Eco morphological effects of large wood in mountain basins, under different level of forest management
138
0
0
Texto completo
(2) Pontificia Universidad Católica de Chile Facultad de Agronomía e Ingeniería Forestal. Eco-morphological effects of large Wood in mountain basins, under different level of forest management. Fernando Javier Ugalde Pascual. Thesis to obtain the degree of. Doctor en Ciencias de la Agricultura. Santiago, Chile, December 2018.
(3) Thesis presented as part of the requirements for the degree of Doctor in Ciencias de la Agricultura, approved by the. Thesis Committee. _____________________ Dr. Luca Mao, Advisor. __________________ Dr. Pablo Becerra. ___________________ Dr. Andrés Iroumé. Santiago, December 2018.
(4) A Pamela, Por su amor y apoyo permanente.
(5) Acknowledgements This study was the thesis project of the author who wishes to thank the Comisión Nacional de Investigación Científica y Tecnológica (CONICYT), Chile, for the scholarship to pursue this Doctoral studies (2012-2016). This study was partially funded by the CONICYT project “Effect of native forest replacement by pine plantations on biodiversity and ecosystem processes of Andean riparian and riverine habitats in the south of Chile” USA 2012-0011(P.I. Luca Mao).. I wish to thank: . My Thesis Committee: Professors Luca Mao, Pablo Becerra and Doctor Andrés Iroumé, for their confidence in and support of this research;. . The Dirección de Investigación y Postgrado, especially Dr. Francisco Meza, Sandra Osorio, Susan Arenas, Claudia Gonzalez and Ximena Ortega;. . The members of Ecohydromorphology laboratory,. Joaquín Lobato,. Ricardo. Carrillo, Rodrigo Pérez, Claudio Godoy for their valuable help and friendship; . Claudio Gomez, Jose Donoso, Diego Silva, Ernesto Cobo, José Miguel Riesco, Francisco Novoa for their support and friendship in the laboratory and fieldwork ;. . My friends and colleagues Anita Paris, Bruna Garretón, Lina María Forero, Valeria Espinoza, Estefanía Torres, María José Sandoval and Nicole Sallaberry for their support and friendship during the development of this thesis;. . The collaboration of Forestal Arauco, Forestal Niblinto, Conaf and Universidad de Talca for letting us work on theirs fields.. . Maxi Ruiz from Fundo Picazo, Misael from Forestal Arauco, Professor at Los Queñes school, Director Minas del Prado school, Esteban Jara and family from Fundo Los Queñes, Miguel from Pejerreyes’s neighbords association,. . Shaw Lacy for all his help, Hector and Noemi Muñoz Jorquera from quebrada las cerezas’s basin and José Ignacio Covarrubias from Fundo Huaraculén..
(6) My ultimate gratitude goes to my wife Pamela, my daughter Ainhoa, my son Mikel, my parents Fernando and María Teresa and my grandparents Miguel, Carmen, and Elisa for all their support and love from this and other dimension.. Fernando.
(7) Contents Table of Contents. Chapter 1. General Introduction. 9. Chapter 2. The effects of replacing native forest on the quantity and impacts of in-channel pieces of large wood in Chilean streams. 30. Chapter 3. The ecomorphological effects of in-channel large wood on resident fishes in headwater streams of central Chile. 69. Chapter 4. Pine plantations as a potential management tool for controlling invasive trout: Aquatic conservation of headwaters in a multi-use landscape in Central Chile.. 100. Chapter 5. Final Remarks. 119. Appendices Appendix 1. Abstract of the poster at the Workshop Eco-hidro- morfología de los ríos Chilenos 2015. Eco-morphological effects of large woody debris in andean watersheds under different forest management. 123. Appendix 2. Abstract of the poster at the Third International Conference on WOOD IN WORLD RIVERS 2015, Large wood characteristics in native forest and pine plantation streams of the costal range of southern Chile. 124. Appendix 3 Abstract of the presentation at the XIV Congreso de la Sociedad Chilena de Limnología 2017, Los efectos de reemplazar bosques nativos en la cantidad de piezas de madera e impactos en el canal en esteros chilenos. 125. Appendix 4 Abstract accepted for the presentation at the Fourth Conference on Wood in World Rivers 2019. 127. Appendix 5 Map from Las Arañas basin. 128. Appendix 6 Map from Los Cerezos basin. 129. Appendix 7 Map from Los Queñes basin. 130.
(8) Appendix 8 Map from Las Cabras basin. 131. Appendix 9 Map from Manqui basin. 132. Appendix 10 Map from Mela basin. 133. Appendix 11 Map from Quebrada Piuquenes basin. 134. Appendix 12 Map from Potrerillos basin. 135. Appendix 13 Map from Sin Puerta basin. 136. Appendix 14 Map from El Duende basin. 137. Appendix 15 Map from El Milico basin. 138.
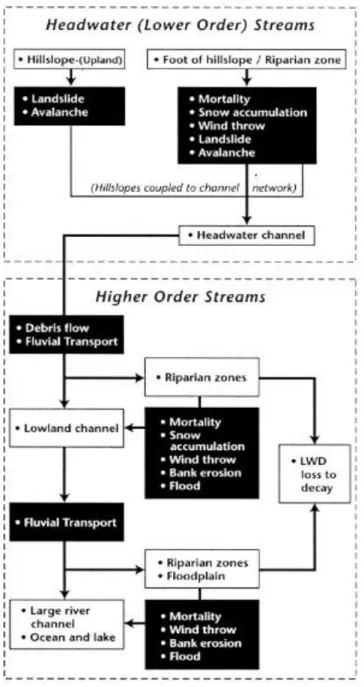
(9) General Introduction Headwater streams. Their morphology and ecology The headwaters of mountain basins are characterized by small drainage areas, steeper slopes, thicker sediment sizes and many non-moving rocks that create waterfalls (Comiti et al., 2007), as seen in figure 1. Headwater reaches exert a strong influence on channel configuration and flow dynamics, creating zones from where the sediments is retained in the channel bed, or in the floodplain and delivered downstream (Meyer, et al., 2007). In these segments, the ex-situ organic matter and nutrients are stored, processed (through physical and biological pathways) and lately supplied downstream. The retention and processing are more pronounced when the structuring of the channels reduces the speed of the flow, improving the hyporreic exchange (Findlay, 1995; Eggert and Wallace, 2007; Stofleth et al., 2008). The "River Continuum Concept" (Vannote et al., 1980), predicts that the functional structure of the macroinvertebrate communities in each section of the river is related to the nature and availability of food resources. In this concept the spatial changes in the functional structure of the macroinvertebrate communities along the head to mouth gradient of a river, respond to changes in the availability of resources. This concept is adjusted with special emphasis on streams of Basque Country (Larrañaga et al., 2009) which have similar climatic conditions to central-south of Chile. The fish fauna of a river is affected directly by the availability of food and by the morphology of the channel, a direct consequence of the availability and organization of large woody on it. Few studies in Chile have been carried out in terms of distribution and abundance of native and exotic fish, but there are reports of areas little human intervention by man (Habit et al., 2003). 9.
(10) The ecological properties of a headwater stream strongly depend on its morphological pattern and morphological changes which changes due to natural and anthropogenic disturbances. From this point of view the fluvial style and its dynamics strongly depends on the type and size of the riparian vegetation (Hupp & Osterkamp, 1996), which influences the cohesion of the banks, resulting in a reduction in erosion and the geomorphic effect of living trees continues even after they reach the channel by different pathways (see figure 2), or bankfull area from the banks or the portion of basin directly connected to the river network, and their transportation within the fluvial network (see figure 3). Dead or living pieces of large wood (LW) (tree, log, branch or root) can exert a tremendous influence on river erosion and sedimentation processes (Jeffries et al., 2003), channel morphology (Abbe & Montgomery, 2003), and channel hydraulics (e.g. Wallerstein et al., 2001). They also regulate water temperature, turbulence, and nutrient fluxes (Van Der Nat et al., 2003).. Riparian vegetation and large wood in headwater streams In-channel logs can accumulate in so-called logjams, and even valley jams which can strongly modify stream channel morphology and hydrology (Montgomery et al., 1995; Manners et al., 2007; Wohl et al., 2011) by the creation of steps, downstream and upstream pools which slow down the speed of the water, increasing the time of contact with the sediments and hyporreic exchange. They also increase the retention of sediment and organic matter (leaves), and small wood pieces. Log jams create biogeochemical hotspots, increasing the river's metabolism and biomass (Warren et al., 2007). LW and its grouped formations has also been found to play a key role in the ecological diversity of river channels providing habitat for a range of fish, macroinvertebrates and vertebrates of riverine fauna (Gurnell et al., 2002) as seen in figure 4. 10.
(11) Consequently, large wood accumulations indirectly alter the dynamics of river populations and communities of streams, increasing abundance, diversity and species composition (Wallace et al., 1995). The organic matter from riparian vegetation is determined by the structure and complexity of streams. It presents a higher energetic input to streams as well as important source of food for macroinvertebrates in headwater streams. Furthermore logjams increase retention of carbon and nitrogen and the amount of stream biogeochemical processing that occurs in situ, thus shortening the spatial scale of carbon and nutrient processing and stream metabolism. The organic matter stored (CPOM) is the food base for many primary consumers, which generates a higher total productivity. In addition, log jams increase the area available for the colonization of macroinvertebrates and provide refuge for fish, especially in droughts. The increase in the production of macroinvertebrates influences the higher trophic levels of trophic chains since they are an important part of the diet of fish, which increases their growth rates and condition factors for fish (Sigourney et al., 2006; Habit et al., 2007). In that sense, changes in the distribution of log jams could alter the nutrient cycle, the metabolism and the production of headwater streams. Several natural phenomenon and processes influence the abundance of LW and the frequency of logjams in the channel over time. Also human activities such as forest plantations, road construction and the cleaning of large wood from the channel lead to a reduction in size and frequency of log jams in the headwaters (Wohl, 2006). Generally it requires a long time to return to the original levels because the wood is gradually recruited into the channel, consequently the loss of functionality. It is known that the geomorphic processes of rivers and their conditions of dynamic equilibrium promote the diversity of habitats and functionality of aquatic and riparian ecosystems. Therefore,. 11.
(12) there is a need to understand these geomorphological processes responsible for the functioning of the river.. Large wood studies in Latin America and Chile In Latin America there is little specific information about in-channel LW abundance and its morphological and ecological effects, as well as studies of the ecology and biodiversity of stream headwater sections under forest management. In Chile, despite the importance of riparian and riverine habitats for water production and biodiversity conservation, the study on ecology and biodiversity of these habitats are available, but still relatively limited. There are studies that describe the large wood abundance and stream morphology in temperate basins (Andreoli et al., 2007c; Comiti et al., 2008) and in Andean basins and in Patagonia (Mao et al., 2008a). In addition, a study in a third order basin located in the southern Chilean coastal range, which reports the abundance of large wood, their longitudinal distribution and mobilization (Iroumé, et al., 2010; Iroumé et al., 2015, 2017). Some studies suggest that the dimensions of woody debris, its storage and morphological effects vary depending on the history of forest disturbances (Andreoli et al., 2007b; Comiti et al., 2008). Other studies suggest that this replacement modifies different abiotic variables and the water flow along the streams (Iroumé and Huber, 2002; Iroumé et al., 2006; Huber, 2008; Huber et al., 2010). Few studies have compared how the fauna of macroinvertebrates varies between watersheds that present forest management (Figueroa et al., 2003; Mancilla et al., 2009) reporting generally a. 12.
(13) lower animal diversity and a greater richness of species in basins lacking such management. However, no study has examined holistically the effect of forest management on the ecomorphology of Andean watersheds. Other studies have recognized the importance of small fragments, generally located along canals within pine plantations, for the conservation of insects and vertebrates (Grez et al, 2005). Therefore, riparian habitats in small fragments within watersheds with forest management should maintain a low diversity of native species compared to riparian habitats located in basins covered only with native forest. It is thus necessary to investigate further to understand the complex dynamics of woody debris, its effects on the morphology of the channel and the ecology of the river in basins that present native forest substitution.. Large wood studies in basins with forest plantations Logjams create biogeochemical hotspots in headwater streams, increasing stream metabolism and animal production. Also, logjams enhance overbank flooding and sedimentation and can store sediments and form pools. The abundance of wood loads and logjams frequency in mountain streams fluctuate through time due to natural recruitment processes (e.g., wildfire, insect infestation, debris flow,) and human disturbances (e.g., extensive timber clear-cuttings, extensive forest plantations). Streams draining basins impacted by human activities usually exhibit low wood load, which require very long time to return to original levels. The effect of substitution of native forests by exotic forestry plantations on different organisms and ecological processes has been largely studied (e.g. Arévalo and Fernández-Palacios, 2005) mainly reporting reductions in biodiversity within plantations depending on the management and silvicultural methods. In Chile (Mancilla et al., 2009a, Vera et al., 2014 ), some studies suggest that native 13.
(14) forest replacement by pine plantations modify different abiotic variables and hydrology (Iroumé et al., 2006), and some works specifically focused on LW suggest that wood dimensions, storage and morphological effects vary considerably as a function of forest disturbance history (Comiti et al., 2008; Ulloa et al., 2011). However, more research is needed to understand the complexity of large wood, recruitment and mobilization processes, and the effects on river channel morphology, and ecology in Chilean basins as consequences of replacement of native forest by forestry plantations. This study reports on a comparison between two streams draining basins of similar size but different land use (i.e. native forest vs. pine planation). Before extensive clearcut and the intensification of agriculture, the Andean basins in the central-southern zone of Chile would have been covered by native forests (Gajardo, 1994). Exotic tree plantation, agricultural activities and urban expansion have changed this landscape especially over the last century, and some basins are partially or completely occupied by exotic species, especially Pinus radiata (San Martín and Donoso 1996; Bustamante and Castor 1998; Grez, et al., 1998). In order to carry out forest plantations and management activities in Chilean watersheds, the law determines the preservation of a certain width of riparian forest (buffer zone) on each side of the riverbanks to avoid the loss of these original ecosystems as well as serious impacts on the ecology, the functionality and the stream biodiversity of fluvial habitats (Curran & Wohl, 2003; Iroumé et al., 2006). In general, the impacts of forest operation and land use change on the ecological functionality of river systems have not been extensively reported or investigated. Also, the effectiveness of this legal requirement regarding the conservation of fluvial and riparian habitats should probably be investigated and revised. One of the potential impacts of forest. 14.
(15) management in riverine environments is on the abundance of large wood pieces (i.e. trunks, logs, and coarse wood particles) in the channel, riverbed and banks, and the degree of organization of them as jams. Indeed, even if there is increasingly body of evidence from relatively unimpacted basin, scarce attention has been devoted to basins with exotic plantations and frequent clearcuttings. In Chile, invasion ecology of Pinus radiata has been studied in several opportunities, either regarding factors affecting invasion (Bustamante & Simonetti, 2005; Becerra & Bustamante, 2009, 2011) or consequences of this invasion on native species (Becerra & Montenegro, 2013). It appears that the interiors of undisturbed fragments are resistant to invasion with abiotic conditions posing constraints to seedling establishment (Bustamante et al, 2003; Bustamante and Simonetti, 2005; Becerra and Bustamante, 2011). However, in Chile no study has assessed the magnitude of invasion by this tree in riparian habitats of small fragments within, nor the impacts of this invasion. Given the closeness of riparian habitats located in small fragments to pine plantations, invasion by P. radiata should be common and have different types of impacts in ecology and biodiversity of these habitats. Similarly, only some few studies have compared fauna and animal ecology between pine plantations and native forests (Estades and Temple, 1999; Grez et al, 2003; Vergara and Simonetti, 2004; Saavedra and Simonetti, 2005; Gonzalez et al, 2006; Becerra and Simonetti, 2013), usually documenting lower animal native species diversity and higher richness of exotic animals within plantations. On the other hand, some studies have assessed the effect of forest fragmentation on animal diversity (Estades & Temple, 1999; Grez et al, 2003; Donoso et al, 2003; Vergara & Simonetti, 2004; Saavedra & Simonetti, 2005; Gonzalez et al, 2006; Kelt, 2006; Jaña et al, 2006), frequently reporting reductions in animal diversity and abundance in fragments compared to continuous forests. However, no study 15.
(16) has examined the effect of replacement of native forest by pine plantations on animal ecology in riparian habitats of remnant fragments within pine plantations. Nevertheless, other studies have acknowledged the importance of small fragments, generally located along streams within pine plantations, for conservation of insects and vertebrates (Smith-Ramírez, 2004; Grez et al, 2005). Therefore, riparian habitats in small fragments within pine plantations should hold lower native species diversity than riparian habitats located within basins covered with only native forests. Although the effect of forest management on the different organisms and ecological processes has been studied, this impact has not been evaluated on an eco-morphological scale in gravelbed headwater streams. Studies have mainly reported the reduction of macroinvertebrates biodiversity within the plantations of Pinus radiata D. Don. (Mancilla et al., 2009a) and reduction of its behavior (Correa-Araneda et al., 2017). The investigation focus on the differences between basins covered by native forest and where native forest has been replaced by pine plantations in an Andean and Coastal region of the south of Chile, between the 7th and 8th region, where the native forest substitution by pine plantations has mostly occurred. Specifically focused on the amount and degree of accumulation of log pieces, the LW morphological effects in terms of number and dimension of pools, channel width, and morphological habitat diversity. Then I focus the study in the consequences for different aquatic organism-types.. Hypothesis and objectives of the thesis The specific hypotheses addressed in this thesis is that the replacement of native forests by exotic plantations at the basin scale would cause a sensitive change in amount and degree of. 16.
(17) organization of logjams along the river network, with consequent change in the dynamic of wood recruitment and displacement along the river network, with a change of the retention of organic matter, animal production, and in situ biogeochemical processes. The general objective of this thesis is to study and increase the knowledge of the consequences of Pinus radiata management on the abundancy, dynamics and functionality of LW and its effect on the aquatic organisms and. biodiversity on headwater streams in an eco-morphological. perspective. 1. The first specific objective of this thesis is to assess the differences on in-stream large wood (volumes and characteristics) and related eco-morphological effects in streams draining catchments covered by native and Pine plantation forests. 2.. The second specific objective of this thesis is to verify the extent to which the amount of LW in a reach in headwater streams affects its morphology, organic matter retention, macroinvertebrate diversity, and how these then affect fish population.. 3. The third specific objective of this thesis is to assess differences in fish condition and abundance of native versus non-native fish species among headwater systems in Central Chile between catchments dominated by pine plantations and native forests.. List of references ABBE, T., G. PESS, D.R. MONTGOMERY & K.L. FETHERSTON 2003. Integrating Engineered Log Jam Technology into River Rehabilitation. In: Restoration of Puget Sound Rivers. 443–482. ABBE, T.B.T.B. and D.R.M.T.B. & D.R. MONTGOMERY 2003. Patterns and processes of wood debris accumulation in the Queets river basin, Washington. Geomorphology 51 (1–3): 81–107.. 17.
(18) ANDREOLI, A., G. CARLIG, F. COMITI & A. IROUMÉ 2007a. Residuos leñosos de gran tamaño en un torrente de la Cordillera de Los Andes , Chile: su funcionalidad e importancia. 28 (2): 83–96. ANDREOLI, A., F. COMITI & M.A. LENZI 2007b. Characteristics , distribution and geomorphic role of large woody debris in a mountain stream of the Chilean Andes. 1692: 1675–1692. ANDREOLI, A., F. COMITI, L. MAO, M.A. LENZI, A. URCIOLO, A.I. ARRAU & L.O. URRUTIA 2007c. Geomorphic Effect of Wood in Southern Andes Streams : Evidences From Three Mountain Basins. 510735pp. ANTÓN, A., A. ELOSEGI, L. GARCÍA-ARBERAS, J. DÍEZ & A. RALLO 2011. Restoration of dead wood in Basque stream channels: effects on brown trout population. Ecology of Freshwater Fish 20 (3): 461–471. ARÉVALO, J.R. & J.M. FERNÁNDEZ-PALACIOS 2005. Gradient analysis of exotic Pinus radiata plantations and potential restoration of natural vegetation in Tenerife, Canary Islands (Spain). Acta Oecologica. ARISMENDI, I., B. PENALUNA & D. SOTO 2011. Body condition indices as a rapid assessment of the abundance of introduced salmonids in oligotrophic lakes of southern Chile. Lake and Reservoir Management 27 (1): 61–69. ARISMENDI, I., D. SOTO, B. PENALUNA, C. JARA, C. LEAL & J. LEÓN-MUÑOZ 2009. Aquaculture, non-native salmonid invasions and associated declines of native fishes in Northern Patagonian lakes. Freshwater Biology 54 (5): 1135–1147. ARRATIA F., G. 1983. Preferencias de habitat de peces siluriformes de aguas continentales de Chile (Fam. Diplomystidae y Trichomycteridae). Studies on Neotropical Fauna and Environment 18 (4): 217–237. BECERRA, P.I. & R.O. BUSTAMANTE 2009. The effect of herbivory on seedling survival of the invasive exotic species Pinus radiata and Eucalyptus globulus in a Mediterranean ecosystem of Central Chile. Forest Ecology and Management. BECERRA, P.I. & R.O. BUSTAMANTE 2011. Effect of a native tree on seedling establishment of two exotic invasive species in a semiarid ecosystem. Biological Invasions. BECERRA, P.I. & G. MONTENEGRO 2013. The widely invasive tree Pinus radiata facilitates regeneration of native woody species in a semi-arid ecosystem. Applied Vegetation Science. BECERRA, P.I. & J. a SIMONETTI 2013. Patterns of exotic species richness of different taxonomic groups in a fragmented landscape of central Chile. Bosque. BELLETTI, B., M. RINALDI, M. BUSSETTINI, F. COMITI, A.M. GURNELL, L. MAO, L. NARDI & P. VEZZA 2017. Characterising physical habitats and fluvial hydromorphology: A new system for the survey and classification of river geomorphic units. Geomorphology 283 (February): 143–157. BUSTAMANTE, R.O. & J.A. SIMONETTI 2005. Is Pinus radiata invading the native vegetation in Central Chile? Demographic responses in a fragmented forest. Biological Invasions.. 18.
(19) CID, N., N. BONADA, S. CARLSON, T. GRANTHAM, A. GASITH & V. RESH 2017. High Variability Is a Defining Component of Mediterranean-Climate Rivers and Their Biota. Water 9 (1): 52. COMITI, F., A. ANDREOLI, M.A. LENZI & L. MAO 2006. Spatial density and characteristics of woody debris in five mountain rivers of the Dolomites (Italian Alps). Geomorphology 78 (1–2): 44–63. COMITI, F., A. ANDREOLI, L. MAO & M.A. LENZI 2008. Wood storage in three mountain streams of the Southern Andes and its hydro-morphological effects. Earth 262 (July 2007): 244–262. COMITI, F., L. MAO, M.A. LENZI & M. SILIGARDI 2009. Artificial steps to stabilize mountain rivers: a post-project ecological assessment. River Research and Applications 25 (5): 639–659. COMITI, F., L. MAO, A. WILCOX, E.E. WOHL & M.A. LENZI 2007. Field-derived relationships for flow velocity and resistance in high-gradient streams. Journal of Hydrology 340 (1–2): 48–62. CORDOVA, J.M., E.J. ROSI-MARSHALL, A.M. YAMAMURO & G.A. LAMBERTI 2007. Quantity, controls and functions of large woody debris in Midwestern USA streams. River Res. Appl 23: 21–33. CORREA-ARANEDA, F., A. BASAGUREN, R.T. ABDALA-DÍAZ, A.M. TONIN & L. BOYERO 2017. Resource-allocation tradeoffs in caddisflies facing multiple stressors. Ecology and Evolution (May): 1–8. CURRAN, J.H.J.H.J. & E.E.E.E.E. WOHL 2003. Large woody debris and flow resistance in step-pool channels, Cascade Range, Washington. Geomorphology 51 (1–3): 141–157. DAVIDSON, S.L. & B.C. EATON 2013. Modeling channel morphodynamic response to variations in large wood: Implications for stream rehabilitation in degraded watersheds. Geomorphology 202. DÍEZ, J.R., S. LARRAÑAGA, A. ELOSEGI & J. POZO 2000. Effect of removal of wood on streambed stability and retention of organic matter. Journal of the North American Benthological Society 19 (4): 621–632. DOMÍNGUEZ, E. & H.R. FERNÁNDEZ 2009. Fundación Miguel Lillo, Tucumán Macroinvertebrados bentónicos sudamericanos: Sistemática y biología. EGGERT, S.L. & J.B. WALLACE 2007. Wood biofilm as a food resource for stream detritivores. Limnology and Oceanography. ELOSEGI, A., J. DÍEZ, J. POZO & J. WILEY 2007. Contribution of dead wood to the carbon flux in forested streams. Earth Surface Processes and Landforms 1228 (1978): 1219–1228. ELOSEGI, A., J.R. DÍEZ, L. FLORES & J. MOLINERO 2017. Pools, channel form, and sediment storage in wood-restored streams: Potential effects on downstream reservoirs. Geomorphology 279. ELOSEGI, A., L. FLORES, D. JOSERRA, F. SCIENCE, B. COUNTRY & P.O. BOX 2011. The. 19.
(20) importance of local processes on river habitat characteristics: A Basque stream case study. Limnetica 30 (2): 183–196. ELOSEGI, A. & S. SABATER 2009. Conceptos y técnicas en ecología fluvial. ESTADES, C.F. & S.A. TEMPLE 1999. Deciduous-forest bird communities in a fragmented landscape dominated by exotic pine plantations. Ecological Applications. FAUSCH, K.D. & T.G. NORTHCOTE 1992. Large Woody Debris and Salmonid Habitat in a Small Coastal British Columbia Stream. Canadian Journal of Fisheries and Aquatic Sciences 49 (4): 682–693. FAUSTINI, J.M., J.A. JONES & J.A. FAUSTINI, J.M., JONES 2003. Influence of large woody debris on channel morphology and dynamics in steep, boulder-rich mountain streams, western Cascades, Oregon. Geomorphology 51 (1–3): 187–205. FERNÁNDEZ, H.R. & E. DOMÍNGUEZ 2001. Guía para la determinación de Artrópodos Bentónicos Sudamericanos, Universidad de Tucumán, Tucumán. FIGUEROA, R., C. VALDOVINOS, E. ARAYA & O. PARRA 2003. Macroinvertebrados bentónicos como indicadores de calidad de agua de ríos del sur de Chile Benthic macroinvertebrates as indicators of water quality of southern Chile rivers. Revista chilena de historia natural 76: 275–285. FLINT, J.O., R.W. HOLZENTHAL & S.C. HARRIS 1999. Catalog of the Neotropical Caddisflies (Insecta:Trichoptera). A special Publication of the Ohio Biological Survey: 239. FLORES, L., J.R. DÍEZ, A. LARRAÑAGA, C. PASCOAL & A. ELOSEGI 2013. Effects of retention site on breakdown of organic matter in a mountain stream. Freshwater Biology doi:10.111. FLORES, L., A. LARRAÑAGA, J. DÍEZ & A. ELOSEGI 2011. Experimental wood addition in streams: effects on organic matter storage and breakdown. Freshwater Biology 56 (10): 2156–2167. GAJARDO, R. 1994. La Vegetación Natural de Chile. Clasificación y Distribución Geográfica. Editorial Universitaria: Sanatiago (Chile). GEORGE ROBISON, E. & R.L. BESCHTA 1990. Coarse woody debris and channel morphology interactions for undisturbed streams in southeast Alaska, U.S.A. Earth Surface Processes and Landforms 15 (2): 149–156. GURNELL, A.M., H. PIEGAY, F.J. SWANSON & S. V GREGORY 2002. Large wood and fluvial processes. Freshwater Biology. GUSTAFSSON, P., L. a. GREENBERG & E. BERGMAN 2012. The influence of large wood on brown trout (Salmo trutta) behaviour and surface foraging. Freshwater Biology 57 (5): 1050–1059. HABIT, E., M.C. BELK & O. PARRA 2007. Response of the riverine fish community to the construction and operation of a diversion hydropower plant in central Chile. Aquatic Conservation: Marine and Freshwater Ecosystems 17 (1): 37–49. HABIT, E., P. PIEDRA, D.E. RUZZANTE, S.J. WALDE, M.C. BELK, V.E. CUSSAC, J.. 20.
(21) GONZALEZ & N. COLIN 2010. Changes in the distribution of native fishes in response to introduced species and other anthropogenic effects. Global Ecology and Biogeography 19 (5): 697–710. HABIT, E., P. VICTORIANO & A. RODRÍGUEZ-RUIZ 2003. Variaciones espacio-temporales del ensamble de peces de un sistema fluvial de bajo orden del centro-sur de Chile. Revista chilena de historia natural 76 (1): 3–14. HASSAN, M.A., D.L. HOGAN, S.A. BIRD, C.L. MAY, T. GOMI & D. CAMPBELL 2006. SPATIAL AND TEMPORAL DYNAMICS OF WOOD IN HEADWATER STREAMS OF THE PACIFIC NORTHWEST 1. 2. HE, Z., W. WU, F. DOUGLAS SHIELDS & F.D.S. JR 2009. Numerical analysis of effects of large wood structures on channel morphology and fish habitat suitability in a Southern US sandy creek. Ecohydrology 2 (3): 370–380. HILDERBRAND, R.H., A.D. LEMLY, C.A. DOLLOFF & K.L. HARPSTER 1997. Effects of large woody debris placement on stream channels and benthic macroinvertebrates. Canadian Journal of Fisheries and Aquatic Sciences 54 (4): 931–939. HUBER, A. 2008. Effect of Pinus radiata plantations on water balance in Chile. 148 (May 2007): 142–148. HUPP, C.R. & W.R. OSTERKAMP 1996. Riparian vegetation and fluvial geomorphic processes. Geomorphology. ILLIES, J. 1963. Revision der Sudamericanischen Gripopterygidae (Plecoptera). Mitt. Schweiz. ent. Ges., Lausanne 36 (3): 135–248. IROUMÉ A, ANDREOLI A, COMITI F, ULLOA H, H.A., A. IROUMÉ, A. ANDREOLI, F. COMITI, H. ULLOA & A. HUBER 2010. Large wood abundance, distribution and mobilization in a third order Coastal mountain range river system, southern Chile. Forest Ecology and Management 260 (4): 480–490. IROUMÉ, A. & A. HUBER 2002. Comparison of interception losses in a broadleaved native forest and a Pseudotsuga menziesii (Douglas fir) plantation in the Andes Mountains of southern Chile. Hydrological Processes. IROUMÉ, A., L. MAO, A. ANDREOLI, H. ULLOA & M.P. ARDILES 2015. Large wood mobility processes in low-order Chilean river channels. Geomorphology 228. IROUMÉ, A., O. MAYEN & A. HUBER 2006. Runoff and peak flow responses to timber harvest and forest age in southern Chile. Hydrological Processes 20 (1): 37–50. IROUMÉ, A., V. RUIZ-VILLANUEVA & L. PICCO 2017. Breakdown of instream wood in low order forested streams of the Southern Chilean mountain ranges. Forest Ecology and Management 401: 17–32. JEFFRIES, R., S.E. DARBY & D.A. SEAR 2003. The influence of vegetation and organic debris on flood-plain sediment dynamics: Case study of a low-order stream in the New Forest, England. Geomorphology 51 (1–3): 61–80. JOCHNER, M., J.M. TUROWSKI, A. BADOUX, M. STOFFEL & C. RICKLI 2015. The role of. 21.
(22) log jams and exceptional flood events in mobilizing coarse particulate organic matter in a steep headwater stream. Earth Surface Dynamics 3 (3). KAIL, J. 2003. Influence of large woody debris on the morphology of six central European streams. Geomorphology 51 (1–3): 207–223. KLEIN, R., R. SONNEVIL & D. SHORT 1987. Effects of woody debris removal on sediment storage in a northwest California stream. Erosion and Sedimentation in the Pacific Rim (165): 403–404. LARRAÑAGA, A., A. BASAGUREN & J. POZO 2009. Impacts of Eucalyptus globulus Plantations on Physiology and Population Densities of Invertebrates Inhabiting Iberian Atlantic Streams. International Review of Hydrobiology 94 (4): 497–511. LARRAÑAGA, S., J.R. DÍEZ, A. ELOSEGI & J. POZO 2003. Leaf retention in streams of the Agüera basin (northern Spain). Aquatic Sciences 65 (2): 158–166. LE LAY, Y.-F., H. PIÉGAY & B. MOULIN 2013. Wood Entrance, Deposition, Transfer and Effects on Fluvial Forms and Processes: Problem Statements and Challenging Issues. In: Treatise on Geomorphology. 20–36. Elsevier. LIBORIUSSEN, L., E. JEPPESEN, M.E. BRAMM & M.F. LASSEN 2005. Periphytonmacroinvertebrate interactions in light and fish manipulated enclosures in a clear and a turbid shallow lake. Aquatic Ecology 39 (1–2): 23–39. LINK, O. & E.M. HABIT 2014. Requirements and boundary conditions for fish passes of nonsport fish species based on Chilean experiences. Reviews in Environmental Science and Biotechnology 14 (1): 9–21. MANCILLA ESCOBAR, G., E. ESPINOZA SILVA, Ó. FERNÁNDEZ PALACIOS, M. LÓPEZ MUÑOZ & M. MIRANDA ALMENDRA 2012. Evaluación de las alteraciones hidromorfológicas en los cuerpos de agua que inciden sobre el estado ecológico del recurso hídrico en Chile. Santiago MANCILLA, G., C. VALDOVINOS, M. AZOCAR, P. JORQUERA & R. FIGUEROA 2009a. Efecto del reemplazo de la vegetación nativa de ribera sobre la comunidad de macroinvertebrados bentónicos en arroyos de climas templados, Chile central. Hidrobiologica 19 (3): 193–203. MANCILLA, G., C. VALDOVINOS, M. AZOCAR, P. JORQUERA & R. FIGUEROA 2009b. Replacement effect of riparian native vegetation on benthic macroinvertebrates community in temperate climate streams, Central Chile. Hidrobiologica 19 (3): 193–203. MANNERS, R.B., M.W. DOYLE & M.J. SMALL 2007. Structure and hydraulics of natural woody debris jams. Water Resources Research. MAO, L., A. ANDREOLI, F. COMITI & M.A. LENZI 2008a. Geomorphic effects of large wood jams on a sub-antarctic mountain stream. River Research and Applications 24 (3): 249–266. MAO, L., S. BURNS, F. COMITI, A. ANDREOLI & A. URCIUOLO 2008b. Acumulaciones de detritos leñosos en un cauce de montaña de Tierra del Fuego : análisis de la movilidad y de los efectos hidromorfológicos LW jams in a mountain stream of Tierra del Fuego : piece movement and hydro-morphological effects analysis. 29 (3): 197–211. 22.
(23) MAO, L., F. UGALDE, A. IROUME & S.N. LACY 2017. The Effects of Replacing Native Forest on the Quantity and Impacts of In-Channel Pieces of Large Wood in Chilean Streams. River Research and Applications 33 (1): 73–88. MARTIN, D.J., R.T. PAVLOWSKY & C.P. HARDEN 2016. Reach-scale characterization of large woody debris in a low-gradient, Midwestern U.S.A. river system. Geomorphology 262. MERRIT, R.W. AND CUMMINS, K.W. 1996. Trophic relations of macroinvertebrates. In: Methods in Stream Ecology. 453–473. Hahuer, R.F. and Lamberti. San Diego, CA. MEYER, J.L., D.L. STRAYER, J.B. WALLACE, S.L. EGGERT, G.S. HELFMAN, N.E. LEONARD & and N.E.L. MEYER, J. L., D. L. STRAYER, J. B. WALLACE, S. L. EGGERT, G. S. HELFMAN 2007. The contribution of headwater streams to biodiversity in river networks. Meyer, J. L., D. L. Strayer, J. B. Wallace, S. L. Eggert, G. S. Helfman, and N. E. Leonard. 43. MINTEGUI AGUIRRE, J., M. LENZI, J.R. SÁNCHEZ & L. MAO 2006. Movilización versus estabilización de los sedimentos en los cursos sometidos a la dinámica torrencial. MONTGOMERY, D.R., BUFFINGTON, J.M. 1997. Channel-reach morphology in mountain drainage basins. Geol. Soc. Am. Bull 109 (5): 596–611. MONTGOMERY, D.R., J.M. BUFFINGTON, R.D. SMITH, K.M. SCHMIDT & G. PESS 1995. Pool Spacing in Forest Channels. Water Resources Research 31 (4): 1097–1105. MOUQUET, N. & M. LOREAU 2003. Community Patterns in Source‐Sink Metacommunities. The American Naturalist 162 (5): 544–557. VAN DER NAT, D., K. TOCKNER, P.J. EDWARDS & J. V. WARD 2003. Large wood dynamics of complex Alpine river floodplains. Journal of the North American Benthological Society. OSEI, N.A., A.M. GURNELL & G.L. HARVEY 2015. The role of large wood in retaining fine sediment, organic matter and plant propagules in a small, single-thread forest river. Geomorphology 235. PALMA, A., R. FIGUEROA & V.H. RUIZ 2009. Evaluación de ribera y habitat fluvia a través de los índices QBR e IHF. Gayana (Concepción) 73 (1): 57–63. PALMA, A., J. GONZÁLEZ-BARRIENTOS, C.A. REYES & R. RAMOS-JILIBERTO 2013. Biodiversidad y estructura comunitaria de ríos en las zonas árida, semiárida y mediterráneanorte de Chile. Revista Chilena de Historia Natural 86 (1): 1–14. PENALUNA, B.E., I. ARISMENDI & D. SOTO 2009. Evidence of Interactive Segregation between Introduced Trout and Native Fishes in Northern Patagonian Rivers, Chile. Transactions of the American Fisheries Society 138 (4): 839–845. PILOTTO, F., G.L. HARVEY, G. WHARTON & M.T. PUSCH 2016. Simple large wood structures promote hydromorphological heterogeneity and benthic macroinvertebrate diversity in low-gradient rivers. Aquatic Sciences 78 (4): 755–766. RALEIGH, R.F., T. HICKMAN, R.C. SOLOMON & P.C. NELSON 1984. Habitat suitability. 23.
(24) information: Rainbow trout. U. S. Department of Interior, Fish and Wildlife Service (82): 64. RONI, P., T. BEECHIE, G. PESS, K. HANSON & B. JONSSON 2015. Wood placement in river restoration: fact, fiction, and future direction. Canadian Journal of Fisheries and Aquatic Sciences 72 (3). RUIZ-VILLANUEVA, V., H. PI??GAY, A.A. GURNELL, R.A. MARSTON, M. STOFFEL, H. PIEGAY, A.A. GURNELL, R.A. MARSTON, M. STOFFEL, H. PIÏ¿½GAY, A.A. GURNELL, R.A. MARSTON & M. STOFFEL 2016. Recent advances quantifying the large wood dynamics in river basins: New methods and remaining challenges. Reviews of Geophysics 54 (3): 611–652. SAS INSTITUTE INC. 2011. SAS. SAS Institute Inc., Cary, NC: 8640. SCHENK, E.R., J.W. MCCARGO, B. MOULIN, C.R. HUPP & J.M. RICHTER 2015. The Influence of Logjams on Largemouth Bass (Micropterus Salmoides) Concentrations on the Lower Roanoke River, a Large Sand-Bed River. River Research and Applications 31 (6): 704–711. SHIELDS, F.D., S.S. KNIGHT & J.M. STOFLETH 2006. Large wood addition for aquatic habitat rehabilitation in an incised, sand-bed stream, Little Topashaw Creek, Mississippi. River Research and Applications 22 (7): 803–817. SIGOURNEY, D.B., B.H. LETCHER & R.A. CUNJAK 2006. Influence of Beaver Activity on Summer Growth and Condition of Age-2 Atlantic Salmon Parr. Transactions of the American Fisheries Society. SILIGARDI M, CIUTTI F, CAPPELLETTI C, M.C. 2000. Studio sulla capacita ` di ritenzione a breve termine in un corso d’acqua alpino. Biologia Ambientale 14 (2): 7–11. SIMPSON, E.H. 1949. Measurement of diversity. Nature 163 (688): 4. SMITH-RAMÍREZ, C. 2004. The Chilean coastal range: A vanishing center of biodiversity and endemism in South American temperate rainforests. In: Biodiversity and Conservation. THEVENET, A., CITTERIO, A., PIEGAY, H. 1998. A new methodology for the assessment of large woody debris accumulations on highlymodified rivers (example of two French piedmont rivers). Regul. Rivers Res. Manag 14: 467–483. ULLOA, H., A. IROUME, M.A. LENZI, A. ANDREOLI, C. ALVAREZ & V. BARRERA 2011. Large wood in two catchments from the coastal mountain range with different land use history. Bosque. VALDES-PINEDA, R., R. PIZARRO, P. GARCIA-CHEVESICH, J.B. VALDES, C. OLIVARES, M. VERA, F. BALOCCHI, F. PEREZ, C. VALLEJOS, R. FUENTES, A. ABARZA, B. HELWIG, R. VALDÉS-PINEDA, R. PIZARRO, P. GARCÍA-CHEVESICH, J.B. VALDÉS, C. OLIVARES, M. VERA, F. BALOCCHI, F. PÉREZ, C. VALLEJOS, R. FUENTES, A. ABARZA & B. HELWIG 2014. Water governance in Chile: Availability, management and climate change. Journal of Hydrology 519 (PC): 2538–2567. VANNOTE, R.L., W.G. MINSHALL, K.W. CUMMINS, J.R. SEDELL, C.E. CUSHING, G.W. MINSHALL, K.W. CUMMINS, J.R. SEDELL & C.E. CUSHING 1980. The River 24.
(25) Continuum Concept. Canadian Journal of Fisheries and Aquatic Sciences 37 (1): 130–137. VERA, M., C. JARA, A. IROUME, H. ULLOA, A. ANDREOLI & S. BARRIENTOS 2014. Reach scale ecologic influence of in-stream large wood in a Coastal Mountain range channel, Southern Chile. Gayana 78 (2): 85–97. VILA, I. & E. HABIT 2015. Current situation of the fish fauna in the Mediterranean region of Andean river systems in Chile. FiSHMED Fishes in Mediterranean Environments (2015): 1–19. VILA, I. & R. PARDO 2009. Peces límnicos. In: Biodiversidad de Chile, Patrimonio y Desafíos especies:. 302–307. WALLACE, J.B., J.R. WEBSTER & J.L. MEYER 1995. Influence of log additions on physical and biotic characteristics of a mountain stream. Canadian Journal of Fisheries and Aquatic Sciences. WALLERSTEIN, N.P., C. V. ALONSO, S.J. BENNETT & C.R. THORNE 2001. Distorted froude-scaled flume analysis of large woody debris. Earth Surface Processes and Landforms. WARREN, D.R., E.S. BERNHARDT, R.O. HALL & G.E. LIKENS 2007. Forest age, wood and nutrient dynamics in headwater streams of the Hubbard Brook Experimental Forest, NH. Earth Surface Processes and Landforms. WOHL, E. 2006. Human impacts to mountain streams. Geomorphology 79 (3–4): 217–248. WOHL, E. & K. JAEGER 2009. A conceptual model for the longitudinal distribution of wood in mountain streams. Earth Surface Processes and Landforms 344 (January): 329–344. WOHL, E., S.N. LANE & A.C. WILCOX 2015. The science and practice of river restoration. Water Resources Research 51 (8). WOHL, E., L.E. POLVI & D. CADOL 2011. Wood distribution along streams draining oldgrowth floodplain forests in Congaree National Park, South Carolina, USA. Geomorphology. WOOD-SMITH, R.D. & J.M. BUFFINGTON 1996. Multivariate geomorphic analysis of forest streams: Implications for assessment of land use impacts on channel condition. Earth Surface Processes and Landforms 21 (4): 377–393.. 25.
(26) Figure.1. Stream parameters. present. different. behaviours. along the longitudinal profile.. Headwater zones present steep slopes with low water discharges as well as big sediment calibre. As the river enters the transfer zone slope losses steepness, while water discharges increase gradually. Finally, deposition zones are composed by high order streams with big water discharges and wide/deep channels. Church (1992).. 26.
(27) Figure 2. Flow Diagram for a Wood Budget in a Watershed. Open squares represent geomorphic areas related to locations for the sources and storages of wood, and filled squares represent processes that affect wood transport (Hassan et al., 2006). 27.
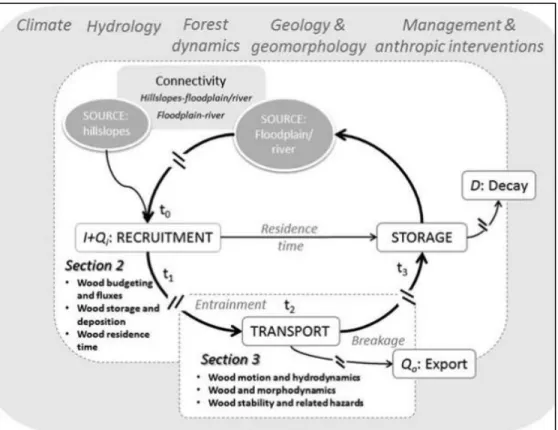
(28) Figure 3. Large wood dynamics in forested river basins illustrated as a cycle where recruitment from wood sources (i.e., hillslopes and fluvial corridor) might be transported and/or stored. The nonlinearity of these processes is illustrated as t0–t3, as recruitment and transport can be induced by steady and episodic disturbances. The time between the processes (t0–t3) may vary among different rivers or within the same river, and this defines the residence time of wood in the system. The two white rectangles show: section 2 focuses on the wood budget, and section 3 focuses on wood transport. I is lateral wood recruitment, D is in situ decay, and Qi and Qo are fluvial transport of wood into and out of the system. (Ruiz-Villanueva et al., 2016). 28.
(29) Figure 4. Conceptual model showing some of the main links between local processes and river habitat characteristics. Modelo conceptual que muestra algunas de las principales relaciones entre procesos locales y características del hábitat fluvial (Elosegi et al., 2011).. 29.
(30) Chapter 2 THE EFFECTS OF REPLACING NATIVE FOREST ON THE QUANTITY AND IMPACTS OF IN-CHANNEL PIECES OF LARGE WOOD IN CHILEAN STREAMS L. MAO a*, F. UGALDEa, A. IROUMEb AND S. N. LACYa. a. Pontificia Universidad Católica de Chile, Department of Ecosystems and Environment, Santiago, Chile b. Universidad Austral de Chile, Faculty of Forest Sciences and Natural Resources, Valdivia, Chile. This chapter was published in River Research and Applications – Copyright © 2016 John Wiley & Sons, Ltd. (vol.33, Issue 1, pages 73-88) 30.
(31) ABSTRACT Dead trees in rivers can significantly affect their morphological and ecological properties by increasing flow resistance, affecting sediment transport, and storing organic matter. Logs are usually recruited from banks or along the entire upstream basin. Although it is generally acknowledged that forested headwater streams feature higher volumes of in-channel pieces of large wood, the influence of forest type and forest management of the potential recruitment zone on the volumes and effects of wood have been less explored, especially in relation to the effects of replacing native forests with pine plantations. This paper presents a comparison of volumes of wood, and characteristics and effects on streams draining paired basins with comparable slopes, areas, and hydrologic regimes, but different in terms of land use. The five selected pairs of basins are located in the Coastal and Andean mountain Ranges in central Chile, in order to compare native forest and pine plantation basins. The results show that logs tend to be shorter and with larger diameters in streams draining native forest basins. Because of their smaller dimensions, logs and jams tend to be more mobile and oriented parallel to the flow. Volumes of in-channel wood in native forest basins are only slightly larger than in pine plantation basins, and no differences have been identified in terms of morphological effects on channel geometry. Also, fish type and biomass were comparable among pairs. Evidence highlights the importance of the width of riparian buffers in mitigating the effects of land use change, especially the substitution of native forest with plantations. key words: native forest; pine plantation; large wood; wood jams; fish; Chile. 31.
(32) INTRODUCTION In forested basins, in-channel pieces of large wood (LW, i.e. logs coarser than 0.1m and longer than 1m) can significantly affect the morphological and ecological properties of rivers. Logs, especially if jammed, can create steps (Rosenfeld and Huato, 2003), increase flow resistance (Cadol and Wohl, 2013), store organic matter (Beckman and Wohl, 2014; Tank et al., 2010), and increase the connectivity with floodplains (Sear et al., 2010). It has also been demonstrated that in-channel wood increases habitats and biological diversity (Cordova et al., 2007; Gerhard and Reich, 2000; Vera et al., 2014), principally by augmenting morphological diversity and complexity (Gurnell et al., 2002). Indeed, the presence of LW can increases fish number and biomass (e.g. Schenk et al., 2015), and engineered LW is commonly used in river restoration practices in order to increase fish biomass (Abbe et al., 2003). On the other hand, when massively recruited and transported during extreme events, LW is potentially dangerous to human infrastructure (Mazzorana et al., 2009). Trees can be delivered to the river by natural mortality or after episodic events such as wildfires, windthrow and forest diseases, or localized mass movements on slopes in the form of landslides or debris flows (Hassan et al., 2005). As well, logs can be recruited into river systems from riparian areas through toppling and bank erosion (Jeffries et al., 2003). When recruited, LW can be transported downstream for a distance that depends on the stream power (Merten et al., 2010), the relative size of the logs compared to the stream (Bocchiola et al., 2006; Wohl and Goode, 2008), and the degree to which logs are already jammed (Gurnell et al., 2002). Log recruitment and transport processes vary spatially and temporally depending on type, magnitude, and frequency of processes occurring at the basin scale (Wohl and Jaeger, 2009). Several attempts have been recently made to organize this into conceptual (e.g. Benda and Sias, 2003) or 32.
(33) numerical models (e.g. Mazzorana et al., 2010; Rigon et al., 2012). Generally, forested headwater streams (<20km2) feature higher volumes of wood, and logs are less jammed than in downstream reaches. This has been related to the considerable recruitment of logs because of high connectivity with colluvial processes occurring on slopes, and with the transport-limited conditions for wood in such environments (Abbe and Montgomery, 2003; Hassan et al., 2005; Marcus et al., 2002; Rigon et al., 2012; Wohl and Jaeger, 2009). The volume of LW in forested basins clearly depends on the type, age, and management of the forest cover. In general, old-growth forests feature a multi-layered canopy with taller and larger diameter trees that supply larger volumes of wood to rivers, which in turn can trap more floated logs, resulting in higher volumes of in-channel wood (Wohl and Beckman, 2014). Jackson and Wohl (2015) recently reported higher wood loads in old growth rather than in younger growth forests in the Southern Rocky Mountains (US). However, there is little evidence in the literature on the effects of land use changes on the loads of in-channel wood. For instance, Burrows et al. (2012) showed that streams draining clear-cut eucalyptus basins have greater abundance and volumes of wood than old-growth basins. Nevertheless, there is less field evidence for the effects of native forest replacement with exotic species. There has been extensive study of the effects of substituting native forests with exotic forest plantations on different biota and ecological processes (Arevalo and Fernández-Palacios, 2005; Brockerhoff et al.,2003; Lindenmayer et al., 2000; Vergara and Simonetti, 2004; Vertessy et al., 2003) by comparing plots established on plantations and adjacent native forests. Evidence show that fish communities can also be affected by pine plantations because of changes in nutrient concentration, shading, amount, and quality of organic matter inputs, and that these negative effects can be mitigated by the maintenance of a riparian vegetative boundary (Davies and Nelson, 1994; Lee et al., 2004). These studies have 33.
(34) mainly reported reductions in biodiversity in plantations depending on the management and silvicultural methods. In spite of these studies, much less evidence is available on the impact of native forest substitution by forestry plantations at the basin scale, and especially on how this is reflected at the channel-reach scale. For example, Baillie and Davies (2002) compared basins with native and pine plantation forest covers in New Zealand and reported higher volumes of LW in pine plantation basins. However, eco-morphological effects appeared to have been greater in the native forest basins. Evidence on this for the Andean region is virtually absent. Pinus radiata D. Don (P. radiata) comprises almost 1.5 of the total 2.7 million hectares of planted forest (INFOR, 2009). Replacement of native forest by pine plantations in Chile has occurred mainly in Andean areas as native forests in coastal areas were largely eliminated before the development of plantations (Donoso and Lara, 1995). Final. harvest of P. radiate plantations in Chile is mainly. based on clearcutting, as in most countries where this species is cultivated. The main objective of this study is to assess the impacts of substituting native forests by pine plantations on in-stream large wood and related eco-morphological effects. To achieve that, we compared the characteristics of in-channel LW in streams draining paired basins that are different in terms of land use. The paired-basin study involved the use of five pairs of basins where the pairs were similar in terms of slope, aspect, soils, drainage area, climate, hydrologic regime, and located as close as possible to each other. Five pairs of basins were selected in this study to maximize the difference in terms of land use (native forest vs. pine plantation). This paper focuses on the impact of vegetation changes at the basin scale on the amount, size, and effects of in-channel wood in the Andean and Coastal Ranges of Chile.. MATERIALS AND METHODS 34.
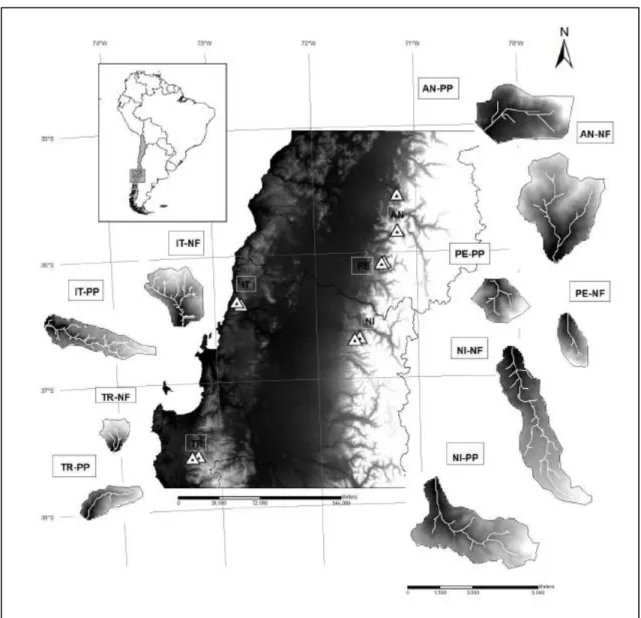
(35) Field sites The study was carried out on five pairs of basins, with draining areas ranging from approximately 2 to 20 km2 (Table I), located on the Andean and Coastal Ranges (three and two pairs, respectively), with latitudes ranging from 35°32′ to 37°34′ (Figure 1). The drainage area, stream order, and main slope of the studied basins were determined with a digital elevation model (30-m size cells) using GIS software. The percentage of forest cover was derived from visual interpretation of recent aerial photos. Although most of the basins with native forest featured nearly the whole area covered with this forest type, nearby basins with plantation that could be selected in this study have areas covered by pine of about 60–75%. Basins with pure pipe plantations are rare in the area, especially because riparian areas are protected by law. According to Gajardo (1994), all the studied sites are classified in the deciduous forest region, a temperate forest area dominated by deciduous species such as Nothofagus obliqua, Nothofagus glauca, Nothofagus alpina, and evergreen species like Nothofagus dombeyi, Cryptocarya alba, Aextoxicon punctatum, among others. All the watersheds with replacement, previous to planting P. radiata on the slopes, were covered with native vegetation, so that any changes registered in the watersheds are attributable to the replacement of native forests by plantations of pine trees and the activities associated with their management. At the time of the surveys, the plantations were between 12 and 18 years old and all had experienced only one rotation, or clear-cut and replanting. In all cases, the method of harvest is clear-cutting. Tress in the pine plantation areas have mean height of approx. 20m and mean diameter of 25 cm. Although the height of trees is. 35.
(36) not available for all study sites, the mean diameter of trees in the riparian area and the slopes along the study reaches is reported on Table II. According to Gutierrez and Becerra (submitted) the most common native species present on the riparian areas of the 10 basins were A. punctatum, Aristotelia chilensis, Citronella mucronata, C. alba, Lomatia dentata, Luma apiculata, Chusquea quila, Boquila trifoliolata, Cissus striata, Hydrangea serratifolia, Lapageria rosea, and Lardizabala biternata. The most common exotic species were Rubus ulmifolius, Rosa moschata, Prunella vulgaris, and Rumex acetosella. Although the median size of trees is comparable on riparian forest along the streams draining basins with native forest and pine plantation, riparian forests in pine plantation basins feature lower density of trees, lower diversity of tree species, lower regeneration of tree species, and higher richness of exotic species than in watersheds without replacement of native forest (Gutierrez and Becerra, submitted). The northernmost pair on the Coastal Range is located at the estuary of the River Itata (IT). The two basins are approximately 5 km2 , and are only 5 km apart. The basin with native forest (IT-NF) features a deciduous Mediterranean coastal forest but 38% of the basin area is covered by almost mature P. radiata. In the paired basin (IT-PP), the forest cover is dominated by pine plantation (75%), and a fourth of this area was bare because of a clear-cut that took place two years before the surveys. There is still native forest in 25% of the basin, especially in the lower part and along the river network, which is quite developed, considering that the stream is an order 4, despite the small size of the basin (5km2 ). The second pair of basins on the Coastal Range is located in the Trongol Valley (TR). The native forest in this area (TR-NF) is completely covered by mixed deciduous coastal forest.. 36.
(37) Although there were no visible scars of landslides or debris flows in recent aerial and satellite images, field evidence (e.g. typical depositional levees, and the presence of large boulders) suggests that debris flows have occurred in the past in the main channel. The pair basin (TR-PP) is located 4 km apart from TR-NF, and 60% of its area is covered by mature pine plantation. On the Andean Range, the southernmost pair of basins is located in the Rio Cato Valley, a tributary of the Ñuble River (NI). Some 71% of the surface area of the plantation pair (NI-PP) is covered with pine plantation, 10% of which was clear-cut at the time of the surveys, mostly in its uppermost part. A further pair on the Andean Range is located in the Achibueno Valley (PE). The basins are around 3 km2, are at similar altitudes, and are less than 5 km apart. The native forest in this area (PE-NF) is completely covered by Andean Mediterranean deciduous forest. Its pine plantation counterpart (PE-PP) is planted with P. radiata on 66% of its surface, which had been clear-cut for the first time just a year before this study. The basins of the last pair on the Andean Range basins are approximately 30km apart, and the range of elevations differs more than in the other pairs. Around 67% of the native forest basin (AN-NF) is covered by the Andean Mediterranean deciduous forest up to 1200m.a.s.l., above which the basin is almost bare. Pine plantations cover 68% of the pair basin (AN-PP), and native forest is concentrated on the highest part of the basin and along the river network.. Field data collection A segment of at least 370m of every studied basin was surveyed between November 2013 and February 2015 (see Table II). Between 6 and 12 reaches were selected at every basin. The reaches were defined as uniform in terms of slope, channel width, channel morphology, and 37.
(38) abundance of in-channel wood. The lengths of the reaches were generally approximately 10 times the bankfull width (Table II). Longitudinal profiles and three cross sections per reach were surveyed using a laser distance meter with clinometer and a prisma pole. The cross sections were measured in order to calculate the average bankfull width and depth, and the average fluvial corridor width of all the reaches. The longitudinal profiles were used to count the number of steps and pools and to calculate the longitudinal area of pools. Pieces of wood lying both in the active channel and the adjacent active floodplain were measured if the diameter was greater than 10 cm and the length was greater than 1m, as previously done in similar field studies (e.g. Comiti et al., 2008; Iroumé et al., 2010; Mao et al., 2008; Wohl et al., 2010). Logs lying alone on the bed were classified as single logs, whereas if two logs were grouped or in contact, they were classified as jammed. All single logs and logs belonging to jams were measured. The length and mid-diameter of every log were measured with a tape and a tree caliper, respectively. The volume of each log was calculated from its middiameter and length, assuming a solid cylindrical shape. The size and volume of rootwads present were measured and the volume was added to the volume of the log. Several other. measurements were recorded for each piece of wood during the field survey,. including the type of piece (log, rootwad, log with rootwads), tree species (broadleafs vs. conifers), orientation to flow (parallel, orthogonal, oblique), state of decay (fresh, semi-decayed or old log), and position (within or outside the bankfull line). The most probable recruitment mechanism that delivered the log into the channel was determined in the field as landslide, debris flow, bank erosion, natural mortality, artificial cutting, or transported from upstream. Jams were defined following a simplification of the Abbe and Montgomery (2003) classification, which distinguishes autochthonous jams (i.e. key wood elements not fluvially 38.
(39) transported from upstream), allochthonous jams (i.e. key elements previously transported from upstream) and combination jams (autochthonous key elements with racked transported pieces). For transversal logs and jams forming a step in the profile, the volume of the downstream pool and the volume of sediment stored upstream were estimated as a solid wedge by measuring their length, width, and depth using a tap measure. At all the studied sites fish was sampled in the summer along a 100-m-long reach blocked with nets at the up- and down-stream ends, using a standard double-pass electrofishing technique (HT2000 Battery Backpack Electrofisher device, Halltech Aquatic Research Inc.). All fish were identified at the species level, measured for total length, and weighed before being returned to the river.. RESULTS. Dimensions and type of log pieces in the studied rivers The average diameters of in-channel logs ranged from 10 to 25 cm, while the maximum diameters range from 15 to 80 cm (Figure 2). Taking average diameter into consideration for comparing native forest- and pine plantation dominated basins, a F-test confirms that the average diameter was significantly greater in native forest basins in the AN and NI pairs, while the maximum diameter was significantly coarser in the native forest basins of the TR, AN, and NI pairs (Figure 2, Table III). In-channel logs were generally longer in basins where pine plantations predominate. Figure 2 shows that the average length of logs surveyed in the studied rivers ranged from 2 to 4 m, and the average length of logs in basins with pine plantation was greater than those from natural forest 39.
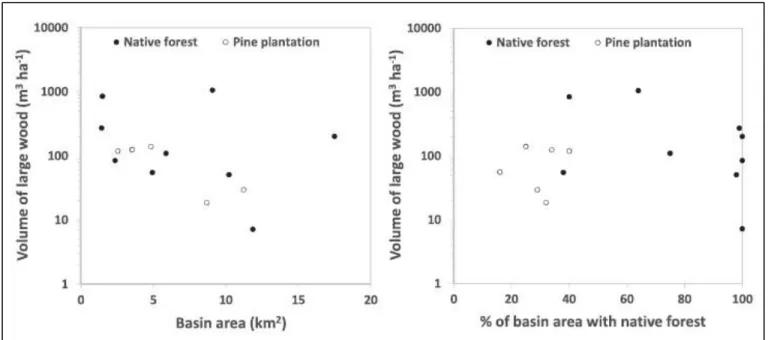
(40) basins (statistically significant differences in pairs IT and TR, Table III). If we take into consideration the longer logs surveyed at the studied reaches, pine plantations basins had significantly longer logs only in the pairs IT and PE (Figure 2, Table III). However, in all but one case, the longest logs per reach (generally between 5 and 20m long) were always pine species, and the longest log surveyed was a pine species of over 30m in the PE-PP channel. With the exception of AN, the percentage of pine logs was higher in basins with pine plantation (Figure 3; Table III). This is especially true for the IT pair, where in the pine plantation basin more than 80% of the in- channel logs were conifers (more rounded and straight than native species, and with smaller branches). Figure 3 also shows that logs in native forest basins were in a poorer state of conservation in three of the five study pairs (AN, PE, and NI).. Volumes, abundance, and degree of accumulation of logs in the studied rivers Figure 4 shows the volume of large wood surveyed in the studied sites. To compare reaches of different lengths and widths, volumes are expressed in terms of channel area units (length and bankfull width of the reach). The volumes of wood at the studied reaches varied by a magnitude of two orders, ranging from 6m3 ha_1 in a reach of the IT native forest basin to 1780m3 ha_1 in a reach of the TR native forest basin. The volume of logs does not appear to depend on the location of the basins (Coastal vs. Andean Range), but in three of the five pairs (TR, PE, and NI) LW volumes in streams draining native basins were nearly double that in the pine plantation basins (Table III). However, LW volumes were significantly higher in the pine plantation basin of the IT pair (Table III). Volumes of wood in the studied sites can be compared to other evidence gathered in Chilean streams over the last decade. In particular, data on volumes of in-channel wood are available for 40.
(41) five other basins comparable to the studied sites: Tres Arroyos (9km2, 64% with native forest; see Andreoli et al., 2007 and Comiti et al., 2008), El Toro (17.5km2; 100% with native forest, see Andreoli et al., 2007), Vueltas de Zorra (in the Coastal Range, 5.87km2, 75% with native forest; see Iroumé et al., 2011, 2015 and Ulloa et al., 2011), Pichun (a pine plantation basin of 4.3km2 in the Coastal Range; see Iroumé et al., 2011, 2015 and Ulloa et al., 2011), and Milico basin, (in the Andean Range, 1.5km2 , 40% with native forest, being the rest above the timberline; Gomez, 2013). Figure 5 shows that in small mountain basins (<15km2 ) in-channel volumes of large wood can vary by three orders of magnitude, ranging from 10 to 1000m3 ha_1, with a weak tendency toward a reduction of large wood storage with greater basin area (R=_0.12; p>0.66). Basins with dominant forest cover tend to feature higher volumes of LW (F test = 1.855; p = 0.19). However, basins with a higher percentage of native forest do not necessarily have higher volumes of LW, as volumes can vary quite dramatically (two orders of magnitude) even in basins with pure pine plantation. There were no significant differences in single log volumes between the native forest and pine plantation forest pairs at any of the five sites (Figure 4, Table III). As well, no significant differences between native and pine plantation basins were found in terms of volumes of jammed logs, and significantly higher volumes of jammed LW were only found in the NI native forest. Because LW volumes depend on both the number and dimension of logs, it is worth considering the number of logs and jams. Figure 6 shows that there were no significant differences between native forest and pine plantation basins in terms of the number of isolated logs, except for the TR basin (Table III). The only site with a significant difference in the number of log jams was AN, where the number of log jams was higher in the plantation forest site.. 41.
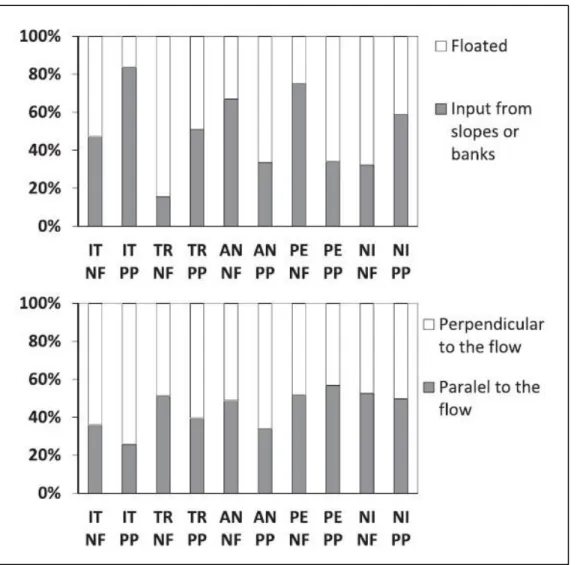
(42) Figure 7 shows the percentage of logs recruited from the banks/slopes or from upstream reaches (i.e. floated by previous flood events). It appears that in the IT, TR, and NI pairs, a higher percentage of the logs were most likely to have been transported in streams draining native forest basins than from the plantation forest basins. Figure 7 also shows that in four of the five pairs, logs in native forest basins tend to be more parallel than those in pine plantation basins (statically significant differences were found for the IT and AN pairs; Table III). Figure 8 shows that jams tended to be more autochthonous in pine plantation basins in the AN, NI, and TR pairs, but this difference was statistically significant only for NI. As to the orientation of jams, Figure 8 shows that logs tended to. accumulate in a direction parallel to the flow in. native forest basins of the TR, AN, and PE pairs (the difference is statistically significant only for AN; Table III).. Eco-morphological effects of in-channel logs In the studied sites, no clear evidence of differences were observed between streams draining native forests or pine. plantations in terms of the number or dimensions of steps and pools.. Retention of sediments because of jams was not significantly different in native forest versus pine plantation basins. Indeed, Figure 9 shows that the ratio between the volume of trapped sediments and the volume of jammed logs is between 1 and 3, with no significant differences among basins, except for TR. The remarkable value (around 7) obtained for the native forest basin of the TR pair is because of the massive amount of sediments coming from debris flows and trapped by jams. A total of three fish species were encountered in all ten watersheds, namely rainbow trout (Oncorhynchus mykiss), brown trout (Salmo trutta), and the Chilean catfish (Nematogenys 42.
(43) inermis). Two of the three species were encountered in all watersheds, except for Las Arañas (TRNF), where no fish were found. Rainbow trout (O. mykiss) were found in all sites except TRNF, brown trout (S. trutta) were encountered in six sites (IT-NF, NI-NF, NI-PP, PENF, PE-PP, and TR-PP), and Chilean catfish (N. inermis) were found in two sites (AN-PP and IT-PP). If the fish biomass per unit effort (BPUE: g 250m-2 ) is considered, a total of 1250.7 g of fish per 250m2. of stream were found in native forest streams, whereas 1088.8 g of fish per 250m-2 of stream. were found in pine plantation streams (Figure 10). The difference in biomass (161.9) is not significant (two sample t-test = 0.42; p = 0.69) between the two land uses. In assessing the associations between fish BPUE and LW volumes, sites TR-NF and NI-PP were both excluded as outliers. The volume of large in-channel LW did not correlate significantly with the number of species or BPUE in any of the watersheds, regardless of dominant land use. Only two factors appear to be significantly related to fish BPUE, namely maximum log diameter (F = 7.57, P= 0.03, R2 = 0.58) and maximum log length (F = 3.75, P= 0.10, R2 = 0.42) (Figure 10).. DISCUSSION. Dimensions and type of log pieces in native forest versus pine plantation basins The results of the comparative analysis of paired native forest versus pine plantation basins suggest that pieces of large in-channel wood tend to have larger diameters in the native forest basins. This can be related to the fact that native forest basins naturally tend to feature older and coarser plants that are unlikely to be left growing in basins with pine plantations. Indeed, in four of the pairs the mean diameter of standing trees on the slopes is larger in the native forest than in the pine plantation basins. Even if the riparian area is left untouched, it is 43.
(44) generally less likely to include very old and large plants in close proximity to the channel (because of higher frequency of flood disturbances) rather than on the slopes. In fact, in four of the pairs the mean diameter of trees growing in the riparian area is larger in the native forest than in the pine plantation basins. On the other hand, in-channel logs tend to be longer in basins with pine plantation. This is probably because of the fact that many pine logs are recruited from the slopes or after clearcutting (many pine logs were artificially cut), and were therefore generally longer than native species, and in a better state of conservation. There was generally a higher percentage of in-channel logs in a poor state of conservation in native forest basins (i.e. without branches and bark, and with porous/rotted wood), which makes it easier for the flow to break up longer logs during floods. In support of this observation, field evidence suggest that at least in three of the five pairs, logs are predominantly transported from upstream reaches in native forest basins, whereas in pine plantation basins logs were recruited more from banks and slopes than transported from upstream. There is no clear explanation as to why the AN and PE pairs feature more floated logs in the pine plantation basins, but at least for the AN pair this could be because of the generally smaller dimensions of logs in the pine plantation basin. Overall, there are weak indications that logs tend to be more parallel to the flow (i.e. logs have been transported from upstream, e.g. Francis, 2007) in native forest basins, while logs in pine plantation basins tend to be more oblique or perpendicular to the flow, indicating that they have been in the channel for less time and are close to the point of recruitment from the banks or slopes. However, the results obtained in our field investigation do not provide unequivocal evidence that there were more transport or combination jams in native forest basins, and that log jams. were not predominantly oriented parallel to the flow in native forest basins, as one might. expect if logs were relatively. smaller and more mobile in native forest basins. Besides, the 44.
(45) presence of complex branches in broadleaf species of native forest basins could reduce the mobility of logs if compared with conifers (Dixon and Sear, 2014). Results show that there were no significant differences between native forest and pine plantation basins in terms of volumes of jammed logs or the number of log jams. The number of log jams per ha of stream ranged from o to more than 200, and significant differences could be identified among pairs. In AN, the very low number of jams in the native forest basins could be caused by the higher discharges (because of the larger basin area) and more likely by the wider bankfull, which reduces the chances of large logs being trapped by the banks and creating jams (as logs tend to be more mobile at smaller piece length/channel width ratios, e.g. Bocchiola et al., 2006; Dixon and Sear, 2014). The considerably higher number of jams in both TR pair basins (around 100 jams ha-1 i.e. around 80 jams km-1 ) helps explain the high LW volumes in both, even though log diameter and length were comparable to those in the other studied pairs. This high longitudinal frequency of jams is because of high recruitment of logs and to the presence of potential ‘trapping’ sites. Montgomery et al. (2003), the number of logs per unit of channel length decreases with increasing basin area, and has been reported to be as high as 400 jams km-1 . The very high number and volumes of LW elements could be related to the fact that the two TR basins are the smallest among the selected pairs. Following the conceptual model of longitudinal distribution of wood proposed by Wohl and Jaeger (2009), higher LW volumes are likely to be found in narrower and steeper streams, and LW volumes tend to decrease downstream. Wohl and Jaeger’s (2009) model also suggests that the percentage of jammed wood increases with drainage area and channel width because of the higher capacity of logs to be transported. The fact that the TR pair featured the highest number of jams may thus be related to an unlimited transport. 45.
Figure

+7




Documento similar