Syndecan 4 and beta 1 integrin are regulated by electrical activity in skeletal muscle: Implications for cell adhesion
10
0
0
Texto completo
(2) 384. G. Ugarte et al. / Matrix Biology 29 (2010) 383–392. 70 µm cell strainer. Cell suspensions were pre-plated to eliminate fibroblasts, and non-adherent cells were plated onto collagen-coated culture dishes at a density of 100,000 cells/ml. Myoblasts were grown at 37 °C with 8% CO2 in DMEM supplemented with 10% fetal bovine serum and 10% horse serum (HS). After 48 h, the medium was changed to DMEM 10% HS to induce myotube formation, with 10 µM cytosine-β-D-arabinofuranoside to inhibit fibroblast proliferation. Under these conditions, primary myotubes showed spontaneous contractions as of day 2 in differentiation medium. To inhibit electrical activity, 1 μg/ml tetrodotoxin (TTX; Alomone, Jerusalem, Israel) was added to cell cultures at day 1 of differentiation.. integrin regulates cell adhesion through the phosphatidylinositol 3kinase (PI3K)/Akt signaling pathway (Wang et al., 2007). In addition, protein phosphatase 2A (PP2A) is associated physically with β1 integrin in muscle cells under differentiation and regulates the phosphorylation state of integrin β1A (Kim et al., 2004). It is well documented that syndecan-4 and integrins have crucial and complementary roles in cell adhesion and migration. In fact, syndecan-4 can act as a co-receptor for α5β1 integrins in the assembly of focal adhesions during the spreading of adherent cells (Mostafavi-Pour et al., 2003). Costameres – striated muscle homologs to focal adhesions in nonmuscle cells – are protein assemblies aligned between the sarcolemma and underlying sarcomeric myofibrils in a periodic array coincident with Z-discs. These macromolecular structures, containing focal adhesion proteins such as vinculin, talin, α-actinin, and β1 integrin, are considered sites where contractile forces from sarcomeres are transmitted across the plasmalemma to the ECM or adjacent cells (Ervasti, 2003). Skeletal muscle determination of these progenitors involves the activity of bHLH transcription factors such as Myf5 and MyoD. Terminal differentiation corresponds to the expression of myogenin and Mef2, and the outcome is the fusion of mononucleated myoblasts in multinucleated myotubes that express muscle-specific proteins such as myosin heavy chain and creatine kinase (Buckingham et al., 2003). In addition, skeletal muscle cells develop excitability and contractile properties during the first stages of myogenesis. This phenotype has been associated to the onset of voltage-dependent channels protein expression (sodium and calcium channels) and the maturation of the excitation–contraction apparatus (Brodie et al., 1989; Grouselle et al., 1991; Cognard et al., 1993). At more advanced developmental stages, muscle activity is controlled by motor innervation (Sanes and Lichtman, 1999). The last steps of the muscle differentiation process can be recapitulated in vitro using murine embryonic or fetal myoblasts induced to differentiate to myotubes in low serum culture conditions. The aim of this work is to determine how membrane molecules involved in cell adhesion such as syndecan-4 and β1 integrin can be modulated by an intrinsic property such as electrical activity in skeletal muscle cells under differentiation. Our results demonstrate that protein levels of both molecules are decreased after the inhibition of activity, disrupting their costameric localization and reducing myotube adhesion. This evidence indicates that the activity-dependent positive regulation of myofiber adhesion might be required for normal muscle physiology.. Proteins were extracted from primary cultures using a Tris/NaCl buffer containing 0.5% Triton X-100 plus protease inhibitors. Protein concentrations were determined using the Micro BCA™ protein assay kit (Pierce, Rockford, IL, USA). Samples were electrophoresed in 10% polyacrylamide gels (or in 4–12% gradient gels for separate HSPGs), transferred onto nitrocellulose membranes and probed with one of the following antibodies: mouse anti-Δ-heparan sulfate 3G10 (1:2500) (USBiological, Swampscott, MA, USA); rabbit anti-TGF-β type I receptor (TβRI) (1:500) (Santa Cruz Biotechnology, Santa Cruz, CA, USA); rabbit anti-β1 integrin (1:1000) (Santa Cruz Biotechnology); rabbit antiphospho-Akt (Ser 473) (1:1000) (Cell Signaling Technology, Beverly, MA, USA); rabbit anti-Akt (1:1000) (Cell Signaling Technology); rabbit anti-alpha 5 integrin (1:1000) (Santa Cruz Biotechnology); rabbit anti-phosphorylated Smad-2 (1:500) (Calbiochem, San Diego, CA, USA); or mouse anti-GADPH (1:5000) (Chemicon, Temecula, CA, USA) (Fuentealba et al., 1999; Ugarte and Brandan, 2006; Cabello-Verrugio and Brandan, 2007; Vial et al., 2008). After incubation with HRPconjugated secondary antibodies (1:10,000) (Pierce, Rockford, IL, USA), reactive proteins were visualized using chemiluminescent substrates (Pierce, Rockford, IL, USA). For the detection of phospho-Akt, proteins were extracted in RIPA buffer plus protease inhibitors.. 2. Materials and methods. 2.5. Immunofluorescence analysis. 2.1. Animals and muscle denervation. Cells were fixed in 3% paraformaldehyde, permeabilized with 0.05% Triton X-100, blocked with BSA and incubated with antisyndecan-4 (1:1000), anti-β1 integrin (1:250), mouse anti-vinculin (1:250) (Sigma, St. Louis, MO, USA) or mouse anti-myosin heavy chain (1:500) (Sigma) primary antibodies. Bound antibodies were detected by incubating the cells with appropriate 1:1000 fluorochrome-conjugated secondary antibodies (Alexa Fluor 488/568, Molecular Probe, Eugene, OR, USA). After rinsing, the slides were viewed through a SW1000 Olympus confocal microscope.. Two-month-old male Sprague–Dawley rats were anesthetized with xylazine/ketamine and right legs were denervated by a sciatic nerve section of 0.5 cm in the hip region of the hind limb, as previously described (Fadic et al., 1990). After 72 h, the tibialis anterior muscle was excised for analysis. Muscles from contralateral legs were used as controls. For protein extracts, muscle tissue was frozen with liquid nitrogen, triturated and homogenized in NaCl/Tris buffer containing 1% SDS, 1% Triton X-100 and protease inhibitors. Extracts were then centrifuged at 20,000 ×g for 10 min, and supernatants collected and analyzed for protein quantification. HSPGenriched fractions from the samples were obtained following the method detailed below. 2.2. Cell cultures Rat skeletal myoblasts were isolated from the hind legs of day 19 Sprague–Dawley rat fetuses. Briefly, muscle mass was dissected from bones, subjected to mechanic dissociation and filtered through a. 2.3. Electrical stimulation Primary cultures were stimulated as described by Chahine et al. (1992). For electrical stimulation the electrodes were immersed in culture medium and myotubes were stimulated using a Grass stimulator with 0.4 ms/4 V pulses (threshold required to induce visible contractions) in 100 Hz trains lasting 1 s, applied once every 100 s for 24 h. 2.4. Western blotting. 2.6. Semi-quantitative RT-PCR analysis Total RNA was isolated from primary myotubes using TRIzolR (Invitrogen) following the protocol described by Chomczynski and Sacchi (1987). cDNA was synthesized from total RNA, previously treated with DNAse I, primed with random primers and reversed transcribed with M-MLV reversed transcriptase. PCR cycling conditions for rat syndecan-4 were 31 cycles consisting of 94 °C for 30 s, 56 °C for 60 s and 72 °C for 60 s, followed by a 10 min final extension at 72 °C. Twenty-five cycles of the same cycling protocol were used for.
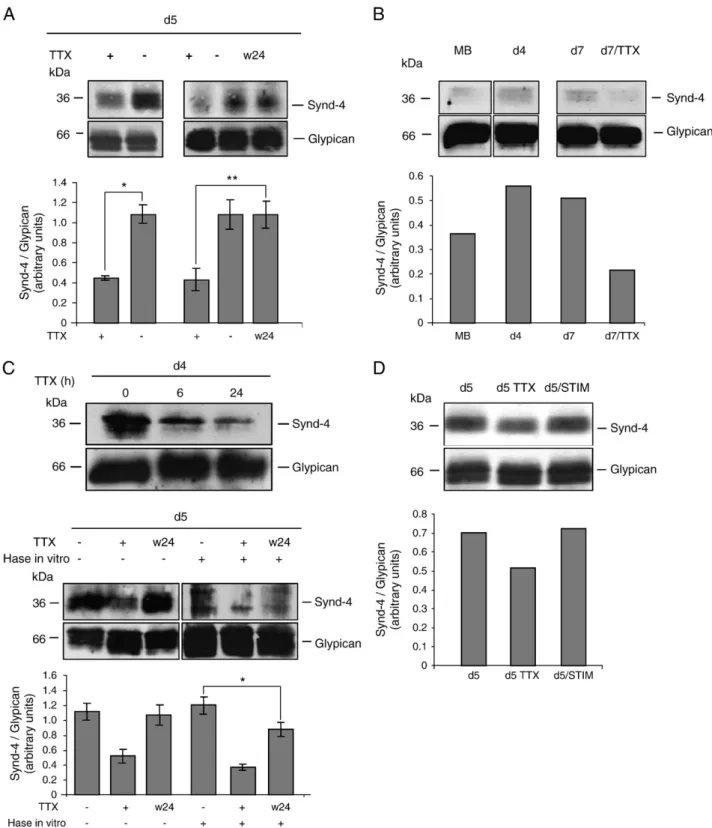
(3) G. Ugarte et al. / Matrix Biology 29 (2010) 383–392. β-actin. The PCR products were analyzed by agarose gel electrophoresis. The primers for syndecan-4 were forward: 5-TTGGAAC CATGGCGCCT-3 and reverse: 5-TGCGTAGAACTCGTTGGTGG-3, which yielded a PCR product of 613 bp. The primers for rat β-actin were forward: 5-TCTACAATGAGCTGCGTGTG-3 and reverse: 5-TAC ATGGCTGGGGTGTTGAA-3, which yielded a PCR product of 131 bp. 2.7. Northern blotting Ten micrograms of total RNA samples was electrophoresed in 1.2% agarose/formaldehyde gels, transferred onto Nytran membranes (Schleicher & Shuell, Germany) and hybridized with random primed 32 [ P]dCTP-labeled cDNA probes for rat syndecan-4 or tubulin in hybridization buffer overnight at 42 or 65 °C, respectively. Hybridized membranes were then washed at 42 °C and exposed to a Phospho Imager (Vial et al., 2008). The cDNA probe for rat syndecan-4 corresponds to a fragment of 609 bp that was amplified by RT-PCR using the following primers: forward: 5′-GAGAAAGCTTATGGCGCCT GTCTGCCTGTT-3′ and reverse: 5′-GCTAGAGGATCCTCATGCGTA GAACTCGTTGGTGG-3′. 2.8. Purification of HSPGs An enriched fraction proteoglycans was obtained from cell extracts prepared in Tris/NaCl buffer containing 0.5% Triton X-100 plus protease inhibitors. Samples were incubated with DEAE weak anion exchange support overnight at 4 °C, eluted with 1.5 M NaCl and then dialyzed (Brandan et al., 1996). HSPGs were detected in western blots with antiΔ-HS after digestion with heparitinase (USBiological) for 6 h at 37 °C (Steinfeld et al., 1996; Fuentealba et al., 1999; Dovas et al., 2006).. 385. after digestion with heparitinase. Fig. 1A shows that syndecan-4 core protein levels (∼36 kDa) exhibit a biphasic behavior as they change during the differentiation of primary myotubes. The levels are high early during the first days of differentiation (day 2), then decrease and increase at later phases (past day 5). Glypican core protein levels do not change and syndecan-1 is down-regulated during this process as previously reported (Larrain et al., 1997). Syndecan-3 core protein (reactive band around 120 kDa) is expressed mainly in early myotubes (day 2) and does not change significantly at more advanced stages. The biphasic behavior of syndecan-4 was also observed in the levels of transcripts by northern blot analysis (Fig. 1B), suggesting that during myoblast differentiation the amount of syndecan-4 core protein correlates with changes in gene expression. Under culture conditions, rat primary myotubes start to develop spontaneous electrical activity associated to visible contractile activity at day 2 of differentiation (Dutton et al., 1993; Ugarte and Brandan, 2006). To determine whether the amount of syndecan-4 is regulated by electrical activity in primary cultures, this activity was inhibited by the addition of TTX to culture media. TTX specifically blocks voltagedependent sodium channels in a reversible manner, demonstrating the recovery of the contractile activity of myotubes a few minutes after the toxin had been washed out. Fig. 2A shows that the inhibition of electrical activity for 72 h strongly decreases the levels of syndecan4 core protein. The protein levels can be recovered in 24 h after the wash-out of TTX (lane w24). Quantitative analysis is shown in the. 2.9. Adhesion assay Cell cultures were washed three times with PBS, incubated in PBS containing 10 mM EDTA for 10 min at 37 °C and then gently shaken for 1 min at RT. Supernatants were collected, and attached cells were scraped for protein and DNA quantification. 2.10. Cell viability Cell viability was measured by the modified 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay (Mosmann, 1983) using MEM medium without phenol red (Sigma). 2.11. Apoptotic assay Myotubes (day 4 differentiation) were incubated with or without TTX. Apoptotic nuclei were induced by H2O2 incubations for 4 h under culture conditions. Then the cells were fixed and stained with Hoechst (Alvarez et al., 2004). 2.12. Cell culture heparitinase digestion Cell cultures were incubated in 600 μL of minimum media containing 1% BSA and 1 mU of heparitinase for 90 min at 37 °C (Gutierrez et al., 2006). 3. Results 3.1. Protein levels of cell surface syndecan-4 are regulated by electrical activity in differentiating myotubes Little is known about how the regulation of syndecan-4 expression changes during the differentiation of skeletal muscle cells. To analyze this question HSPG protein levels were analyzed by western blot using an antibody (anti-Δ-HS) that recognizes an epitope generated in HSPGs. Fig. 1. Syndecan-4 changes with a biphasic behavior during the differentiation of skeletal muscle cells. A) Western blot (anti-Δ-HS) of post-DEAE-enriched fractions of HSPGs corresponding to the different days of differentiation of rat primary cultures. The band corresponding to the core protein of glypican, unmodified during myotube differentiation, is shown as a loading control. B) Northern blot of total RNA samples of primary myotubes at day 2, 4 and 6 of differentiation, using a specific probe for rat syndecan-4, as described in Materials and methods. The reactive band corresponding to the syndecan-4 transcript is revealed at 2.1 kb. A tubulin probe is shown as a loading control revealed at 1.7 kb..
(4) 386. G. Ugarte et al. / Matrix Biology 29 (2010) 383–392. Fig. 2. Syndecan-4 at the cell surface is regulated by electrical activity during the differentiation of skeletal muscle cells. A) Top, western blot (anti-Δ-HS) of post-DEAE-enriched fractions of HSPG corresponding to TTX-treated and control cultures at day 5 of differentiation. Right panel, western blot (anti-Δ-HS) of post-DEAE-enriched fractions of HSPGs of total extracts, corresponding to TTX-treated cultures, control and TTX-treated for 48 h, washed and lysed 24 h afterwards (w24). Bottom, the values shown are the results obtained from three independent experiments performed in triplicate and correspond to the mean and standard deviation. *P b 0.05, **P b 0.02. B) Top, western blot (anti-Δ-HS) of post-DEAEenriched fractions of HSPG corresponding to RMT461 myoblasts, myotubes at day 4 of differentiation, myotubes at day 7 and TTX-treated myotubes at day 7. Bottom, the values shown are the results obtained from one experiment. C) Upper panel: western blot (anti-Δ-HS) of total cell extracts from cultures (at day 4 of differentiation) treated with TTX for 6 or 24 h and then incubated with heparitinase (Hase) in vitro. Lower panel: Top. Western blot (anti-Δ-HS) of post-DEAE-enriched fractions of HSPGs of total extracts (at day 5 of differentiation) corresponding to control, TTX-treated and TTX-treated for 48 h, washed and lysed 24 h afterwards (w24) (lanes 1–3). Lanes 4–6: western blot (anti-Δ-HS) of total cell extracts from cultures in the same three conditions but cells were incubated with heparitinase (Hase) in vitro. Bottom. The values shown are the results obtained from three independent experiments performed in triplicate and correspond to the mean and standard deviation. *P b 0.02. D) Top, western blot (anti-Δ-HS) of post-DEAE-enriched fractions of HSPG corresponding to control and stimulated cultures for 24 h at day 5 of differentiation. The band corresponding to the core protein of glypican, unmodified during myotube differentiation, is shown as a loading control. Bottom, the values shown are the results obtained from one experiment..
(5) G. Ugarte et al. / Matrix Biology 29 (2010) 383–392. same figure. This activity-dependent regulation of syndecan-4 was also observed in the immortalized cell line RMT461 generated from rat primary myoblasts, which also develops spontaneous electrical activity (Macpherson et al., 2002). The levels of syndecan-4 are lower in TTXtreated myotubes than control cultures at day 7 of differentiation (see Fig. 2B). Quantitative analysis is shown in the same figure. The inhibition of electrical activity with TTX does not affect myoblast fusion or the expression of late markers of skeletal muscle differentiation such as myosin (data not shown). To determine whether the activity-dependent regulation of syndecan-4 correlates to modifications in the amount of HSPG present at the cell surface, living myotube cultures were incubated with heparitinase. In this way, only the surface-exposed molecules were digested by the enzyme added to the culture medium and then detectable by the anti-ΔHS antibody in western blots. Fig. 2C, top shows that syndecan-4 core protein starts to decrease 6 h after the inhibition of spontaneous electrical activity and becomes more evident after 24 h, suggesting that electrical inactivity affects the amount of syndecan-4 present at the cell surface. Fig. 2C, low panel shows that 24 h after restoring the electrical activity by the wash-out of TTX, the levels of the total pool of the core protein were recovered (Fig. 2C, lower panel, lanes 1-3). Noteworthy, the syndecan-4 core protein at the cell surface, evaluated by the digestion with heparitinase in living cultures, is not completely restored after this time (Fig. 2C, lower panel, lanes 4-6). As loading control the figure shows the unchanged levels of glypican-1 core protein. Quantitative analysis is shown in the same figure. To test whether enhanced electrical activity in primary myotubes could positively regulate syndecan-4 protein levels, we studied the effect of electrical stimulation with extracellular electrodes over syndecan-4 core protein levels. This method has been successfully employed to study the electrical activity-dependent expression of several muscle proteins such as the nicotinic acetylcholine receptor and myogenin (Chahine et al., 1993; Macpherson et al., 2002). Fig. 2D shows that after 24 h of stimulation, syndecan-4 core protein levels are specifically increased. As loading control the figure shows the unchanged levels of glypican-1 core protein. Quantitative analysis of this experiment is shown in the same figure. These experiments confirm that syndecan-4 can be regulated by myotube excitability during the differentiation process. To determine whether the activity-dependent regulation of syndecan-4 core protein is correlated with changes at the transcriptional level, semi-quantitative RT-PCR and northern blot assays were performed. Fig. 3A shows that the fragment amplified by PCR corresponding to syndecan-4 is unmodified in TTX-treated myotubes compared with control cultures. A similar result was obtained in the northern blot assay (Fig. 3B). These results suggest that transcripts for this HSPG are unchanged in the inactive myotubes and that the regulation of syndecan-4 by electrical activity does not involve modifications of transcriptional activity of the gene. 3.2. Syndecan-4 is down-regulated in denervated muscles To evaluate if the regulation of protein levels of syndecan-4 by electrical activity also occurs in adult skeletal muscle in vivo, the levels of syndecan-4 after a short-term motor denervation of rat hind limb muscles were analyzed. Fig. 4A shows that syndecan-4 core protein levels are decreased in the denervated tibialis anterior compared with the contralateral control muscle. By contrast, in the denervated muscle TβRI is up-regulated as previously reported (Fig. 4B) (Ugarte and Brandan, 2006). This result indicates that the inhibition of muscle activity can also affect syndecan-4 levels in vivo. 3.3. The levels of β1 integrin are regulated by electrical activity during skeletal muscle differentiation Considering that syndecan-4 and β1 integrins can cooperate in the cell migration and adhesion processes, the β1 integrin levels in. 387. Fig. 3. Syndecan-4 transcripts are unmodified in inactive myotubes. A) Semi-quantitative RT-PCR for syndecan-4 (613 bp) from total RNA extracted from control and inactive cultures (TTX) at day 4 of differentiation. PCR product for β-actin (131 bp) is shown below as the control of the amount of RNA used in reversed transcription. B) Northern blot of total RNA samples from control myotubes (day 5 of differentiation), TTX-treated, and TTXtreated, washed out and lysed 24 h afterwards (w24), using a specific probe for rat syndecan-4, as described in Materials and methods. The reactive band corresponding to the syndecan-4 transcript is revealed at 2.1 kb. The tubulin probe is shown as a loading control revealed at 1.7 kb.. control and TTX-treated primary myotubes were analyzed. Fig. 5A shows that β1 integrin protein levels increase between day 2 and 4 of myoblast differentiation (lanes 1, 3 and 5) and progressively decrease after the inhibition of electrical activity with TTX (lanes 2, 4 and 6). The band detected by the anti-pan-β1 antibody used in this blot probably corresponds to a 1D isoform, which is strongly induced during myotube differentiation (Kim et al., 2004). This regulation seems to be specific for β1 integrin since the amount of α5 integrin is unaffected (data not shown). To determine whether the levels of β1 integrin can be positively regulated by electrical activity, the modification of protein levels following the recovery of spontaneous electrical activity after the wash-out of TTX was analyzed. Fig. 5B shows that protein levels of β1 integrin after the inhibition of electrical activity for 24 h increase progressively from 6 h after restoring the spontaneous activity and almost reaches control levels after 24 h. The lower panel shows that the recovery of β1 integrin protein levels is accompanied of the down-regulation of TβRI, which is negatively regulated by spontaneous electrical activity (Ugarte and Brandan, 2006). To determine whether intermediates of the intracellular cascades associated with syndecan-4 and β1 integrin are affected by the inhibition of electrical activity, the phosphorylation state of Akt kinase was evaluated. Fig. 5C shows that TTX-treated myotubes contain Akt.
(6) 388. G. Ugarte et al. / Matrix Biology 29 (2010) 383–392. 3.5. Syndecan-4 and β1 integrin co-localize with vinculin in costameric structures, which are lost by the inhibition of spontaneous electrical activity. Fig. 4. Syndecan-4 is down-regulated in vivo after the denervation of adult muscles. A) Western blot (anti-Δ-HS) of post-DEAE-enriched fractions of HSPG corresponding to rat tibialis anterior muscle (TA) 72 h after denervation (Den) or from control-innervated muscles (Ctrl). B) Immunodetection of TβRI as a denervation control for the same samples (21). Core protein of glypican and GAPDH are shown as loading controls.. kinase in a hyperphosphorylated state compared with control cultures. The basal level of the phosphorylation of Akt can be recovered after the wash-out of TTX (w48), without affecting total Akt protein levels. The specificity of the reactive band for phospho-Akt was verified in C2C12 myoblast cultures after the addition of IGF, a canonical activator of Akt-dependent intracellular cascade (right panel). Although the relationship between the TGF-β-dependent signaling cascade and Akt is documented (Zhang, 2009), the hyperphorylation of Akt cannot be explained by the inactivity-triggered up-regulation of TβRI. The inhibition of the kinase activity of this receptor with SB 431542 (Laping et al., 2002) does not affect the state of phosphorylation of Akt in control and TTX-treated cultures (Fig. 5D), suggesting that an autocrine loop of TGF-β or basal activity of the receptor in differentiating fetal myoblasts is negligible in these conditions. The effect of the inhibitor SB 431542 on Smad-2 phosphorylation in response to TGF-β is shown in Fig. 5D, right panel. 3.4. Myotubes cell adhesion depends on electrical activity To evaluate the impact of the decrease in protein levels of syndecan-4 and β1 integrin, the adhesion capacity of active and paralyzed myotubes was determined. Fig. 6A shows that after incubating cultures with PBS-EDTA, TTX-treated myotubes detach from the substrate to a greater extent that control myotubes, as determined by the amount of proteins and DNA contained in both supernatants and culture plate-associated fractions. The effect of TTX cannot be attributed to a reduction in cell viability, since no effect was observed by MTT reduction assay (Fig. 6B). Also, the effect of TTX cannot be attributed to an increase in apoptosis of the myotubes since the number of apoptotic nuclei was similar in TTX-treated or untreated myotubes (Fig. 6C). These results suggest that the adhesion capacity of differentiating myotubes strongly decreases after the inhibition of spontaneous electrical activity.. Next, we performed indirect immunofluorescence analyses to study the effect of primary myotube activity on the cellular localization of syndecan-4 and β1 integrin. Fig. 7 shows that syndecan-4 reactivity co-localizes with vinculin, a marker of costameric structures of differentiated myotubes, showing a striated pattern (left panels, top row). Images obtained at higher magnification confirm that syndecan-4 labeling co-localizes with vinculin in costameres corresponding to Z-lines. This labeling is localized between A bands defined by the reactivity for sarcomeric myosin (Fig. 7). By contrast, in paralyzed myotubes the reactivity to syndecan-4 is diffused and the striated pattern is lost, suggesting that the costameric organization is disrupted (Fig. 7, left panels, middle row). The same panels show that vinculin is also lost from costameres in TTX-treated myotubes, suggesting that these structures are disorganized in paralyzed cells. Vinculin protein levels evaluated in western blots are unmodified in TTX-treated cultures, suggesting that the inhibition of spontaneous electrical activity only affects the assembly of vinculin in costameres (data not shown). Similarly, β1 integrin is co-localized with vinculin in costameric structures in differentiated myotubes (Fig. 7, right panels, top row) and this distribution is lost in inactive myotubes (middle row). Fig. 7 also shows that the loss of localization of syndecan-4 and β1 integrin in costameric structures can be recovered in 48 h after the wash-out of TTX (w48). The assembly of costameric proteins in the striated pattern was not observed after 24 h after restoring spontaneous electrical activity, suggesting that a longer time is required for the reassembly of these molecular components on the cell surface. Altogether, these results suggest that the inhibition of electrical activity of myotubes as well as affecting protein levels also disrupts the costameric distribution of syndecan-4 and β1 integrin, thereby explaining the defect in the adhesion phenotype of inactive myotubes. 4. Discussion Our results demonstrate that syndecan-4 and β1 integrin, molecules involved in cell adhesion and migration, are regulated by electrical activity during the differentiation of rat fetal myoblasts. The inhibition of electrical activity strongly decreases, in a reversible manner, the levels of both proteins. Immunodetection analysis shows that syndecan-4 and β1 integrin are localized in costameric structures on differentiated myotubes and that this sub-cellular distribution is lost in paralyzed myotubes. These myotubes have a clear defective adhesion phenotype compared with control cultures. TTX treatment neither affects cell viability nor apoptosis. Altogether, these results strongly suggest that the inactivity-dependent-modifications in the amount and distribution of syndecan-4 and β1 integrin in skeletal muscle cells determine this defective adhesion phenotype. Although the regulation of muscle protein expression by electrical activity has been extensively documented as nAChR and myogenin and TGF-β receptor type I in the same cell system (Ugarte and Brandan, 2006), our experiments suggesting that syndecan-4 and β1 integrin are regulated by electrical activity cannot exclude that a downstream process, such as the calcium influx-triggered contractile activity of muscle cells, could participate in the regulation of syndecan-4 and β1 integrin. The amount of syndecan-4 core protein shows a biphasic behavior during primary myoblast differentiation, with high levels during the first days, decreasing in the next and elevating again from day 5 of differentiation. This temporal regulation was also observed in the amount of transcripts along the myoblast differentiation, suggesting a transcriptional regulation of the syndecan-4 gene during this process. This behavior can reflect the different physiological roles of syndecan4 during the process of myotube formation. In the myoblast stage,.
(7) G. Ugarte et al. / Matrix Biology 29 (2010) 383–392. 389. Fig. 5. β1 integrin protein levels are decreased in TTX-treated myotubes. A) Western blot for β1 integrin of total cell extracts corresponding to control and TTX-treated cultures at day 2, 3 and 4 of differentiation. GAPDH immunoblot is shown as a loading control. B) Western blot for β1 integrin of total cell extracts corresponding to control and TTX-treated cultures for 24 h (lanes 1 and 2) and TTX-treated cultures, washed out and lysed after 6, 12 and 24 h. The last two lanes (6 and 7) correspond to extracts of control cultures and TTX-treated for 48 h. As an internal control, the lower panel shows the western blot for TβRI of total cell extracts corresponding to the same samples described above. C) Western blot for phospho-Akt kinase corresponding to control cultures (day 5 of differentiation), TTX-treated cultures and TTX-treated cultures, washed out and lysed after 48 h (w48). The IGFdependent phosphorylation of Akt in C2C12 myoblasts is shown as a positive control. Myoblast cultures were starved for 4 h in DMEM and then incubated with IGF-1 (20 ng/ml) for 30 min. Lower panels show the immunodetection for total Akt corresponding to each sample. D) Left panel, corresponds to western blot for phospho-Akt kinase corresponding to control cultures (day 5 of differentiation), TTX-treated cultures and TTX-treated cultures, washed out and lysed after 48 h (w48), in the presence or absence of 5 μM SB 431542 and 1.0 ng/ml of TGF-β1 for 30 min. The right panel shows a western blot for p-Smad-2 with a GAPDH immunoblot shown as a loading control.. syndecan-4 can be important for proliferative growth factor-dependent signaling such as FGF-2, but at more differentiated states this HSPG can adopt a structural role in the costameric structures of myofibers. In addition, although the total pool of syndecan-4 can be totally recovered after 24 h by restoring electrical activity, the recovery of the surface pool takes longer. This result is consistent with the immunofluorescence analysis showing that the reassembly of syndecan-4 organization in costameres is only observed 48 h after restoring contractile activity. RT-PCR and northern assays showed that the amount of transcripts for syndecan-4 are unmodified in inactive myotubes, suggesting that the activity-dependent regulation of syndecan-4 is likely to occur after the transcriptional level. Several reports have shown that members of the syndecan superfamily can be cleaved from the plasma membrane by processes of shedding (Subramanian et al., 1997). It is possible to suggest that this phenomenon is involved in the regulation of surface syndecan-4 by electrical activity in skeletal myotubes. It is tempting to suggest two kinds of mechanisms of the regulation of syndecan-4 protein levels in muscle cells: one involved in long-term modifications of gene expression during the differentiation process and the second, shortterm regulation, as triggered by electrical inactivity, involving the. modification of the amount of mature molecules at the cell surface as shedding or endocytosis followed by degradation. It has been reported that satellite cells extracted from syndecan-4 null mice in culture have defects in cellular association and migration, the expression of MyoD and myosin heavy chain and differentiation. However, the phenotype of syndecan-4−/− adult muscle tissue is similar to wild type (Cornelison et al., 2004). Our results show that the inhibition of electrical activity of primary myotubes disrupts the costameric localization of syndecan-4 and β1 integrin. Because many different proteins are components of costameres (Ervasti, 2003), it cannot be excluded that in syndecan-4−/− mice a compensatory mechanism might occur with some of the molecules involved. This compensatory effect might not be developed in primary cultures, whereas the down-regulation of syndecan-4 and β1 integrin is induced by a short-term inhibition of spontaneous electrical activity. A compensatory mechanism of the proteoglycan decorin during skeletal muscle regeneration has been documented in biglycan null mice (Casar et al., 2004a,b). Previously, we have shown that decorin is also regulated by the presence of the motor nerve in rat skeletal muscles (Fadic et al., 1990). In this case, denervation as well as the in vivo blockage of the electrical.
(8) 390. G. Ugarte et al. / Matrix Biology 29 (2010) 383–392. Fig. 6. Cell adhesion is decreased in TTX-treated primary myotubes. A) The plot is generated from protein and DNA quantification of supernatants and cell fractions obtained after the incubation of control and TTX-treated cultures (day 4 of differentiation) in PBS-EDTA, and represents the percentage of cells detached from substrate after treatment. Data are expressed as the means ± SD for percentages calculated for the three assays made in parallel for control and inactive cultures. * Indicates P b 0.01. B) The cell viability was evaluated by the 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) reduction assay. Results correspond to the percentage of viable cells in the presence of TTX or control (gray) with respect to cells incubated with H2O2 (white). The bars correspond to the mean ± SE for three experiments each carried out in triplicate. * Indicates P b 0.02 between the treatment with TTX or vehicle with respect to H2O2. C) Left, a, myotubes (day 4 differentiation) untreated (vehicle); b, untreated myotubes incubated with H2O2; c, myotubes incubated with TTX; d, myotubes incubated with TTX and H2O2. In each case the myotubes were fixed and stained with Hoechst. Apoptotic nuclei were quantified in 15 fields for two independent preparations. A representative apoptotic nucleus is shown. Bar corresponds to 50 μm. Right, correspond to the percentage of apoptotic nuclei in the presence or absence of TTX (gray) or in the presence of H2O2 (white). The bars correspond to the mean ± SE for three experiments each carried out in triplicate. * Indicates P b 0.01 between the treatment with TTX or vehicle with respect to H2O2.. activity by TTX induced an increase of this ECM proteoglycan. In contrast with the regulation of syndecan-4, the increase of decorin was triggered at the transcriptional level (Brandan et al., 1992). It has been reported that β1 integrin can regulate adhesion through the PI3K/Akt signaling pathway (Wang et al., 2007). By contrast, PP2A is associated physically with β1 integrin in muscle cells under differentiation and regulates the phosphorylation state of. integrin β1A (Kim et al., 2004). Our results showing that TTX-treated myotubes contain Akt in a hyperphosphorylated state can be explained by the inactivity-triggered decrease in β1 integrin levels that could decrease PP2A phosphatase activity, resulting in higher levels of phosphorylated downstream targets such as Akt. PP2A is physically associated to Akt and is the main phosphatase involved in the regulation of this kinase in rat adipocytes (Resjo et al., 2002). TGF-.
(9) G. Ugarte et al. / Matrix Biology 29 (2010) 383–392. 391. Fig. 7. Syndecan-4 and β1 integrin reactivity co-localize with vinculin and is disrupted in TTX-treated myotubes. Left and right panels, indirect immunofluorescence images showing the sub-cellular localization of both syndecan-4 and β1 integrin with vinculin in control cultures at day 5 of differentiation. The left and right columns in each panel correspond to merged images matching to high magnification selected areas of corresponding images (scale bar = 2.5 μm). Middle rows in each panel, indirect immunofluorescence images showing the disrupted sub-cellular localization of both syndecan-4 and β1 integrin with vinculin in TTX-treated cultures at day 5 of differentiation. Lower rows in each panel, indirect immunofluorescence images of TTX-treated cultures for 2 days, washed out and fixed 48 h afterwards (w48), showing the recovery of the co-localization of both syndecan-4 and β1 integrin with vinculin. Scale bar = 10 μm. The inset shows indirect immunofluorescence images at high magnification: the sub-cellular co-localization of syndecan-4 and vinculin between the myosin heavy chain in a control myotube at day 4 of differentiation. Scale bar = 1.5 μm.. β responds through non-Smad dependent pathway signals through several kinase-activated pathways, including PI3K/Akt pathways (Zhang, 2009). Because we have shown that the amount of TGF-β1 receptors on the cell surface is modulated by electrical activity (Ugarte and Brandan, 2006), we inhibited TGF-β signaling by blocking TβRI kinase activity (Laping et al., 2002). Under this experimental condition we saw no change in the TTX-induced hyperphosphorylation of Akt. Immunolocalization analysis shows that in differentiated primary myotubes, syndecan-4 and β1 integrin reactivity are distributed in a striated pattern and co-localize with vinculin, suggesting that these molecules are localized in costameres. This structure has a crucial role in muscle physiology by forming a structural link that transmits the force generated in sarcomeres to the ECM and neighboring fibers. In deconvolution microscopy studies, a similar costameric distribution of syndecan-4 in rat cardiomyocytes has been reported (VanWinkle et al., 2002). β1 integrin has been detected in costameres and muscle– tendon junctions of adult skeletal muscle (Belkin et al., 1996). Our analysis of TTX-treated myotubes shows that the costameric organization is disrupted (determined by vinculin immunostaining), affecting syndecan-4 and β1 integrin localization. A decrease in protein levels of both molecules and the loss of costameric distribution correlate with lower cell adhesion capacity in inactive myotubes. In this context, electrical activity can act as a positive regulation factor for the expression of costameric transmembrane. molecules such as syndecan-4 and β1 integrin in myotubes under differentiation. This regulation can acquire relevance during the formation of primary and secondary fibers during fetal myogenesis as well as in regeneration processes of adult muscle when events of growth and fusion of new fibers and ECM remodeling take place. In fact, although syndecan-4 has been defined as a molecular marker of satellite cells in adult muscle tissue, syndecan-4 is transiently upregulated both during the regeneration of adult muscle and during limb myogenesis (Olguin and Brandan, 2001; Cornelison et al., 2004). Our results showing that syndecan-4 is down-regulated in denervated muscles support an additional role for this HSPG in the physiology of adult muscle fibers.. Acknowledgements The authors are indebted to Dr. Hugo Olguín (P. Universidad Católica de Chile) for critical reading of the manuscript, Dr. Alan Rapraeger (University of Wisconsin-Madison, USA) for providing the anti-syndecan-4 antibody and Dr. Daniel Goldman (University of Michigan, USA) for providing the RMT461 cell line. This study was supported by research grants from FONDAP-Biomedicine # 13980001, CARE PFB12/2007 and Muscular Dystrophy Association # 89419. The postdoctoral fellowship to G.U. was supported by Millenium Institute for Fundamental and Applied Biology (MIFAB) financed in part by.
(10) 392. G. Ugarte et al. / Matrix Biology 29 (2010) 383–392. Ministerio de Planificación y Cooperación (MIDEPLAN, Chile) and by FONDAP-Biomedicine. References Alvarez, A.R., Sandoval, P.C., Leal, N.R., Castro, P.U., Kosik, K.S., 2004. Activation of the neuronal c-Abl tyrosine kinase by amyloid-beta-peptide and reactive oxygen species. Neurobiol. Dis. 17, 326–336. Belkin, A.M., Zhidkova, N.I., Balzac, F., Altruda, F., Tomatis, D., Maier, A., Tarone, G., Koteliansky, V.E., Burridge, K., 1996. Beta 1D integrin displaces the beta 1A isoform in striated muscles: localization at junctional structures and signaling potential in nonmuscle cells. J. Cell. Biol. 132, 211–226. Brandan, E., Fuentes, M.E., Andrade, W., 1992. Decorin, a chondroitin/dermatan sulfate proteoglycan is under neural control in rat skeletal muscle. J. Neurosci. Res. 32, 51–59. Brandan, E., Carey, D.J., Larrain, J., Melo, F., Campos, A., 1996. Synthesis and processing of glypican during differentiation of skeletal muscle cells. Eur. J. Cell Biol. 71, 170–176. Brodie, C., Brody, M., Sampson, S.R., 1989. Characterization of the relation between sodium channels and electrical activity in cultured rat skeletal myotubes: regulatory aspects. Brain Res. 488, 186–194. Buckingham, M., Bajard, L., Chang, T., Daubas, P., Hadchouel, J., Meilhac, S., Montarras, D., Rocancourt, D., Relaix, F., 2003. The formation of skeletal muscle: from somite to limb. J. Anat. 202, 59–68. Cabello-Verrugio, C., Brandan, E., 2007. A novel modulatory mechanism of transforming growth factor-beta signaling through decorin and LRP-1. J. Biol. Chem. 282, 18842–18850. Casar, J.C., Cabello-Verrugio, C., Olguin, H., Aldunate, R., Inestrosa, N.C., Brandan, E., 2004a. Heparan sulfate proteoglycans are increased during skeletal muscle regeneration: requirement of syndecan-3 for successful fiber formation. J. Cell. Sci. 117, 73–84. Casar, J.C., McKechnie, B.A., Fallon, J.R., Young, M.F., Brandan, E., 2004b. Transient upregulation of biglycan during skeletal muscle regeneration: delayed fiber growth along with decorin increase in biglycan-deficient mice. Dev. Biol. 268, 358–371. Chahine, K.G., Walke, W., Goldman, D., 1992. A 102 base pair sequence of the nicotinic acetylcholine receptor delta-subunit gene confers regulation by muscle electrical activity. Development 115, 213–219. Chahine, K.G., Baracchini, E., Goldman, D., 1993. Coupling muscle electrical activity to gene expression via a cAMP-dependent second messenger system. J. Biol. Chem. 268, 2893–2898. Chomczynski, P., Sacchi, N., 1987. Single-step method of RNA isolation by acid guanidium thiocyanate–phenol–chloroform extraction. Anal. Biochem. 162, 156–159. Cognard, C., Constantin, B., Rivet-Bastide, M., Imbert, N., Besse, C., Raymond, G., 1993. Appearance and evolution of calcium currents and contraction during the early post-fusional stages of rat skeletal muscle cells developing in primary culture. Development 117, 1153–1161. Cornelison, D., Filla, M., Stanley, H., Rapraeger, A., Olwin, B., 2001. Syndecan-3 and syndecan-4 specifically mark skeletal muscle satellite cells and are implicated in satellite cell maintenance and muscle regeneration. Dev. Biol. 239, 79–94. Cornelison, D.D., Wilcox-Adelman, S.A., Goetinck, P.F., Rauvala, H., Rapraeger, A.C., Olwin, B.B., 2004. Essential and separable roles for Syndecan-3 and Syndecan-4 in skeletal muscle development and regeneration. Genes Dev. 18, 2231–2236. Dovas, A., Yoneda, A., Couchman, J.R., 2006. PKCbeta-dependent activation of RhoA by syndecan-4 during focal adhesion formation. J. Cell. Sci. 119, 2837–2846. Dutton, E.K., Simon, A.M., Burden, S.J., 1993. Electrical activity-dependent regulation of the acetylcholine receptor delta-subunit gene, MyoD, and myogenin in primary myotubes. Proc. Natl. Acad. Sci. U. S. A. 90, 2040–2044. Ervasti, J.M., 2003. Costameres: the Achilles' heel of Herculean muscle. J. Biol. Chem. 278, 13591–13594. Fadic, R., Brandan, E., Inestrosa, N.C., 1990. Motor nerve regulates muscle extracellular matrix proteoglycan expression. J. Neurosci. 10, 3516–3523. Fuentealba, L., Carey, D.J., Brandan, E., 1999. Antisense inhibition of syndecan-3 expression during skeletal muscle differentiation accelerates myogenesis through a basic fibroblast growth factor-dependent mechanism. J. Biol. Chem. 274, 37876–37884. Grouselle, M., Koenig, J., Lascombe, M.L., Chapron, J., Meleard, P., Georgescauld, D., 1991. Fura-2 imaging of spontaneous and electrically induced oscillations of intracellular free Ca2+ in rat myotubes. Pflugers Arch. 418, 40–50.. Gutierrez, J., Osses, N., Brandan, E., 2006. Changes in secreted and cell associated proteoglycan synthesis during conversion of myoblasts to osteoblasts in response to bone morphogenetic protein-2: role of decorin in cell response to BMP-2. J. Cell. Physiol. 206, 58–67. Horowitz, A., Tkachenko, E., Simons, M., 2002. Fibroblast growth factor-specific modulation of cellular response by syndecan-4. J. Cell. Biol. 157, 715–725. Hynes, R.O., 2002. Integrins: bidirectional, allosteric signaling machines. Cell 110, 673–687. Kim, S.M., Kwon, M.S., Park, C.S., Choi, K.R., Chun, J.S., Ahn, J., Song, W.K., 2004. Modulation of Thr phosphorylation of integrin beta1 during muscle differentiation. J. Biol. Chem. 279, 7082–7090. Laping, N.J., Grygielko, E., Mathur, A., Butter, S., Bomberger, J., Tweed, C., Martin, W., Fornwald, J., Lehr, R., Harling, J., Gaster, L., Callahan, J.F., Olson, B.A., 2002. Inhibition of transforming growth factor (TGF)-beta1-induced extracellular matrix with a novel inhibitor of the TGF-beta type I receptor kinase activity: SB-431542. Mol. Pharmacol. 62, 58–64. Larrain, J., Cizmeci-Smith, G., Troncoso, V., Stahl, R.C., Carey, D.J., Brandan, E., 1997. Syndecan-1 expression is down-regulated during myoblast terminal differentiation modulation by growth factors and retinoic acid. J. Biol. Chem. 272, 18418–18424. Macpherson, P., Kostrominova, T., Tang, H., Goldman, D., 2002. Protein kinase C and calcium/calmodulin-activated protein kinase II (CaMK II) suppress nicotinic acetylcholine receptor gene expression in mammalian muscle. A specific role for CaMK II in activity-dependent gene expression. J Biol Chem 277, 15638–15646. Mayer, U., 2003. Integrins: redundant or important players in skeletal muscle? J. Biol. Chem. 278, 14587–14590. Mayer, U., Saher, G., Fassler, R., Bornemann, A., Echtermeyer, F., von der Mark, H., Miosge, N., Poschl, E., von der Mark, K., 1997. Absence of integrin alpha 7 causes a novel form of muscular dystrophy. Nat. Genet. 17, 318–323. Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Methods. 65, 55–63. Mostafavi-Pour, Z., Askari, J.A., Parkinson, S.J., Parker, P.J., Ng, T.T., Humphries, M.J., 2003. Integrin-specific signaling pathways controlling focal adhesion formation and cell migration. J. Cell. Biol. 161, 155–167. Olguin, H., Brandan, E., 2001. Expression and localization of proteoglycans during limb myogenic activation. Dev. Dyn. 221, 106–115. Rapraeger, A.C., 2001. Molecular interactions of syndecans during development. Semin. Cell. Dev. Biol. 12, 107–116. Resjo, S., Goransson, O., Harndahl, L., Zolnierowicz, S., Manganiello, V., Degerman, E., 2002. Protein phosphatase 2A is the main phosphatase involved in the regulation of protein kinase B in rat adipocytes. Cell. Signal. 14, 231–238. Sanes, J.R., Lichtman, J.W., 1999. Development of the vertebrate neuromuscular junction. Annu. Rev. Neurosci. 22, 389–442. Saoncella, S., Echtermeyer, F., Denhez, F., Nowlen, J.K., Mosher, D.F., Robinson, S.D., Hynes, R.O., Goetinck, P.F., 1999. Syndecan-4 signals cooperatively with integrins in a Rho-dependent manner in the assembly of focal adhesions and actin stress fibers. Proc. Natl. Acad. Sci. U. S. A. 96, 2805–2810. Steinfeld, R., Van Den Berghe, H., David, G., 1996. Stimulation of fibroblast growth factor receptor-1 occupancy and signaling by cell surface-associated syndecans and glypican. J. Cell. Biol. 133, 405–416. Subramanian, S.V., Fitzgerald, M.L., Bernfield, M., 1997. Regulated shedding of syndecan-1 and -4 ectodomains by thrombin and growth factor receptor activation. J. Biol. Chem. 272, 14713–14720. Tsuzuki, S., Kojima, T., Katsumi, A., Yamazaki, T., Sugiura, I., Saito, H., 1997. Molecular cloning, genomic organization, promoter activity, and tissue-specific expression of the mouse ryudocan gene. J. Biochem. 122, 17–24. Ugarte, G., Brandan, E., 2006. Transforming growth factor beta (TGF-beta) signaling is regulated by electrical activity in skeletal muscle cells TGF-beta type I receptor is transcriptionally regulated by myotube excitability. J. Biol. Chem. 281, 18473–18481. VanWinkle, W.B., Snuggs, M.B., De Hostos, E.L., Buja, L.M., Woods, A., Couchman, J.R., 2002. Localization of the transmembrane proteoglycan syndecan-4 and its regulatory kinases in costameres of rat cardiomyocytes: a deconvolution microscopic study. Anat. Rec. 268, 38–46. Vial, C., Zuniga, L.M., Cabello-Verrugio, C., Canon, P., Fadic, R., Brandan, E., 2008. Skeletal muscle cells express the profibrotic cytokine connective tissue growth factor (CTGF/CCN2), which induces their dedifferentiation. J. Cell. Physiol. 215, 410–421. Wang, H.Q., Bai, L., Shen, B.R., Yan, Z.Q., Jiang, Z.L., 2007. Coculture with endothelial cells enhances vascular smooth muscle cell adhesion and spreading via activation of beta1-integrin and phosphatidylinositol 3-kinase/Akt. Eur. J. Cell Biol. 86, 51–62. Zhang, Y.E., 2009. Non-Smad pathways in TGF-beta signaling. Cell. Res. 19, 128–139..
(11)
Figure

Documento similar
In mammalian cells, the highly regulated expression of nuclear Myc family transcription factors (c-, N- and L-Myc) is closely tied to cell growth and proliferation as well
Thus, while SF significantly increases cell adhesion on the channel surface due to the supply by SF of additional binding motifs, still cell···cell binding is preferred
En cambio, nosotros mostramos que la inhibición de p38α resulta en la acumulación de ROS y activación de JNK, sensibilizando a las células tumorales a apoptosis
As expected for a protein with a crucial role in cell cycle control, p21 Cip1 expression is tightly regulated by several mechanisms, including p53-dependent and
Also, a very recent report demonstrates that the loss in self-renewal detected in satellite cells from aged mice can be overcome by p38α/β inhibition (Bernet and Rudnicki,
Mammalian embryonic development comprises a plethora of tightly regulated biological processes (spatiotemporal control of specific gene regulation, cell signalling, cell migration
Toll-like receptor-4 (TLR4) signaling augments chemokine- induced neutrophil migration by modulating cell surface expression of chemokine receptors. Overexpression of G
In this work, we have undertaken the study of SUZ12, a Polycomb group protein and the microRNAs (miRNA) expressed by the oncogenic Epstein Barr Virus (EBV) in
