TítuloLong term stability of Pseudomonas putida cultures suring the off gas treatment of toluene
10
0
0
Texto completo
(2) 448. RAÚL MUÑOZ, ANTONIA ROJAS, LUIS FELIPE DÍAZ, SERGIO BORDEL AND. SANTIAGO VILLAVERDE. nature of some of these VOCs and their excreted metabolic intermediates. This toxicity and mutagenic nature might impose irreversible or temporary looses of the degradation capacity of microorganisms during the long-term exposure at low concentration mutagenic contaminants, or as a result of short-term episodes of high inlet pollutant concentration (Jones et al., 1997; Mirpuri et al., 1997; Oliveira and Livingston, 2003). Thus, operational problems derived from microbial instability in processes treating toluene have been recently reported in literature. For instance, Song and Kinney (2005) reported a decline in the elimination capacity (EC) of biofilters subjected to high toluene loadings, likely due to the deterioration of the toluene degrading community. In their study, the process was maintained stable during 3 weeks at sub-critical conditions (RE = 100 %) and then overloaded. The EC immediately increased (concomitantly with a decrease in RE) reaching a pseudo-stationary state and started to decrease approx. 10-12 days after process overload. However, the mechanisms responsible of microbial deterioration were not identified. Likewise, Jones et al. (1997) and Villaverde et al. (1997) observed a decreasing fraction of toluene degrading Pseudomonas putida 54G in both suspended and biofilm-based bioreactors during the off-gas treatment of 0.5-3 g toluene m-3. This reduction, measured as cellular culturability on toluene, correlated with decreasing toluene degradation rates, being more pronounced the higher the toluene inlet concentration was. In addition, the fraction of respiring non-toluene culturable cells (supposed to grow at the expenses of leakage and lysis products) increased up to 95.5 %. Practical implications of this reduction in the fraction of toluene degrading cells might be the increase in culture oxygen requirements, which itself is problematic when treating high toluene loads (Villaverde et al., 1997). In this context, Leddy et al. (1995) and Brinkmann et al. (1994) reported that when present at high concentrations, toluene can induce irreversible defects in both plasmid and chromosomally encoded toluene degradation pathways in P. putida strains. Therefore, further research on the implications that toluene-mediated mutations in Pseudomonas culture impose on process performance, i.e EC, is needed in order to overcome operational problems derived from microbial instability in the long-term operation. In this context, Pseudomonas putida mt-2, harbouring the genes for toluene degradation in the TOL plasmid pWW0, and Pseudomonas putida F1, harbouring the chromosomally encoded TOD degradation pathway, were compared in terms of process stability under high toluene loadings. Culture stability was evaluated using process parameters such as EC, CO2 production, biomass concentration, fraction of toluene degrading cells, and specific toluene respiration rates..
(3) LONG-TERM STABILITY OF PSEUDOMONAS PUTIDA. 449. 2 MATERIALS AND METHODS 2.1 MICROORGANISMS AND CULTIVATION Pseudomonas putida mt-2 [DSM 3931] and P. putida F1 [DSM 6899] were selected for its well known capability to mineralize toluene using the TOL and TOD pathways, respectively (Worsey and Williams, 1975; Bordel et al., 2007). The MSM used for P. putida cultivation was composed of (g·l-1): NaHPO4· 12H2O, 6.15; KH2PO4, 1.52; (NH4)2SO4, 1.0; MgSO4·7H2O, 0.2; CaCl2, 0.038; and 10 ml·l-1 of a trace element solution containing (g·l -1): EDTA, 0.5; FeSO 4·7H 2O, 0.2; ZnSO 4·7H 2O, 0.01; MnCl2·4H2O, 0.003; H3BO3, 0.03; CoCl2·6H2O, 0.02; CuCl2·2H2O, 0.001; NiCl2·6H2O, 0.002; NaMoO4·2H2O, 0.003. The final pH of medium was 7.0. Pseudomonas putida mt-2 was cultivated in MSM where trace elements were increased by 100 %. 2.2 EXPERIMENTAL SETUP The influence of toluene concentration on process stability was investigated under aseptic conditions in a magnetically stirred 1-L glass bioreactor (Afora S.A, Spain) operated as a chemostat. Temperature and agitation rate were maintained constant at 25 °C and 500 rpm, respectively. The reactor was filled with 900 ml of sterile MSM and inoculated with 40 ml of the tested P. putida strain to attain an initial biomass concentration of approx. 4-7 mg Dry Weight l-1. Toluene was supplied in the gas phase through the aeration (1100 ml min-1 of synthetic air filtered through a 0.2 m Millex® –FG membrane filter) by mixing a toluene-saturated stream with a toluenefree air stream at different proportions (Fig. 1). When inoculating with P. putida mt-2, two series of continuous experiments were carried out during 4 days. In the first series of experiments (D = 0.2 h-1) toluene inlet concentration was set at 9.1 ± 0.9 g m-3. In the second series (D = 0.1 h-1 in order to avoid bacterial washout) the suspended growth reactor was fed with 3.2 ± 0.3 g toluene m-3. When testing P. putida F1, two series of continuous experiments were carried out during 32 days at D of 0.1 and 0.3 h-1, respectively. In the first series of experiments (D = 0.1 h -1) toluene inlet concentration was set at 3.3 ± 0.2 g m-3 during 21 days and thereafter increased up to 6.2 ± 0.4 g m-3. In the second series (D = 0.3 h-1) the chemostat was fed with 11.7 ± 0.7 g toluene m-3 during the first 21 days and thereafter with 20.2 ± 1.9 g toluene m-3. Gaseous toluene and CO2/O2 concentrations were daily monitored through valves A and C (Figure 1). Excreted metabolites, dissolved total organic carbon (TOC) content, pH, absorbance at 650 nm, and CFU plate counts in selective medium (toluene) and non-selective medium (peptone) were also daily recorded by withdrawing a 10 ml liquid sample through valve B under sterile conditions (Figure. 1). Dissolved Oxygen Concentration (DOC) and Temperature (T) were monitored on line. In addition, the specific O 2 consumption and the presence of catechol-2-3-dioxygenase were periodically monitored during toluene biodegradation by P. putida mt-2..
(4) 450. RAÚL MUÑOZ, ANTONIA ROJAS, LUIS FELIPE DÍAZ, SERGIO BORDEL AND. SANTIAGO VILLAVERDE. Figure. 1. Schematic representation of the experimental set up. 1, Compressed air; 2, Mass flow controller; 3, toluene evaporator; 4, mass flow controller; 5, air-filter; 6, stirred tank reactor, 7 MSM reservoir, 8 MSM pump. PC, pressure control; PI, pressure indicator.. 2.3 ANALYTICAL METHODS Toluene, CO 2 /O 2 , and benzyl alcohol (BA) and benzoic acid (BAc) concentrations were analyzed by GC-MS, GC-TCD, and HPLC, respectively, according to Bordel et al. (2007). The determination of dissolved Total Organic Carbon (TOC), pH, DOC and dry weight were also performed out according to Bordel et al. (2007). Respirometric assays were carried out to determine the time course of the specific O2 consumption during toluene biodegradation. Reaction vessels were filled with 15.6 ml of MSM, 2 ml of P. putida mt-2 (previously centrifuged at 6000 rpm during 15 min and resuspended in fresh MSM) and supplied with toluene at 25 mg l-1 (from a toluene saturated MSM stock solution). The total reaction volume was 18.5 ml. Tests in the absence of metabolites and toluene were also carried out under similar conditions to serve as controls for endogenous respiration. In P. putida F1 culture the fraction of viable bacteria degrading toluene was determined by standard plate count of colony-forming units (CFU) on non-selective medium (casein peptone 15 g l-1, soymeal peptone 5 g l-1, NaCl 5 g l-1, and agar 15 g l-1) and selective medium (MSM supplied with toluene vapors). Cells were incubated in.
(5) LONG-TERM STABILITY OF PSEUDOMONAS PUTIDA. 451. sealed containers for 24 h in triplicate at 30ºC. In P. putida mt-2 the fraction of toluene degrading cells was determined by periodically withdrawing samples from the bioreactor, diluting and spreading on agar plates with glucose as the sole carbon and energy source. After 1 day of growth, colonies were transferred and grid-arranged in triplicate to agar plates under non- selective (2 plates of glucose) and selective (1 plate grown on saturated toluene atmosphere) conditions. After 1 or 2 days of incubation at 30 ºC, colonies grown under non-selective conditions were subjected to the catechol2,3-dioxygenase test (see below) Plates grown under toluene atmosphere showed the TOL+ colonies. Absence of growth in selective medium plates indicated TOL- cells, which also corresponded with white colonies obtained in the catechol-2,3-dioxygenase (C23O) test. The presence of C23O was assayed according to Duetz et al. (1991). Between 450 and 750 colonies (after 1 day of growth at 30 ºC) on agar plates with glucose as the sole carbon and energy source) were screened by spraying the plates with a 250 mM catechol solution. Positive colonies (those with an intact TOL plasmid) turned bright yellow almost immediately, while those that had lost C23O activity remained white. Polymerase chain reaction (PCR) was used to search for the gene responsible for catechol-2,3-dioxygenase activity (xylE). The following specific oligonucleotides were designed: xylE-forward: 5’-TACTGGACATGAGCAAGGC-3’ and xylE-reverse: 5’-GATAGATGTGTCGGTCATGG-3’ (670-bp fragment). Nucleotide sequence was retrieved from Genebank, accession number AJ344068.1 (region 50914-51837) (Greated et al., 2002). 25-μL reactions had the following composition: 1 mM oligonucleotides (Roche-TIB MOLBIOL, Berlín, Germany); 80 μM dNTPs (Q-Biogen, Austin, Texas, USA); 0.1 U/25 μL Taq-polymerase (Roche, Barcelona, Spain); 1:10 dilution of Taq-polymerase buffer (Roche, Barcelona, Spain); volume up to 25 μL was completed with extra pure water. Temperature cycles were: initial denaturing step of 94ºC, 3 min; 30 cycles of 94ºC (40 sec), 62 ºC (40 sec), 72 ºC (40 sec); final elongation step 72ºC, 3 min. Cleaner PCR results were obtained when purified plasmid DNA was used as template (0.2 ng per reaction), than with direct colony-PCR. The 10-minute protocol for plasmid DNA isolation (Zou et al., 1990) was carried out for C23O- and C23O+ colonies. PCR results were visualized in 1.2%-agarose gels with 1xTAE buffer, according to standard protocols (Sambrook and Russel, 2002). Size changes on TOL plasmid was confirmed by restriction analysis of purified plasmid DNA. HindIII -fragment pattern of C23O- and C23O+ colonies was visualized on 0.6% agarose gels on 1xTAE (Sambrook and Russel, 2002)..
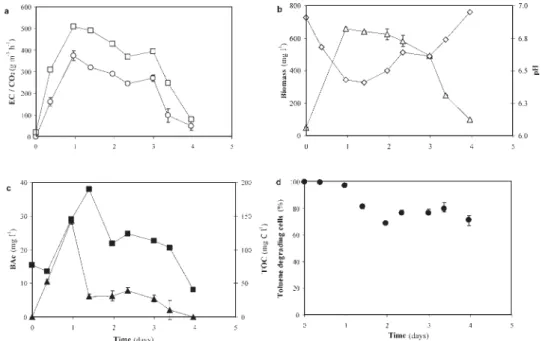
(6) 452. RAÚL MUÑOZ, ANTONIA ROJAS, LUIS FELIPE DÍAZ, SERGIO BORDEL AND. SANTIAGO VILLAVERDE. 3 RESULTS AND DISCUSSION When inoculated with P. putida mt-2 process collapse occurred after approx 3.5 days, regardless the toluene gas inlet concentration (9.1 and 3.2 g m-3). In both cases, bacterial activity (evaluated based on toluene elimination capacity (EC), CO2 production rate and specific respiration rate on toluene) increased during the first day of cultivation followed by a gradual decrease in both toluene EC and CO2 production (Figure 2a). This decay of bacterial activity corresponded with a steady decrease in biomass, TOC and benzoic acid concentrations, and an increase in the pH of the cultivation medium (Figure 2b,c). Likewise, the fraction of toluene degrading P. putida mt-2 started to decrease during the second day of cultivation reaching a steady state value of 73 ± 4 % until the end of the experimentation, which was independent of the loading rate scenario (Figure 2d). Non-toluene degrading variants of P. putida mt-2 did not become yellow after spraying with catechol, indicating no basal activity of the catechol 2,3 dioxygenase (Brinkmann et al., 1994). PCR and plasmid restriction analyses of non toluene degrading isolates indicated deletion of a significant portion of the pWW0 plasmid (Data not shown).. Figure 2. Time course of EC ( ), CO2 production ( ), biomass concentration (Δ), pH ( ), benzoic acid ( ), TOC ( ), and fraction of toluene degrading cells ( ) during toluene biodegradation by P. putida mt-2 operated at 9.1 g m-3 and D = 0.2 h-1..
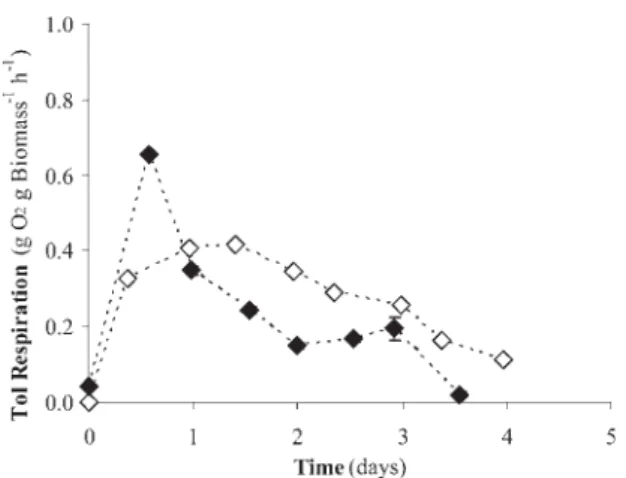
(7) LONG-TERM STABILITY OF PSEUDOMONAS PUTIDA. 453. These results are in agreement with previous observations reported in literature. For instance, Villaverde et al. (1997) reported a decrease in the toluene culturable cell fraction down to 2 % after 3 month of operation in flat plate biofilm reactor exposed to toluene liquid concentrations of 7 mg l-1. In a similar study, Jones et al. (1997) observed that the number of toluene-mediated injured cells correlated negatively with the time of exposure and toluene concentration, although in both cases the reason of this significant bacterial deterioration were not identified. Several authors have pointed out that some intermediates from toluene biodegradation might induce severe irreversible defects in both plasmid and chromosomal-associated pathways in Pseudomonas cultures. Partial deletions or curing of the TOL plasmid have been already reported in P. putida mt-2 during growth on toluene, BA or BAc (Brinkmann et al., 1994; Leddy et al., 1995). Likewise, Duetz et al. (1991) recorded a total loss of the C23O activity in P. putida mt-2 grown on benzoic acid in a 120-h phauxostat culture. In our study, both benzoic acid and benzyl alcohol were always detected when operating at very low concentration as a result of toluene biodegradation via the TOL pathway, which might have promoted the loss of toluene catabolic capacity likely due to the deterioration of both plasmidic and chromosomal pathways (Brinkmann et al., 1994; Leddy et al., 1995). However, the decrease in the fraction of toluene degrading P. putida mt-2 cells observed in our study can not explain itself the severe decrease in process performance recorded in the process. Indeed, a constant rate of segregation of P. putida variants would finally render to a stable steady state, but never to bacterial washout. Further analysis of the physiological state of the culture based on respirometric tests suggested an accumulative toluene-mediated microbial damage, which could explain the deterioration of the microbial culture and therefore process instability. Thus, the specific respiration rate on toluene steadily decreased after 1 days of cultivation regardless the toluene inlet concentration (Figure 3). Aromatic compounds such as toluene can act as uncoupplers diminishing the efficacy of the respiratory chain and therefore reducing microbial metabolic activity (de Smet et al., 1978). These aromatic pollutants are also known to increase cell permeability by damaging the cytoplasmatic membrane, which contributes to the loss of ions, cytoplasmatic material and proton gradients. In this regard, Dominguez-Cuevas et al. (2006) have recently suggested that membrane damage in a pWW0 bearing P. putida KT2440 (similar to P. putida mt-2) strain induced high levels of oxidative stress and inhibition of the most productive steps of the aerobic metabolism. On the other hand, P. putida F1 exhibited stable performance at all tested toluene inlet concentrations (up to 20.2 g m-3) as shown by the constant values of EC, CO2 production, pH and biomass concentration within each steady state (Figure 4a,b). Thus, no mutant population established in the culture at the expense of the organic carbon excreted by the wild type P. putida F1 cells. This was confirmed by two facts:.
(8) 454. RAÚL MUÑOZ, ANTONIA ROJAS, LUIS FELIPE DÍAZ, SERGIO BORDEL AND. SANTIAGO VILLAVERDE. Figure 3. Time course of the specific toluene respiration rate in P. putida mt-2 cultures operated at high ( ) and low ( ) toluene loadings.. Figure 4. Time course of EC ( ), CO2 production ( ), biomass concentration (Δ), pH ( ), benzoic acid ( ), TOC ( ), and fraction of toluene degrading cells ( ) during toluene biodegradation by P. putida F1 operated at D = 0.1 h-1 and 3.3 g m-3 and 6.2 g m-3..
(9) LONG-TERM STABILITY OF PSEUDOMONAS PUTIDA. 455. Firstly, TOC and BA remained constant within each steady state (Figure 4c), and secondly, no significant differences in the number of CFU growing on selective and non-selective medium was observed throughout the experimentation (Figure 4d). Benzyl alcohol, a curing agent causing TOL plasmid deletion and damage at chromosomal level in Pseudomonas strains, was present in the cultivation medium as a result of the monooxygenation of toluene by the dioxygenase system of P. putida F1 (Bordel et al., 2007). However, despite being exposed to BA and inlet gas toluene concentrations of up to 60 mg l-1 and 20.2 g m-3, respectively, there was no significant effect on the overall process performance over 1 months of operation. Both P. putida strains exhibited similar performances in terms of EC and CO2 production. Thus, when the process was operated at D = 0.1 h-1 and 3.3 g toluene m-3, toluene was degraded at rates of approx. 200 g m-3 h-1. However, toluene is sensed as a stressor rather than a nutrient by P. putida mt-2. Based on expression profiles, Dominguez-Cuevas et al. (2006) reported that in the presence of toluene, the bulk of the available transcriptional machinery of a pWW0-bearing P. putida is reassigned to endure general stress, while only a small share is redirected to the degradation of the aromatic compound.. 4 CONCLUSIONS Despite the numerous studies on the effect of toluene-mediated mutations in Pseudomonas putida cultures, no systematic studies of the influence of these mutations on process performance (EC, CO2 production) have been, to the best of our knowledge carried out. This work demonstrated that under high toluene loadings P. putida F1 exhibited a much more stable performance than P. putida mt-2. Process collapse in P. putida mt-2 cultures occurred in approx 4 days as a result of membrane damage and further inhibition of the aerobic metabolism as shown by a decaying toluene respiratory activity. No significant decrease in the overall performance of the biodegradation process was however recorded over 32 days of operation when using P. putida F1. Therefore, the selection of highly resistant bacterial strain is crucial in order to ensure not only the viability but also the long term stability of the biodegradation process specially when operating at high loading rates.. 5 ACKNOWLEDGEMENTS This research was supported by the Spanish Ministry of Education and Science (PPQ2006-08230 and JCI-2005-1881-5 contracts)..
(10) 456. RAÚL MUÑOZ, ANTONIA ROJAS, LUIS FELIPE DÍAZ, SERGIO BORDEL AND. SANTIAGO VILLAVERDE. REFERENCES Bordel, S., Diaz, L.F., Muñoz, R. and Villaverde, S. (2007) New insights on toluene biodegradation by Pseudomonas putida F1: Influence of pollutant concentration and excreted metabolites. Appl. Microbiol. Biotechnol. 74: 857-866. Brinkmann, U., Ramos, J.L. and Reineke, W. (1994) Loss of the Tol meta-cleavage pathway functions of Pseudomonas putida strain PaW1 (pWW0) during growth on toluene. J. Basic Microbiol. 34 (5): 303-309. de Smet, M.J., Kingma, J. and Witholt, B. (1978) The effect of toluene on the structure and permeability of the outer and cytoplasmic membranes of Escherichia coli. Biochim. Biophys. Acta 506: 64-80. Dominguez-Cuevas, P., Gonzalez-Pastor, J.E., Marques, S., Ramos, J.L. and de Lorenzo, V. (2006) Transcriptional tradeoff between metabolic and stress-response programs in Pseudomonas putida KT2440 Cells exposed to toluene. J. Biol. Chem. 281(17): 11981-11991. Duetz, W.A., Winson, M.K., van Andel, J.G. and Williams, P.A. (1991) Mathematical analysis of catabolic function loss in a population of Pseudomonas putida mt-2 during non-limited growth on benzoate. J. Gen. Microbiol. 137: 1363-1368. Greated, A., Lambertsen, L., Williams, P.A. and Thomas, C.M. (2002) Complete sequence of the IncP-9 Tol plasmid pWW0 from Pseudomonas putida. Environ. Microbiol. 4(12): 856-871. Jones, W.L., Mirpuri, R.G., Villaverde, S., Lewandowski, Z. and Cunningham, A.B. (1997) The effect of bacterial injury on toluene degradation and respiration rates in vapor phase bioreactor. Water Sci. Technol. 36(1): 85-92. Leddy, M.B., Phipps, D.W. and Ridgway, H.F. (1995) Catabolite-mediated mutations in alternate toluene degradative pathways in Pseudomonas putida. J. Bacteriol. 177(16): 4713-4720. Mirpuri, R., Jones, W. and Bryers, J.D. (1997) Toluene degrading kinetics for planktonic and biofilm grown cells of Pseudomonas putida 54G. Biotechnol. Bioeng. 53: 535-46. Oliveira, T.A.C. and Livingston, A.G. (2003) Bioscrubbing of waste gas-substrate absorber to avoid instability induced by inhibition kinetics. Biotechnol. Bioeng. 84(5): 552-563. Sambrook, J. and Russel D.W. (2002) Molecular cloning: a laboratory manual. Third edition. Cold Spring Harbour aboratory Press, Cold Spring Harbor, New York. Song, J.H. and Kinney, K.A. (2005) Microbial response and elimination capacity in biofilters subjected to high toluene loadings. Appl. Microbiol. Biotechnol. 68: 554-559. Villaverde, S., Mirpuri, R.G., Lewandowski, Z. and Jones, W.L. (1997) Physiological and chemical gradients in a Pseudomonas putida 54G biofilm degrading toluene in a flat plate vapor phase bioreactor. Biotechnol. Bioeng. 56: 361-71. Worsey, M.J. and Williams, P.A. (1975) Metabolism of toluene and the xylenes by Pseudomonas putida arvilla (mt-2): evidence for a new function of the TOL plasmid. J. Bacteriol. 124: 7-13. Zou, C., Yang, Y. and Y. Jong A.Y. (1990). Miniprep in Ten minutes. Biotechniques 8 (2): 172-173..
(11)
Figure
Documento similar
More specifically, Theorem 1 shows that under any work-conserving HL service discipline, a subcritical multi-class queueing net- work satisfying state space collapse (in fluid
Mönch [19] employs the term agent-based manu- facturing to emphasize the impact of this paradigm because activities such as process planning [13, 14], holonic control, and
After compound 1 was shaped into pellets and membranes, the stability of these processed materials was examined against possible operating industrial conditions such as moisture
We propose that dark matter is stable as a consequence of an accidental Z 2 that results from a flavour symmetry group which is the double-cover group of the symmetry group of one
Liquid–liquid extraction/headspace/gas chromatographic/mass spectrometric determination of benzene, toluene, ethylbenzene, (o-, m- and p-)xylene and styrene in olive oil using
I have been inspired by Jameson as the leading analyst of the reflection of our mode of production in contemporary culture, as an investigator of realism, and as a fundamental
Previous studies on the single treatment of strawberry waste extrudate by anaerobic digestion have reported an improvement in methane production, stability, and the allowed organic
In this work, a systematic experimental and theoretical analysis of the vapor-liquid equilibrium of {aromatic hydrocarbon (toluene) + ionic liquid} binary mixtures
