Entanglement and Drowning of a Magellanic Penguin (Spheniscus magellanicus) in a Gill Net Recorded by a Time depth Recorder in South central Chile
6
0
0
Texto completo
(2) Entanglement and Drowning of a Magellanic Penguin ( Spheniscus magellanicus ) in a Gill Net Recorded by a Time-depth Recorder in South-central Chile KLEMENS PÜTZ1,*, LUCIANO HIRIART-BERTRAND2, 3, ALEJANDRO SIMEONE3, VICTORIA RIQUELME4, RONNIE REYES-ARRIAGADA4, 5 AND BENNO LÜTHI6 1. Antarctic Research Trust, Am Oste-Hamme-Kanal 10, 27432 Bremervörde, Germany. 2. Center for Advanced Studies in Ecology and Biodiversity (CASEB), Departamento de Ecología, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, Casilla 114-D, Santiago, Chile 3. Universidad Andrés Bello, Departamento de Ecología y Biodiversidad, República 470, Santiago, Chile. 4. 5. Instituto de Zoología, Facultad de Ciencias, Universidad Austral de Chile, Casilla 567, Valdivia, Chile. Instituto de Ecología y Evolución, Facultad de Ciencias, Universidad Austral de Chile, Casilla 567, Valdivia, Chile 6. Antarctic Research Trust, c/o Zoo Zürich, Zürichbergstr, 221, 8044 Zürich, Switzerland *Corresponding author; E-mail: [email protected]. Abstract.—Various mitigation measures have been implemented to reduce incidental seabird mortality in longline and trawl fisheries but little attention has been given to artisanal fishing. In the 2008/09 breeding season, during a study of foraging of Humboldt, Spheniscus humboldti, and Magellanic Penguins, S. magellanicus, breeding on Puñihuil islets, southern Chile, a Magellanic Penguin equipped with a time-depth recorder became entangled and subsequently drowned in a gill net set for Corvina Drum (Cilus gilberti). The device was returned by fishermen and the data appear to be the first documented case of such a drowning in a marine, air-breathing vertebrate. According to the data, while diving to a depth of more than 50 m, the bird became entangled and drowned, remaining below 60 m for nearly 21 hours until the net was hauled. Although only a single incident is reported, there are indications that incidental mortality of penguins, other seabirds and marine mammals is more common in artisanal fisheries than previously anticipated. Received 20 April 2010, accepted 5 July 2010. Key words.—artisanal fishery, data-logger, entanglement, gill net, penguin mortality. Waterbirds 34(1): 121-125, 2011. Approximately 80% of seabird species are in decline globally (Birdlife International 2009). A major factor contributing to this trend is mortality by drowning in fishing gear deployed in both commercial and artisanal fishing operations (Moreno et al. 2006; Pütz and Poncet 2009; Zydelis et al. 2009). Consequently, mitigation measures have been developed to reduce albatross and petrel bycatch in longline and trawl fishing (e.g. Cooper et al. 2006; Sullivan et al. 2006a; 2006b; Gonzáles-Zevallos et al. 2007; Watkins et al. 2008). However, bycatch of seabirds other than Procellariiformes, such as gulls, shags and penguins, has raised only local or regional, if any, attention (Zydelis et al. 2009) with only anecdotal reports on penguin mortality due to coastal fishing operations (Gandini et al. 1999; Simeone et al. 1999; Yorio and Caille 1999; Dar-. by and Dawson 2000; Norman 2000; Majluf et al. 2002; Taylor et al. 2002). The Puñihuil islets in south-central Chile are the only place where Humboldt (Spheniscus humboldti) and Magellanic (S. magellanicus) Penguins breed sympatrically (Simeone and Schlatter 1998). However, over the past decades breeding pair numbers have declined, bolstered at least partly by incidental mortality in the unregulated gillnet fishery undertaken in the area (Skewgar et al. 2009). Therefore, a study investigating potential interactions between the penguins’ foraging behavior and the artisanal fishery was initiated. Here, we report on one Magellanic Penguin equipped with a time-depth-recorder that became incidentally entangled in a gill net and subsequently drowned. To our knowledge, this is the first reported case of an incidental mortality actually recorded by a time-depth-recorder.. 121.
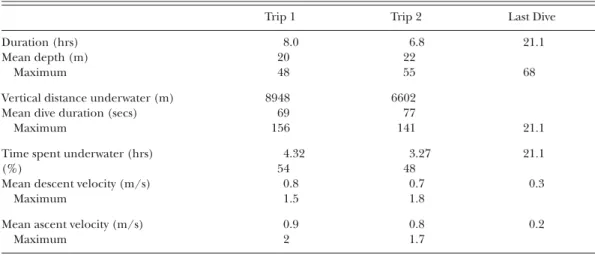
(3) 122. WATERBIRDS METHODS. RESULTS AND DISCUSSION. Field work was conducted between 11 November and 15 December 2008 at the Natural Reserve “Puñihuil islets” (41°55.4’S, 74°2.3’W), located off the northwest coast of Isla Grande de Chiloé, southern Chile. The islets consist of three small islands where ca. 80 Humboldt and ca. 460 Magellanic Penguin breeding pairs nest sympatrically between September and April. In order to investigate the foraging behavior of sympatrically breeding Humboldt and Magellanic Penguins during the breeding season, birds guarding chicks were equipped with MK9 archival tags (Wildlife Computers, Redmond, USA). Devices were attached on the midline of the back using black tape (tesa, Beiersdorf AG, Hamburg, Germany) and glue according to the method described by Wilson et al. (1997). The whole process took less than 15 min per bird. The MK9 used weighed approximately 32 g, equivalent to ca. 1% of the mean penguin body mass, and their maximum dimensions were 88 × 18 × 18 mm with a cross-sectional area of 3.25 cm2, corresponding to about 2% of the penguins’ body cross-sectional area. They were programmed with MK9HOST (Vers. 1.09.1022, Wildlife Computers), with a measurement interval of 3 seconds. The devices were each equipped with a pressure sensor, a light sensor and a temperature sensor. A saltwater switch prevented data being recorded while at the surface. Temperature and light measurements were not considered in this study. For the purpose of this study, only the results obtained from a bird incidentally drowned in fishing gear are considered. The male Magellanic Penguin was equipped with a MK9 on 28 November while brooding two chicks. The device, but not the penguin, was returned on 1 December by local fishermen who had removed it from the drowned bird in the net. Dive data were analyzed using INSTRUMENT HELPER (Vers. 1.0.55, Wildlife Computers). After correcting for a drifting surface level, the following dive parameters were recorded for all dives ≥3 m: onset, duration, maximum depth, descent and ascent rate. Times given are in local summer time, which was GMT -3 hours.. Analysis of the recorded dive data revealed that the first foraging trip of the Magellanic Penguin started on 29 November 2008 at 14:20 local summer time (Table 1). The bird returned to the colony on that day at 22:20 and departed again on 30 November at 06:00. The penguin showed continuous diving activity until 12:55 (Fig. 1), following a series of dives to depths greater than 50 m. The last dive recorded was initially to a depth of 53 m, reached at 12:56, presumably the time when entanglement occurred. The recorded depth then increased continuously to 67 m (first reached at 21:00) and remained at this maximum depth ± 1 m for nearly 13 hours. The ascent phase started at 09:55 on 1 December, lasted seven minutes until 10:03 and was characterized by short pauses at depths of 52 m (60 secs), 38 m (30 secs) and 22 m (15 secs) before reaching the surface. Overall, descent and ascent rates were 0.3 and 0.2 m/s, respectively, and the dive lasted 21 hrs 7 min in total. To our knowledge, this is the first time that entanglement and subsequent drowning of a marine, air-breathing vertebrate has been recorded using a time-depth recorder. For several reasons, we are confident that the penguin studied became entangled in a gill net. First, the device was returned by a local fisherman, albeit without the penguin car-. Table 1. Summary of dive parameters (>3 m) of two foraging trips recorded from a Magellanic Penguin from Puñihuil islets, Chile, drowned in a gill net. The second foraging trip and the last dive recorded where the bird may have drowned are separated.. Duration (hrs) Mean depth (m) Maximum Vertical distance underwater (m) Mean dive duration (secs) Maximum Time spent underwater (hrs) (%) Mean descent velocity (m/s) Maximum Mean ascent velocity (m/s) Maximum. Trip 1. Trip 2. 8.0 20 48. 6.8 22 55. 8948 69 156. 6602 77 141. Last Dive 21.1 68. 21.1. 4.32 54 0.8 1.5. 3.27 48 0.7 1.8. 21.1. 0.9 2. 0.8 1.7. 0.2. 0.3.
(4) DROWNING OF A MAGELLANIC PENGUIN IN A GILL NET. Figure 1. Diving activity of a Magellanic Penguin from Pu huil islets, southern Chile, drowned in a gill net: (a) entire d profile, (b) onset of last dive and (c) end of last dive. Timi given are local summer time (GMT-3).. cass, after being out at sea hauling gill nets. However, the fisherman stated that he had found the bird drowned in the net. Secondly, the ascent phase of the penguin’s diving profile was characterized by several periods without any changes in depth, unlikely to occur in a bird naturally ascending to the surface. Normally, descent and ascent rates were greater than 1 m/s in dives deeper than 50 m (Pütz et al., unpubl. data), whereas in this instance the ascent rate, at 0.2 m/s, was much lower. Thirdly, gill nets are commonly used to catch Corvina Drum Cilus gilberti, hake Merluccius spp., Cusk-eels Genypterus spp. and feral salmon around the Puñihuil islets (Sepulveda et al. 2007; Skewgar et al. 2009). Gill nets used are made of monofilament nylon strings and are kept in vertical position by floats and anchors. The length of the net is variable (from 200 to 800 m) with a mesh size of 5 cm, although nets used to catch Cor-. 123. vina Drum may have a mesh size of up to 20 cm (Simeone et al. 1999). According to local fishermen, nets are set within 20-30 m depth if left at sea during the day and at deeper depths of 60-70 m when left overnight. The latter situation is consistent with our data, where the bird remained at the respective water depths overnight. Furthermore, according to information provided by fishermen (pers. comm. to A. S.), as fish and perhaps other marine animals get entangled over time, nets become progressively heavier and sink to greater depths which could explain the slow, but continuous sinking of the net after the study penguin became entangled. Finally, the frequent stops during hauling are typical as fishermen retrieve their nets manually and need to stop several times to rest. These pauses become shorter during the hauling operation as the net becomes lighter. Thus, we are confident that the penguin did indeed become entangled in a gill net. Incidental entanglement and drowning of penguins in this area has been previously reported in Puñihuil, where at least 50 Humboldt and Magellanic Penguins died during summer 2006 (Skewgar et al. 2009). Although high numbers of drowned birds are not common, chronic entanglement of small numbers (such as the one reported here) is common in Puñihuil (Simeone, pers. obs.). Incidental mortality of penguins has been reported in trawl nets (Gandini et al. 1999; Yorio and Caille 1999; Gonzáles-Zevallos and Yorio 2006; Gonzáles-Zevallos et al. 2007) and longlining gear (Nel et al. 2002; Moreno et al. 2006). Also, entanglement in discarded fishing gear such as nets and nylon fishing lines poses a potential threat and is an occasional source of penguin mortality (Pütz, pers. obs.; Harrigan 1992). However, mortality associated with gill nets appears much more common in Chile (Simeone et al. 1999, Schlatter et al. 2009) and Peru (Majluf et al. 2002), but also elsewhere (Darby and Dawson 2000), than previously anticipated. For example, in March 2009 a mass mortality of nearly 1,380 Magellanic Penguins, both adults (48%) and juveniles (52%), occurred in south-central Chile on a beach near.
(5) 124. WATERBIRDS. Queule (39°23’S, 73°13’W), about 300 km to the north of the Puñihuil islets (Schlatter et al. 2009). Examination of carcasses revealed that the most likely cause of death was asphyxia due to drowning (Schlatter et al. 2009), which was confirmed by several reports from fishermen. However, the real extent of this mortality remains unknown and it can be assumed that many carcasses either sank before being washed ashore or drifted elsewhere. All these incidents imply that fishery-related mortality may reach unacceptable levels not only for local, but also regional and even global penguin populations (c.f. Majluf et al. 2002). Consequently, as in other fishing operations, mitigation measures should be deployed to decrease penguin and also other seabird and marine mammal mortality associated with artisanal fishing along the Chilean and Peruvian coasts (Majluf et al. 2002). In contrast to recent advances in mitigation measures for longline and trawler fishing, only a few studies have investigated potential measures to reduce seabird bycatch in artisanal and gill net fishing (Bull 2007). However, the deployment of acoustic pingers and also an increase in the visibility of the net, for example by using metal oxide/barium sulphate nets, have the potential to significantly reduce seabird and marine mammal bycatch (Melvin et al. 1999; Trippel et al. 2003). Further, the establishment of marine protected areas where human activities are regulated in an appropriate manner without compromising either the survival of seabirds and marine mammals or the economic viability of local fisheries (c.f. Boersma and Parrish 1999) can reduce seabird bycatch. Such has already been proposed for Puñihuil to specifically protect penguins (Skewgar et al. 2009). ACKNOWLEDGMENTS Financial support derived from the Antarctic Research Trust. We are particularly grateful to “Ecoturismo Puñihuil” for organizing the trips to the islands and help with logistics, including lodging. Sphenisco e.V. helped making contacts at Puñihuil. CONAF (Chilean Forest Service), through Hernán Rivera, provided permits to work at Puñihuil Islets and the Subsecretaría de Pesca (Undersecretariat of Chilean Fisheries) provided. permits for handling penguins. The cooperation of the project FONDAP-FONDECYT 1501-0001 is appreciated. Thanks to L. Baxter for proofreading. LITERATURE CITED BirdLife International. 2009. http://www.birdlife.org/ news/news/2009/05/seabird_tracking.html, accessed 18 November 2009. Boersma, P. D. and J. K. Parish. 1999. Limiting abuse: Marine protected areas, a limited solution. Ecological Economics 31: 287-304. Bull, L. S. 2007. Reducing seabird bycatch in longline, trawl and gillnet fisheries. Fish and Fisheries 8: 31-56. Cooper, J., G. B. Baker, M. C. Double, R. Gales, W. Papworth, M. L. Tasker and S. M. Waugh. 2006. The Agreement on the Conservation of Albatrosses and Petrels: Rationale, history, progress and the way forward. Marine Ornithology 34: 1-5. Darby, J. T. and S. M. Dawson. 2000. Bycatch of Yelloweyed Penguins (Megadyptes antipodes) in gillnets in New Zealand waters 1979-1997. Biological Conservation 93: 327-332. Gandini, P. A., E. Frere, A. D. Pettovello and P. V. Cedrola. 1999. Interaction between Magellanic Penguins and shrimp fisheries in Patagonia, Argentina. Condor 101: 783-789. Gonzáles-Zevallos, D. and P. Yorio. 2006. Seabird use of discards and incidental captures at the Argentine hake trawl fishery in the Golfo San Jorge, Argentina. Marine Ecology Progress Series 316: 175-183. Gonzáles-Zevallos, D., P. Yorio and G. Caille. 2007. Seabird mortality at trawler warp cables and a proposed mitigation measure: A case study in Golfo San Jorge, Patagonia, Argentina. Biological Conservation 136: 108-116. Harrigan, K. E. 1992. Causes of mortality of Little Penguins Eudyptula minor in Victoria. Emu 91: 273-277. Majluf, P., E. A. Babcock, J. C. Riveros, M. A. Schreiber and W. Alderete. 2002. Catch and bycatch of seabirds and marine mammals in the small-scale fishery of Punta San Juan, Peru. Conservation Biology 16: 13331433. Melvin, E. F., J. K. Parrish and L. L. Conquest. 1999. Novel tools to reduce seabird bycatch in coastal gillnet fisheries. Conservation Biology 13: 1386-1397. Moreno, C. A., J. A. Arata, P. Rubilarm, R. Hucke-Gaete and G. Robertson. 2006. Artisanal longline fisheries in Southern Chile: Lessons to be learned to avoid incidental seabird mortality. Biological Conservation 127: 27-36. Nel, D. C., P. G. Ryan and B. P. Watkins. 2002. Seabird mortality in the Patagonian Toothfish longline fishery around the Prince Edward Islands 1996-2000. Antarctic Science 14: 151-161. Norman, F. I. 2000. Preliminary investigation of the bycatch of marine birds and mammals in inshore commercial fisheries, Victoria, Australia. Biological Conservation 92: 217-226. Pütz, K. and S. Poncet. 2009. Interactions between seabirds and longline fisheries. Pages 181-184 in Biological studies in Polar Oceans—exploration of life in icy waters (G. Hempel and I. Hempel, Eds.). Wirtschaftsverlag N.W. Gmbh, Bremerhaven, Germany. Schlatter, R. P., E. H. Paredes, J. H. Ulloa, J. Harris, A. Romero, J. Vasquez, A. Lizama, C. Hernandez and A. Simeone. 2009. Mortality of Magellanic Penguins.
(6) DROWNING OF A MAGELLANIC PENGUIN IN A GILL NET (Spheniscus magellanicus) at Queule, Araucania region, Chile. Boletin Chileno de Ornitologica 15: 78-86. Sepulveda, M., M. J. Perez, W. Sielfeld, D. Oliva, L. R. Duran, L. Rodrıguez, V. Araos and M. Buscaglia. 2007. Operational interaction between South American sea lions Otaria flavescens and artisanal (small-scale) fishing in Chile: Results from interview surveys and onboard observations. Fisheries Research 83: 332–340. Simeone, A. and R. P. Schlatter. 1998. Threats to a mixedspecies colony of Spheniscus Penguins in southern Chile. Colonial Waterbirds 21: 418-421. Simeone, A., M. Bernal and J. Meza. 1999. Incidental mortality of Humboldt penguins Spheniscus humboldti in gill nets, central Chile. Marine Ornithology 27: 157-161. Skewgar, E., A. Simeone and P. D. Boersma. 2009. Marine reserve in Chile would benefit penguins and ecotourism. Ocean Coast Management 52: 487-491. Sullivan, B. J., T. A. Reid and L. Bugoni. 2006a. Seabird mortality on factory trawlers in the Falkland Islands and beyond. Biological Conservation 131: 495-504. Sullivan, B. H., P. Brickle, T. A. Reid, D. G. Bone and D. A. J. Middleton. 2006b. Mitigation of seabird mortality on factory trawlers: Trials of three devices to reduce warp cable strikes. Polar Biology 29: 745-753. Taylor, S. S., M. L. Leonard, D. J. Boness and P. Majluf. 2002. Foraging by Humboldt Penguins (Spheniscus. 125. humboldti) during the chick-rearing period: general patterns, sex differences, and recommendations to reduce incidental catches in fishing nets. Canadian Journal of Zoology 80: 700-707. Trippel, E. A., N. L. Holy, D. L. Palka, T. D. Shepherd, G. D., Melvin and J. M. Terhune. 2003. Nylon barium sulphate gillnet reduces porpoise and seabird mortality. Marine Mammal Science 19: 240-243. Yorio, P. and G. Caille. 1999. Seabird interactions with coastal fisheries in northern Patagonia: Use of discards and incidental captures in nets. Waterbirds 22: 207-216. Watkins, B. P., S. L. Petersen and P. G. Ryan. 2008. Interactions between seabirds and deep-water hake trawl gear: An assessment of impacts in South African waters. Animal Conservation 11: 247-254. Wilson, R. P., K. Pütz, G. Peters, B. M. Culik, A. J. Scolaro, J.-B. Charrassin and Y. Ropert-Coudert. 1997. Long term attachment of transmitting and recording devices to penguins and other seabirds. Wildlife Society Bulletin 25: 101-106. Zydelis, R., J. Bellebaum, H. Österblom, M. Vetemaa, B. Schirmeister, A. Stipniece, M. Dagys, M. van Eerden and S. Garthe. 2009. Bycatch in gillnet fisheries – an overlooked threat to waterbird populations. Biological Conservation 142: 1269-1281..
(7)
Figure

Documento similar
In the preparation of this report, the Venice Commission has relied on the comments of its rapporteurs; its recently adopted Report on Respect for Democracy, Human Rights and the Rule
A similar observation was made by Abu-Elwan (1999) in his article about the development of mathematical problem posing strategies for prospective middle school teachers. Indeed,
Most of the researchers argued that classical economist’s view of planned savings is equal to planned investment applicable only in the era of closed economy because of low
This in turn invites a re-drawing of Giannobile’s catalogue (see below) to allow the possibility that the central elements are in fact the figure appearing on the obverse
In this description iot included differen- ces in activity rates associated to different levels of s t icnption according to education achievements of the labour population
The following figures show the evolution along more than half a solar cycle of the AR faculae and network contrast dependence on both µ and the measured magnetic signal, B/µ,
17,18 Contrary to graphene, the band gap in ML-MDS separating the valence and conduction bands is naturally large and due to the absence of inversion symmetry in ML-MDS the
First, by taking advantage of the modular description of gene networks, we have proposed a simple three component mod- ule as a general platform to test the influence that
