Differential expression of pro inflammatory cytokines, endothelin 1 and nitric oxide synthases in the rat carotid body exposed to intermittent hypoxia
12
0
0
Texto completo
(2) BR A IN RE S E A RCH 1 3 95 ( 20 1 1 ) 7 4 –8 5. body (CB) to hypoxia contributes to the OSA-induced hypertension (Iturriaga et al., 2009; Prabhakar et al., 2010; Smith and Pacchia, 2007; Weiss et al., 2007). Patients with recently diagnosed OSA show enhanced sympathetic and cardiorespiratory responses to hypoxia, attributed to an enhanced CB chemoreflex sensitivity to hypoxia (Lusina et al., 2006; Narkiewicz et al., 1998; Narkiewicz et al., 1999). Moreover, studies performed in animals, confirmed that CIH selectively enhances CB chemosensory and ventilatory responses to acute hypoxia (Del Rio et al., 2010; Pawar et al., 2009; Peng et al., 2003; Reeves et al., 2003; Rey et al., 2004), producing autonomic alterations (Dick et al., 2007; Marcus et al., 2010; Rey et al., 2004, 2008; Zoccal et al., 2008) and long-term potentiation of motor ventilatory activity (McGuire et al., 2003; Mitchell et al., 2001). It is widely accepted that the primary oxygen sensors in the CB are the glomus cells, which make synaptic contact with nerve terminals of chemosensory petrosal neurons (Iturriaga and Alcayaga, 2004; Iturriaga et al., 2007). The current model of CB chemotransduction states that hypoxia induces the depolarization of glomus cells, the entry of Ca2+ and the subsequent release of excitatory transmitters, which in turn increases the discharges of the chemosensory neurons (Iturriaga and Alcayaga, 2004; Iturriaga et al., 2007). Among several molecules present in glomus cells, acetylcholine and adenosine triphosphate fulfill most of the criteria to be considered as excitatory transmitters between glomus cells and petrosal nerve endings (Iturriaga et al., 2007). However, other molecules such as serotonin, GABA, nitric oxide (NO) and endothelin-1 (ET-1) modulate the chemosensory process acting on CB blood vessels or glomus cells (Iturriaga et al., 2007; Rey and Iturriaga, 2004). More recently, it have been found that pro-inflammatory cytokines such as interleukin 1β (IL-1β), interleukin 6 (IL-6), and the tumor necrosis factor-α (TNF-α), are excitatory modulators of CB chemoreception in rats (Lam et al., 2008; Liu et al., 2009; Shu et al., 2007). The mechanisms underlying the enhanced CB chemosensory reactivity to hypoxia induced by CIH are not entirely known. Nevertheless, the evidence indicates that cyclic episodes of hypoxia–reoxygenation produce oxidative stress due to the accumulation of reactive oxygen species (ROS), which are involved in the chemosensory facilitation induced by CIH. Indeed, Peng et al. (2003) found that pretreatment of rats for 10 days before the exposure to CIH with a superoxide dismutase mimetic prevents the CB chemosensory potentiation. Recently, we tested the hypothesis that oxidative stress contributes to the CB chemosensory potentiation and the progression of the hypertension in rats exposed to CIH for 21 days. We found that ascorbic acid, which impedes the systemic and local oxidative stress in the CB, prevents the enhanced CB chemosensory responses to hypoxia and the hypertension induced by CIH (Del Rio et al., 2010). Although, these observations support a main contribution for ROS in the generation of the CB chemosensory potentiation and the cardiovascular pathological consequences of CIH, a direct role for ROS in the chemotransduction process is a matter of debate (Park et al., 2009). Peng et al. (2009) found that exogenous H2O2 mimicked the long-term facilitation of CB. 75. chemosensory discharges in normoxic control rats and mice, similarly to what is found in CIH-treated animals, but the administration of H2O2 does not produce per se CB chemosensory excitation in the rat (Peng et al., 2009) or cat CB (Osanai et al., 1997). Furthermore, modifications of the ROS production in glomus cells does not modify its neurosecretory activity, suggesting a lack of a causal link between ROS levels and glomus cells activity (Gomez-Niño et al., 2009; Gonzalez et al., 2007). Another plausible explanation is that the enhancing effects of ROS on oxygen CB chemoreception are mediated by other molecules, produced downstream of the ROS signal, which act upon the CB blood vessels and/or glomus cells. It is well known that an enhanced ROS production induced by hypoxia–reoxygention evokes the activation of transcription factors such as the nuclear factor kappa beta (NF-κβ), and the hypoxic inducible factor 1α (HIF-1α), increasing the expression of pro-inflammatory molecules such as IL-1β, IL-6, TNF-α and iNOS (Janssen-Heininger et al., 2000; Semenza and Prabhakar, 2007). Previously, we found that CIH increases the expression and the functional vasoconstrictor tone of ET-1 in the CB of cats exposes to short-term CIH (Rey et al., 2006). In addition, new evidence showing excitatory effects of cytokines on CB chemoreception (Fan et al., 2009; Lam et al., 2008; Liu et al., 2009; Shu et al., 2007), supports a possible role for pro-inflammatory cytokines as mediators of the CB chemosensory potentiation induced by CIH. However, there is no information regarding changes of the expression of TNFα, IL-1β and IL-6 in the CB of animals exposed to CIH. Based on the discussed evidence, we hypothesized that CIH may increase the expression of pro-inflammatory cytokines and other chemosensory modulators, such as ET1 and NO, which may potentially contribute to enhance the carotid chemosensory responses to hypoxia. Recently, we reported that CIH potentiates the CB chemosensory responses to hypoxia, and increases the expression of 3nitrotyrosine (3-NT) and iNOS in the CB of rats exposed to intermittent hypoxia for 21 days (Del Rio et al., 2010). However, we did not study if the expression of these molecules was temporarily related to the progression of the enhanced CB chemosensory responses to hypoxia. Thus, the aim of the present study was to determine if the CB chemosensory potentiation to hypoxia shows a temporal association with changes in the expression of TNF-α, IL-1β, IL-6, 3-NT, ET-1, iNOS, and eNOS in the CB from rats following CIH for 7, 14 and 21 days. We performed such study in a suitable model of rats that developed hypertension following 21 days of CIH exposure (Del Rio et al., 2010; Iturriaga et al., 2009).. 2.. Results. 2.1. Effect of chronic intermittent hypoxia on TNF-α, IL-1β and IL-6 immunoreactivity and immune cell invasion in the rat CB The CBs from Sham rats exhibited a weak positive immunoreactive staining for TNF-α, IL-1β and IL-6, mainly confined to clusters of glomus cells as shown in Fig. 1A. Nevertheless, it is worth noting that not all glomus cells showed positive. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
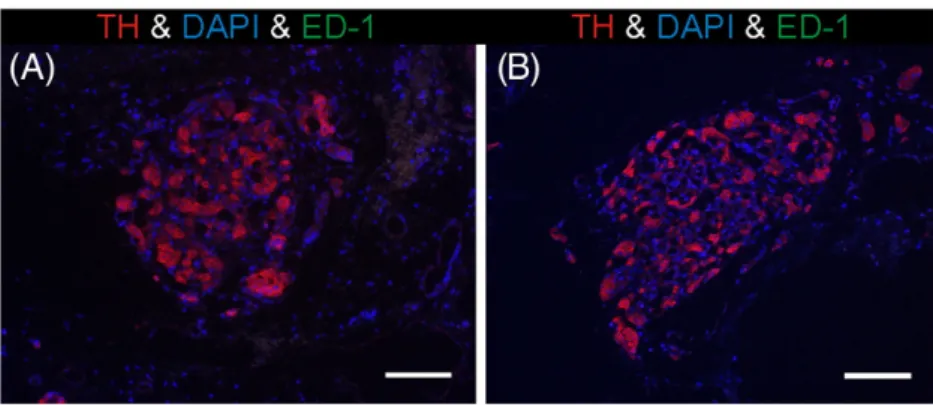
(3) 76. BR A IN RE S EA RCH 1 3 95 ( 20 1 1 ) 7 4 –85. Fig. 1 – Relative expression of TNF-α, IL-1β and IL-6 the CB of rats exposed to 7, 14 and 21 days of CIH. A, Positive immunoreactivity for TNF-α, IL-1β and IL-6 in the CB from a Sham rat and rats exposed to 7, 14 and 21 days of CIH. Sections were counterstained with Harris-Hematoxylin. Inset, negative control by omission of the primary antibody. Scale bar, 20 μm. B, Quantification of changes for TNF-α, IL-1β and IL-6 in the CBs from rats exposed to 7, 14 and 21 days of CIH. ***, P < 0.001; **, P < 0.01; *, P < 0.05 compared to sham condition. Newman–Keuls test after one-way ANOVA. n = 4–6.. immunoreactive staining for TNF-α, IL-1β and IL-6. CIH produced a progressive increase in the relative expression level of TNF-α-ir (Figs. 1A and B), which was significantly different from the Sham group levels at 14 days of CIH exposure. The positive IL-1β-ir showed a similar behavior; a significant increase in IL-1β-ir was detected at 21 days of CIH exposure (Fig. 1B). On the contrary, CIH did not modify the relative expression of IL-6-ir in the CB tissue (Figs. 1A and B).. To determine if the increased expression of TNF-α and IL-1β-ir depends on the local cytokine production in the CB, or if the increased levels were produced by the tissue infiltration of immune cells, we measured the specific biomarker ED-1 to detect activated macrophages in the CB from rats exposed to CIH or sham conditions for 21 days. We did not detect any ED-1 positive cells in the CB parenchyma from CIH-treated (Fig. 2, n = 4) or Sham rats (n = 4). Thus, the increase in cytokines. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
(4) BR A IN RE S E A RCH 1 3 95 ( 20 1 1 ) 7 4 –8 5. 77. Fig. 2 – Effects of CIH on macrophages infiltration in the rat CB. No ED-1 positive immune cells were found in the CB after 21 days of CIH exposure. A, CB from a sham rat. B, CB from a 21 days exposed rat. TH positive cells (red); ED-1 cells (green); nuclei DAPI (blue). Scale bar, 100 μm.. expression induced by CIH was not be explained by the infiltration of immune cells in the CB tissue from rats exposed to 21 days of CIH.. 2.2. Effects of chronic intermittent hypoxia on ET-1, eNOS and iNOS immunoreactivity in the rat CB Fig. 3 shows the effects of CIH on ET-1, eNOS and iNOS immunoreactivity in the CB tissue from rats exposed to 7, 14 and 21 days of CIH. ET-1-ir was found in Sham and CIH-treated CBs. Positive ET-1-ir signal was present in perilobular areas, which contain capillaries and nerve fibers encircling the glomus cell clusters (glomoids), and in glomus cells defined by the ovoid-like morphology and prominent nuclei (Fig. 3A). Exposure to CIH for 7 days induces a 2-fold increase in the ET1-ir positive staining in the rat CB, but at 14 days of CIH, the ET-1-ir staining returned to the level observed in the Sham group and remained close to the sham values throughout the 21 days of CIH exposure (Fig. 3B). Thus, CIH produced a transient increased of ET-1-ir, suggesting that ET-1 may contribute to the enhanced CB chemosensory excitability during the early phase of the intermittent hypoxic challenge. The eNOS-ir positive staining was present in the CB tissue around the chemoreceptor cell clusters and in endothelial cells from capillaries and arterioles. However, as is shown in Fig. 3A, the eNOS-ir was mainly confined to the glomus cells. CIH induced a significant decrease in eNOS-ir by the 7 day of exposure (P < 0.001), which was followed by a progressive but incomplete recovery towards the level observed in Sham rats after 21 days of CIH (Fig. 3B). We found a weak positive staining for iNOS-ir in the glomus cells from Sham rats (Fig. 3A), which was 3-fold lower than the eNOS-ir integrated optical intensity. Contrarily to what happened with eNOS-ir, rats exposed to CIH showed a progressive increase in iNOS-ir positive staining in the CB, being significantly different from the Sham level at 21 days of CIH (Fig. 3B, P < 0.05). Note that exposure to CIH for 21 days induced a significant increased in the positive iNOS-ir that was not only confined to glomus cells, but also other cells showed positive iNOS-ir.. 2.3. Plasma levels of ET-1 and inflammatory cytokines in CIH-treated rats Table 1 shows the effects of CIH on plasma concentrations of ET-1, TNF-α, IL-1β and IL-6. No significant changes in the ET1 plasma concentrations were found in rats exposed to 7, 14 or 21 days of exposure to intermittent hypoxia when compared to Sham ET-1 plasma levels. No detectable levels of TNF-α and IL-1β were found in the Sham or CIH-treated rats. The IL-6 plasma levels presented a transient increase at 7 days of CIH compared to the Sham rats, followed by a progressive return towards Sham levels by 21 days of CIH exposure.. 2.4. Chronic intermittent hypoxia induces oxidative stress in the CB Nitrotyrosine formation was used as an index of CB oxidative/ nitrosative stress as previously described (Del Rio et al., 2010). The CB of Sham rats showed a weak 3-NT-ir positive staining mainly in glomus cells, although some blood vessels showed positive staining (Fig. 4A). CIH strongly increases the 3-NT-ir positive staining in the CB tissue. The statistical analysis reveals that the optical integrated intensity for 3-NT increased at 7 days of CIH and remained elevated at 14 and 21 days of CIH, as compared to the Sham group (Fig. 4B). Thus, the increased formation of 3-NT suggests that the CB tissue is continuously exposed to oxidative stress during the CIH exposure.. 2.5. Effects of chronic intermittent hypoxia on CB chemosensory responses to acute hypoxia Exposure to CIH for 7 days produced potentiation of rat carotid chemosensory responses to acute hypoxia, an effect that persisted until the 21 days of CIH. Fig. 5A illustrates representative recordings of carotid chemosensory responses induced by short hypoxic challenges (PO2 ~5 mm Hg) in rats exposed to CIH for 7, 14 and 21 days. The effects of CIH on the chemosensory responses measured at several PO2 levels (from 5 to 670 mm Hg) are summarized in Fig. 5B. The two-. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
(5) 78. BR A IN RE S EA RCH 1 3 95 ( 20 1 1 ) 7 4 –85. Fig. 3 – Time-course of the effects of CIH on ET-1, eNOS and iNOS in the rat CB. A, Positive immunoreactivity for ET-1, eNOS and iNOS in the CB from a Sham rat and rats exposed to 7, 14 and 21 days of CIH. Sections were counterstained with Harris-Hematoxylin. Inset, negative control by omission of the primary antibody. Scale bar, 20 μm. B, Quantification of the effects of CIH on the expression ET-1, eNOS and iNOS. ***, P < 0.001; *, P < 0.05 compared to sham condition. Newman–Keuls test after one-way ANOVA. n = 6–8.. way ANOVA analysis showed that the overall chemosensory curve for PO2 was different in CIH-rats at the day 7 of CIH exposure (P < 0.001). The Newman–Keuls test showed that carotid chemosensory discharges were higher for the normoxic and hypoxic PO2 values in rats exposed to 7, 14 and 21 days of CIH, as compared with the discharges recorded in the Sham group (Fig. 5B).. 3.. Discussion. 3.1.. General. We studied the changes in the immunohistological expression of pro-inflammatory molecules in the CB and the progression of. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
(6) BR A IN RE S E A RCH 1 3 95 ( 20 1 1 ) 7 4 –8 5. Table 1 – Effects of CIH on plasma concentration of ET-1, TNF-α, IL-1β and IL-6 in rats exposed to 7, 14 and 21 days of intermittent hypoxia. Sham ET-1 (pg/ml) TNF-α (pg/ml) IL-1β (pg/ml) IL-6 (pg/ml). CIH 7 days CIH 14 days CIH 21 days. 1.8 ± 0.2. 2.0 ± 0.3. 1.6 ± 0.2. 1.5 ± 0.3. N.D.. N.D.. N.D.. N.D.. N.D.. N.D.. N.D.. N.D.. 139.10 ± 88.6. 218.6 ± 35.7*. 159.9 ± 11.9. 68.5 ± 8.1*. Data expressed as means ± SE. N.D., non detectable levels were found in the samples; *, P < 0.05 CIH vs. Sham. Newman–Keuls test after one way ANOVA. n = 6–8.. the enhanced carotid chemosensory responses to hypoxia in rats exposed to CIH for 7, 14 and 21 days. Rats exposed to CIH developed a stable potentiation of basal chemosensory discharges and carotid chemosensory responses to acute hypoxia at 7 days of exposure. The main findings of this study showed that the exposure of rats to CIH, which elicits a local CB oxidative/nitrosative stress confirmed by the accumulation of 3-NT, modified the relative expression of pro-inflammatory cytokines, eNOS and iNOS, and ET-1 in the rat CB with different temporal courses. Indeed, CIH produced an early but transient increase of the ET-ir positive staining at 7 days, a decrease in eNOS immunoreactivity and a delayed but progressive increase of the immunorreactive levels of TNF-α, IL-1β and iNOS. The CIH-induced increase of TNF-α and IL-1β immunoreactivity was not associated with increased plasma levels or to the CB tissue invasion of immune cell, suggesting that CIH augment the local production of cytokines in the CB.. 3.2. Augmented expression of pro-inflammatory cytokines induced by CIH The progression of the hypertension in OSA patients and animals exposed to CIH has been associated to increased levels of pro-inflammatory cytokines (Garvey et al., 2009; Jelic and Le Jemtel, 2008; Minoguchi et al., 2005; Williams and Scharf, 2007). It is known that an enhanced production of ROS induced by hypoxia–reoxygenation evokes the expression of several genes, and the synthesis and secretion of proinflammatory cytokines. In response to oxidative stress, HIF1α induces the expression of several genes including ET-1 and iNOS, but ROS also produces the translocation of NF-κβ to the nucleus, potentiating the expression of IL-1β and TNF-α, iNOS and ET-1 (Janssen-Heininger et al., 2000). In the last years, some studies found that the pro-inflammatory cytokines IL-1β, IL-6, and TNF-α, are excitatory modulators of the rat CB chemoreception (Lam et al., 2008; Liu et al., 2009; Shu et al., 2007). Indeed, the rat glomus cells express these cytokines, as well as the IL-1R1 receptor for IL-1β and the TNF-R1 receptor for TNF-α (Lam et al., 2008; Wang et al., 2002, 2006). Shu et al. (2007) reported that the exogenous application of IL-1β to the isolated glomus cells of the rat CB inhibits the O2-dependent voltage-gated K+ currents and the concomitant increase of intracellular Ca2+. Furthermore, Shu et al. (2007). 79. showed that IL-1β produced CB chemosensory excitation in rats, effect blocked by the IL-1β receptor antagonist IL-1ra. In addition, pro-inflammatory cytokines have been involved in the enhanced rat CB chemosensory responses induced by sustained hypoxia. Indeed, Lam et al. (2008) found that exposure of rats to sustained hypobaric hypoxia increases the mRNA expression of IL-1β, TNF-α and IL-6, as well as the expression of the IL-1R1, IL-6R and TNF-R1 receptors. Recently, Liu et al. (2009) reported that chronic hypobaric hypoxia recruits macrophages to the CB, and increases the expression of mRNAs for IL-1β, TNF-α, and the monocyte chemoattractant protein-1 after 1 day of hypoxia. The increased expression of cytokine was observed not only in monocytes, but also in glomus cells and sustentacular cells, the glial-like cells of the CB. To our knowledge, this is the first report showing that CIH increases the expression of TNF-α and IL-1β in the rat CB. Present data shows that cytokine immunoreactivity was mainly confined to CB cells, and the increased positive staining was not associated to the invasion of immune cell within the rat CB. In contrast to what was found in the CB from rats exposed to sustained hypobaric-hypoxia (Liu et al., 2009), we did not detect any evidence of immune cell invasion in the CB tissue after 21 days of CIH exposure. Since a small population of resident ED-1 positive cells has been found in the normal rats CB (Dvorakova et al., 2000), we performed positive controls to confirm the specificity and quality of the antibody used in our study. We detected positive immunostaining for ED-1 cells in the rat spleen (data not shown). Thus, our results suggest that CIH increases the local expression of TNF-α and IL-1β in the rat CB. In addition, we measured the plasma levels of TNF-α, IL-1β and IL-6 in rats exposed to 7, 14 and 21 days of CIH. While plasmatic concentrations of TNF-α and IL-1β remained beyond the detection limits of the rat specific ELISA kits for TNF-α and IL-1β in all the experimental conditions, we found that IL-6 levels increased transiently during the first week of CIH. The plasmatic levels of cytokines measured in the Sham rats agree with previous reports showing similar levels of IL-6 (Ling et al., 2001) and no detectable levels of TNF-α and IL-1β in plasma samples from normoxic rats (Anuar et al., 2006; Ling et al., 2001; Morita et al., 2009). Since plasmatic cytokine detection was performed in samples obtained after 12–16 h from the last hypoxic exposure, we cannot preclude that CIH may have evoked transient elevations of plasma cytokines levels. Indeed, Morita et al. (2009) found that rat TNF-α and IL-1β plasma levels increased one hour after the lung injury induced by a mechanical ventilatory insult, while IL-6 plasma level increased after 6 h. Thus, we cannot rule out acute effects of CIH on plasma TNF-α, IL-1β and IL-6 concentrations.. 3.3. Contribution of vasoactive molecules to the enhanced CB chemosensory response to hypoxia induced by CIH Endothelin-1 has been proposed to contribute to the CB chemosensory potentiation induced by CIH (Pawar et al., 2009; Rey et al., 2006, 2007). This vasoconstrictor peptide is expressed in the endothelium of CB blood vessels and in glomus cells of the rat and cat CB (Chen et al., 2007; McQueen et al., 1995; Rey et al., 2006, 2007) and increases its expression. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
(7) 80. BR A IN RE S EA RCH 1 3 95 ( 20 1 1 ) 7 4 –85. Fig. 4 – Exposure to 7, 14 and 21 days of CIH increased 3-NT-ir in the rat CB. A, Positive 3-NT signal in the CB from a Sham rat and rats exposed to 7, 14 and 21 days. Sections were counterstained with Harris-Hematoxylin. Inset, negative control by omission of the primary antibody. Scale bar, 20 μm. B, Quantification of the effects of CIH on the expression of 3 NT-ir. *, P < 0.05 compared to Sham condition; N.S., P > 0.05, Newman–Keuls test after one-way ANOVA. n = 6–8.. in the cat CB exposed to CIH for 4 days (Rey et al., 2006). The administration of ET-1 produces chemosensory excitation in the in situ and in vitro perfused cat CB, but not in the superfused preparation (Rey and Iturriaga, 2004), indicating that the excitatory effect is associated with its vasomotor effect. The enhanced carotid chemosensory responses to hypoxia were reduced by bosentan in CIH-treated cats, but not in the controls (Rey et al., 2006), indicating that a local increase of ET-1 contributes to the enhanced carotid chemosensory responses. Pawar et al. (2009) tested the hypothesis that ET-1 induced by ROS plays an essential role in CIH-evoked chemosensory potentiation of the neonatal rat CB. In neonatal. rats exposed to CIH for 10 days, which exhibited enhanced carotid chemosensory responses to hypoxia, they did not detect any increase in the expression of ET-1 in the CB, but found a potentiated release of ET-1 during normoxia and upregulation of the ET-A receptor. The systemic administration of MnTMPyP prevented the elevation of ROS level, the increased release of ET-1, the upregulation of ET-A and the enhanced CB chemosensory response (Pawar et al., 2009). Thus, these results support the idea that a ROS-dependent ET-1 release as well as the up-regulation of ET-A receptor are involved in the potentiation of the CB chemosensory response to hypoxia elicited by CIH in neonatal rats. However, it is worth. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
(8) BR A IN RE S E A RCH 1 3 95 ( 20 1 1 ) 7 4 –8 5. 81. Iturriaga, 2004), a reduced NO level may contribute to enhance the CB chemosensitivity, as well as to amplify the vasoconstrictor effect of ET-1. This interpretation is supported by the recent finding of Marcus et al. (2010), who found that CIH decreases the expression of the neuronal NO synthase in the rat CB, suggesting that the removal of the inhibitory NO influence may also contribute to enhance the CB chemosensory responses to hypoxia. Nevertheless, our results also showed that iNOS-ir levels were higher at 21 of CIH exposure. Since iNOS produce higher amounts of NO, it is plausible that the NO level in the CB will increase during long-term CIH. It is worth noting that NO has a dual effect on carotid chemosensory discharges. Indeed, Iturriaga et al. (2000) found that at low levels NO is predominantly an inhibitor of chemosensory discharges, whereas at high concentration NO increases carotid chemosensory discharges. Thus, it is plausible that high NO levels in the CB of rats exposed to 21 days CIH may contribute to maintained CB chemosensory potentiation. Our results showed that intermittent hypoxia produce a transient increase of ET-1-ir in the rat CB by ~ 2-fold, while ET-1 plasma levels remains unchanged. The ET-1 plasma levels measured in this study were in the normal detectable range for rats (Troncoso-Brindeiro et al., 2007). Nevertheless, Kanagy et al. (2001) using radioimmunoassay found a 2-fold increase in the plasma ET-1 levels from rats exposed to 14 days of eucapnic intermittent hypoxia. Since we used a CIH paradigm characterized by short episodes of intermittent hypoxia without raising the PCO2 (Dick et al., 2007; Fletcher et al., 1992; Marcus et al., 2010; Peng et al., 2003; Zoccal et al., 2008), we cannot rule out if the combined effect of hypoxia and hypercapnia may be an effective stimuli for elevating the ET-1 plasmatic levels. Fig. 5 – Time-course of the effects of CIH on the rat CB chemosensory response to acute hypoxia. A, Representatives recordings of CB chemosensory responses to brief hypoxic stimulus (fill bar, inspired PO2 ~ 5 mm Hg) in one Sham rat, and in rats exposed to CIH for 7, 14 and 21 days, respectively. B, Summary of the effects of CIH on CB chemosensory discharges induced by several levels of inspired PO2 (5–670 mm Hg). ƒx, Carotid chemosensory frequency of discharge in Hz. ***, P < 0.001, ** P < 0.01 compared with Sham condition. Newman–Keuls test after 2-way ANOVA. n = 8–10.. noting that present results and the findings of Lam et al. (2006) show that CIH transiently increases the expression of ET-1 in the adult rat CB during the first week of CIH, but later ET-1-ir returned to the sham levels. This observation suggests that ET-1 may contribute to the enhanced CB responsiveness to hypoxia in the early phase of CIH. In addition to the transient changes in ET-1 expression, present results showed a significant decrease in the eNOS expression in the CB at 7 days of CIH that lasted until the 21 days of CIH, suggesting that CIH may decrease the NO levels in the CB. In preliminary experiments, using in vitro CB from rats exposed to 7 days of CIH we found a significant decrease in NO production within the CB (unpublished data). Since NO at low concentration is considered an inhibitory modulator of the CB chemosensory activity (Iturriaga et al., 2000; Rey and. 3.4.. Oxidative stress and nitrotyrosine formation. Our results showing a marked increase of 3-NT immunoreactivity (3-NT-ir) in the CB from rats exposed to CIH for 7 days, which correlates with the enhanced carotid chemosensory responses to hypoxia, support and extend the idea that oxidative/nitrosative stress plays a critical role in CB chemosensory potentiation (Iturriaga et al., 2009; Prabhakar et al., 2010). Although, it has been proposed that ROS contributes to the CB chemosensory potentiation and the cardiovascular alterations induced by CIH, a direct effect for ROS in the CB oxygen chemotransduction process is still unclear (GomezNiño et al., 2009; Gonzalez et al., 2007). Present results suggest that 3-NT formation may contribute to the carotid chemosensory potentiation through the nitration of protein residues. This interpretation agrees with our previous finding that treatment of CIH-rats with ascorbic acid prevents both the carotid chemosensory potentiation and the increased 3-NT formation in the CB of rats exposed to CIH for 21 days (Del Rio et al., 2010). The early increase in 3-NT-ir found in the CB from rats exposed for 7 days for CIH, which remained elevated during the 21 days CIH of treatment, suggests that peroxynitrite formation due to the reaction of NO with the superoxide radical is present during the whole length of the CIH treatment. However, since all NOS isoforms have the potential capacity to synthesize superoxide instead of NO, we cannot preclude the possibility that the increased nitrotyrosine. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
(9) 82. BR A IN RE S EA RCH 1 3 95 ( 20 1 1 ) 7 4 –85. production may simply derive from uncoupling NOS in CBs from rats exposed to CIH. Peroxynitrite is a potent oxidant agent (Ferrer-Sueta and Radi, 2009), with a high rate of formation (up to 16 × 109 M/s). Peroxynitrites can modify tyrosine and tryptophane residues, iron sulfur clusters, zinc thiolates and other residues. Moreover, peroxynitrites may react with inorganic molecules such as CO2 producing other free radicals that may modify DNA, lipids or proteins (FerrerSueta and Radi, 2009). Further studies are required to determine which protein or enzyme complexes involved in the CB chemosensory process are nitrated in animals exposed to CIH. In summary, our results showed that exposure to CIH for 7 days enhances the rat CB chemosensory responses to acute hypoxia, and the 3-NT formation, effects which persisted until 21 days of CIH. CIH produces a transient increase of the ET-1-ir along with a decrease in eNOS-ir detectable at 7 days of CIH, and a progressive late increase of TNF-α, IL-1β and iNOS immunoreactivity in the CB. Thus, present results showed that CIH induced the expression of pro-inflammatory cytokines in the rat CB, suggesting a potential contribution of these chemosensory modulators to the CB chemosensory potentiation.. 4.. Experimental procedures. 4.1.. Animals. Experiments were performed on male Sprague-Dawley rats, weighting initially ~ 200 g. Rats were fed with standard rat chow diet ad libitum, and kept on a 12-hour light/dark schedule (8:00 am–8:00 pm). The experimental procedures to obtain CB tissue samples and to record physiological variables and carotid sinus chemosensory discharges were performed under sodium pentobarbitone anesthesia (40 mg/kg ip), followed by additional doses when necessary to maintain a level of surgical anesthesia. The experimental protocol was approved by the Bio Ethical Committee of the Facultad de Ciencias Biológicas of the Pontificia Universidad Católica of Chile.. 4.2.. Exposure to chronic intermittent hypoxia. Unrestrained freely moving rats were housed in individual chambers (12 cm × 35 cm, 2.2 l) and exposed to a CIH protocol consisting of hypoxic cycles of 5% inspired O2 for 20 s, followed by room air for 280 s, applied 12 times/h for 8 h/day. The chambers were equipped with a rear N2 inlet to produce hypoxic episodes, and a front air extractor, which enables to recover the normoxic level. A computerized system controls the solenoid valve inlets and the alternating cycles of the extractors. During hypoxic exposure, the extractors were stopped, while rear solenoid valves allow 100% N2 flows into the chambers. In the sham condition, the hypoxic cycles were replaced by flushing compressed air into the chambers at the same flow and pressure. The O2 level in the chambers was continuously monitored with an oxygen analyzer (Ohmeda 5120, USA). The CO2 in the chamber was maintained low by. continuous air extraction. The room temperature was kept at 22–25 °C and the hypoxic and sham patterns were applied from 8:00 am to 4:00 pm.. 4.3.. Recordings of carotid body chemosensory discharges. Experiments were performed ~15 to 20 h after the end of the last hypoxic cycle. Pentobarbitone anesthetized rats were placed in supine position and the rectal temperature was maintained at 38.0 ± 0.5 °C with a regulated heating pad. The CB chemosensory discharge was measured as previously described (Del Rio et al., 2010). Briefly, one carotid sinus nerve was dissected and placed on a pair of platinum electrodes and covered with warm mineral oil. The neural signal was pre-amplified (Grass P511, USA), filtered (10–500 Hz) and fed to an electronic spike-amplitude discriminator allowing the selection of action potentials of given amplitude above the noise. The selected potentials were counted with a frequency meter to assess the CB chemosensory frequency of discharge (ƒx), expressed in Hz. Carotid sinus barosensory activity was eliminated by crushing the common carotid arterial wall between the carotid sinus and the CB and confirmed by the disappearance of large barosensory discharges in the electroneurogram. In all CB recording experiments, the contra lateral carotid sinus nerve was cut to prevent vascular and ventilatory effects caused by the activation of chemosensory reflexes. The chemosensory discharge was measured at normoxia and during acute hypoxic stimuli (PO2 ≈ 5 to 670 mm Hg) maintained for 20–30 s. Rats breathed spontaneously during the whole experiment. At the end of the experiments, rats were killed by an overdose of sodium pentobarbitone (100 mg/kg ip.).. 4.4. Immunohistochemichal detection of ET-1, eNOS, iNOS, TNF-α, IL-1β, IL-6 and 3-NT in the rat CB The quantification of the relative expression of ET-1, eNOS and iNOS, TNF-α, IL-1β, IL-6 and 3-NT was performed as previously described (Del Rio et al., 2010). Anesthetized rats were perfused intracardially with phosphate saline buffer (PBS) at pH 7.4 for 10 min followed by buffered 4% paraformaldehyde (PFA, Sigma, USA). The carotid bifurcations containing the CB were dissected and post-fixed in cold 4 °C PFA 4% for 12 h. Samples were then dehydrated in ethanol, included in paraffin, cut in 5 μm sections and mounted on silanized slides. Deparaffinized samples were submitted to microwave based antigen retrieval protocol (700 W for 6 min in citrate buffer 1 M pH 6.0) for unmasking antigens which have become modified by the tissue fixation process. Samples were incubated with 0.3% H2 O2 to inhibit endogenous peroxidase and then in blocking normal horse serum solution (Vectastain Elite ABC Kit, Vector Lab, USA). The slides were incubated with specific antibodies overnight at 4 °C in humidity chambers for detection of ET-1 (T-4050, 1:500, rabbit anti-ET-1, Peninsula Labs., USA), eNOS (N 3893, 1:200, rabbit anti-eNOS, Sigma, USA), iNOS (N 7782, 1:200, rabbit anti-iNOS, Sigma, USA), TNF-α (sc-1350, 1:20, goat anti-TNF-α, Santa Cruz Biotech., USA), IL-1β (sc-7884, 1:100, rabbit anti-IL-1β, Santa Cruz Biotech., USA), IL-6 (sc-1265, 1:50, goat anti-IL-6, Santa Cruz Biotech., USA) and 3-NT (A-21285, 1:20, rabbit anti-3-NT,. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
(10) BR A IN RE S E A RCH 1 3 95 ( 20 1 1 ) 7 4 –8 5. Molecular Probes, USA). Negative controls were performed by omission of the primary antibody. After rinse slides in cold PBS, samples were incubated with proper secondary antibodies conjugated to biotin followed by a ready-to-use stabilized ABC reagent (Vectastain Elite ABC Kit, Vector Lab, USA), and revealed at 37 °C in a dark chamber with 3,3diaminobenzidine tetrahydrochloride (DAB; Sigma, USA). To avoid false positives during DAB chromogen quantification, special attention was kept to prevent DAB signal saturation. For each antibody used, the immunohistochemical procedure from all the four experimental groups was run in parallel to maintained similar conditions. Finally, samples were counterstained with Harris Hematoxylin and permanently mounted with Entellan® (Merck, USA). Photomicrographs of the CB tissue were taken at 100× with a CCD-camera coupled to an Olympus CX 31 microscope (Olympus Corp, Japan), digitized and analyzed with the ImageJ software (NIH, USA). A color deconvolution algorithm (Ruifrok and Johnston, 2001) was used to separate the Harris Hematoxylin and DAB staining, and quantify the immunoreactive mark (− ir) from the RGB (red-green-blue) images. The algorithm deconvolved the color information acquired with the RGB camera and calculated the contribution of each applied stains based on a stain-specific RGB absorption. The positive immunoreactive intensity, averaged from four CB fields (9200 μm2 each) was expressed as optical integrated intensity. Since the mean sectional area of an adult rat CB is around 45,000 μm2, the quantification of the positive-ir covers a representative 80% of the total CB sectional area.. 4.5. Detection of immune cell invasion in the CB of rats exposed to CIH To study if the CB was invaded by immune cells we detected the immune antigen ED-1 by immunofluorescence. The CB tissue samples for immunoflourescence studies were obtained as previously described (Del Rio et al., 2008). Briefly, the carotid artery bifurcations were excised from anesthetized rats, fixed with 4% PFA, and included in paraffin wax as previously described. The CBs were then serially sectioned at a thickness of 5 μm and mounted on silanized slides. Glomus cells were stained with antibodies against tyrosine hydroxylase (TH), a recognized marker of chemoreceptor cells. Tissue sections were exposed to UV light for 30 min and treated with NaBH4 0.1% to quench the tissue autofluorescence. The CB histological sections were incubated for 1 h in blocking serum solution (Vector Lab, USA) followed by incubation with a mixed of an anti-tyrosine hydroxylase antibody (AB152, 1:100, rabbit anti-TH, Millipore, USA) and anti-ED-1 antibody (MCA341GA, 1:100, mouse anti-ED-1, AbD Serotec, USA) overnight at 4 °C in a humidity chamber. After washing with PBS, tissue sections were incubated for 1 h with Alexa-fluor 488 donkey antimouse IgG (A-21202, 1:200, Molecular Probes, USA) and Alexa-flour 594 goat anti rabbit IgG (A11012, 1:200, Molecular Probes, USA) in PBS with 1% BSA and 0.5% Triton X-100. Finally, sections were mounted in DAPI-containing media (Vectashield, Vector Lab, USA) and visualized using a fluorescent microscope (Eclipse E, Nikon, Japan).. 4.6.. 83. Plasma ET-1, TNF-α, IL-1β and IL-6 levels. Enzyme-linked immunoabsorbent assays (ELISA) were used to measure the systemic plasma levels of ET-1, TNF-α, IL-1β and IL-6. Blood samples were collected from one carotid artery and placed in EDTA ice-cold microcentrifuge tubes. The plasma was separated by centrifugation and stored at − 80 °C until used. The ET-1 plasma level was assessed as previously described (Rey et al., 2006) using an ELISA ET-1 kit (#BBE5, R&D Systems, USA). The cytokines plasma concentrations were measured using ELISA kits developed for detection of rat cytokines (TNF-α #ER3TNFA, IL-1β #ER2IL1B, and IL-6 #ER2IL6, Thermo Scientific, USA).. 4.7.. Statistical analysis. The data was expressed as means ± SE. Paired comparisons between 3 or more groups were assessed with one or two-way ANOVA tests, followed by Newman–Keuls posthoc comparisons. All analyses were done with a statistical significance of P < 0.05.. Acknowledgments This work was supported by grant 1100405 from the National Fund for Scientific and Technological Development of Chile (FONDECYT). We would like to thank the assistance of Dr. Julio Alcayaga in the construction and operation of the hypoxic chambers. Rodrigo Del Rio was supported by a CONICYT AT24091043 fellowship.. REFERENCES. Anuar, F., Whiteman, M., Siau, J.L., Kwong, S.E., Bhatia, M., Moore, P.K., 2006. Nitric oxide-releasing flurbiprofen reduces formation of proinflammatory hydrogen sulfide in lipopolysaccharide-treated rat. Br. J. Pharmacol. 147, 966–974. Chen, J., He, L., Liu, X., Dinger, B., Stensaas, L., Fidone, S., 2007. Effect of the endothelin receptor antagonist bosentan on chronic hypoxia-induced morphological and physiological changes in rat carotid body. Am. J. Physiol. Lung Cell. Mol. Physiol. 292, L1257–L1262. Del Rio, R., Moya, E.A., Koenig, C.S., Fujiwara, K., Alcayaga, J., Iturriaga, R., 2008. Modulatory effects of histamine on cat carotid body chemoreception. Respir. Physiol. Neurobiol. 164, 401–410. Del Rio, R., Moya, E.A., Iturriaga, R., 2010. Carotid body and cardiorespiratory alterations in intermittent hypoxia: the oxidative link. Eur. Respir. J. 36, 143–150. Dick, T.E., Hsieh, Y.H., Wang, N., Prabhakar, N.R., 2007. Acute intermittent hypoxia increases both phrenic and sympathetic nerve activities in the rat. Exp. Physiol. 92, 87–97. Dvorakova, M., Hohler, B., Vollerthun, R., Fischbach, T., Kummer, W., 2000. Macrophages: a major source of cytochrome b558 in the rat carotid body. Brain Res. 852, 349–354. Fan, J., Zhang, B., Shu, H.F., Zhang, X.Y., Wang, X., Kuang, F., Liu, L., Peng, Z.W., Wu, R., Zhou, Z., Wang, B.R., 2009. Interleukin-6 increases intracellular Ca2+ concentration and induces catecholamine secretion in rat carotid body glomus cells. J. Neurosci. Res. 87, 2757–2762.. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
(11) 84. BR A IN RE S EA RCH 1 3 95 ( 20 1 1 ) 7 4 –85. Ferrer-Sueta, G., Radi, R., 2009. Chemical biology of peroxinitrite: kinetics, difusion and radicals. ACS Chem. Biol. 4, 161–177. Fletcher, E.C., Lesske, J., Behm, R., Miller, C.C., Stauss, H., Unger, T., 1992. Carotid chemoreceptors, systemic blood pressure, and chronic episodic hypoxia mimicking sleep apnea. J. Appl. Physiol. 72, 1978–1984. Garvey, J.F., Taylor, C.T., McNicholas, W.T., 2009. Cardiovascular disease in obstructive sleep apnoea syndrome: the role of intermittent hypoxia and inflammation. Eur. Respir. J. 33, 1195–1205. Gomez-Niño, A., Agapito, M.T., Obeso, A., González, C., 2009. Effects of mitocondrial poisons on glutathione redox potential and carotid body chemoreceptor activity. Respir. Physiol. Neurobiol. 165, 104–111. González, C., Agapito, M.T., Rocher, A., González-Martin, M.C., Vega-Agapito, V., Gomez-Niño, A., Rigual, R., Castañeda, J., Obeso, A., 2007. Chemoreception in the context of the general biology of ROS. Respir. Physiol. Neurobiol. 157, 30–44. Gozal, D., Kheirandish-Gozal, L., 2008. Cardiovascular morbidity in obstructive sleep apnea, oxidative stress, inflammation, and much more. Am. J. Respir. Crit. Care Med. 177, 369–375. Iturriaga, R., Alcayaga, J., 2004. Neurotransmission in the carotid body: transmitters and modulators between glomus cells and petrosal ganglion nerve terminals. Brain Res. Brain Res. Rev. 47, 46–53. Iturriaga, R., Villanueva, S., Mosqueira, M., 2000. Dual effects of nitric oxide on carotid body chemoreception. J. Appl. Physiol. 89, 1005–1012. Iturriaga, R., Varas, R., Alcayaga, J., 2007. Electrical and pharmacological properties of petrosal ganglion neurons that innervate the carotid body. Respir. Physiol. Neurobiol. 157, 130–139. Iturriaga, R., Moya, E.A., Del Rio, R., 2009. Carotid body potentiation induced by intermittent hypoxia: implications for cardiorespiratory changes induced by sleep apnoea. Clin. Exp. Pharmacol. Physiol. 36, 1197–1204. Janssen-Heininger, Y.M., Poynter, M.E., Baeuerle, P.A., 2000. Recent advances towards understanding redox mechanisms in the activation of nuclear factor kappaB. Free Radic. Biol. Med. 28, 1317–1327. Jelic, S., Le Jemtel, T.H., 2008. Inflammation, oxidative stress, and the vascular endothelium in obstructive sleep apnea. Trends Cardiovasc. Med. 18, 253–260. Kanagy, N.L., Walker, B.R., Nelin, L.D., 2001. Role of endothelin in intermittent hypoxia-induced hypertension. Hypertension 37, 511–515. Lam, S.Y., Tipoe, G.L., Liong, E.C., Fung, M.L., 2006. Hypoxia-Inducible factor (HIF)-1α and Endothelin-1 expression in the rat carotid body during Intermittent. Adv. Exp. Med. Biol. 580, 21–27. Lam, S.Y., Tipoe, G.L., Liong, E.C., Fung, M.L., 2008. Chronic hypoxia upregulates the expression and function of proinflammatory cytokines in the rat carotid body. Histochem. Cell Biol. 130, 549–559. Ling, Y.L., Meng, A.H., Zhao, X.Y., Shan, B.E., Zhang, J.L., Zhang, X. P., 2001. Effect of cholecystokinin on cytokines during endotoxic shock in rats. World J. Gastroenterol. 7, 667–671. Liu, X., He, L., Stensaas, L., Dinger, B., Fidone, S., 2009. Adaptation to chronic hypoxia involves immune cell invasion and increased expression of inflammatory cytokines in rat carotid body. Am. J. Physiol. Lung Cell. Mol. Physiol. 296, L158–L166. Lusina, S.J., Kennedy, P.M., Inglis, J.T., McKenzie, D.C., Ayas, N.T., Sheel, A.W., 2006. Long-term intermittent hypoxia increases sympathetic activity and chemosensitivity during acute hypoxia in humans. J. Physiol. 575, 961–970. Marcus, N.J., Li, Y.L., Bird, C.E., Schultz, H.D., Morgan, B.J., 2010. Chronic intermittent hypoxia augments chemoreflex control of sympathetic activity: role of the angiotensin II type 1 receptor. Respir. Physiol. Neurobiol. 171, 36–45.. McGuire, M., Zhang, Y., White, D.P., Ling, L., 2003. Chronic intermittent hypoxia enhances ventilatory long-term facilitation in awake rats. J. Appl. Physiol. 95, 1499–1508. McQueen, D.S., Dashwood, M.R., Cobb, V.J., Bond, M.S., Marr, C.G., Spyer, K.M., 1995. Endothelins and rat carotid body: autoradiographic and functional pharmacological studies. J. Auton. Nerv. Syst. 53, 115–125. Minoguchi, K., Yokoe, T., Tazaki, T., Minoguchi, H., Tanaka, A., Oda, N., Okada, S., Ohta, S., Naito, H., Adachi, M., 2005. Increased carotid intima-media thickness and serum inflammatory markers in obstructive sleep apnea. Am. J. Respir. Crit. Care Med. 172, 625–630. Mitchell, G.S., Baker, T.L., Nanda, S.A., Fuller, D.D., Zabka, A.G., Hodgeman, B.A., Bavis, R.W., Mack, K.J., Olson Jr., E.B., 2001. Invited review: intermittent hypoxia and respiratory plasticity. J. Appl. Physiol. 90, 2466–2475. Morita, Y., Oda, S., Sadahiro, T., Nakamura, M., Oshima, T., Otani, S., Hirasawa, H., 2009. The effects of body temperature control on cytokine production in a rat model of ventilator-induced lung injury. Cytokine 47, 48–55. Narkiewicz, K., van de Borne, P.J., Montano, N., Dyken, M.E., Phillips, B.G., Somers, V.K., 1998. Contribution of tonic chemoreflex activation to sympathetic activity and blood pressure in patients with obstructive sleep apnea. Circulation 97, 943–945. Narkiewicz, K., Van De Borne, P.J., Pesek, C.A., Dyken, M.E., Montano, N., Somers, V.K., 1999. Selective potentiation of peripheral chemoreflex sensitivity in obstructive sleep apnea. Circulation 99, 1183–1189. Osanai, S., Mokashi, A., Rozanov, C.H., Buerk, D.G., Lahiri, L., 1997. Potential role of H2O2 in chemoreception in the cat carotid body. J. Auton. Nerv. Syst. 63, 39–45. Park, S.J., Chun, Y.S., Park, K.S., Kim, S.J., Choi, S.O., Kim, H.L., Park, J.W., 2009. Identification of subdomains in NADPH oxidase-4 critical for the oxygen-dependent regulation of TASK-1 K channels. Am. J. Physiol. Cell Physiol. 297, C855–C864. Pawar, A., Nanduri, J., Yuan, G., Khan, S.A., Wang, N., Kumar, G.K., Prabhakar, N.R., 2009. Reactive oxygen species-dependent endothelin signaling is required for augmented hypoxic sensory response of the neonatal carotid body by intermittent hypoxia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 296, R735–R742. Peng, Y.J., Overholt, J.L., Kline, D., Kumar, G.K., Prabhakar, N.R., 2003. Induction of sensory long-term facilitation in the carotid body by intermittent hypoxia: implications for recurrent apneas. Proc. Natl. Acad. Sci. U. S. A. 100, 10073–10078. Peng, Y.J., Nanduri, J., Yuan, G., Wang, N., Deneris, E., Pendyala, S., Natarajan, V., Kumar, G.K., Prabhakar, N.R., 2009. NADPH oxidase is required for the sensory plasticity of the carotid body by chronic intermittent hypoxia. J. Neurosci. 29, 4903–4910. Prabhakar, N.R., Kumar, G.K., Nanduri, J., 2010. Intermittent hypoxia augments acute hypoxic sensing via HIF-mediated ROS. Respir. Physiol. Neurobiol. 174, 230–234. Reeves, S.R., Gozal, E., Guo, S.Z., Sachleben Jr., L.R., Brittian, K.R., Lipton, A.J., Gozal, D., 2003. Effect of long-term intermittent and sustained hypoxia on hypoxic ventilatory and metabolic responses in the adult rat. J. Appl. Physiol. 95, 1767–1774. Rey, S., Iturriaga, R., 2004. Endothelins and nitric oxide: vasoactive modulators of carotid body Chemoreception. Curr. Neurovasc. Res. 1, 464–473. Rey, S., Del Rio, R., Alcayaga, J., Iturriaga, R., 2004. Chronic intermittent hypoxia enhances cat chemosensory and ventilatory responses to hypoxia. J. Physiol. 560, 577–586. Rey, S., Del Rio, R., Iturriaga, R., 2006. Contribution of endothelin-1 to the enhanced carotid body chemosensory responses induced by chronic intermittent hypoxia. Brain Res. 1086, 152–159. Rey, S., Corthorn, J., Chacón, C., Iturriaga, R., 2007. Expression and immunolocalization of endothelin peptides and its receptors,. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
(12) BR A IN RE S E A RCH 1 3 95 ( 20 1 1 ) 7 4 –8 5. ETA and ETB, in the carotid body exposed to chronic intermittent hypoxia. J. Histochem. Cytochem. 55, 167–174. Rey, S., Tarvainen, M.P., Karjalainen, P.A., Iturriaga, R., 2008. Dynamic time-varying analysis of heart rate and blood pressure variability in cats exposed to short-term chronic intermittent hypoxia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 295, R28–R37. Ruifrok, A.C., Johnston, D.A., 2001. Quantification of histochemical staining by color deconvolution. Anal. Quant. Cytol. Histol. 23, 291–299. Semenza, G.L., Prabhakar, N.R., 2007. HIF-1-dependent respiratory, cardiovascular, and redox responses to chronic intermittent hypoxia. Antioxid. Redox Signal. 9, 1391–1396. Shu, H.F., Wang, B.R., Wang, S.R., Yao, W., Huang, H.P., Zhou, Z., Wang, X., Fan, J., Wang, T., Ju, G., 2007. IL-1βeta inhibits IK and increases [Ca2+]i in the carotid body glomus cells and increases carotid sinus nerve firings in the rat. Eur. J. Neurosci. 25, 3638–3647. Smith, M.L., Pacchia, C.F., 2007. Sleep apnoea and hypertension: role of chemoreflexes in humans. Exp. Physiol. 92, 45–50. Somers, V.K., White, D.P., Amin, R., Abraham, W.T., Costa, F., Culebras, A., Daniels, S., Floras, J.S., Hunt, C.E., Olson, L.J., Pickering, T.G., Russell, R., Woo, M., Young, T., 2008. Sleep apnea and cardiovascular disease. J. Am. Coll. Cardiol. 52, 686–717.. 85. Troncoso Brindeiro, C.M., da Silva, A.Q., Allahdadi, K.J., Youngblood, V., Kanagy, N.L., 2007. Reactive oxygen species contribute to sleep apnea-induced hypertension in rats. Am. J. Physiol. Heart Circ. Physiol. 293, H2971–H2976. Wang, X., Wang, B.R., Duan, X.L., Zhang, P., Ding, Y.Q., Jia, Y., Jiao, X.Y., Ju, G., 2002. Strong expression of interleukin-1 receptor type I in the rat carotid body. J. Histochem. Cytochem. 50, 1677–1684. Wang, X., Zhang, X.J., Xu, Z., Li, X., Li, G.L., Ju, G., Wang, B.R., 2006. Morphological evidence for existence of IL-6 receptor alpha in the glomus cells of rat carotid body. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 288, 292–296. Weiss, J.W., Liu, M.D., Huang, J., 2007. Physiological basis for a causal relationship of obstructive sleep apnoea to hypertension. Exp. Physiol. 92, 21–26. Williams, A., Scharf, S.M., 2007. Obstructive sleep apnea, cardiovascular disease, and inflammation—is NFkappaB the key? Sleep Breath. 11, 69–76. Zoccal, D.B., Simms, A.E., Bonagamba, L.G., Braga, V.A., Pickering, A.E., Paton, J.F., Machado, B.H., 2008. Increased sympathetic outflow in juvenile rats submitted to chronic intermittent hypoxia correlates with enhanced expiratory activity. J. Physiol. 586, 3253–3265.. Downloaded from ClinicalKey.com at University of Chile Catholic ALERTA May 09, 2016. For personal use only. No other uses without permission. Copyright ©2016. Elsevier Inc. All rights reserved..
(13)
Figure
Documento similar