus era también una reescritura y
Abstract: This paper reviews the dialogue and controversies between the paratexts of a corpus of collections of short novels –and romances– publi- shed from 1624 to 1637:
De hecho, este sometimiento periódico al voto, esta decisión periódica de los electores sobre la gestión ha sido uno de los componentes teóricos más interesantes de la
Ciaurriz quien, durante su primer arlo de estancia en Loyola 40 , catalogó sus fondos siguiendo la división previa a la que nos hemos referido; y si esta labor fue de
Para denegación hegeliana del mal: «Así como no existe lo fal- so, no existe el mal, es objetada primero por Sade y luego por la subjetividad romántica: en la mé- dula de la
