Empleo del herbicida Glufosinato de Amonio en la selección de líneas transformadas de banano cv Grande naine (Musa spp AAA) in vitro y en casa de cultivo
104
0
0
Texto completo
(2) Dedicatoria. A mi hija que me ha acompañado en todos los momentos decisivos de mi vida A mi esposo por su apoyo y ayuda diaria..
(3) Agradecimientos. Todo el que ha tenido la oportunidad de realizar un trabajo de esta magnitud, sabe cuan difícil resulta la redacción de estas líneas por el temor de olvidar a alguna persona que nos haya ofrecido su ayuda y también porque se nos hace necesario decirles gracias a todos. A mis tutores Dra Idalmis Bermudez Caraballoso y Dr Rafael Gómez Kosky por apoyarme en todo momento y brindarme sus conocimientos científicos. Al colectivo del laboratorio de Embriogénesis y transformación por su gran cooperación y amistad. A todo el colectivo del IBP por su cooperación y estímulo en todo momento. A la Dra Novisel Veitía por su gran ayuda en el procesamiento estadístico y conducción del documento de tesis. A mis amigas Maite y colegas de la Maestría. A todos, MUCHAS GRACIAS.
(4) ____________________________________________________Resumen. RESUMEN La presente investigación tuvo como objetivo con el empleo del glufosinato de amonio seleccionar in vitro y en casa de cultivo líneas transformadas de banano cv. Grande naine (Musa spp. AAA), que tienen como marcador de selección el gen. bar. Se determinó la concentración mínima inhibitoria de glufosinato de. amonio al estudiar diferentes concentraciones sobre agregados celulares embriogénicos de banano (5,0; 10,0; 15,0; 20,0 y 25,0 mg.l-.1), en brotes cultivados in vitro (0; 1,0; 1,5; 2,0; 3,0 y 4,0 mg.l-.1) y plantas en casa de cultivo de cultivo (0; 5,0; 10,0; 20,0; 30,0.y 40,0 g.l-1) para su uso en la selección. Se evaluó la respuesta con la concentración mínima inhibitoria en los brotes cultivados in vitro y plantas en casa de cultivo de líneas de banano modificadas genéticamente con los plasmidios pHCA58, pHCG59 y pHGA91. Se evaluó la respuesta de nuevas líneas transformadas de banano que portan el plasmidio pHCG59, en fragmentos de hojas de plantas de campo empleando la concentración mínima inhibitoria de glufosinato de amonio y concentraciones menores (7,5; 15,0 y 30,0 g.l-1). Como resultado se determinó que la concentración mínima que inhibe el crecimiento en agregados celulares embriogénicos de banano cv. Grande naine fue de 20,0 mg.l-1 de glufosinato de amonio, en brotes in vitro 3,0 mg.l-1 y en plantas en casa de cultivo 30,0 g.l-. Se demostró la no expresión del gen bar mediante la evaluación de la respuesta de los brotes in vitro y plantas en casa de cultivo de líneas de banano cultivar Grande naine (Musa spp. AAA) transformadas frente a la concentración mínima inhibitoria, previamente determinada, de glufosinato de amonio, lo cual fue comprobado a través del chequeo molecular Reacción en cadena de la polimerasa (RCP). Además a partir del protocolo desarrollado con el empleo de fragmentos de hojas de plantas de campo cultivados in vitro en medio de cultivo agar-agua al 1% y 30,0 g.l-1de glufosinato de amonio se logró diferenciar las dos líneas transformadas del control no transformado, a partir de la expresión del gen bar. Ambas líneas fueron positivas en el análisis molecular de RCP. Estos resultados permiten disponer de una herramienta útil en la transformación genética del banano..
(5) Índice. INDICE Pag.. 1. INTRODUCCIÓN. 1. 2. REVISIÓN BIBLIOGRÁFICA. 5. 2. 1 Origen y distribución. 5. 2.1.2 Clasificación. 5. 2.1.3 Generalidades del cultivo de la Papaya. 6. 2.1.4 Importancia económica del cultivo de la papaya. 7. 2.2 Principales variedades cultivadas en Cuba. 8. 2.3 Cultivo de la papaya. 9. 2.4 Propagación de la papaya. 10. 2.4.1 Propagación por semillas y esquejes. 10. 2.5 Cultivo in vitro. 13. 2.5.1 Organogénesis. 14. 2.5.2 Embriogénesis somática. 14. 2.6 Embriogénesis Somática Secundaria o Repetitiva. 15. 2.7 Factores que afectan la Embriogénesis Somática. 16. 2.7.1 Genotipo. 16. 2.7.2 Explante. 16. 2.7.3 Medio de Cultivo. 17. 2.7.4 Condiciones de Cultivo. 17. 2.8 Ambiente in vitro. 18. 2.8.1 El ambiente in vitro y la embriogénesis somática. 18. 2.9 Embriogénesis somática en papaya. 20.
(6) Índice 2.9.1 Embriogénesis Directa. 21. 2.9.2 Obtención y multiplicación de los embriones somáticos. 21. 2.9.3 Germinación de los embriones somáticos. 22. 2.9.4 Enraizamiento y aclimatización del cultivo de la papaya. 22. 2.9.4.1 Enrraizamiento in vitro. 23. 2.9.4.2 Enraizamiento ex vitro. 23. 2.9.4.3 Aclimatización. 24. 3. MATERIALES Y MÉTODOS. 26. 3.1 Obtención de los embriones somáticos a partir de embriones cigóticos.. 28. 3.1.1 Influencia de la época del año. 28. 3.1.2. 29. Influencia del tipo de frasco de cultivo. 3.2 Multiplicación secundaria de los embriones somáticos. 30. 3.2.1 Efecto de la auxina 2,4-D.. 30. 3.2.2 Influencia del número de grupos de embriones por frasco. 31. 3.3 Germinación de los embriones somáticos. 31. 3.3.1 Influencia del tipo de frasco. 31. 3. 4 Conversión. 33. 3.4.1 Influencia del tipo de cobertor en la conversión.. 33. 3.4.2 Influencia de la altura de las plantas en la conversión. 35. 4-RESULTADOS Y DISCUSIÓN. 37. 4.1 Obtención de los embriones somáticos a partir de embriones cigóticos. 37. 4.1.1 Influencia de la época del año. 37. 4.1.2 Influencia del tipo de frasco de cultivo. 40.
(7) Índice 4.2 Multiplicación secundaria de los embriones somáticos.. 43. 4.2.1 Efecto de la auxina 2,4-D.. 43. 4.2.2 Influencia del número de grupos de embriones por frasco. 45. 4.3 Germinación de los embriones somáticos. 47. 4.3.1 Influencia del tipo de frasco. 47. 4.4 Conversión. 52. 4.4.1 Influencia del tipo de cobertor en la conversión.. 52. 4. 4.2 Influencia de la altura de las plantas en la conversión. 56. 5. CONCLUSIONES. 60. 6. RECOMENDACIONES. 61. 7. BIBLIOGRAFÍA. 62.
(8) Índice.
(9) Introducción.. 1. Introducción. La papaya (Carica papaya L.) es oriunda de América Central y se caracteriza por ser un cultivo productivo en corto tiempo y de forma continua durante todo el año. La producción mundial en el 2006 ascendió a 6 708 551 toneladas (FAO, 2006). Por lo tanto tiene gran importancia económica a nivel mundial, debido a que se puede consumir como fruta fresca o procesarse para obtener otros productos como dulces, jaleas, licuados y encurtidos. También posee un gran potencial de industrialización en el área farmacéutica, culinaria, médica, industria cervecera y bebidas no alcohólicas (Acuña, 2005). En Cuba en los últimos años se ha incrementado el área de cultivo de la papaya, sin embargo, los niveles de producción son bajos y la oferta no se corresponde con la demanda, esto ha sido causado principalmente por las enfermedades virales, las cuales son capaces de disminuir en más de un 50% la producción de las plantaciones (Arocha et al., 2002). A esto se adicionan las lesiones provocadas por hongos sobre los frutos post-cosecha, debido a la rápida maduración que presentan los mismos, lo cual limita su aceptación en el mercado, por este motivo existe la necesidad de buscar alternativas para aumentar la propagación y comercialización de esta especie. La principal variedad que se cultiva en Cuba es la Maradol rojo, que a pesar de presentar numerosas ventajas tiene entre sus limitantes, la susceptibilidad a las principales enfermedades virales, además de la rápida maduración de los frutos post-cosecha (Pestano, 2001). La forma tradicional de propagación de la papaya, ha sido la reproducción sexual (por semillas), también puede ser propagada por medio de estacas o injertos aunque este. 1.
(10) Introducción. método de propagación no brinda los efectos deseados, las estacas son de lento desarrollo y los injertos degeneran y no mantienen las características deseadas. Para obtener semillas de calidad los frutos deben provenir del cruzamiento entre plantas hermafroditas, de esta manera se puede lograr un 66% de plantas hermafroditas y 33% de plantas femeninas, con esta selección existe la certeza de no aparición de plantas masculinas no productivas (Otero, 2003). La biotecnología puede acelerar los programas convencionales de propagación masiva de plantas y dar soluciones cuando los métodos convencionales fallan. En la actualidad se han desarrollado diferentes métodos de regeneración de plantas a partir del cultivo de tejidos tanto por organogénesis (Hossain et al., 1993, Vegas et al., 2003) como por embriogénesis somática (Posada, 1995; Del Sol et al., 2001, Gallardo, 2006), sin embargo por ser los métodos biotecnológicos hasta el presente más costosos respecto a la vía por semilla botánica su empleo esta limitado solamente para genotipos híbridos que lo justifiquen (Elder y Macleod, 2000). La organogénesis compite con la semilla botánica por los altos costos, por lo que es necesario buscar un sistema de propagación eficiente que permita la propagación de esta especie, la embriogénesis somática puede ser una vía alternativa, ya que, ofrece mayores posibilidades de obtener volúmenes de producción superiores en un menor período de tiempo, lo cual la convierte en un método potencialmente más eficiente que la regeneración vía organogénesis (Villalobos y Torpe, 1991). Durante la evaluación en campo de plantas proveniente del cultivo de tejido se observó que el 100% de las plantas de papaya obtenidas por embriogénesis somática evaluada en campo, han resultado ser hermafroditas, este resultado es de gran importancia ya que los frutos obtenidos de estas plantas presentan una mayor calidad, en cuanto a. 2.
(11) Introducción. tamaño, color de la pulpa,. muy buen sabor y alto contenido de azúcares lo que. conlleva a una mayor demanda en el mercado comercial Gómez (2008) comunicación personal. En el cultivo de la papaya varias metodologías de regeneración de plantas vía embriogénesis somática han sido establecidas, pero, su fin fundamental ha estado dirigido al mejoramiento genético, una se basa en el uso como explante inicial de embriones cigóticos inmaduros (Posada, 1995) y la otra con el empleo de plantas in vitro de papaya (Gallardo, 2006). Sin embargo este proceso de la morfogénesis in vitro también puede ser utilizado para la producción de semillas de plantas élites en este cultivo. No obstante no se encontraron en la literatura científicas consultadas referencias sobre el uso de este método de propagación a nivel comercial. Contar con procedimientos eficientes para la propagación masiva de esta especie permitirá mejorar la propagación comercial y constituirá una herramienta para el desarrollo de nuevas variedades mediante la transformación genética. (Vargas et al., 2004) Si se tiene en cuenta estos criterios y los antecedentes mencionados, se justifica realizar estudio para realizar innovación a un protocolo mejorado vía embriogénesis somática en papaya variedad Maradol rojo para regenerar plantas a nivel comercial. Por lo antes expuesto se planteó la siguiente hipótesis de trabajo: ” La estandarización a escala productiva de un protocolo para la obtención de plantas de Carica papaya L. a partir de embriones cigóticos permitirá contar con una vía alternativa para la propagación masiva de esta especie” A partir de esta hipótesis planteada se definieron los siguientes objetivos:. 3.
(12) Introducción. 1. Determinar la influencia de la época del año y del tipo de frasco de cultivo en la formación de embriones somáticos. 2. Determinar la concentración de 2,4-D, así como la cantidad de explantes por frasco de cultivo más adecuada para la multiplicación de los embriones somáticos y la influencia del tipo de frasco de cultivo durante la fase de germinación. 3. Evaluar el tipo de cobertor y la influencia de la altura, el tipo de raíz de las plantas de papaya procedentes de embriogénesis somática en la supervivencia en casa de cultivo.. .. 4.
(13) Introducción.. 5.
(14) 2. Revisión Bibliográfica 2.1 Generalidades del cultivo Se ha considerado a la península Malaya en Asia como probable centro de origen primario, tanto de Musa balbisiana como de Musa acuminata cuyos cruzamientos dieron origen a todas las variedades comestibles conocidas en América (Belalcázar, 1991; Jones 2000). 2. 2 Botánica La planta a pesar de tener una apariencia de árbol, se trata en realidad de una herbácea perenne del género Musa, con falso tallo de forma cilíndrica que está formado por las vainas de las hojas superpuestas, un cormo y un sistema radicular fibroso. Pertenece a la clase monocotiledónea, orden Zingiberales y a la familia Musaceae, la cual incluye los géneros Musa y Ensete según (Simmonds., 1962). Las musáceas comestibles usualmente se dividen en dos tipos: bananos y plátanos 2.3. Importancia Los bananos y los plátanos son un componente importante en la alimentación de millones de personas en el mundo tropical, que encuentran en ellos una fuente excelente de carbohidratos, minerales y vitaminas para satisfacer parcialmente sus necesidades diarias, y una fuente de divisas para aquellos países que exportan estas frutas frescas a los mercados antes mencionados. (FHIA 2007). La generalización de este cultivo en América fue debido a la facilidad de propagación, diversas formas de consumo y a la aptitud para producir bebidas fermentadas a partir de la pulpa madura (López, 1989). 2.4. Cultivar Grande naine.
(15) El cultivar Grande naine es hoy la principal variedad de banano que se planta a nivel mundial. Este cultivar tiene entre sus principales ventajas: la altura de la planta alrededor de los 2.5 m de altura, que lo hace más fácil de manejar y más estable al daño del viento, un índice de cosecha más alto que otros cultivares y el intervalo de emergencia-floración y floración-madurez fisiológica es de 196 y 122 días respectivamente, lo que hace un ciclo completo de 318 días (Soto, 1985).. 2.5. Cultivo de tejidos en plátanos y bananos. El principal inconveniente en plátanos y bananos para su mejoramiento genético por métodos tradicionales es el hecho de su esterilidad y poliploidía, lo que implica muchos años de trabajo para crear un nuevo cultivar o diseminar un material genético con interés para la agricultura (Vuylsteke, 2001). Esto crea la necesidad de buscar nuevas alternativas basadas en las técnicas de cultivo de tejidos e ingeniería genética, para complementar programas de mejora genética, así como introducción de nuevos clones. Dos principales métodos de regeneración in vitro han sido utilizados para la propagación, la organogénesis y la embriogénesis. La regeneración de plantas por una y otra vía depende de las características genéticas de las plantas y del manejo del cultivo in vitro, que tiene en cuenta la manipulación, los medios de cultivo y otras condiciones ambientales (Tisserat, 1991; Gómez, 1997). La embriogénesis somática no es un fenómeno artificial y es conocido en la naturaleza como una forma de apomixis llamada embrionía adventicia, por primera vez descrita por Strasburges en 1878. Aunque fueron Steward et al. (1958) y Reinert (1958), quienes dieron créditos por primera vez a la descripción de la embriogénesis somática. Este método ha sido descrito para un gran número de especies (Tisserat et al., 1979; Krishnaraj y Vasil, 1995); sin embargo la inducción del embrión somático y la regeneración de plantas todavía no son procesos eficientes (Parrot, 1993). A pesar de ello este método es considerado como el más eficiente para la producción masiva de.
(16) plantas in vitro, (Preil, 1991), el proceso productivo puede ser automatizado, logrando altos coeficientes de multiplicación en cortos períodos de tiempo, al poder aplicarse los principios de la cinética microbiana y la posibilidad de encapsular estas estructuras y obtener semillas artificiales (Redenbaugh, 1986).. Además es un. sistema de. regeneración a nivel celular útil para la transformación genética e hibridación somática (Georget et al., 2000) En Musa corresponde el primer informe sobre la embriogénesis somática a Cronauer y Krikorian (1983), quienes lograron embriones a partir de suspensiones celulares derivadas de ápices cultivados in vitro de los triploides Saba y Pelipita (ABB). Novak et al. (1989), con callos formados a partir de rizoma en cultivares diploides y triploides, lograron suspensiones embriogénicas. Después se publicó la obtención de suspensiones celulares y regeneración de plantas a partir de yemas neoformadas del cultivar triploide Bluggoe (ABB) (Dhed´a et al., 1991). Dhed´a (1992) desarrolló una metodología para el establecimiento de suspensiones celulares embriogénicas a partir de scalps, estos son explantes derivados de brotes adventicios muy proliferantes; después de un mes en el medio de cultivo de inducción, las estructuras nodulares llamados glóbulos meristemáticos, aparecieron sobre la superficie del scalps. El cultivo continuo de estos glóbulos meristemáticos en medio de cultivo líquido dio lugar a la formación de suspensiones celulares embriogénicas. Dicha metodología fue mejorada y empleada en otros cultivares por Schoofs (1997). Escalant et al. (1994) y Gómez et al. (2002), utilizando sistemas de inmersión temporal, lograron aumentar los porcentajes de embriones somáticos germinados, mientras que Grapin et al. (1996) y Cote et al. (1996), del CIRAD-FHLOR obtuvieron suspensiones celulares con adecuadas porcentajes de regeneración cuando emplearon como explante inicial flores masculinas. Resultados importantes en la embriogénesis somática en plátanos y bananos han sido logrados por Daniels (2003) en el cultivar híbrido de plátano FHIA-21 utilizando este mismo explante. También se han formado callos con células embriogénicas a partir de explantes de flores.
(17) femeninas inmaduras, las cuales son extraídas de la planta en la etapa de transición de la fase vegetativa a la floral, se eliminan las hojas y se abre el pseudotallo para extraer la inflorescencia joven (Grapin et al., 2000). Chong (2003) logró obtener suspensiones celulares embriogénicas al cultivar directamente en medios de cultivo líquidos los agregados florales masculinos del cultivar de banano Gran Enano (Musa AAA). La micropropagación vía organogénesis mediante yemas axilares, a pesar de no ser el método más rápido, es el más ampliamente utilizado para la propagación comercial de Musa. Un factor importante a tener en cuenta en la propagación por esta vía es, la edad del cultivos in vitro o el número de subcultivos, incrementándose con él, el número de brotes por explante y la formación de yemas adventicias (Orellana, 1998). 2.6. Transformación genética en plátanos y bananos La transformación genética de plantas puede ser definida como la introducción o integración estable de genes en el genoma de una planta y su expresión en la planta transgénica (Sagi et al., 1998). El proceso involucra la introducción de por lo menos dos genes (un gen marcador de selección y un gen de interés), pero el número de genes introducidos simultáneamente puede ser mayor (Sági et al., 1995). En cultivares de bananos y plátanos ofrece una vía atractiva para la introducción de genes simples agronómicamente importantes (Khanna et al., 2004) y de genes capaces de codificar para nuevas funciones en el organismo (resistencia a plagas y enfermedades o herbicidas, resistencia al frío o la sequía) Una oportunidad para la transformación genética eficiente de bananos es la introducción directa del ADN por electroporación en protoplastos viables y altamente regenerables (Fromm et al., 1985). La expresión transciende de los genes introducidos por electroporación en protoplastos de banano de cocción cultivar “Bluggoe” fue de una frecuencia de 1.8 % (Sagi et al., 1994)..
(18) El uso de meristemos y la transformación por Agrobacterium tumefaciens resulta altamente riesgoso por la regeneración de plantas quiméricas, por lo que es necesario un segundo ciclo de propagación de plantas bajo selección con antibióticos, lo que trae consigo la reducción diez veces del número de plantas transgénicas obtenidas durante el primer ciclo selectivo (May et al., 1995). Swennen et al., (2003) transformaron más de 12 000 meristemos de los cultivares Gran Enano y William con Agrobacterium tumefaciens y la frecuencia de posibles plantas transgénicas fue de 0.5 % estos valores indicaron que este tipo de tejido no son el blanco ideal para la transformación genética en este cultivo. La transformación de meristemos puede producir quimeras que no son fácilmente identificadas ni disociadas (Roux et al., 2001). La técnica más utilizada para la generación de bananos transgénicos ha sido el uso de suspensiones celulares y transformación por biobalística (Becker et al., 2000), que se basa en la aceleración a altas velocidades de micropartículas generalmente de tungsteno u oro, recubiertas con ADN, las cuales pueden atravesar la pared y membranas celulares (Sagi, 2000). Los agregados celulares de suspensiones de células embriogénicas bombardeadas del cultivar “Bluggoe” se desarrollaron en el medio selectivo y regeneraron transformantes que fueron confirmados en el análisis de hibridación del ADN (Sagi et al., 1995). Sin embargo, ha sido posible combinar exitosamente las suspensiones celulares y Agrobacterium tumefaciens. Hernández et al., (1999) investigaron el movimiento quemotáctico y el ataque de Agrobacterium tumefaciens a las células y tejidos de bananos indicando que esos procesos no limitan la transformación de bananos y plátanos. Ganapathi et al. (2001) reportaron la transformación mediada por Agrobacterium tumefaciens de suspensiones de células embriogénicas del cultivar Rasthali (AAB). Pineda et al. (2002) transformaron células embriogénicas en suspensión de “Dominico Hartón” (AAB), basado en la expresión transitoria del gen de la -glucoronidasa (GUS) mediante Agrobacterium tumefaciens..
(19) Khanna et al. (2004) desarrollaron un sistema eficiente de transformación de suspensiones de células embriogénicas de los cultivares de banano Gran Enano (AAA) y Lady Finger (AAB) mediante Agrobacterium tumefaciens. Este aparece como el primer reporte de obtención de banano transgénico por esta vía en estos cultivares usando. los. genes. hpt. (Higromicina. fosfotransferasa). y. npt. II. (Neomicina. fosfotransferasa). Estos autores lograron una alta eficiencia de transformación 65 plantas en 50 mg del VCS, sin embargo, estos solo fueron estudios preliminares para el establecimiento del protocolo de transformación no llegando a obtener plantas con resistencia a una enfermedad determinada y mucho menos el estudio en condiciones de campo de las plantas transformadas. Hasta el momento. solo se ha reportado el trabajo de Gómez Lim et al., (2004). relacionado con la transformación genética usando biobalística con genes para la resistencia a la enfermedad Sigatoka negra que incluyen quitinasas, glucanasas y un gen antifúngico de Capsicum anuum Las técnicas de transformación genética representan una alternativa para obtener los genotipos de Musa resistentes a la raya negra de la hoja (Sweenen, 2003) El desarrollo de suspensiones celulares embriogénicas ha dado la oportunidad para el establecimiento del cultivo de protoplastos regenerables que pueden utilizarse también como otro tipo de explante para la transformación genética (Megia et al., 1993; Panis et al., 1993; Sagi et al., 1994). De acuerdo con Pérez (2000), las suspensiones de células embriogénicas y la técnica de cultivo de protoplastos abren una tercera ruta alternativa para la generación de plantas de bananos transgénicas. La transformación genética se inició desde los años 90 en China, que fue el primer país que cultivó tabaco transgénico a Virus (Roush, 2001), en 1944, la compañía Calgene desarrolló un tomate de maduración retardada (Flavr-Sarr), el cual se convirtió en el primer productor alimenticio de plantas transgénicas. En los últimos años los cultivos transgénicos han llegado a los campos del agricultor en por lo menos.
(20) 10 países industrializados y 11 en desarrollo. En el 2005, el área total de cultivos transgénicos plantados en campos alcanzó 90 millones de hectáreas, 9 millones hectáreas más que en el 2004 (James, 2005). Se estima que en los mercados de Estados Unidos y otros países, más del 60% de los alimentos procesados como las pizzas, hojuelas de papa, galletas, helados, la miel de maíz contengan desde los años 90, ingredientes derivados de soya, maíz y canola transgénicos (Roca, 2002).. 2.6.1. Métodos de transformación En los vegetales la información genética se puede introducir por cuatro tipos de técnicas: Transformación de protoplastos, Transformación por electroporación, por biobombardeo y Transformación mediada por Agrobacterium. A pesar de las diferentes técnicas, la opción más utilizada para la introducción en las plantas del material genético adicional es la mediada por la bacteria agrobacterium tumefaciens. El mecanismo de integración de este ADN está dirigido por un conjunto de genes bacterianos (los llamados genes vir, por virulencia) cuya expresión se induce en presencia de compuestos fenólicos liberados por las propias plantas en crecimiento, como la acetosiringona (Hansen y Chilton, 1999). 2.6.2. Marcadores de selección. Los marcadores de selección pueden dividirse en varias categorías, dependiendo de si confieren selección positiva (marcadores que permitan el desarrollo de las plantas transformadas) o negativas (estos por el contrario, causan la muerte de las células transformadas), o de si esa selección es condicional (requiere) o no condicional (no requiere) la presencia de sustratos externos (Babwah y Wadell, 2000; Miki y Mchugh, 2004). Los primeros marcadores desarrollados fueron los de tipo positivo y condicionales que necesitan de la presencia de antibióticos, herbicidas o compuestos químicos, normalmente tóxico para las plantas sin transformar. De las publicaciones científicas relacionadas con este tema en el año 2002, el 90 por ciento utilizaban tres.
(21) de estos sistemas, resistencia a Kanamicina, a Higromicina (ambos antibióticos) y resistencia al herbicida fosfinotricina (Miki y Mchungh, 2004) Más recientemente se han desarrollado marcadores de selección positivos condicionales, donde se utilizan agentes no tóxicos que pueden ser sustratos que producen crecimiento y diferenciación de las células o tejidos transformados (Fernández, 2006). Este es el caso, del gen manA que codifica para la fosfomanosa isomerasa; si no se introduce este gen de origen bacteriano no puede ser utilizada la D-manosa como fuente de carbono por la planta (Joersbo et al., 1998). Leyman et al., (2005) utilizaron la sobre expresión del gen AtTPS1 que codifica para la trialosa 6fosfato sintetasa, en brotes de tabaco (Nicotiana tabacum) y en la germinación de semillas de Arabidopsis taliana, para reducir la sensibilidad a glucosa en plantas transgénicas sin causar efectos negativos. Otros ejemplos de este tipo son los genes xyla y uida, que codifican respectivamente para la xilosa isomerasa y -glucoronidasa y. permiten. el. crecimiento. sobre. D-xilosa. y. benciladenil-N-3-glucorónido,. respectivamente (Joersbo y Okkels., 1996; Haldrup et al., 1998a y 1998b). Ejemplo. de. este. tipo. son. los. genes. ipt. y. pgazz. que. codifican. para. isopenteniltransferasa y modifican el nivel endógeno de fitohormonas, y así promueven el desarrollo de tallos (Endo et al., 2001; Zuo et al., 2002a) Otros ejemplos de este tipo son los genes rol, ESRI y CKII (Ebinuma et al., Banno y Chua., 2002; Zuo et al., 2002b). Algunos de los efectos generados por este tipo de marcadores, aunque no tóxicos, tampoco son recomendables para el desarrollo vegetal, sobre todo si se tiene en cuenta que el fenotipo de selección sólo es necesario hasta que ésta concluye. Por este motivo, estos genes normalmente se introducen junto con promotores de naturaleza inducible, que permiten la expresión del gen sólo en determinadas condiciones. Para la transformación se recomienda eliminar todos los genes de resistencia a antibióticos como marcadores de selección en plantas (FAO/WHO, 2000; EFB, 2001). Aunque la presencia de estos genes, no tuviera repercusiones desde el punto de vista.
(22) de la salud o implicaciones ecológicas, la presencia de un marcador de selección con otras características, es algo que puede disminuir la aceptación pública de los organismos geneticamente modificado. Es por ello que ha cobrado impulso la investigación con el empleo de herbicidas.. 2.6.3. Herbicidas como marcadores de selección en plantas transformadas El descubrimiento de los dos genes, bar o pat, que confieren resistencia al herbicida FInalé, le ha permitido a la biotecnología desarrollar cultivos transgénicos, a partir de la inserción de estos genes en las plantas y utilizarlos como marcadores de selección en los procesos de transgénesis (Zeng et al., 2004; Hu et al., 2005). El gen bar que codifica para la enzima fosfinotricina acetiltransferasa (PAT), que confiere resistencia a la fosfinotricina, compuesto activo del herbicida Finalé®, es usado en las transformaciones de varios cultivos, ya sea como gen de interés o de selección. (OECD, 1999). Este gen fue aislado de cultivos de Streptomyces hygroscopicus, y algunos investigadores plantean que su nombre se debe a la resistencia que le confieren al tripéptido bialaphos. Por otra parte, esta el gen pat, que aunque similar al gen bar, codifica para la misma enzima, pero fue aislado de cultivos de Streptomyces viridochromogenes (OECD, 1999). Estos genes son responsables de la destoxificación del herbicida glufosinato de amonio, ya que la enzima provoca una acetilación que convierte a la molécula de glufosinato en una sustancia no tóxica (Mulwa et al., 2006). 2. 7 Características generales del herbicida Finalé® El glufosinato es un compuesto que actualmente se usa como herbicida no-selectivo que ha tenido gran aceptación en la agricultura moderna por las ventajas que ofrece, ya que solo actúa sobre las partes verdes de la planta, por esta propiedad se encuentra predestinado para uso en fruticultura y en cultivos altos. También debido a.
(23) su amplio espectro de acción, el glufosinato de amonio es utilizado para eliminar malas hierbas competidoras (Bayer CropScience Iberia, 2006). En comparación con otros herbicidas de contacto, de acción más rápida, el glufosinato de amonio es más duradero, lo que permite reducir el número de aplicaciones para el control de las malas hierbas. Además, su uso es adecuado en zonas con peligro de erosión y en áreas agrícolas con elevadas exigencias de seguridad ecológica (Bayer CropScience Iberia, 2006). El glufosinato de amonio es actualmente el compuesto activo de varios herbicidas, pues este contiene intrínsecamente al isómero L- de la fosfinotricina (L-PPT), responsable de la actividad herbicida, puede ser obtenido tanto por un proceso de fermentación de dos tripéptidos u obteniendo de forma sintética el glufosinato de amonio. Este compuesto fue registrado en EE.UU. para ser usado como herbicida en 1993 por la compañía Hoechet Celanese (US, EPA, 1993). En la actualidad, el glufosinato de amonio está registrado en más de 80 países bajo diferentes nombres tales como Basta, Liberty, Finale, Rely, entre otros, y se aplica con éxito en más de 100 cultivos (Bayer CropScience, 2005).. 2.7.1. Modo de acción del herbicida Finalé® La fosfinotricina es un aminoácido, 4-[hidroxil-(metil) fosfinoil]-D,L-homoalanina. El Lisómero de fosfinotricina (L-PPT) es ampliamente usado como agente controlador de un amplio espectro de hierbas y está registrado en varios países, su uso como herbicida. El D-isómero (D-PPT) no exhibe actividad herbicida. Por otra parte, el glufosinato de amonio es un equimolar, una mezcla racémica de los dos isómeros L- y D- de la fosfinotricina. El L-PPT es sintetizado solo por dos géneros de bacterias Gram-positivas Streptomyces y Kitasatosporia. También este aminoácido ha sido reportado como componente de solo dos tripéptidos, el bialaphos y phosalacina. El bialaphos es un tripéptido producido naturalmente por Streptomyces hygroscopicus y por Streptomyces.
(24) viridochromogenes. Mientras que la phosalacina es un tripéptido producido por Kitasatosporia (OECD, 1999). El glufosinato de amonio se considera un herbicida de contacto con un efecto sistémico vía floema, cuya acción se extiende a un amplio espectro de malas hierbas tanto anuales como perennes (Labrada et al., 1996). Este se usa después emergida la cosecha o para el control total de la vegetación en tierras no cultivadas. También es empleado para desecar las cosechas. Es absorbido por las hojas y en menor medida por los tallos verdes. No se absorbe por los tallos cuya corteza esté suberificada, ni tampoco por las raíces, la materia activa sufre una rápida biodegradación en contacto con el suelo (Labrada et al., 1996). El mecanismo primario de acción del glufosinato de amonio sobre las plantas es inhibir la enzima glutamina sintetasa (Mulwa et al., 2006). Está enzima cataliza la síntesis de la glutamina a partir de glutamato de amonio y juega un papel central en el metabolismo del nitrógeno de las plantas, principalmente en el proceso de asimilación de amoníaco en nitrógeno orgánico. Por tanto, la eficiencia del herbicida es mayor en las plantas en activo crecimiento. La inhibición de la glutamina sintetasa conduce a múltiples interferencias en el Metabolismo de las plantas y como consecuencia a estas, la muerte: • Daños de las funciones de la membrana causada por la acumulación de amoníaco (Beriault et al., 1999). • Disminución de la síntesis de proteínas, péptidos y nucleótidos debido a la falta de N orgánico para las reacciones de transaminación y transamidación. (Wendler et al., 1990) • Rápida inhibición de la fijación del CO fotosintético (dentro de 2-8 horas), 2. seguido por un daño permanente al aparato fotosintético debido a los daños en la vía foto-respiratoria. (Wendler et al., 1990).
(25) A pesar de la ausencia de los síntomas visuales, las plantas tratadas detienen su crecimiento a los primeros días después de la aplicación del glufosinato de amonio (Finalé®). El desarrollo de los síntomas visuales depende de las especies de malas hierbas y de las condiciones ambientales. Alrededor de1 a 2 días después de la aplicación, una decoloración verde pálida imperceptible o amarillenta en las hojas, comienza a observarse. Estos síntomas iniciales seguidamente se convierten en clorosis y finalmente, necrosis. La muerte completa de las malas hierbas ocurre entre la primera y segunda semana después del tratamiento. Algunos. herbicidas. inhiben. vías. metabólicas. encontradas. en. plantas. y. microorganismos. Debido a la conservación de varios sitios de unión enzimáticos cruciales en estos, los inhibidores de enzimas de plantas pueden actuar como inhibidores de sus respectivas enzimas microbianas. Esto abre la posibilidad de que herbicidas actúen como inhibidores de patógenos en algunos instantes (Pline et al., 2001). Un ejemplo que muestra como actúa el glufosinato de amonio o PPT en algunos microorganismos que poseen la enzima glutamina sintetasa, que también se encuentra en plantas, es el caso de algunas especies del género Trichoderma. Investigaciones realizadas en estos hongos mostraron que la fosfinotricina inhibía a la enzima glutamina sintetasa y que además estimulaba la actividad de otras enzimas como la aspartato aminotransferasa y la alanina aminotransferasa, así como también se detectó un aumento en los niveles de NADPH-glutamato deshidrogenada (Ahmad et al., 1995).. 2.7.2. Métodos de selección a herbicidas in vitro y ex vitro. Varios son los métodos para la selección a herbicidas en dependencia del cultivo con el cual se trabaja, ya que en base a esto será la concentración del agente selectivo en el medio de cultivo, así como el número de subcultivos que se necesitan con el herbicida para eliminar el mayor porcentaje de células o tejido no transformado..
(26) En condiciones in vitro el herbicida es fundamentalmente utilizado en la fase de multiplicación celular (agregados celulares en suspensión o callos). Respecto a su uso en la fase de regeneración de plantas existe diferencia en su uso o no. Parveez et al., (1996) señalan no realizar la selección en esta etapa pues la producción de fenoles por los explantes ante la respuesta a la presencia del herbicida en el medio de cultivo selectivo puede afectar los porcentajes de regeneración o formación de plantas. Para la fase de crecimiento de los brotes in vitro y el enraizamiento de estos es más comúnmente utilizado. Para los plátanos no se encontró en la literatura trabajos relacionados con este tema, pero sí en otros cultivos como el trigo (Triticum aestivum L.), Weeks, Anderson y Blechl 1993 y Melchiorre et al., 2002. También es importante el uso de la misma concentración del herbicida en el medio de cultivo, pues algunos autores plantean comenzar la selección con una concentración menor y después aumentar la misma. En condiciones ex vitro todos los protocolos coinciden en la aplicación del herbicida en distintas concentraciones en dependencia del cultivo que se trate. Las formas de aplicación son por rociado o asperjado y en otros casos aplicados sobre la hoja con un pincel. Todo esto empleando plantas en fase de aclimatización en macetas o bolsas. Toki et al., 1992 y Armes et al., 2004 en el cultivo del arroz (Oriza sativa L.) en casa de cultivo emplea como marcador para la selección de líneas transgénicas al herbicida Basta. En otros cultivos como caña de azúcar (Falco et al., 2000), en pasto perenne Leymus chinensis (Trin.) Shu et al., 2005, en trigo Weeks et al., 1993. 2.7.3. Cultivos transgénicos con resistencia al herbicida Finalé® Los cultivos que son mejorados genéticamente buscando tolerancia a herbicidas, seguido por los cultivos que son resistentes a insectos, representan la avanzada en la biotecnología agrícola que ha sido exitosamente explotada a escala mundial (Mayer et al., 2004). La cantidad de plantas transgénicas resistentes a herbicidas liberadas para pruebas en campos ocupa el segundo lugar (21%) en los Estados Unidos, antecedido.
(27) por las plantas resistentes a insectos que liderean las pruebas en campo hasta el 5 de abril de 2006. Ambas características han ocupado la mayoría de las veces desde 1987 más del 50% de las pruebas en campo liberadas en Estados Unidos (ISB, 2006). En el campo de la biotecnología, específicamente en la obtención de plantas transgénicas resistentes a herbicidas, se han obtenidos una gran variedad de cultivos resistentes a glufosinato y glifosato. Existen alrededor de treinta cultivos resistentes a este herbicida y se continuan los estudios. Varios de estos cultivos han sido aprobados por la Administración de Alimentos y Drogas (FDA) de los Estados Unidos para la producción comercial, entre los que se encuentran la soya, el maíz, el algodón, la remolacha y la canola (Mulwa et al., 2006). Esta gran variedad de cultivos resistentes a glufosinato refleja como se comportará en el futuro la biotecnología en este aspecto, ya que cada año el número de cultivos con estas características ha aumentado, ya no solo vale pensar en cultivos de gran importancia económica como la soya, el algodón entre otros, sino ya se obtienen otros cultivos resistentes a glufosinato no tan importantes pero sí con intereses comerciales. Estos antecedentes permiten profundizar en el estudio del empleo de marcadores de selección tipo herbicida como agente de selección en banano (Musa spp AAA)..
(28) 3. MATERIALES Y MÉTODOS El presente trabajo se realizó en el Instituto de Biotecnología de las Plantas (IBP), adscrito a la Universidad Central “Marta Abreu” de Las Villas (UCLV), Santa Clara, Cuba, en el período comprendido desde Enero 2006 hasta Diciembre 2007 Técnicas y procedimientos generales de trabajo Los medios y frascos de cultivo utilizados en este trabajo fueron esterilizados en autoclave a 121 ºC y 1.2 kg.cm-2 de presión por tiempos que variaron en dependencia del volumen de medio de cultivo a esterilizar, según información técnica de la firma SIGMA (1991). Los platos metálicos para el manejo de los explantes, los tamices para filtrado, placas de Petri y pipetas, fueron esterilizados en la estufa a 180 ºC durante dos horas. Los medios de cultivo en estado semisólido se dosificaron en frascos de vidrio con capacidad total de 250 ml, a los que se les adicionaron 30 ml y en placas de Petri, con capacidad de 10.0 y 20.0 ml. El instrumental (pinzas y espátulas) se desinfectó en una solución de hipoclorito de sodio al 1.0% (v/v) durante 15 minutos (Agramonte et al., 1993). En los trabajos con suspensiones celulares se utilizaron pipetas cortas de vidrio de 15.0 ml de volumen y una pipeta automática modelo PIPETBOY (TECNOMARA). El manejo de los cultivos en medio semisólido y de las suspensiones celulares, se realizó en condiciones de esterilidad empleando una cámara de flujo laminar horizontal. Análisis estadístico.
(29) Para el procesamiento estadístico de los datos se empleó la Prueba no paramétrica de Kruskall Wallis previa comprobación de los supuestos normalidad de los datos de los grupos homogéneos y homogeneidad de varianza. Para ello se utilizó el paquete estadístico computacional SPSS7 PC + versión 15.0 para Windows. 3.1 Determinación de la concentración mínima inhibitoria de glufosinato de amonio sobre agregados celulares embriogénicos, brotes y plantas de banano cv Gran Enano (Musa spp. AAA). 3.1.1 Determinación de la concentración mínima inhibitoria de glufosinato de amonio sobre agregados celulares, de banano cv Gran Enano (Musa sp. AAA) Este experimento tuvo como objetivo determinar la concentración mínima inhibitoria e glufosinato de amonio sobre agregados celulares embriogénicos de banano, para su uso en la selección de células o grupos de células posibles transformadas. Los. agregados. celulares. fueron. tomados. de. suspensiones. celulares. embriogénicas de ocho a diez meses después de haber sido establecidas a partir de embriogénesis somática obtenidas del cultivo in vitro de flores masculinas inmaduras, según la metodología de Gómez et al 1999. Para la multiplicación de las suspensiones se utilizó una densidad de cultivo inicial de 3.0% del volumen de células sedimentadas, siguiendo la metodología propuesta por Gómez et al. (1999) FOTO.
(30) Para el uso del glufosinato de amonio se preparó una solución a una concentración de 15 mg.ml, a partir de la cual se tomaron distintas cantidades en (ml) para establecer las concentraciones 0, 5.0, 10.0, 15.0, 20.0, 25.0 mg.l-1 empleando como fuente el herbicida comercial Finalé® (Bayer Co). La esterilización de esta solución de glufosinato de amonio se realizó por filtración con un filtro de celulosa con tamaño de poro de 0,22 µm. Se utilizaron 5 placas de Petri de 5,0 cm de diámetro por cada tratamiento que contenían 10 ml de medio de cultivo descrito por Bieberach (1995), compuesto por: Sales MS 100%, vitaminas MS, biotina 1.0 mg.l-1, extracto de malta 100 mg.l-1, 2,4-D 1.0 mg.l-1, sacarosa 45 g.l-1, el pH fue ajustado a 5.6 antes de la esterilización por autoclave y solidificado con 3 glL de Gelrite (Dúchefa. ).. Después de autoclaveado, fue añadido el glufosinato de amonio al medio de cultivo en la cabina de flujo laminar cuando este tenía una temperatura aproximada de 45ºC, sin modificar el pH del medio de cultivo Posteriormente en platos metálicos estériles se colocó papel de filtro y encima mallas de poliestireno de 1.0 cm2. En tubos cónicos de 15 mL se ajustó el porcentaje de volumen de células sedimentadas al 33% y luego, con una micropipeta (SOCOREX) de 1000 µL con puntas azules con la punta cortada, se homogenizó la suspensión y se depositaron 150. L de células (20-25 mg de. masa fresca) sobre la malla de nylon, la que retuvo las células y el papel de filtro absorbió el exceso de medio de cultivo. Las mallas con las células fueron colocadas en las placas de Petri, las cuales fueron selladas con Parafilm. (R). y.
(31) colocadas en la oscuridad a una temperatura de 27 ± 2 ºC. Este experimento se repitió dos veces en el tiempo FOTO Durante dos meses fueron realizados cada 15 días los cambios a medio fresco con el agente selectivo. Las evaluaciones se realizaron en el momento de cada subcultivo y se evaluó masa fresca inicial y final, para determinar el incremento en peso respecto al testigo y calcular el índice medio (Im) según Labrada (1999) que se describe a continuación: Im = 100 (Ii / Ic) Donde: Ii = Valor de las concentraciones del agente selectivo Ic = Valor del control Además a las 4 y 8 semanas se evaluó la mortalidad celular de los agregados celulares embriogénicos a las diferentes concentraciones del agente selectivo, de acuerdo con la escala de grados según Bermudez (2006) Escala de idalmis Se partió de diez miligramos de masa fresca de agregados celulares embriogénicos que fueron colocados sobre un porta objeto y teñidos con 100 µl de diacetato de fluoresceína (0.1 % v/v en acetona). Se realizó el conteo de los agregados celulares vivos y muertos y con el empleo del microscopio óptico OPTON (Axioskop) con una cámara digital OLYMPUS DP 70 acoplada, instalada con software para Windows XP. masa seca a las ocho semanas de cultivo-.
(32) Análisis estádist. Y dif entre las medias 3.1.2 Determinación de la concentración mínima inhibitoria sobre brotes in vitro Con el objetivo de determinar la concentración mínima inhibitoria de glufosinato de amonio sobre brotes in vitro, para su uso. como agente selectivo sobre. líneas de plantas posibles transformadas es que se realizó este experimento. Los explantes utilizados fueron brotes in vitro de banano provenientes de yemas axilares vía organogénesis en el sexto subcultivo de multiplicación. Estos brotes cultivados in vitro tenían una altura de 4.0 cm y 4.0 hojas completamente abiertas las que fueron decapitadas entre 2.0 y 5.0 mm por encima del microcormo. Los frascos de vidrio utilizados son de capacidad 250 ml, a los que se le adicionó 30 ml de medio de cultivo de multiplicación descrito por Orellana (1994), formado por sales MS, 4 mg.l-1 de 6-BAP,0.65 mg.l-1 de AIA, 30 g.l-1 de sacarosa y 1 mg.l-1 de tiamina y al que se le adicionó el glufosinato de amonio a las concentraciones de 0 ,1.0, 1.5, 2.0, 3.0, 4.0 mg.l-1 utilizando el mismo procedimiento descrito en el acápite anterior, ajustado a un pH de 5.6 --------En el medio de cultivo se colocaron cuatro plantas por frasco y cuatro repeticiones por cada concentración de glufosinato de amonio utilizada. Los frascos con los brotes fueron colocados en cámaras de crecimiento con luz solar y fotoperíodo propio de la época del año, temperatura de 25 ± 2 C y a una densidad de flujo de fotones fotosintéticas (DFFF) de 50 - 62.5 mol.m-2.s-1..
(33) A los 30 días de cultivo se evaluó la altura de las plantas (cm), número de hojas por planta, número y longitud de las raíces (cm) (las cuales aparecieron a partir de los 23 días de cultivo). Además, se confeccionó una escala para la evaluación cualitativa del material vegetal basada en la descripción de los daños provocados por el glufosinato de amonio y con el empleo de esta se compararon los diferentestratamientos para ser utilizados en la selección in vitro de líneas posibles transformadas. Estadística 3.1.3 Determinación de la concentración mínima inhibitoria de glufosinato de amonio para plantas en fase de aclimatización Este experimento se realizó con el objetivo de determinar la concentración mínima inhibitoria de glufosinato de amonio sobre plantas cultivadas in vitro en casa de cultivo para evaluar el nivel de expresión del gen de selección al herbicida que permite seleccionar plantas supuestamente transformadas Como material vegetal se utilizaron plantas en bolsas provenientes de fase de aclimatización con 60 días de crecimiento, altura promedio de 15.0 cm, diámetro del pseudotallo 1.8 cm y 6.0 hojas activas, en condiciones ambientales controladas, las que previamente fueron aclimatizadas en casa de cultivo por 45 días en bandejas de polieturano (70 alveolos) con sustrato compuesto por una mezcla de 50% de casting, 30% de compost y 20% de zeolita. El cultivo se realizó a temperatura entre 32-34 ºC durante el día y entre 22-24 ºC durante la noche y el riego se realizó manual una vez al día. El sustrato usado fue una.
(34) mezcla de 50% de casting, 30% de compost y 20% de zeolita colocado en bolsas de diámetro----. El glufosinato de amonio se aplicó con la ayuda de un pincel a todas las hojas de las plantas por ambos lados (haz y envés), a las siguientes concentraciones (05.0-10.0-20.0-30.0 y 40.0 g.l-1), Se utilizaron ocho plantas por tratamiento, La evaluación cualitativa del material vegetal basada en la descripción de los daños provocados por el glufosinato de amonio, se realizó a los 30 y 45 días. A partir de las observaciones realizadas se elaboró una escala de evaluación.. Estadística 3.2 Evaluación de la respuesta de líneas transformadas de banano cv Gran Enano con el empleo del glufosinato de amonio a partir del Herbicida Finalé® 3.2.1. Respuesta a nivel de brotes in vitro de líneas transformadas El objetivo de este experimento fue evaluar la respuesta de los brotes in vitro de líneas de banano modificadas genéticamente frente a la concentración mínima inhibitoria de glufosinato de amonio previamente determinada en el acápite 3.1.2..
(35) Como material vegetal se utilizaron brotes de líneas modificadas genéticamente que habían sido obtenidas según protocolo de transformación genética desarrollado por Bermudez (2006), donde se emplearon tres construcciones que portaban al gen bar como marcador de selección. Estas líneas habían sido evaluadas durante cuatro y cinco ciclos de cultivo en condiciones de campo (42 meses), frente a la enfermedad Sigatoca negra e introducidas al laboratorio y multiplicadas in vitro durante doce meses según metodología propuesta por Orellana (1994). Las construcciones utilizadas fueron combinaciones de proteínas antifúngicas, las cuales se describen a continuación: El plásmido pHCA58 formado por la combinación del gen de la I quitinasa (PR 3) de frijol y el gen de la proteína AP-24, una osmotina vegetal (PR 5) de tabaco; el plasmido pHCG-59 formado por la combinación del gen de la ß-1,3endoglucanasa (PR 2) de tabaco y el gen de la I quitinasa (PR 3) de frijol y el plasmido pHGA-91 formado por la combinación del gen de la ß-1,3endoglucanasa (PR 2) de tabaco y el gen de la proteína AP-24, una osmotina vegetal (PR 5) de tabaco. Por construcción se estableció el siguiente número de líneas: pHCA58 - 4 líneas (2-4-25-27) pHCG59 – 4 Líneas (1-4-16-48) pHGA91- 4 Lineas (7-17-22-23) Se emplearon 25 plantas de cada línea transformada genéticamente en la fase de multiplicación, en el octavo subcultivo, al que se adicionó el glufosinato de.
(36) amonio a la concentración obtenida en el acápite 3.1.2. Como controles se utilizaron 25 plantas del cv Gran Enano con y sin el agente selectivo. A los 30 días de cultivo se realizó la evaluación cualitativa del material vegetal basada en la descripción. de los daños provocados. por el glufosinato de. amonio, según la escala de evaluación in vitro que se confeccionó en el acápite 3.1.2. 3.2.2 Respuesta a nivel de plantas en casa de cultivo de líneas de plantas transformadas Este experimento tuvo como objetivo evaluar el efecto del glufosinato de amonio sobre líneas de plantas modificadas genéticamente de banano cv Gran enano, en casa de cultivo y comprobar la expresión del gen bar para la resistencia a este herbicida. Como material vegetal se emplearon 10 plantas por líneas de cada construcción provenientes de 10 subcultivos, las cuales fueron previamente aclimatizadas en casa de cultivo por 45 días en bandejas de polieturano (70 alveolos) con sustrato compuesto por una mezcla de 50% de casting, 30% de compost y 20% de zeolita. Posteriormente se transfirieron a bolsas de ‘’’’’’’’ de diámetro con igual sustrato por 45 días más hasta alcanzar como mínimo 20 cm de altura y al menos tres hojas activas. Transcurrido este tiempo las plantas fueron colocadas en la casa para transgénicas y se le aplicó el glufosinato de amonio con el mismo procedimiento y a la concentración determinada en el acápite 3.1.3. Como controles se emplearon 20 plantas de banano cv Gran enano provenientes de la fase de aclimatización con las características descritas.
(37) anteriormente y a diez de ellas se les aplicó el agente selectivo de la misma forma que a las plantas de los distintos tratamientos. A los 30 días se realizó la evaluación cualitativa del material vegetal basada en la descripción de los daños provocados por el glufosinato de amonio, según la escala de evaluación descrita en el presente trabajo (acápite 3.1.3).. 3.2.3. Efecto del glufosinato de amonio en fragmentos de hojas in vitro de plantas de campo Tomando en cuenta los resultados presentados por Ishida y Komari (2007) para determinar la expresión del gen bar en líneas de maíz transformadas genéticamente, se estudió la posible aplicación del protocolo desarrollado para plantas transgénicas de banano cv Gran Enano. Es por ello, que este experimento tuvo como objetivo evaluar el efecto del glufosinato de amonio en los fragmentos de hojas de plantas de líneas de campo frente a la concentración mínima inhibitoria de glufosinato de amonio determinada en el acápite 3.1.3. Los fragmentos de hojas de líneas transformadas de banano cv. Gran enano provenientes de nuevos eventos de transformación fueron tomados de plantas de las líneas GE 59-11 y 59-17, las cuales portaban la construcción -------------------------- Las mismas tenían 12 meses de cultivo en campo, en áreas del Instituto Nacional de Viandas Tropicales (INIVIT), MINAGRI, municipio de Santo Domingo y habían sido seleccionadas por su resistencia a la enfermedad Sigatoka negra en el año 2007..
(38) Se tomó la hoja 1 de la planta en cada línea supuestamente modificada genéticamente y el control sin transformar del cv Gran enano. Las hojas se trasladaron al laboratorio, se lavaron con detergente, abundante agua y en la cabina de flujo laminar se desinfectó con alcohol al 1% durante tres minutos. Posteriormente se realizaron tres enjuagues con agua deshionizada estéril. Seguidamente en platos metálicos estériles se procedió a cortar sobre papel de filtro seis fragmentos de hojas de 2.0 cm2. Estos se colocaron en placas de Petri de 10.0cm de diámetro que contenían agar-agar al 1.0 % + agua, esterilizado en autoclave y en la cabina de flujo laminar se le añadió la concentración de glufosinato de amonio determinada en el epígrafe 3.1.3 Se evaluó a las 72 horas la necrosis de los fragmentos de hojas según protocolo propuesto por Ishida y Komari (2007).. 3.2.3.1 Efecto de diferentes concentraciones de glufosinato de amonio en fragmentos de hojas in vitro de plantas de campo El experimento se desarrolló con el objetivo de evaluar el efecto de diferentes concentraciones de glufosinato de amonio sobre fragmentos de hojas de líneas transformadas genéticamente de banano cv Gran Enano provenientes de protocolo. Bermudez. (2006),. procedentes. de. campo. con. diferentes. concentraciones de glufosinato de amonio Se tomaron los fragmentos de la hoja 1 de cada línea transformada genéticamente de igual forma que en el acápite 3.2.3 a diferentes concentraciones del agente selectivo: 7.5, 15.0 y 30.0, g.l- 1.
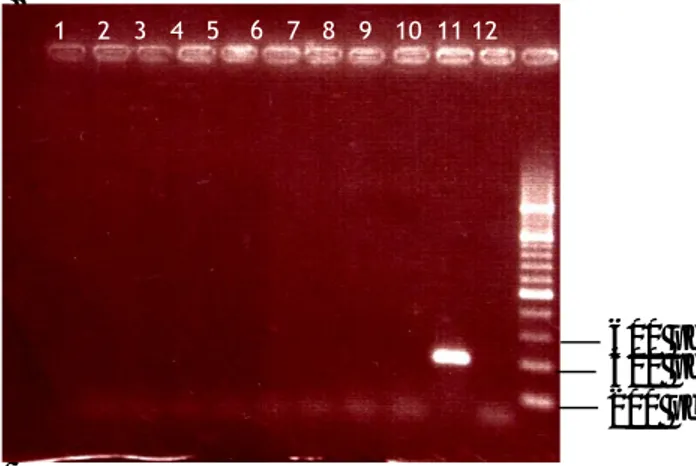
(39) Las evaluaciones se realizaron a los tres y cinco días, de forma similar a lo descrito en el acápite 3.2.3 en cada concentración de glufosinato de amonio 3.3. Análisis en Reacción en Cadena de la Polimerasa (RCP) El presente experimento tuvo como objetivo detectar las líneas que integraron en su genoma ADN proveniente de las construcciones genéticas utilizadas mediante la amplificación del marcador de selección gen bar Se partió del material recolectado en el Instituto Nacional de Viandas Tropicales (INIVIT), MINAGRI, municipio de Santo Domingo y habían sido seleccionadas por su resistencia a la enfermedad Sigatoka negra en el año 2007. (hoja cigarro) El análisis por RCP fue usado para detectar las líneas que hubieran integrado en su genoma ADN proveniente de las construcciones genéticas utilizadas, mediante la amplificación del marcador de selección gen bar. Se utilizó un termociclador personal Mastercycle Eppendorf. La mezcla de la reacción para la amplificación del ADN incluyó: 200 µm de dNTPs, 1.5 mM MgCl2, 1.0 U de Taq polimerasa en solución tampón 1X en un volumen final de 25 µl. Para reconocer la secuencia del gen bar, se usaron los siguientes oligonucleótidos: bar 5’ (5’ CGA GAC AAG CAC GGT CAA CTT C 3’) y bar 3’ (5’ GAA ACC CAC GTC ATG CCA GTT C 3’) a una concentración de 1 µM. Las reacciones se llevaron a cabo mediante 35 ciclos de: 1 min de desnaturalización a 94 ºC, 45 s de hibridación a 62 ºC, 1 min de extensión a 72 ºC y una extensión final de siete minutos a 72 ºC..
(40) Los productos de la reacción fueron analizados mediante electroforesis en gel de agarosa al 1.5% (p/v) con tampón TBE 1X (40 mM Tris-acetato, 1mM EDTA). Diez µl de cada producto se mezclaron con 1.0 µl del tampón cargador (0.25 % (p/v) de bromofenol azul, 0.25 % (p/v) de xilen cianol FF y 40 % (p/v) de sacarosa). Luego se realizó la tinción con bromuro de etidio por 10 minutos y se observó en un transiluminador MacroVue, Pharmacia LKB mediante la incidencia de radiación ultravioleta. Como control positivo se realizó una reacción adicional, con las mismas condiciones experimentales antes descritas, y se usó como ADN molde 10 ng del plasmidio pHCA58 que porta el gen bar. Como marcador de peso molecular se usó el O’GeneRuler™ 1Kb DNA Ladder, Fermentas Co. En el caso del control negativo el ADN se sustituyó por un volumen equivalente de agua y se siguieron las mismas condiciones experimentales..
(41) Resultados y Discusión. 4. RESULTADOS Y DISCUSIÓN 4.1. Determinación de la concentración mínima de glufosinato de amonio que inhibe el crecimiento en agregados celulares embriogénicos, brotes in vitro y plantas de banano cv Grande naine (Musa spp. AAA) 4.1.1. Determinación de la concentración mínima de glufosinato de amonio que inhibe el crecimiento de agregados celulares embriogénicos En las diferentes semanas de cultivo el crecimiento de los agregados celulares embriogénicos fue afectado por las diferentes concentraciones de glufosinato de amonio empleadas. Los mayores valores en el índice medio de crecimiento se alcanzaron en el tratamiento control y con las concentraciones de 5,0 y 10,0 mg.l-1 de glufosinato de amonio, sin diferencias significativas entre ellos. Mientras que, con las concentraciones de 15,0; 20,0 y 25,0 mg.l-1 del agente selectivo se produjeron los menores valores en el índice medio de crecimiento, los cuales difirieron significativamente con el control y las concentraciones de glufosinato de amonio antes mencionadas. Sin embargo, estas altas. concentraciones no mostraron. diferencias significativas entre ellas (Figura 3 A y B). Las reducciones en el crecimiento en los agregados celulares embriogénicos provocadas por las diferentes concentraciones de glufosinato de amonio se incrementaron en la medida que aumentaron las semanas de cultivo (Figura 3 A y B). A las seis y ocho semanas de cultivo se registraron los menores valores en el índice medio de crecimiento en los agregados celulares embriogénicos ya que con las concentraciones de 15,0; 20,0 y 25,0 mg.l -1 de glufosinato de amonio se produjo una reducción del crecimiento entre. 76,4 y 81,22 por ciento. Estos. resultados indican que la selección a este nivel celular debe realizarse a las ocho semanas de cultivo ya que en este tiempo se produjo mayor reducción del crecimiento celular con las concentraciones más altas de glufosinato de amonio. De forma general, a las ocho semanas. 32.
(42) Resultados y Discusión de cultivo todas las concentraciones de glufosinato de amonio estudiadas como principal afectación produjeron la necrosis del tejido celular (Figura 4).. A). Indice de crecimiento (%). 0. 100. 100a. 15. 99,98a. 20. -1 25 (mg.l ). 100 a. 96,59a. 88,88a. 78,75a. 80 60 40,68b 36,38b 33,08b. 40. 38,63b 37,99b 34,99b. 20 0 2 semanas. 0. Indice de crecimiento (%). 10. 120. A). B). 5. 4 semanas. evaluaciones. 5. 10. 15. 20. -1 25 (mg.l ). 120 100. 100a. 94,87a. 100 a 86,52a. 92,74a 78,44a. 80 60 40. 30,76b 27,81b 28,04b. 20. 23,53b 19,11b 16,98b. 0 6 semanas. evaluaciones. 8 semanas. Medias con letras distintas difieren estadísticamente según prueba de Kruskal Wallis, para P 0.05. Figura 3. Efecto de las diferentes concentraciones de glufosinato de amonio sobre el crecimiento en agregados celulares embriogénicos del cultivar de banano Grande naine (Musa spp. AAA). A) dos y cuatro semanas de cultivo y B) seis y ocho semanas de cultivo.. 33.
(43) Resultados y Discusión. A). B). C). E). F). A). D)D). Figura 4. Afectaciones provocadas en los agregados celulares embriogénicos de Grande naine a las ocho semanas de cultivo por las diferentes concentraciones de glufosinato de amonio. A) control (sin glufosinato de amonio), B) 5,0 mg.l-1, C) 10,0 mg.l-1, D) 15,0 mg.l-1, E) 20,0 mg.l-1 y F) 25,0 mg.l-1. (Bar = 1,0 cm). Por otro lado, al analizar los valores de masa seca de los agregados celulares embriogénicos a las ocho semanas de cultivo se encontró, que estos disminuyeron en la medida que se incrementaron las concentraciones del agente selectivo. Los resultados más bajos se registraron con las concentraciones de 15,0; 20,0 y 25,0 mg.l-1 las cuales mostraron una reducción de la masa seca entre 2,1 y 2,5 veces respecto a los valores alcanzados por el control. Estas tres concentraciones del agente selectivo difirieron significativamente con el control y las concentraciones de 5,0 y 10,0 mg.l -1, sin embargo estas no difirieron entre ellas (Tabla1).. Estos resultados se corresponden con los obtenidos en el índice medio de. crecimiento descritos anteriormente donde los menores valores coincidieron con las concentraciones de 15,0, 20,0 y 25,0 mg.l -1 de glufosinato de amonio. Tabla 1. Efecto de las distintas concentraciones de glufosinato de amonio sobre la masa seca (g) en agregados celulares embriogénicos del cultivar de banano Grande naine a las ocho semanas de cultivo.. 34.
(44) Resultados y Discusión. Concentración de Glufosinato de amonio (mg.l-1) 0,0 5,0 10,0 15,0 20,0 25,0 E. estándar. Masa seca (g) 0,036 b 0,034 b 0,033 b 0,017 a 0,015 a 0,014 a ±0,00203. Medias con letras distintas en una misma columna difieren según la prueba de Duncan para p 0.05.. Iguales resultados se obtuvieron en el análisis de la viabilidad de los agregados embriogénicos donde a las ocho semanas de cultivo en las concentraciones más alta del herbicida (15,0, 20,0 y 25,0 mg.l-1) se observó un 100 % de mortalidad (Figura 5). Estos resultados apoyan los anteriormente obtenidos.. A). D). B). E). C). F). Figura 5 . Efecto de las distintas concentraciones de glufosinato de amonio 1) 0, Control; 2) 5,0 mg.l-1 3) 10,0 mg.l-1;4) 15,0 mg.l-1; 5) 20,0 y 6) 25.0 mg.l-1 ,sobre la vitalidad celular a las ocho semanas de cultivo del cultivar de banano Grande Naine (Musa spp. AAA) (40 x). Bar =50 m. De acuerdo con los resultados alcanzados se seleccionó la concentración de 20,0 mg.l -1 de glufosinato de amonio como la mínima inhibitoria, ya que a las ocho semanas de cultivo, produjo una reducción en el crecimiento de más del 80 por ciento y 2,4 veces la masa seca respecto al control, además de tener una mortalidad celular del 100 %.. 35.
(45) Resultados y Discusión Estos resultados coinciden con lo informado por Sreeramanan et al. (2006), quienes emplearon en el cultivar de banano Pisang Rastali (AAB), similares concentraciones de glufosinato de amonio a las del presente trabajo (0; 5,0; 10,0; 15,0; 20,0 y 25,0 mg.l-1) y utilizaron yemas aisladas in vitro como explante vegetal para la búsqueda de un agente selectivo para la transformación genética en este cultivo. Estos autores seleccionaron como mejor concentración mínima inhibitoria 15,0; 20,0 y 25,0 mg.l-1sin diferencias significativas entre ellas. En este trabajo los autores ensayaron además concentraciones superiores de glufosinato de amonio (50,0; 100,0; 150,0; 200,0; 250,0 y 300,0 mg.l-1) en todos estos casos obtuvieron una mortalidad total de las yemas in vitro. Además estudiaron el efecto de otros cuatro posibles agentes de selección, los antibióticos (Geneticina, Kanamicina, Higromicina, Paromomicin) y concluyeron que el glufosinato de amonio a partir del herbicida Basta y la higromicina mostraron ser los mejores agentes selectivos para banano. Distintos autores informan también el uso del herbicida glufosinato de amonio como agente selectivo para la transformación genética de bananos y plátanos, pero utilizando su compuesto activo (la fosfinotricina). Busot et al. (1999) señala como la mejor concentración inhibitoria para agregados embriogénicos del cultivar de banano Grande naine (AAA) fue 6,0 mg.l -1 de fosfinotricina. Iguales resultado obtuvo al utilizar el mismo material vegetal y cultivar, Bermúdez (2006) a la concentración de 6,0 mg.l-1 de fosfinotricina, por ser ésta la mínima concentración que logró el 100 % de mortalidad de los agregados celulares. Similares resultados obtuvo Daniels (2003) al emplear suspensiones celulares embriogénicas de plátano cultivar híbrido ´FHIA-21 (AAAB). También de García et al. (2004) utiliza para la selección fosfinotricina a la concentración de 5,0 mg.l-1 usando como material vegetal meristemos del cultivar de plátano vianda Hartón (Musa spp. AAB). El uso de estas menores concentraciones para la selección están basadas en que todos los autores emplearon como agente selectivo la Fosfinotricina reactivo, en estado puro sin los aditivos que conforman la formulación de un herbicida comercial. En tejidos con metabolismo activos el efecto del herbicida glufosinato de amonio puede ser detrimental, ya que las células transformadas que puedan dar lugar a un tejido quimérico no. 36.
(46) Resultados y Discusión tienen la fuerza o el poder para continuar desarrollándose, debido a la acumulación de amonio en las células no transformadas (Sreeramanan et al., 2006). La selección de células transformadas es el factor clave en el desarrollo de un método exitoso de transformación genética. El uso de una concentración mínima puede hacer al sistema de transformación más económico teniendo en cuenta el agente selectivo para lo cual representa casi la mitad del costo de la transformación (Parveez et al., 1996). La determinación de la concentración mínima inhibitoria óptima es muy importante ya que puede hacer el proceso de selección de células transformadas más eficiente, resultando en una mayor o muy baja ocurrencia de quimeras. La concentración de fosfinotricina para el proceso de selección varía entre 1,0 y 100,0 mg.l-1, la cual depende del tejido vegetal y la composición del medio de cultivo (Enríquez, 2002). Este autor regeneró plantas transgénicas resistentes al herbicida BASTA ® (Bayer Co.) de papa y caña de azúcar con concentraciones de 2,0 y 4,0 mg.l-1 respectivamente. De acuerdo con Hadi et al. (2002) en cualquier procedimiento de transformación genética solo una pequeña fracción del tejido blanco es transformada, mientras que la mayoría resulta no transformada, por lo cual los sistemas de selección son necesariamente para identificar las células transformadas. La selección usualmente involucra el crecimiento de posibles transformantes en un compuesto químico que inhibirá el crecimiento de las células no transformadas. El más ampliamente usado como gen marcador de selección en plantas es la neomicin fosfotransferasa II (npt II) (Beck et al., (1982); Bevan et al., (1983); Herrera-E et al. (1983) el cual confiere resistencia a antibióticos como kanamicina, neomicina, G 418 y a la higromicina (Waldron et al., 1985). Existe otro gen marcador de selección que es el de la Fosfinotricina a partir del glufosinato de amonio obtenido del herbicida comercial BASTA (De Block et al., 1987). A nivel mundial se está tratando de evitar el uso de antibióticos como agentes selectivos, en los protocolos de transformación genética, ya que una de las mayores preocupaciones de los cultivos modificados genéticamente es la presencia de genes que confieren resistencia a. 37.
Figure




+7





Documento similar