Propagación in vitro de Pinus caribaea var caribaea por organogénesis
72
0
0
Texto completo
(2) y o. c u -tr a c k. .c. RESUMEN Con el objetivo de desarrollar la organogénesis de Pinus caribaea var. caribaea como una alternativa para su propagación, se realizaron experimentos para establecer in vitro brotes obtenidos de plantas donantes cultivadas en casa de cultivo, determinar el efecto del 6BAP y agentes gelificantes en la fase de multiplicación y definir la influencia de las concentraciones de sacarosa y nutrientes inorgánicos en el desarrollo de las plantas obtenidas in vitro. Los resultados demostraron que se logró establecer in vitro brotes apicales obtenidos de plantas donantes cultivadas en casa de cultivo y se definieron las características que debe tener este tipo de brote. En la fase de multiplicación, el 6-BAP influyó en el desarrollo de las plantas in vitro, con una concentración de 6,66 µM se lograron los mejores resultados en cuanto al número (6,75) y longitud de los brotes por plantas (2,70 cm), con un coeficiente de multiplicación de 2,38. Se demostró que el agente gelificante y su concentración, fueron factores que influyeron en el desarrollo in vitro de las plantas en la fase de multiplicación, con 4,0 g.L-1 de Gelrite se obtuvo el mayor coeficiente de multiplicación (2,87). Al incrementar la concentración de sacarosa a 60 g.L-1 en el último subcultivo de la fase de multiplicación, se produjeron plantas con un color verde más intenso, acículas más desarrolladas y diferenciadas, y el olor característico de los aceites esenciales que se puede percibir al macerar tejido de árboles adultos, estos cambios, le confirieron a estas plantas mejores características para ser subcultivadas a la fase de enraizamiento. Al reducir la concentración de nutrientes inorgánicos en el medio de cultivo de enraizamiento, se lograron los mayores porcentajes de residuos en la paredes celulares de los tallos. Con 50% de nutrientes inorgánicos, se logró el mayor porcentaje de residuos acumulados en la pared celular (47,95%) y con ello plantas con una mayor diferenciación celular.. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(3) y o. c u -tr a c k. .c. ÍNDICE Página 1.. INTRODUCCIÓN. 1. 2.. REVISIÓN BIBLIOGRÁFICA. 4. 2.1 Generalidades sobre el cultivo. 4. 2.1.1 Origen y distribución. 4. 2.1.2 Clasificación taxonómica. 5. 2.1.3 Características botánicas. 6. 2.1.4 Importancia económica. 7. 2.2 Propagación en forestales 2.3 Propagación in vitro 2.3.1 Regeneración de plantas por organogénesis. 10 11. 2.3.1.1 Establecimiento in vitro. 13. 2.3.1.2 Multiplicación in vitro. 14. 2.3.1.2.1 Etapa de elongación. 3.. 7. 16. 2.3.1.3 Fase de enraizamiento. 18. 2.3.1.4 Fase de aclimatización. 25. MATERIALES Y MÉTODOS. 28. 3.1 Fase de establecimiento. 28. 3.1.1 Selección del tipo de brote. 29. 3.1.2 Efecto del 6-BAP. 30. 3.2 Fase de Multiplicación. 31. 3.2.1 Efecto del 6-BAP. 32. 3.2.2 Efecto de diferentes agentes gelificantes y su concentración. 32. 3.2.3 Efecto de diferentes concentraciones de Gelrite. 33. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(4) y o. c u -tr a c k. .c. 3.2.4 Efecto de la sacarosa 3.3 Fase de enraizamiento. 4.. 33 34. 3.3.1 Efecto de diferentes concentraciones de AIB. 34. 3.3.2 Efecto de la concentración de nutrientes inorgánicos. 35. RESULTADOS Y DISCUSIÓN. 38. 4.1 Fase de establecimiento. 38. 4.1.1 Selección del tipo de brote. 38. 4.1.2 Efecto del 6-BAP. 40. 4.2 Fase de Multiplicación. 43. 4.2.1 Efecto del 6-BAP. 43. 4.2.2 Efecto de diferentes agentes gelificantes y su concentración. 45. 4.2.3 Efecto de diferentes concentraciones de Gelrite. 48. 4.2.4 Efecto de la sacarosa. 50. 4.3 Fase de enraizamiento. 53. 4.3.1 Efecto de diferentes concentraciones de AIB. 53. 4.3.2 Efecto de la concentración de nutrientes inorgánicos. 56. 5.. CONCLUSIONES. 61. 6.. RECOMENDACIONES. 62. 7.. REFERENCIAS BIBLIOGRÁFICAS. 63. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(5) y o. c u -tr a c k. .c. Introducción. _________________________________________________________________________ 1. INTRODUCCIÓN Pinus caribaea Morelet var. caribaea Barret y Golfari, es una variedad endémica de la región occidental de Cuba, específicamente de la provincia de Pinar del Río y del municipio especial Isla de la Juventud. Es la principal especie de pino plantada en el país. Sin embargo, por el declinar marcado de sus plantaciones debido a la deforestación, está clasificada como una variedad vulnerable en la lista roja publicada por la Unión Internacional para la Conservación de la Naturaleza y los Recursos Naturales (IUCN, 2010). Económicamente resulta importante para la producción de madera y resina, aunque se han desarrollado metodologías para la obtención de cera conífera, pasta clorofila-caroteno y residuo forrajero a partir del empleo del follaje después de la tala (Díaz et al., 2007). La Empresa Forestal, en Cuba, propaga el pino de manera tradicional a partir de la siembra de posturas obtenidas de semillas seleccionadas, pero este sistema no garantiza la demanda del país (Cantillo et al., 2006 a), y no siempre se logra un desarrollo homogéneo y con calidad de muchos de los árboles plantados. Una alternativa que podría contribuir a solucionar esta problemática, es el empleo del cultivo in vitro de órganos y tejidos vegetales. En Cuba, existen algunas investigaciones sobre el tema en el género Pinus (Cantillo et al., 2006 a,b), pero no se han desarrollado protocolos que se apliquen comercialmente para la propagación in vitro de ninguna de las especies de pinos sembradas en el país. A nivel mundial, la mayoría de los trabajos realizados para la propagación in vitro del género Pinus, han utilizado embriones cigóticos inmaduros y maduros como material vegetal inicial (Nehra et al., 2005; Lelu et al., 2006; Pullman y Skryabina, Tesis de Maestría. 1. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(6) y o. c u -tr a c k. .c. Introducción. _________________________________________________________________________ 2007), a partir de los cuales, se han regenerado plantas por organogénesis (Tang et al., 2004; Tang y Newton, 2005) y embriogénesis somática (Nehra et al., 2005; Vales et al., 2007). La regeneración de plantas por organogénesis, ha sido un método que ha demostrado ser eficiente en la propagación comercial de muchas especies vegetales (Zhang et al., 2006). Sin embargo, el establecimiento in vitro de brotes apicales de pino, ha sido una alternativa poco abordada. Fundamentalmente porque la respuesta in vitro del material vegetal depende en muchos casos del genotipo, la edad en campo y las características de las plantas donantes (Li et al., 2010). Por otra parte, los coeficientes de multiplicación y los porcentajes de formación de raíces in vitro que se han obtenido para muchas especies, han sido bajos (Oliveira et al., 2003; Stojicic y Budimir, 2004). Desarrollar la propagación in vitro por organogénesis a partir de plantas donantes obtenidas de semillas, permitirá mantener la diversidad biológica de P. caribaea var. caribaea fruto de muchos de años de evolución, moldeada por los procesos naturales y cada vez más, por la influencia del ser humano y multiplicar los nuevos clones de esta variedad, obtenidos mediante trabajos de selección y mejoramiento genético durante más de 40 años, los cuales presentan incrementos de hasta 30% en la producción de madera y rinden entre 20-40% más de oleorresinas que los árboles originales (Pérez, 2008, comunicación personal). Atendiendo a la problemática abordada y los antecedentes presentados, se planteó como hipótesis de trabajo la siguiente:. Tesis de Maestría. 2. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(7) y o. c u -tr a c k. .c. Introducción. _________________________________________________________________________ “Es posible a partir de brotes obtenidos de plantas donantes cultivadas en casas de cultivo, desarrollar la organogénesis de Pinus caribaea var. caribaea como una alternativa para su propagación”. Objetivos: 1. Lograr el establecimiento in vitro de brotes obtenidos de plantas donantes cultivadas en casa de cultivo. 2. Determinar el efecto de las concentraciones de 6-BAP y agentes gelificantes en la multiplicación in vitro de esta variedad. 3. Definir la influencia de las concentraciones de sacarosa y nutrientes inorgánicos en el desarrollo de las plantas obtenidas in vitro.. Tesis de Maestría. 3. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(8) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ 2. REVISIÓN BIBLIOGRÁFICA. 2.1 Generalidades sobre el cultivo 2.1.1 Origen y distribución Pinus caribaea Morelet var. caribaea Barret y Golfari, es una variedad endémica del oeste de Cuba (específicamente de la provincia de Pinar del Río y el municipio especial Isla de la Juventud), es la principal especie de pino plantada en el país, fundamentalmente sobre suelos arenosos y latosólicos (García et al., 2009). La figura 1 muestra, según García et al. (2009), ocho de las zonas de distribución natural más importantes de la variedad en la provincia de Pinar del Río.. Figura 1. Zonas de distribución natural de Pinus caribaea var. caribaea en el 2009 en la provincia de Pinar del Río.. Esta limitada y escasa área de distribución natural comprende sólo 811 ha, que también comparte con le especie Pinus tropicalis. Tesis de Maestría. 4. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(9) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ Es de destacar las afectaciones que han sufrido las áreas con poblaciones naturales de esta variedad, provocadas fundamentalmente por factores como: los incendios naturales o artificiales y el aprovechamiento maderero, convirtiéndose los incendios forestales en el principal enemigo de estas poblaciones (García et al., 2009), por lo que sería recomendable redoblar los esfuerzos en aras de conservar todo el fondo genético y poner en práctica acciones que contribuyan de manera sustancial a su conservación, por la importancia particular que tiene para la variedad continuar habitando en estos sitios. Por todo lo explicado anteriormente, a finales del 2006 la variedad fue clasificada como vulnerable en la lista roja publicada por la Unión Internacional para la Conservación de la Naturaleza y los Recursos Naturales (IUCN, 2010).. 2.1.2 Clasificación taxonómica En 1851 el naturalista francés Pierre Marie Arthur Morelet clasificó esta especie de pino como Pinus caribaea. Barrett y Golfari (1962), teniendo en cuenta el gran interés que había despertado la especie en los planes de reforestación de numerosos países del hemisferio sur, hicieron un minucioso estudio de la misma y llegaron a la conclusión de que existían suficientes razones para considerar las poblaciones geográficamente aisladas de Pinus caribaea Morelet como entidades diferentes, aunque no merezcan el rango específico. Por lo tanto, se subdividió a la especie en tres variedades: P. caribaea Morelet var. caribaea Barrett y Golfari, de Cuba (típica); P. caribaea Morelet var. hondurensis Barrett y Golfari, de Centroamérica (Pino de Honduras); y P. caribaea Morelet var. bahamensis Barrett y Golfari, de las Islas Bahamas (Pino de Bahamas). Tesis de Maestría. 5. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(10) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ Desde el punto de vista taxonómico la variedad cubana está clasificada de la siguiente forma: Reino: División: Clase: Orden: Familia: Género: Especie: Nombre binomial Nombre científico Nombre común. Plantae Tracheophyta Coniferopsida Coniferales Pinaceae Pinus caribaea Pinus caribaea Pinus caribaea Morelet var. caribaea Barrett y Golfari Pino macho. 2.1.3 Características botánicas Según Dobler y Torres (1995) y Betancourt (1999), las características botánicas de esta variedad son las siguientes: Las hojas están comúnmente en grupos de dos a tres por fascículo, raramente cuatro, de 15 a 25 cm de largo, de 0,1 a 0,13 cm de espesor, agudas, con bandas estomáticas en todas las caras, de tres a seis canales resiníferos internos, hipodermis biforme con tres a cinco hileras; vainas de 1,0 a 1,3 cm de largo, castañas a negruzcas cuando adultas. La corteza de los árboles jóvenes es grisácea, rugosa y resquebrajada en surcos más o menos profundos; en los adultos se puede mantener esta característica o bien formar placas grandes de color castaño, con fisuras poco profundas, descascarándose en finas láminas. Las características generales de la madera son: textura media, grano típicamente recto, con albura poco diferenciable del duramen. La madera recién aserrada tiene un lustre medio y es grasienta al tacto, en consonancia con la cantidad de resina. Tesis de Maestría. 6. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(11) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ que posea. Esta madera presenta anillos visibles, diferenciándose notablemente la madera tardía de la temprana. La flor es monoica. Las flores masculinas en amentos tienen de 2,0 a 3,0 cm de longitud; las flores femeninas (estróbilos) son reflejos. Las flores masculinas abundan más en las ramas bajas, las femeninas en la parte superior del árbol. Las flores femeninas se transforman en conos (frutos), que son ligeramente asimétricos, de 12 a 15 cm de longitud y entre 1,3 y 4,0 cm de diámetro; cónicos cuando están cerrados, oblongos cuando abiertos. Los conos permanecen en el árbol, si no se tumban, durante un año o más después de la diseminación de las semillas. Los frutos contienen un promedio de entre 60 y 75 semillas. Las semillas son de 0,6 cm de largo, 0,3 cm de ancho y 0,2 cm de espesor, angostamente ovoides y triangulares, de color gris moteado a pardo claro. Las semillas contienen de cuatro a ocho cotiledones.. 2.1.4 Importancia económica Económicamente resulta importante por su producción de madera y resina, aunque se han desarrollado metodologías para la obtención de cera conífera, pasta clorofila-caroteno y residuo forrajero a partir del empleo del follaje después de la tala (Díaz et al., 2007).. 2.2 Propagación en forestales La clonación o propagación vegetativa de árboles ha sido una herramienta útil en el mejoramiento forestal tradicional, y según Gronroos (1987), prometía ser la base de la revolución en lo concerniente a la producción forestal. Tesis de Maestría. 7. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(12) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ Si bien, la mejora genética clásica involucró selecciones, cruzamientos y pruebas genéticas (de familias, híbridos o clones). Según López (2000), a corto plazo con los nuevos avances en el campo de la biotecnología vegetal (organogénesis, embriogénesis somática, mapeo de genes, selección asistida por marcadores moleculares e ingeniería genética), sería posible incrementar aun más la productividad y la competitividad que demandaba el mercado regional e internacional al mejorar significativamente la homogeneidad de las plantaciones e incrementar los rendimientos. La aplicación de las técnicas de propagación vegetativa para la industria forestal es importante porque permite aumentar el número de hectáreas plantadas con material genético superior. Esto implica incrementar el rendimiento por hectárea de plantación y mejorar así la rentabilidad del proceso (Niella y Rocha, 2004). En el ámbito forestal, la mayor parte de los métodos de cultivo de tejidos han sido utilizados para producir árboles directamente para reforestación, pero estas técnicas son también un requisito básico para la producción de árboles genéticamente modificados (Yanchuk, 2001). Las técnicas de cultivo de tejidos comúnmente utilizadas en especies leñosas se basan en la regeneración de plantas por organogénesis y embriogénesis somática. Estas técnicas permiten obtener distintas respuestas, existiendo diferencias en función del genotipo, incluso dentro de una misma especie. En la propagación in vitro de pino por organogénesis, se han utilizado brotes juveniles de 0,5 a 1,0 cm de longitud que son cultivados en medios de cultivo para promover la brotación lateral a partir de yemas pre-formadas. Estos nuevos brotes obtenidos pueden continuar siendo multiplicados y enraizados para lograr la Tesis de Maestría. 8. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(13) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ formación de una planta normal (Niella y Rocha, 2004). La micropropagación tiene el potencial de multiplicar rápidamente genotipos de alto valor para la reforestación. Según Nandwani et al. (2001), estudios relacionados con la regeneración de especies coníferas a partir de brotes adventicios habían permitido obtener importantes progresos. La embriogénesis somática es una técnica que utiliza embriones inmaduros extraídos de las semillas (Lelu et al., 2006; Pullman y Skryabina, 2007) que son cultivados en medios de cultivo específicos para obtener embriones somáticos. Esto permite acelerar el proceso de multiplicación dado que no existe la etapa de enraizamiento posterior como en el caso de organogénesis o multiplicación axilar. Con la regeneración de plantas por embriogénesis somática se habilita también la posibilidad de encapsular los embriones somáticos dando lugar a las semillas artificiales, con las consecuencias. positivas. que esto significa. para el. mantenimiento, transporte del material genético, automatización y mecanización de los diferentes procesos. Los tejidos embriogénicos obtenidos pueden ser crioconservados en nitrógeno líquido a -196°C y ser guardados indefinidamente hasta su nueva utilización. Simultáneamente el material vegetal crioconservado puede ser sometido a ensayos clonales a campo. Luego de la evaluación en campo, los clones superiores pueden recuperarse de los tanques de crioconservación y ser utilizados para la producción de plántulas clonales de alto valor genético (Park, 2002; Niella y Rocha, 2004; Pullmann et al., 2005). La aplicación de estas técnicas a escala comercial está limitada a pocas coníferas, en la última década han existido avances en la mayoría de las especies forestales Tesis de Maestría. 9. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(14) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ de interés, y ya es una tecnología disponible para algunas especies del género Pinus (Klimaszewska et al., 2007). Sin embargo, los gastos de desarrollo de tales tecnologías de cultivo de tejido avanzadas son altos comparado con los de macropropagación, lo que no la convierte en una tecnología disponible para todos (Yanchuk, 2001).. 2.3 Propagación in vitro La propagación in vitro en coníferas se logrado por organogénesis y embriogénesis somática. En el caso de la organogénesis, a través de la brotación de yemas axilares y la inducción de yemas adventicias. La primera emplea ápices, yemas laterales y microestacas. La inducción de yemas adventicias se produce sobre el explante, previa formación de callo, a partir de meristemos preexistentes o tejido no meristemático. Estas yemas se originan de una o varias células cuando se cultivan con concentraciones elevadas de citoquininas. En la embriogénesis somática se pueden emplear como explante embriones cigóticos maduros e inmaduros, que en la mayoría de los casos se originan en forma indirecta a partir de callos embriogénicos, o bien, directamente desde el explante (Echenique et al., 2004). A nivel mundial, la mayoría de los trabajos realizados para la propagación in vitro del género Pinus, han utilizado embriones cigóticos inmaduros y maduros como material vegetal inicial (Nehra et al., 2005; Lelu et al., 2006; Pullman y Skryabina, 2007), a partir de los cuales, se han regenerado plantas por organogénesis (Tang et al., 2004; Tang y Newton, 2005) y embriogénesis somática (Nehra et al., 2005; Vales et al., 2007).. Tesis de Maestría. 10. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(15) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ En pino, la embriogénesis somática ha sido el método de regeneración de plantas más estudiado y desarrollado (Pullman et al., 2005; Salanova et al., 2007; Klimaszewska et al., 2007). Sin embargo, se ha descrito que en ocasiones se limita la formación del callo y se producen pocos embriones somáticos por gramo de masa fresca (Lelu et al., 2006; Maruyama et al., 2007). Estos autores explicaron, que entre otras razones, estos resultados se deben a la influencia del genotipo, la composición del medio de cultivo y al difícil control del desarrollo uniforme de todas semillas en un lote de conos. Además, para aplicar con fines comerciales la regeneración de plantas por embriogénesis somática a partir de embriones cigóticos es necesario haber desarrollado programas de mejoramiento genético a partir de pruebas de progenies, realizar selecciones genéticas y luego establecer huertos para producir las semillas de las plantas mejoradas genéticamente (Klimaszewska et al., 2007). En este sentido, en Cuba este programa aun no está disponible y se están estableciendo los huertos semilleros de segunda generación a partir de plantas de P. caribaea var. caribaea seleccionadas de las mejores combinaciones y de los individuos más sobresalientes en cada familia (Pérez, 2008, comunicación personal).. 2.3.1 Regeneración de plantas por organogénesis Los órganos como tallos, raíces y flores son inducidos a partir de una célula o grupo de células, que según las condiciones de cultivo, tienen la propiedad de mantenerse en activa división. Esta totipotencia de las células somáticas puede regenerar brotes, raíces y flores. La regeneración comprende diferentes fases: Tesis de Maestría. 11. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(16) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ adquisición de competencia, inducción, y realización (De Klerk, 2002). En la primera fase las células no responden al estímulo organogénico, pero adquieren la competencia durante la desdiferenciación; en la fase de inducción, las células son receptivas al estímulo morfogenético y hay una relación directa entre el tipo, concentración y combinación de reguladores del crecimiento adicionados al medio de cultivo y el órgano a desarrollar; y en la fase de realización la célula sufre las sucesivas divisiones (Echenique et al., 2004). La organogénesis se caracteriza por la formación de un primordio unipolar a partir de una yema con el subsecuente desarrollo de este en un brote vegetativo, existiendo siempre una conexión entre los nuevos brotes y el tejido paterno. Se reconocen cinco fases: Fase 0: Preparativa: se selecciona la planta madre y se le realizan tratamientos fitosanitarios, con el objetivo de mejorar la eficiencia en la implantación y el desarrollo posterior de los cultivos in vitro. Fase I: Establecimiento in vitro: el objetivo de esta fase es establecer cultivos asépticos y viables con los cuales iniciar el proceso de propagación. Fase II: Multiplicación in vitro: aquí se garantiza la propagación de los brotes y la estabilidad genética de las plantas producidas. Se realiza el escalado. Fase III: Enraizamiento: Puede ser in vitro o ex vitro y es donde se preparan las plantas para su transferencia al suelo. Fase IV: Aclimatización: es la fase final del proceso y por lo tanto, su meta es lograr plantas listas para el transplante definitivo a campo.. Tesis de Maestría. 12. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(17) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ 2.3.1.1 Establecimiento in vitro La propagación in vitro a partir de árboles adultos siempre ha sido difícil debido a problemas. tales. como. el. establecimiento. de. cultivos. asépticos. (severa. contaminación microbiana), la influencia de la época del año en la respuesta in vitro, o a la acumulación de determinados metabolitos secundarios que provocan fenolización (Agrawal et al., 2002). Según Korban y Sul (2007), en algunas especies de coníferas las plantas obtenidas de semillas pueden servir como plantas madres en casas de cultivo y ser fuentes de material vegetal, si se mantienen y crecen bien, ya que proporcionan gran cantidad de nuevos brotes como material vegetal para establecer in vitro. En varias especies e híbridos del género Larix, un árbol conífero que se caracteriza por su rápido crecimiento, Ewald (2007) empleó plantas de semilleros para obtener los brotes que finalmente estableció in vitro. Este autor también demostró que fue posible establecer plantas in vitro a partir de árboles adultos pero al final del invierno ya que por la pérdida de las agujas en esa época del año se facilitó y fue más efectiva la desinfección del material vegetal. Entre los reguladores del crecimiento en plantas, las citoquininas han sido las que mejores respuestas han permitido obtener durante el establecimiento in vitro de muchas especies vegetales. En árboles adultos de Holarrhena antidysenterica, Kumar et al. (2005) estudiaron el efecto de varias citoquininas en el establecimiento in vitro de los brotes y las mejores respuestas las lograron con 6BAP.. Tesis de Maestría. 13. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(18) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ 2.3.1.2 Multiplicación in vitro Según Bonga y Von Aderkas (1992), en algunas coníferas, la regeneración de plantas por organogénesis a partir de yemas axilares, fue eficaz. En años más recientes se ha podido desarrollar este proceso incluso a partir de árboles adultos (De Diego et al., 2008), pero en general, a lo largo de los años se ha observado que no ha ocurrido así con la mayoría de las especies coníferas y en la práctica, con el empleo de este método de regeneración, no se ha podido llegar a una etapa de aplicación (Bonga et al., 2010). La regeneración de plantas por organogénesis, ha sido un método que ha demostrado ser eficiente en la propagación comercial de muchas especies vegetales. Sin embargo, en Cuba, con respecto al género Pinus existen pocas investigaciones sobre el tema (Cantillo et al., 2006 a,b) y no se han desarrollado protocolos que se apliquen comercialmente para la propagación in vitro de ninguna de las especies de pino plantadas en el país. Esto se debe fundamentalmente, a que la respuesta in vitro del material vegetal durante el proceso de propagación in vitro, depende en muchos casos del genotipo, de la edad en campo y las características de las plantas donantes, y a que los coeficientes de multiplicación y los porcentajes de formación de raíces in vitro que se han obtenidos para muchas especies, han sido bajos. Existen numerosas investigaciones que refieren la regeneración de plantas de pino por organogénesis, pero a partir de embriones cigóticos (Tang et al., 2004; Tang y Newton, 2005; Alonso et al., 2006). Sin embargo, el empleo de explantes tales como brotes apicales, yemas axilares, acículas, etc., para el establecimiento in vitro de plantas de pino han sido alternativas menos estudiadas. Por lo tanto, Tesis de Maestría. 14. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(19) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ pocos trabajos describen resultados en la fase de multiplicación a partir de haber establecido in vitro estos tipos de explantes. Las citoquininas son reguladores del crecimiento muy eficaces para estimular la iniciación directa o indirecta de brotes in vitro (van Staden et al., 2008). Según estos autores, sus efectos en el cultivo de tejidos y órganos pueden variar según el tipo de citoquinina, la concentración empleada en el medio de cultivo, si el material vegetal a establecer in vitro se obtiene de tejido juvenil o tejido adulto, puede variar además en función de la especie vegetal y del método de regeneración de plantas empleado. Según van Staden et al. (2008), cuando se utilizan altas concentraciones de citoquininas en la fase de multiplicación, los brotes que se producen reducen su desarrollo en longitud. Autores como Kumar et al. (2005), estudiaron el efecto de varias citoquininas en la multiplicación in vitro de brotes apicales obtenidos de árboles adultos de Holarrhena antidisentérica y las mejores respuestas las lograron con 6-BAP. Estos autores, al evaluar el efecto de diferentes concentraciones de 6-BAP, comprobaron que al igual que en muchas otras especies, las respuestas in vitro para la mayoría de las variables evaluadas dependieron de las concentraciones estudiadas. Una determinada concentración de Agar puede ser considerada inadecuada si no ayuda al desarrollo in vitro de los brotes o si favorece la aparición de plantas con síntomas de hiperhidricidad. (Thorpe et al., 2008). Estos autores plantean que la hiperhidricidad se puede reducir si se incrementa la concentración de Agar en el. Tesis de Maestría. 15. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(20) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ medio de cultivo, pero este incremento, regularmente está acompañado de una disminución en el desarrollo y crecimiento de las plantas in vitro. No obstante, se ha demostrado que existen diferencias en la composición y características de cada uno de estos compuesto, por ejemplo, el Gelrite como producto comercial está libre de impurezas orgánicas que si se encuentran en el Agar (Thorpe et al., 2008) y tiene entre otras ventajas para la propagación in vitro de plantas a gran escala, que su costo por litro es menor al del Agar y se emplea en menor concentración que este, además, produce un gel más claro que el Agar y con ello permite detectar más fácil cualquier contaminación microbiana.. 2.3.1.2.1 Etapa de elongación Muchos estudios en la propagación in vitro por organogénesis en el género Pinus mencionan el desarrollo de esta etapa dentro del proceso y es que para lograr la iniciación y crecimiento de brotes vigorosos para enraizar y aclimatizar existen tres pasos necesarios: a) Estimular de la división celular. b) Desarrollar los brotes. c) Lograr el crecimiento de los brotes. Esta tricotomía es necesaria mencionarla dado que los tratamientos que estimulan la división celular son antagonistas con los que provocan el crecimiento y desarrollo. Lo mismo sucede durante el enraizamiento; aquí también los tratamientos que estimulan la división celular en la base de los tallos son opuestos a los que producen el crecimiento de las raíces in vitro (Amerson et al., 1982). Existen varios factores que tienen un papel determinante en esta etapa: Tesis de Maestría. 16. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(21) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ •. Tipo de explante inicial. Con respecto al tamaño de brote que se utiliza para iniciar la etapa de elongación, Coke (1996); Frampton et al. (1998) y Mathur et al. (2001) coinciden en que es necesario partir de brotes que tengan un tamaño mayor a 0,5 cm. Otros autores en trabajos más recientes sugieren utilizar brotes mayores de 2,0 cm para iniciar la etapa de elongación (Prehn et al., 2003; Zhang et al., 2006); y esto permitirá la obtención de brotes con mayor desarrollo en longitud para enraizar, en menor tiempo. Mathur et al. (2001) explican que cultivar grupos de brotes en lugar de brotes individuales actúa en detrimento del desarrollo en longitud de los brotes. •. Tipos y concentración de nutrientes inorgánicos. Según Gamborg et al. (1976) el medio de cultivo basal es uno de los factores más importantes que determinan el éxito del cultivo in vitro. Muchos de los medios de cultivo utilizados en Pinus spp., han sido creados para otras especies, incluso angiospermas, como el MS (Murashige y Skoog, 1962), el SH (Schenk y Hildebrandt, 1972), el DCR (Gupta y Durzan, 1985) y el LP (Aitken Christie et al., 1986). También se han utilizado esos medios de cultivo basales con distintas concentraciones de nutrientes; y a partir de experimentos se han seleccionado los medios de cultivo que mejores respuestas han dado. En la actualidad, uno de los medios de cultivo basales más utilizado en pino es el Westvaco (WV5) descrito por Coke (1996), con buenos resultados en el cultivo de tejidos in vitro, con una mejor combinación de nutrientes inorgánicos, apropiada fuente de carbohidratos, reguladores de crecimiento y carbón activado.. Tesis de Maestría. 17. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(22) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ •. Reguladores del crecimiento. No es común encontrarlos en el medio de elongación; sin embargo, Tang et al. (1998) suplementa el medio de cultivo con 0.5 mg.L-1 de ácido 3-indolbutírico (AIB) y 1 mg.L-1 de ácido giberélico (AG3) obteniendo en seis semanas brotes de Pinus taeda de 1,0 cm para enraizar in vitro. Con estos brotes obtuvo el 46% de enraizamiento. La adición de auxinas al medio de elongación podría generar un brote con mayor potencial de enraizamiento en la siguiente fase por inducir la formación de raíces adventicias; además junto con el AG3 podría favorecer la elongación de brotes ya que actúan en el alargamiento celular.. 2.3.1.3 Fase de enraizamiento •. Enraizamiento in vitro. El enraizamiento de brotes de gimnospermas en condiciones in vitro, es ampliamente reconocido como difícil de lograr (Nandwani et al., 2001; Schestibratov et al., 2003; Parasharami et al., 2003; Prehn et al., 2003; Zhang et al., 2006). Es un proceso lento; debido a que los brotes de la mayoría de las coníferas, especialmente Pinus spp. enraízan a partir de un callo o tejido cicatricial o directamente del tejido vascular, después de un tratamiento con auxinas. El enraizamiento directo pocas veces se produce espontáneamente (Stojicic y Budimir, 2004). Las raíces se generan siempre a partir de tejido meristemático, ya sea por fuera de los conductos resiníferos en el enraizamiento directo, o por el tejido meristemático que rodea el callo, en el indirecto. El cultivo in vitro no impide la iniciación radicular en coníferas; los tratamientos con auxinas usualmente tienen. Tesis de Maestría. 18. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(23) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ un efecto positivo en el enraizamiento; es más rápido, más sincronizado y resulta en más raíces por brote; pero la elongación radicular es severamente inhibida. Algunos autores afirman que el desarrollo de raíces in vitro usualmente aumenta la supervivencia al transplante dado que las raíces funcionales generan un balance hídrico favorable. Estas raíces compensarían la pérdida de agua causada por estomas no funcionales, y facilitarían la absorción de nutrientes. Una buena respuesta y un aumento en la masa seca de estas plantas enraizadas in vitro pueden favorecer una mejor toma de nutrientes. Sin embargo la presencia de raíces durante el cultivo in vitro no siempre mejoraría el éxito al trasplante (Seelye et al., 2003). •. Enraizamiento ex vitro. Un rápido enraizamiento mejoraría la calidad de las plantas, incluso es probable que las plantas con un buen sistema radicular y una vigorosa conexión vascular tengan una mayor tasa de crecimiento inicial en la fase de aclimatización. La mayor parte de la bibliografía consultada cita el enraizamiento ex vitro de estacas de Pinus taeda vía macropropagación, con distintos porcentajes de éxito en función de la técnica empleada. Todos los trabajos mencionan la aplicación de un tratamiento inductivo corto con auxinas en distintas concentraciones. Es importante mencionar que entre los factores que podrían afectar el enraizamiento se encuentran los siguientes: •. Tipo y concentración de la auxina. Durante estudios realizados para la formación de raíces in vitro, se demostró que es fuertemente dependiente del tipo de auxina utilizada y su concentración. Varias combinaciones, concentraciones y duración de tratamientos con auxinas han sido Tesis de Maestría. 19. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(24) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ probadas con diferentes porcentajes de éxito. Las auxinas más utilizadas son el AIB y el ANA para las distintas especies del género Pinus spp. (Kalia et al., 2007). En los trabajos que utilizan ANA, las concentraciones oscilan desde 0,1 a 20 mg.L-1 desde minutos hasta varias horas (Nandwani et al., 2001) o tratamientos continuos con concentraciones desde 0,01 mg.L-1 a 0,5 mg.L-1 de ANA pero generalmente combinado con AIB en concentraciones de 1,0 a 20 mg.L-1 (Schestibratov et al., 2003; Prehn et al., 2003), aunque son numerosos los trabajos que utilizan solamente AIB para obtener mejores resultados en concentraciones que varían de 0,1 a 1000 mg.L-1 (Tang y Ouyang, 2000; Parasharami et al., 2003; Zhang et al., 2006). El ácido 3-indolacético (AIA) es la auxina menos potente siendo necesaria su adición en concentraciones superiores a 5,0 mg.L-1, debiéndose su baja actividad a la degradación que la luz produce de la misma. Los resultados son variables, y mayormente se refieren a enraizamiento in vitro con pulsos de auxinas seguidos de un periodo de elongación de raíces in vitro sin reguladores, logrando porcentajes de supervivencia entre 0,0 y 90% en función del tratamiento y la especie. Se observa en muchos de estos trabajos que los porcentajes de enraizamiento pueden ser muy bajos o casi nulos si no se encuentra la concentración óptima del regulador de crecimiento para la especie o variedad con que se trabaja. •. Formas de aplicación de la auxina. Al comparar tratamientos de enraizamiento por pulsos o continuos, debe medirse no solo el porcentaje de enraizamiento sino también el tiempo requerido para enraizar. Los resultados con exposición continua varían entre 20% y 47% de Tesis de Maestría. 20. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(25) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ enraizamiento en 12 semanas en un medio de cultivo con la mitad de los nutrientes inorgánicos GD o DCR con 0,1 mg.L-1 de 6-BAP y 0,1 mg.L-1 de ANA o 2,0 mg.L-1 de AIB. En contraste, un tratamiento en forma de pulso con el mismo medio de cultivo con 0,1 mg.L-1 de 6-BAP y 0,5 mg.L-1 de ANA durante seis a trece días hasta el subcultivo en un medio de cultivo sin reguladores del crecimiento, produjo un 85% de enraizamiento. El enraizamiento posterior a un pulso se completa en cinco o seis semanas, por lo que el proceso completo requiere entre seis y ocho semanas. En general los tratamientos por pulso brindan un mayor porcentaje de enraizamiento en un período más corto de tiempo (Mathur et al., 2001). Tratamientos cortos (10-30 minutos) con altas concentraciones de AIB (200 mg.L-1) han sido suficientes para inducir la formación de raíces. La respuesta declina rápidamente con un tiempo de exposición más prolongado a la mencionada concentración, causando daños a los tejidos e inhibiendo el enraizamiento. Una incubación prolongada en un medio de cultivo rico en auxinas puede causar una callosidad excesiva, iniciación radicular retardada o una inhibición de la elongación de las raíces iniciadas (Selby et al., 1991). •. Número de subcultivos. Se ha demostrado que la formación de raíces puede declinar con los subcultivos sucesivos que han sufrido los brotes antes de su enraizamiento. Sin embargo, la formación de raíces adventicias sería mas difícil de lograr de tejidos maduros que de tejidos juveniles; y las condiciones in vitro probadas para diferentes especies leñosas serían mas favorables para enraizar brotes maduros que las condiciones ex vitro. Esto se debe a la posibilidad de rejuvenecer el tejido maduro a través de sucesivos. subcultivos. Tesis de Maestría. en. medios. de. cultivo. apropiados,. que. inducen 21. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(26) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ progresivamente a un aumento del potencial de formación de raíces adventicias (Parasharami et al., 2003). •. Tipo de brote. Manteniendo todos los parámetros iguales, el tamaño del brote tiene una marcada influencia en el enraizamiento. Los brotes mas largos enraízan mejor que los brotes cortos, por lo que se recomiendan los mayores a 1,5 cm para enraizar. Sin embargo, Coke (1996) demuestra que para realizar el enraizamiento ex vitro, son necesarios brotes de longitud mayor a 4,0 cm, es decir, de mayor tamaño que el requerido para enraizar en laboratorio. •. Concentración de las sales. Las etapas de inducción y crecimiento radicular se realizan, según la bibliografía, en medios de cultivo con la concentración de nutrientes inorgánicos reducidos a la mitad y cuando las raíces alcanzan los 0,5 cm de longitud ya están preparadas para su trasplante a la fase de aclimatización (Mathur et al., 2001; Nandwani et al., 2001; Parasharami et al., 2003). •. Influencia de la sacarosa. En las plantas, los carbohidratos tienen varias funciones esenciales. Ellos constituyen sustratos para la respiración, juegan un importante papel en la vía de síntesis de muchos compuestos, son elementos básicos de las macromoléculas y controlan además, otros muchos procesos relacionados con el desarrollo de las plantas (Gibson, 2000; Smeekens, 2000). La sacarosa probablemente ha sido la fuente de carbohidratos más utilizada en el cultivo in vitro de tejidos vegetales y numerosos estudios la han señalado como la. Tesis de Maestría. 22. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(27) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ fuente de carbono óptima (Alkhateeb, 2001). No obstante, no se debe olvidar que existen enzimas invertasas que son liberadas al medio de cultivo por las plantas cultivadas in vitro y que actúan en la hidrólisis de la sacarosa dando lugar a la glucosa y la fructosa (Thorpe et al., 2008), con lo cual, es importante tener en cuenta que las plantas in vitro dispondrán para su desarrollo no sólo de la sacarosa, sino también de sus dos monosacáridos constituyentes. Se conoce que los azúcares intervienen en diferentes procesos morfogenéticos, una de las funciones más interesante se ha descrito en el desarrollo de las semillas (Calamar y De Klerk, 2002). Otros estudios han demostrado que la glucosa se ha asociado con la división celular y la sacarosa con la acumulación de sustancias de reservas (Weber et al., 1998) y la inducción de la floración (Hong et al., 2006). La capacidad de las plantas para metabolizar los diferentes tipos de carbohidratos es diferente (Alkhateeb, 2008). Se ha descrito por algunos investigadores, que la respuesta in vitro de los cultivos a diferentes tipos y concentraciones de carbohidratos parece ser, en cierta medida, genotipo dependiente (Cuenca y Vieitez, 2000) y se han realizado estudios para definir estas posibles dependencias. Se conoce que las altas concentraciones de sacarosa (>6.0%) en el medio de cultivo han sido capaz de reducir la capacidad fotosintética de las plantas (Arigita et al., 2002). También se ha descrito que estas altas concentraciones pueden reprimir la expresión de genes y reducir el contenido de clorofila, afectar el Ciclo de Calvin, así como reducir la actividad Rubisco y la concentración de Rubisco, lo que conlleva a bajas tasas fotosintéticas (Premkumar et al., 2002; Sinha et al., 2002).. Tesis de Maestría. 23. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(28) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ Diferentes estudios han demostrado que existen efectos opuestos al aplicar sacarosa al medio de cultivo ya que en algunos casos hay una respuesta favorable y en otros se produce una inhibición del crecimiento (Van et al., 2001). A pesar de que se ha examinado el efecto regulador de los azúcares, en particular, el papel de la sacarosa en el desarrollo de la latencia, en la formación de órganos de almacenamiento y la maduración de embriones somáticos, su papel como molécula reguladora aun no ha sido totalmente dilucidado (Calamar y De Klerk, 2002). Con relación a la acción de la sacarosa en la formación de órganos adventicios se han realizado pocos estudios (Calamar y De Klerk, 2002). Según Warren et al. (1994) la sacarosa incrementó la regeneración de tejido vascular en Lactuca sativa (Lechuga), mientras que, en Malus domestica (Manzano), Pawlicki y Welander, (1995) demostraron que el tipo de azúcar influyó en la formación de raíces. Varios han sido los estudios encaminados a comprender mejor la respuesta de diferentes especies coníferas para formar raíces in vitro. La adición al medio de cultivo de auxinas, por lo general, en forma de AIB o ANA, ha sido una práctica general para inducir la formación de raíces in vitro en coníferas (Niemi et al., 2002). Sin embargo, se conoce que el éxito final de esta fase del proceso de propagación in vitro, no solo depende de la adición al medio de cultivo de determinadas concentraciones de auxinas. Por ejemplo, es importante tener en cuenta el efecto que pueden ejercer las altas concentraciones de citoquininas (0.5-10 mg.L-1) empleadas durante la fase de multiplicación, pues pueden inhibir o retrazar la formación de raíces y también limitar los efectos estimuladores de las auxinas en esta fase del proceso (Ben-Jaacov et al., 1991). Tesis de Maestría. 24. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(29) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ En ocasiones, ha sido necesario realizar más de un subcultivo en medio de cultivo libre de citoquininas, hasta que las concentraciones endógenas de este regulador del crecimiento se han reducido lo suficiente en el tejido de la planta (van Staden et al., 2008). Otro elemento que se ha demostrado que es importante evaluar, está relacionado con la influencia de la concentración de sacarosa, pues influye en la calidad funcional de las plantas cuando estas son transferidas a condiciones ex vitro para su aclimatización (Fuentes et al., 2005). Se conoce que los carbohidratos juegan un papel importante en el cultivo in vitro como fuentes de energía y carbono, así como agentes osmóticos, pero también, están vinculados con la diferenciación de los elementos del xilema y floema (Thorpe et al., 2008). Ha sido descrito que al aumentar la concentración de sacarosa en el medio de cultivo se incrementa el porcentaje de materia seca en las plantas (Kubota et al., 2002; Shim et al., 2003). Lo anterior se debe, a que al aumentar el contenido de sacarosa, el potencial osmótico del medio de cultivo disminuye (Cárdenas y Villegas, 2002), se limita la absorción de agua, pero se favorece la asimilación de sacarosa y con ello el incremento de materia seca. Este incremento en el porcentaje de materia seca, está asociado al desarrollo de los tejidos vasculares de las plantas, en particular con el proceso de lignificación.. 2.3.1.4 Fase de aclimatización Manipulando el ambiente in vitro las hojas, con mayor resistencia al estrés hídrico y competentes fotosintéticos, pueden desarrollarse en el laboratorio, en una Tesis de Maestría. 25. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(30) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ preaclimatización, preparando las plantas para la transferencia fuera del laboratorio. Las raíces formadas in vitro pueden favorecer el crecimiento inicial ex vitro; aunque la tasa de crecimiento óptima no ocurrirá hasta que las nuevas hojas y raíces se desarrollen en el ambiente ex vitro (Seelye et al., 2003). La aclimatización induce cambios morfológicos, anatómicos y fisiológicos que hacen a la planta parecerse a una planta normal. Esta etapa es casi siempre estresante y suele asociarse a la muerte de las plantas, generalmente por fuerte deshidratación. En angiospermas la principal causa de muerte al transferir las plantas a condiciones no estériles es la cutícula reducida y una pobre regulación estomática ante la perdida de agua. Sin embargo, en algunas gimnospermas como el género Pinus la regulación cuticular y estomática ante la pérdida de agua es relativamente normal. De esto se desprende que son otros los factores que producen la muerte de las gimnospermas durante la fase de aclimatización. Se le ha prestado atención a la morfología, anatomía interna y fisiología de las plantas obtenidas in vitro, concluyendo que para lograr el éxito en la supervivencia y el crecimiento hay características a tener en cuenta, como la longitud del brote; la calidad de los brotes y la morfología del sistema radicular. Lo interesante es determinar la morfología de plántula óptima y su relación con la supervivencia y crecimiento. Para Pinus taeda, la calidad del brote es mucho más importante en la supervivencia, que el número de raíces. En P. caribaea var. hondurensis x P. tecunumanii, Haines et al. (2000), encontraron que porcentaje de enraizamiento y número de raíces por planta estuvo relacionado con el genotipo; mientras que, dentro de los parámetros morfológicos, la longitud de acículas primarias fue el Tesis de Maestría. 26. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(31) y o. c u -tr a c k. .c. Revisión Bibliográfica. _________________________________________________________________________ mejor indicador. Estos autores concluyeron, que los brotes con mayor enraizamiento, fueron aquellos que presentaron acículas primarias mayores a 2,5 cm de longitud, con el ápice activo y un diámetro basal mayor a 0,1 cm. Existe poca información en relación al crecimiento en suelo de plántulas de coníferas originadas in vitro, dado que muy pocas especies tuvieron éxito en este sentido.. Tesis de Maestría. 27. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(32) y o. c u -tr a c k. .c. Materiales y Métodos ________________________________________________________________________________________. 3. MATERIALES Y MÉTODOS. El presente trabajo se desarrolló en el Instituto de Biotecnología de las Plantas de la Universidad Central “Marta Abreu” de Las Villas entre los años 2008 y 2010.. 3.1 Fase de establecimiento Condiciones generales Brotes de P. caribaea var. caribaea con aproximadamente 3,0 cm de longitud fueron cortados en horas tempranas de la mañana y se colocaron en 200 mL de una solución estéril de ácido cítrico con una concentración de 500 mg.L-1. En el laboratorio, los brotes fueron lavados con detergente comercial (2,0 g.L-1) y agua común, el enjuague se realizó también con una solución de ácido cítrico similar a la descrita anteriormente. Como agente desinfectante se empleó el Hipoclorito de sodio al 2,0% y los brotes se expusieron a su acción durante 20 minutos en un agitador orbital a una velocidad constante de 180 rpm. Trascurrido este tiempo, se realizaron en una cabina de flujo laminar tres enjuagues con la solución de ácido cítrico y antes de ser colocados los brotes en contacto con el medio de cultivo en los tubos de ensayo, se eliminó la parte de la base que se afectó por la acción del Hipoclorito de sodio. El medio de cultivo basal estuvo compuesto por los nutrientes inorgánicos propuestos por Coke (1996), los cuales se conocen comercialmente como Westvaco (WV5) según la presentación comercial del catalogo Duchefa (2008), este medio de cultivo también incluyó en su formulación inicial 1,0 g.L-1 de mioinositol y 0,4 mg.L-1 de tiamina y le fueron adicionados 0,6 mg.L-1 de tiamina para Tesis de Maestría. 28. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(33) y o. c u -tr a c k. .c. Materiales y Métodos ________________________________________________________________________________________. completar a 1,0 mg.L-1 la concentración de este compuesto, 1,0 g.L-1 de Lglutamina, 3,0 g.L-1 de carbón activado, 20 g.L-1 de sacarosa y 2,5 g.L-1 de Gelrite con pH ajustado a 5,8 antes del proceso de esterilización. Se dosificaron 10 mL de medio de cultivo por tubo de ensayo y se esterilizaron durante 15 minutos en autoclave a 1,2 kg.cm-2 de presión y 121°C. Los experimentos fueron repetidos tres veces en el tiempo. La temperatura de la cámara de crecimiento fue de 28 ± 2,0°C, con una densidad de flujo de fotones fotosintéticos que osciló entre 38 y 47,5 µmol.m-2.s-1. Para el procesamiento estadístico de todos los resultados se empleó el Statistical Package for the Social Sciences (SPSS) para Windows versión 18.0. En cada experimento se especificó el tipo de análisis y las pruebas aplicadas.. 3.1.1 Selección del tipo de brote Con el objetivo de lograr el establecimiento in vitro de P. caribaea var. caribaea, se evaluó la respuesta in vitro de cuatro tipos de brotes que fueron cortados de las plantas donantes con diferentes tiempos (días) de brotados, los cuales presentaron diferentes características morfológicas (Figura 2). El brote tipo (A) se caracterizó por tener un color verde más intenso, mayor número de acículas y una menor distancia de inserción al tallo entre las acículas (Figura 2). Los brotes tipo (A1) y (A2) tenían el mismo origen que el tipo (A), pero con 28 y 35 días de brotados en la planta donante, mientras que, el brote tipo (B) en comparación con los anteriores, presentó un color verde menos intenso, un menor. Tesis de Maestría. 29. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(34) y o. c u -tr a c k. .c. Materiales y Métodos ________________________________________________________________________________________. número de acículas en igual longitud del tallo y por consiguiente una mayor separación entre las acículas.. (A) 20 días. (A1) 28 días. (A2) 35 días. (B) 20 días. Figura 2. Características morfológicas de los diferentes tipos de brotes de P. caribaea var. caribaea utilizados como material vegetal de partida para el establecimiento in vitro de esta variedad.. En este experimento, se colocaron 30 brotes por tratamiento y al medio de cultivo basal descrito anteriormente se le adicionaron 4,44 µM de 6-BAP, a los 30 días de cultivo se evaluó, el número de brotes vivos y el número de brotes sin afectaciones por necrosis. Por no cumplirse los supuestos de normalidad y homogeneidad de varianza de los datos se aplicó de Mann Whitney.. 3.1.2 Efecto del 6-BAP Este experimento tuvo como objetivo evaluar la influencia de diferentes concentraciones de 6-BAP en la respuesta in vitro de los brotes durante la fase de establecimiento. Los tratamientos fueron 0,0; 2,22; 4,44; 6,66 y 8,88 µM de 6-BAP y se colocaron un total de 30 brotes por tratamiento. Tesis de Maestría. 30. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
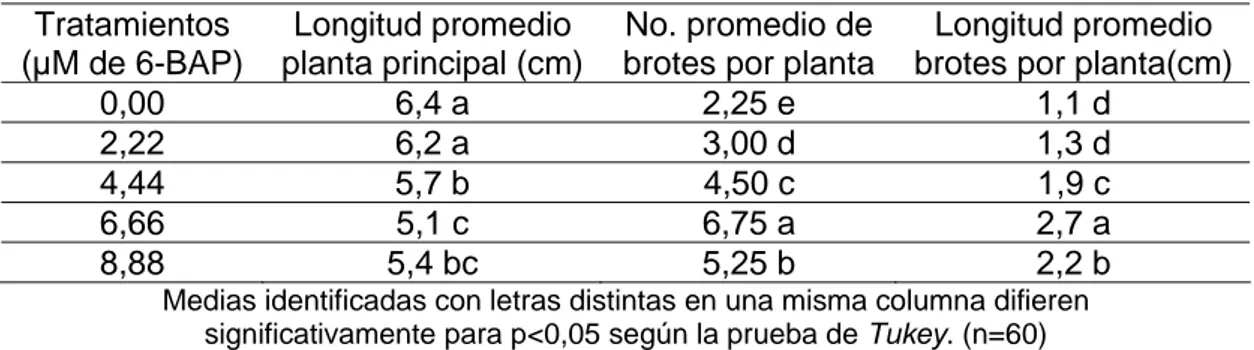
(35) y o. c u -tr a c k. .c. Materiales y Métodos ________________________________________________________________________________________. Las evaluaciones se realizaron a los 30 días de cultivo y se determinó la longitud de los brotes (cm) y el número de brotes desarrollados. Como brotes desarrollados se consideraron aquellos que presentaron cambio en su longitud, engrosaron o desarrollaron sus yemas axilares. Para la comparación de la longitud media de los brotes se realizó un análisis de varianza simple y se empleó la prueba de Tukey. El número de brotes desarrollados se expresó en porcentaje y en este caso, para el procesamiento estadístico de los resultados por no existir normalidad de los datos se empleó la prueba de Kruskal Wallis y la comparación entre parejas de grupo se realizó con prueba de Student Newman Keuls (SNK).. 3.2 Fase de Multiplicación Condiciones generales Como material vegetal para iniciar los estudios en esta fase del proceso, se emplearon plantas con 30 días de establecidas in vitro. El medio de cultivo basal fue similar al descrito para la fase de establecimiento, pero con 30 g.L-1 de sacarosa, en todos los tratamientos se colocaron 60 plantas y se consideró cada planta como una réplica. Se dosificaron 30 mL de medio de cultivo por frasco y se esterilizaron durante 20 minutos en autoclave a 1,2 kg.cm-2 de presión y 121°C. Los experimentos fueron repetidos tres veces en el tiempo, se colocaron tres plantas por frasco de cultivo y la temperatura de la cámara de crecimiento fue de 28 ± 2,0°C, con una densidad de flujo de fotones fotosintéticos que osciló entre 38 y 47,5 µmol.m-2.s-1.. Tesis de Maestría. 31. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
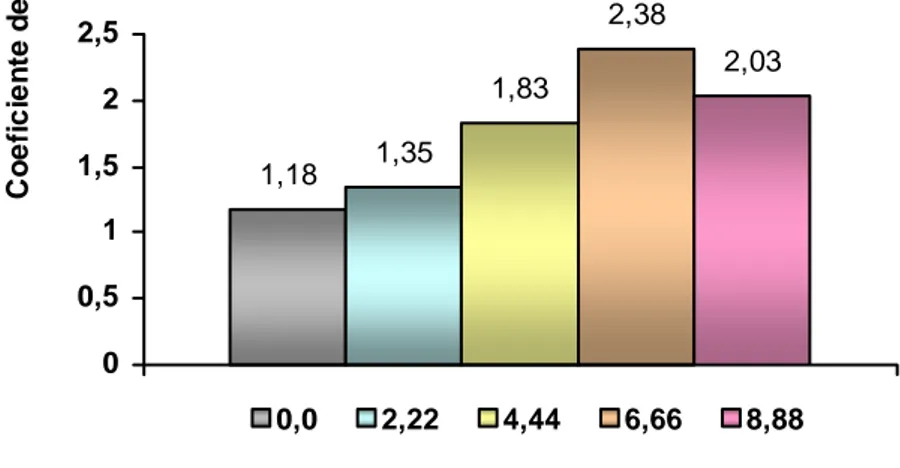
(36) y o. c u -tr a c k. .c. Materiales y Métodos ________________________________________________________________________________________. 3.2.1 Efecto del 6-BAP Este experimento tuvo como objetivo determinar la influencia de diferentes concentraciones de 6-BAP en la multiplicación in vitro de las plantas. Los tratamientos fueron 0,0; 2,22; 4,44; 6,66 y 8,88 µM de 6-BAP. Las evaluaciones se realizaron a los 35 días de cultivo y se determinó en cada tratamiento, el número de brotes por planta, la longitud de los brotes (cm), la longitud de la planta principal (cm) y se determinó el coeficiente de multiplicación. Para la comparación de las medias se realizó un análisis de varianza simple y se empleó la prueba de Tukey.. 3.2.2 Efecto de diferentes agentes gelificantes y su concentración Con el objetivo de evaluar la incidencia de dos agentes gelificantes y diferentes concentraciones de estos en el desarrollo y respuesta in vitro de plantas de pino, se estudiaron tres concentraciones de Gelrite (Duchefa Biochemie) (2,5; 3,0 y 3,5 g.L-1) y tres de Agar E (BIOCEN) (4,3; 4,8 y 5,3 g.L-1). El medio de cultivo fue similar al utilizado en el acápite anterior y a los 35 días de cultivo se evaluó en cada tratamiento, el número de plantas con necrosis total y parcial y se determinó el coeficiente de multiplicación. Por no existir normalidad de los datos en el caso del número de plantas necrosadas, se empleó la prueba de Kruskal Wallis. La comparación entre parejas de grupo, se realizó con la prueba de Student Newman Keuls (SNK).. Tesis de Maestría. 32. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(37) y o. c u -tr a c k. .c. Materiales y Métodos ________________________________________________________________________________________. 3.2.3 Efecto de diferentes concentraciones de Gelrite Teniendo en cuenta que el costo por litro del Gelrite es menor al del Agar, que se emplea en menor concentración y que además produce un gel más claro y con ello permite detectar más fácil la contaminación microbiana. El siguiente experimento tuvo como objetivo evaluar el efecto de cinco concentraciones de Gelrite en la respuesta in vitro y desarrollo de las plantas de pino. El medio de cultivo fue similar al empleado en el acápite anterior y los tratamientos fueron 2,5; 3,0; 3,5; 4,0 y 4,5 g.L-1. Después de 35 días de cultivo se evaluó en cada tratamiento, el número de plantas con necrosis total y parcial, la masa fresca y seca de 30 plantas por tratamiento y se determinó además el coeficiente de multiplicación. Por no existir normalidad de los datos en el caso de los porcentajes de materia seca, se empleó la prueba de Kruskal Wallis. La comparación entre parejas de grupo, se realizó con la prueba de Student Newman Keuls (SNK).. 3.2.4 Efecto de la sacarosa Este experimento tuvo como objetivo, evaluar en el subcultivo previo a la fase de enraizamiento, la influencia de diferentes concentraciones de sacarosa (30; 40; 50 y 60 g.L-1) en las características morfológicas de las plantas obtenidas en un medio de cultivo que fue similar al utilizado en el experimento anterior, pero sin adicionarle 6-BAP y con 3,5 g.L-1 de Gelrite. Las evaluaciones se realizaron a los 35 días de cultivo y se determinó en cada tratamiento, el número de plantas que por sus características morfológicas podían Tesis de Maestría. 33. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(38) y o. c u -tr a c k. .c. Materiales y Métodos ________________________________________________________________________________________. ser subcultivadas a la fase de enraizamiento, el número de nuevos brotes que por su desarrollo en longitud y grosor del tallo, no debían ser transferidos a la fase de enraizamiento y se determinó el porcentaje de materia seca de 30 plantas. Para el procesamiento estadístico de los resultados, por no existir normalidad de los datos se empleó la prueba de Kruskal Wallis. La comparación entre parejas de grupo, se realizó con la prueba de Student Newman Keuls (SNK).. 3.3 Fase de enraizamiento 3.3.1 Efecto de diferentes concentraciones de AIB Con el objetivo de lograr la formación de raíces in vitro, se obtuvieron plantas con 30 y 60 g.L-1 de sacarosa en un medio de cultivo similar al descrito para el experimento anterior y a los 35 días de cultivo, antes de realizar el subcultivo a la fase de enraizamiento, se evaluó la longitud de las plantas y se determinó el porcentaje de materia seca de 30 plantas obtenidas con cada concentración de sacarosa. Para analizar los porcentajes de materia seca se realizó un análisis de varianza simple, por cumplirse con los supuestos de normalidad y homogeneidad de los datos. El resto de las plantas se subcultivaron a un medio de cultivo de enraizamiento compuesto por los nutrientes inorgánicos WV5 a los cuales le fueron adicionados 0,6 mg.L-1 de tiamina para completar a 1,0 mg.L-1 la concentración de este compuesto, 1,0 g.L-1 de L-glutamina, 3,0 g.L-1 de carbón activado, 30 g.L-1 de sacarosa y 3,5 g.L-1 de Gelrite con pH ajustado a 5,8 antes de realizar el proceso de esterilización. Tesis de Maestría. 34. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
(39) y o. c u -tr a c k. .c. Materiales y Métodos ________________________________________________________________________________________. Los tratamientos de AIB estudiados fueron: 0,0; 4,90; 9,80; 14,70 µM y después de 30 días de cultivo, se evaluó para cada tratamiento, el número de plantas con raíces, el número de raíces por planta, la longitud de las raíces y la longitud de las plantas. En el caso de la longitud de las plantas, por existir normalidad, pero no homogeneidad de los datos, se aplicó la prueba de comparación de medias C de Dunnett.. 3.3.2 Efecto de la concentración de nutrientes inorgánicos Este experimento. tuvo. como. objetivo,. determinar. la influencia de. dos. concentraciones de nutrientes inorgánicos (50 y 100% de WV5) en la respuesta in vitro de las plantas en la fase de enraizamiento. Al igual que en el experimento anterior, como material vegetal se emplearon plantas obtenidas in vitro con dos concentraciones de sacarosa (30 y 60 g.L-1) en el subcultivo previo. En el caso del tratamiento con 50% de WV5, fue necesario adicionar 500 mg.L-1 de mio-inositol y 0,8 mg.L-1 de tiamina para completar a 1,0 g.L-1 y 1,0 mg.L-1 respectivamente la concentración de ambos compuestos. De esta forma, ambos tratamientos, solo presentaron diferencias en la concentración de los nutrientes inorgánicos. Después de 30 días de cultivo se evaluó en cada tratamiento, el número de plantas con raíces, el número de raíces por planta, la longitud de las raíces, la presencia de lignina a partir de visualizar las deposiciones de este compuesto en las paredes celulares, así como la cuantificación del contenido de lignina en las bases de segmentos de tallos (1,0 cm de longitud) de 18 plantas seleccionadas al azar.. Tesis de Maestría. 35. .d o. m. o. w. w. w. .d o. C. lic. k. to. bu. y bu to k lic C. w. w. w. N. O W. !. h a n g e Vi e. N. PD. !. XC. er. O W. F-. w. m. h a n g e Vi e. w. PD. XC. er. F-. c u -tr a c k. .c.
Figure




+7




Outline
Documento similar
Así, se estableció un protocolo de establecimiento alternativo para la variedad de color amarillo a partir de explantes conseguidos de hojas jóvenes; también se vio que en la
Analizar el impacto de la quimiocina CXCL12 en la propagación de virus con tropismo X4 en un modelo de infección in vitro en un cultivo de células dendríticas y linfocitos
Para reducir futuros problemas causados por dichos aspectos, los objetivos del trabajo se centraron en mejorar el proceso de transformación y cultivo in vitro de plantas
Development and viability of bovine embryos derived from oocytes matured and fertilized in vitro and co-cultured with bovine oviducal epithelial cells. Mechanisms of fertilization
Con base en lo anterior, la presente investigación tuvo como objetivo desarrollar un método de micropropaga- ción en cinco diferentes genotipos de guayabo evaluados en Tabasco
A los 6 años de edad, los valores medios de esbeltez fueron significativamente superiores con la aplicación de una dosis única de 300 g árbol -1 de (NPK); a los 35 años el
Se evaluaron dieciocho extractos vegetales y microorganismos eficientes (EM) sobre el crecimiento micelial in vitro y formación de esclerocios de Sclerotium rolfsii..
4 Diversos estudios se han realizado para cada una de las fases de la propagación in vitro pero existe una fase intermedia entre la germinación y la aclimatación, la cual
