The effects of methodological limitations in the study of butterfly behavior and demography: a daily study of Vanessa atalanta (Lepidoptera: Nymphalidae) for 22 years
9
0
0
Texto completo
(2) REVISTA DE BIOLOGÍA TROPICAL. 606. How buUerflies reduce the effect of eco. MATERIALS AND METHODS. logical fadors: Butterflíes can control density dependent and density independent factors in. Individual butterfly visitation records (and. two ways: (1) locally by behavioral and phy. a qualitative microclimatic classification) were. siological mechanisms, and (2) ex situ by mi. made daily by the senior author in an urban. gration. Behavioral mechanisms inelude actí. clearing devoíd of hostplants in Winter Park,. vity pattern coordination with the hostplant's. near Orlando, Florida,. USA (28°34'57" N,. tations even involve larval monitoring of day. 81°20'04" W) for 22 years (April 15, 1977April 14, 1999: 8035 days). Details oí the site. length to control development (Young 1983,. and the unusual conditions that allowed daily. Kunte 1997, Nylin 1997).. observations íor such a long period appear in. reactions to cHmate, while physiological adap. Migration: Butterfly travelling varies. Swanson (1998).. from small changes in microhabitat distribu. Climatic data are from monthly means pro. tion to long range migratíon. Migration seems. vided by government databases available in the. to be genetically determined and relates with. Internet (http://water.dnr.state.sc.us/elimatelMay. lifespan (Baker 1984, Ehrlich 1984). Solar ra. 1999). Besides the inferential statistics presented. diation, temperature and wind may (Calvert et. here, graphs of daily changes in number of visi. al. 1992, Monge-Nájera et al. 1998) or may not (Frey et al. 1992) he}p predict the micro. ting butterflies and their arrival time (available in. habitats where populations concentrate. It seems clear that topography (and the micro. visually for non-linear associations (Kozlov et al. 1997) but none were found. Non-parametric. elimate pattern that topography creates) affect. statistics were used to avoid potential conflict. the on-line edition oí this journal) were analyzed. both butterfly population size and biodiver. with the less realistic requirements oí parametric. sity (Gutiérrez 1997).. tests. "Visitation" and "number oí daily visitors". Mid-range movements: Mid-range mo vements are associated with isolated habitat. are defined as total number of occupants and in truders arriving each day.. patches that predispose a species to the shifting mosaic metapopulation model (Harrison et al.. 1998), with relatively frequent local extinc tions and re-establishments. Stepping-stone gene flow is important at least in sorne metapo pulations (Neve et al. 1997, Peterson 1997, Singer and Thomas 1997).. Long-mnge migrations: Long-range mi grations are known in tropical and temperate butterflies, among them the Red Admiral, Vanes. sa atalanta L., a species that appears to fit the re quirements for survival in urban areas (Swanson. 1979, Kitahara and Fujii 1997, Srygley el al. 1997, Thomas 1984). It is a widespread nympha lid íound in the American continent from central Canada to Guatemala, often in elearings (Bitzer and Shaw 1979, TubervilJe el al. 1996). This paper reports on an intensive (daily) long term study (22 years). of a V. atalanta po. pulatíon in a subtropical habitat with emphasis on the association of number of individuals and arrival time with elimate.. RESULTS. Natural history: Individuals were active even at 10° C and with 9 mis winds. Individuals with wings in fresh condition were most com mon in the first half of year (January-June) when they represented roughly three quarters of the population. Smaller individuals (general impression: no measurements were taken) were seen from late December to late March. There were 82 atypical cases of individuals arriving before 1200 hr (not included in the graphs). Ae ríal interactions were seen whenever there was more than one individual in the site U.e. 41. %. of days, N=7634 total days, Fig. 1). A total of 4 794 visits were recorded, with an overall attendance rate of 59.5. %. During the. 2 hr aftemoon visit period there were fre quently 1-3 individuals in the site, two times there were eight and once (March 15, 1982) there were ten. Only once in these 22 years was.
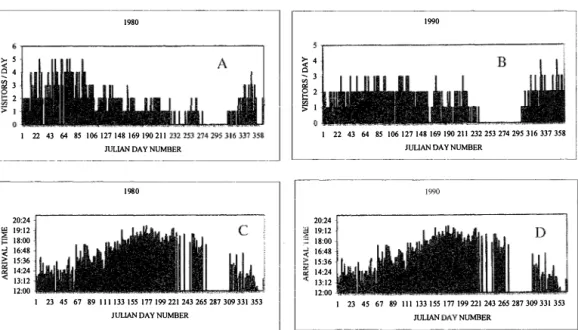
(3) INTERNATIONAL JOURNAL OF TROPICAL BIOLOGY AND CONSERVATION. 607. Fig. 1. Aerial interactions of Vanessa atalanta and study site (from a video).. 19 80. ,. 5. «. :: iQ. �. I A. 4 3 2. >1. 1. 22. 4364. 85 106 127148 169 190 211 2 32 25327 4295316 337358. JULlAN DAy NUMBER. 19 80. 20:24. '" 19:12. ;;¡ r-. ; �. 1990. I. �. 1. '. ;;; '" ¡::. �. 16:48 142 : 4 1l:12 12:00 1. 23 45 67 89 III 1 33155 171199 221 2432652 87309 331 353 JULlAN DAY NUMBER. 1. 22 43 64. 4295 316 337358 85·106 127148 169 190 2ll 2 32 25327 JUUAN DAY NUMBER. ,. I. 1>. 15:36. 3 2. 1. I!. 1 8:00. 4. I� I. 1990. 20:24 19:12 18:00 16:48 15: 36 14:24 1 3:12 12:00 1. 7199 221 243265287309 331 353 23 45 67 89 111 133155 17. mLlAN DAy NUMBER. Fig. 2. Two typical visitation and first daily arriva1 time pattems throughout the year for Vanessa atalanta in a subtropical habitat in Florida. Visitation is shown as number oI visitors per day and arrival time of first visitar is given in hours. A.,e. Visitation rate and initial daily arrival times for 1980. B.,D. Visitation rate and initial daily al'rival times for 1990..
(4) REVISTA DE BIOLOGÍA TROPICAL. 608. 0:00. .4 .. , ;,. •. 3.5. .' .. g. �. .•.. I. ..... 0:00. •. •. • • • • • ,• • .. 2.5. ... ti.. • •. •. •.... • •.. 1. �. \o. J e;. ... 0.5 .. I . .. •. , �" • •. • ... ., •• • ;, .. �. •. : ... .1. ... . •. 0:00. •. •. .. 0:00 O. 15. 10. 20. 4�-------, •. 19 •. 3. • •. : .. .. •. !. .... \)} .. ..." � •. .. .. .... .. •. •. •. .. -. . •. . .. ' •••. .. él. . . ... .q.. -... ;. ... O.S·. •. •. 18. •. •. •. 15. 10. S. PRECIPIT ATlON (mm). PREClPIT ATlON (mm). 3.S. .. • • •• •. . .·'"t''' • •. ••. S. O. 0:00. <[. •. . .... . . o··����""���� .. •. i. •. , ... , I't ·.1· .' '!ti' • • •. ,. •. •. •. •. 3:. •. •. • •. .-... . . ... -.¡ •. ,. •. ..:1 íI. ;. � 16 �I. i. •. •. •. •.. � I. �. lS. •. ,. •. 14. 13·. •. •. 17 ·1. • •. !. 12 i. . ---�" '--" " " " " " " '. " " " " " " " '_r---� " " " " ... 45. SS. 65. 7S. TEMPERATURE (C). 4S. SS. 65. 7S. í. 8S. TEMPERATURE (C). Fig. 3. Relationship of visitation (left) and arrival time (right) of Vanessa atalanta with precipitation (above) and tempe rature in a subtropical habitat (statistical values in text)..
(5) 609. INTERNATIONAL JOURNAL OF TROPICAL BIOLOGY AND CONSERVATION. 23 22 21. 18:00. j. 16:48 15:36. i. 201. I I. +-r-··l"·--,--.,.-····,.----:-·-r-.......,.-�__r__·_.,_,-.---,-+ _,,__�r__r__,.___,.____,__. ,....., 1750. §. T. 1. '-'. !. 14:24 13:12 18:00 16:48. � � �-<. ¡,¡¡. ,....., p::: ::t: '-' ¡,¡¡. 15:36 � t-< 14:24 ¡·...·....,. -T �· ... -11- 'TEMPERATURE. ···· .. T. 13:12. t. 1500 1000. __ PRECIPITATION. 0.0. (mm). -.-VISITORS (N). 2. ¡! 1.5. ¡- 1. !. :.�� 1+-:. p::: -<. 2.0 1.5 . . 1.0 0.5. --r---r--,-r-r-...-1·····.,.··.. .. r . ...-r",-,. ....-r. j.. pa. (C). -.- VISITORS (N). WOO. �. �......,. �r¡ _.-r-... ...,.......,-".... ...... .....,..'---,---,-,¡ "..-r-.... "...'Y ..... "."¡--'r¡ I T T T ...... 00 0\ ....... <"l 00 0\. ..... Ir) 00 0\ ,....;. ['-00 0\. ....... 0\ 00 0\. ....... --,-..-+...,. ·t :. .5. Year. Fig 4. Yearly arrival time and visitation patterns of Vanessa atalanta versus temperature and precipitation in a subropical habitat (statistical values in text)..
(6) 610. BIOLOGÍA TROPICAL. REVISTA DE. predation seen: an individual that flew with dif. There was no significant difference in. ticulty was captured by a bird (Cardinalis car. number of visitors between El Niño (mean. lacked parts of wings;antennae or Pfoboscis.. range 0.90-1.50, N=lI, years 1977, 1978,. plants on the site� The population is composed. non-El Niño years (mean 1.25, SD 0.28, range. dinalis). Some.spechpens had missing legs or. 1.13 visitors/day, standard deviation SD 0.21,. VlSitation pattern: There· are ·no hcJst. 1982, 1983, 1986, 1987 and 1991-1995) and. of vlsitdrs that perch and participate in aerial. 0.80-1.70 visitors/day).. interactions sensu Bitzer and Shaw (1979). The number of visitors begins to increase in au. tumo, reaches a peak near the end of winter and. DISCUSSION. decreases almost constant1y during the spring and surnmer (Fig. 2a, 2b). Generally, there. �. .. two visitation pattems.'PatteQ} 1 (1977 to. 1985): a marked curve with zero population. around Octobe,r, e,.g. Fig. 2a). Pattern 11 (1986-. 1996): a flatter curve with 1-5 months without visitors, e.g. Fig; 2b. Arrival time of tirst indivi. dual to enter the area had the same pattern du. 1Jfe migratory routes of V. atalanta in. North America are poorly known, but a greater. number of srndies in Europe have shown that the migratory population follows te�perature. layers (Benvenuti et al. 1996), is subjected to. significant wind drift and travels with other. ring the study period, with a soft increase from. species (Hansen 1997); has time-compensated. December (Julian dates, Figs. 2b and 2 d).. ters, feeds in winter aód sometimes reproduces. cular month was(inversely correlated. with mean. of the year, around June (Baker 1972, 1984,. tion, SRC: r =-0.43, p< 0.01, n=261 months;. vist 1998). It is now clear that migrations are. January through June anda .softdecline. until The mean daily number of visitorS a parti. monthly temperature(Spearman Ránk Cotrela mean monthly values for number of visitors and .. climatic factors were used in this and following tests) and precipitation (SRC: r = -0.34, p< 0.01,. N=261) (Fig. 3A,C).There were .occasional ab. sences of 1-14 days that represented gaps in the general trend most1y during stormy weather. (Fig. 2 A-D, a few represent the rare instances in. which observer was absent). Arrival time of the. first visitor was positively correlated with mean monthly temperature (SRC: r = 0.84, p< O.OI,. N=238) and with mean monthly precipitation (SRC: r = 0.49, p< 0.01, N=231) (Fig. 3B,D).. Total daily number of individuals arriving at the site was associated with a subjective classitica tion. of. climate (Kruskal-Wallis ANOVA,. p<O.oool, N=5658 days; mean ranks: 4229. windy, 3152 good climate, 2614 cold, 743 over cast or rainy) but not with arrival time (Kruskal. sun orientation, migrates thousands of kilome. while traveling northward in the warmer part Stefanescu et al. 1996, Hansen 1997, Palmq. bidirectional (Benvenuti et al. 1996) but how. the travel information is passed among but terfly generations in general is not known (Pas. teur 1984) and future studies of V. atalanta. should stress this aspect.. .. In this study, seasonal changes in the num. ber of visitors, with fewer visitors in the mois. ter and warmer months and predominance of fresh individuals from January to June suggest. that V. atalanta overwinters in Florida and mi. grates to northerly regions when high surnmer. temperatures affect Florida (R. Bitzer 2000,. pers. cornm.). A similar migration from Africa to Europe is known (Larsen 1993). This would. tit with the smaller individuals seen from late. December to late March: fresher individuals. may emerge in Florida as the offspring of the. Wallis ANOVA 1.7, p=0.63, N=4 681 days).. larger butterflies first seen in late October -. (SRC values: visitors versus temperature -0.17,. could have migrated from more northerly re. Yearly mean values were not correlated. visitors versus precipitation -0.40; arrival time versus temperature 0.17, arrival time versus. precipitation 0.02; in all cases N=22 years and p>O.05). (Fig. 4). early November. These larger individuals. gions earlier in the fall, according to the para llel migration pattern of V. atalanta in Europe. described by Larsen (1993) and Benvenuti et al. (1996) (R. Bitzer 2000, pers. comm.). For V..
(7) INTERNATIONAL JOURNAL OF TROPICAL BIOLOGY AND CONSERVATION. 611. atalanta temperature is a more important limi. general impossibility of daily observatíon for. ting factor than host plant availability (Bryant. long periods, but at least for smaU areas, oc. Simon et al. 1997, Maier and Shreeve 1995).. casional population counts covering less than. The activity period span in Florida ís. two weeks are unreliable: even total absence. greater than in the more seasonal c1imate of. of individuals for such periods can simply re. Iowa, where V. atalanta is active from April to. present gaps in the general trend. Furthermo. October (Bitzer and Shaw 1979, 1995). The. re, the analysis of yearly values hid the asso. greater number of visitors on windy days sug. ciations of number of individuals and arrival. gests that the site was selected because it was. time with climate that were found when a. surrounded by vegetation and buildings (see. smaller scale was used. Long term studies ba. Swanson 1998).. sed on yearly samplings should bere-evalua. The El Niño Migration Model predicts. ted for this aspect.. that overpopulation increases butterfly migra tion to outbreak level during El Niño years (Larsen 1984, Myres 1985). We found no dif. of visitors for El Niño years, apparently because V. atalanta is a regular rat. ACKNOWLEDGMENTS. ference in number. her than an outbreak migrator.. We thank Shirley Gregory and Pat Miller for assistance and support, María Isabel Gon. The limited temperature range that cha. zález for the statistical analysis, Marisol Ro. racterizes sites where V. atalanta occurs (Br. dríguez Pacheco and Ana Araya Anchetta for. yant-Simon et al. 1997, Maier and Shreeve. data digitalization and Royce Bitzer, Depart. 1995) and butterflies' inability to fly under. ment of Entomology, Iowa State University,. heavy rain can explaín why individuals arrive. William Ramírez, Museo de Insectos, Univer. at their perching spots later in the hot, rainy. sidad de Costa Rica, Per-Olof Wickman, Dpt.. V. atalanta males establish courtship. Mathematics and Science, Stockholm Institu. territories 2-4 hours before sunset (Wiltshire. te of Education and one annonymous revie. 1997, Bitzer and Shaw1979, 1995) and use. wer, for suggestions that greatly improved the. neural motion detectors adapted to great. manuscript.. months.. speed (O'Carroll et al. 1996) in aerial interac tions (Bitzer and Shaw 1979). Territories ha ve distinct characteristics (sunlit c1earings. REFERENCES. and ridge tops) and are used by different indi" viduals over the years (Bitzer and Shaw 1979,. Baker, R.R. 1972. The geographicaI origin of the British. Wiltshire 1997). Despite its methodological. spring individuals of the butterflies Vanessa atalanta. difference with previous studies, this study. (L.) and V. cardui (L.). J. Entorno!. (A) 46: 185-196.. produced the same conclusions about territo rial behavior, and also agrees with the eco10-. Baker, R.R. 1984. The dilemma: when and how to go or. gical model of perching behavior which pro. stay, p. 279-296. In R.L Vane-Wright & P.R. Ackery. poses that perching is associated with relati. (eds.). The biology of butterflies. Royal Entomologi. vely high temperature, lekking and long life. cal Society, London.. (Wickman 1992a,b, van Dyck et al. 1997a,b, van Dyck and Matthysen 1998. No data are. Benvenuti, S., P. Dall' Antonia & P. Ioale. 1996. Directio. available regarding the wing color and body. nal preferences in the autumn migration of the red. proportion elements of the model).. admiral (Vanessa atalanta). Ethology 102: 177-186. If our results can be generalized (and that will be unknown until more long term inten. Bitzer, RJ. & K.C. Shaw. 1979 (1980). Territorial beha. sive studies beco me available) behavioral stu. vior of the Red Admiral, Vanessa atalanta (L.) (Le. dies of butterflies do not suffer from the. pidoptera: Nymphalidae). J. Res. Lepid. 18: 36-49..
(8) REVISTA DE BIOLOOÍA TROPICAL. 612. Bitzer, RJ. & K.C. Shaw. 1995. Territorial behavior of the. Kunte-Krushnamegh, J. 1997. Seasonal patterns in but. Red Admiral, Vanessa atalanta (Lepidoptera: Nymp. terfly abundance and species diversity in four tropi. , halidae). I. The role of climatic factors and early in. cal habitats in northern Western Ohats. J. Biosci.. teraction frequency in territorial start time. J. Insect Behav. . 8: 47-66. Bryant-Simon, R., D. Thomas-Chrls & S. Bale-Jeffrey. 1997. Ñettle-feeding nymphalid butterflies: Tempe rature, development and distribution. Eco!. Ent. 22: 390-398. Calvert, W.H., L.P. Brower & R.O. Lawton. 1992. Mass flight respOnse of overwintering mOÍlarch butterflies (Nymphalidae) to cloud-induced changes in solar ra diation intensity inMexico. J. Lepid. Soco 46: 97-105.. Bangalore 22: 593-603. Larsen, T.B. 1984. Butterflies of Saudi Arabia and its neighbours. Sracey, London, 160 p. Larsen, T.B. 1993. Butterfly mass transit. Nat. History 102: 30-39. Maier, C. & T.O. Shreeve. 1995 (1996). Endothermic heat production in three species of Nymphalidae (Lepi doptera). Nota Lepid.18: 127-137 Monge-Nájera, J., F. Hernández, M.I. Oonzález, J. Soley,. Daily, O.C., P.R. EhrIich & D. Wheye. 1991. Determinants. JA Pochet & S. Zolla. 1998. Spatial distribution and. of spatial distribution in a population of the subalpi ne butterfly Oeneis chrysus. Oecologia 88: 587-596.. sound production in five species of cryptic tropical butterflies (Hcimadryas, Lepidoptera: Nymphalidae): Implication for the industrial melanism debate. Rev.. Ehrlich. p.R. 1984. The structure and dynamics of butterfly populations, p. 25-40. In R.I. Vane-Wright & P.R. Ackery (eds.). The biology of butterflies. Royal En tomological Society, London.. Bio!. Trop. 46:297-330. Myres, M.T. 1985. A southward return migration of Pain ted Lady butterflies, Vanessa cardui, over southern Alberta in the fall of 1983, and biometeorological as. Frey, D.K., L.H. Leong, D. Fredericks & S. Raskowitz. 1992. Clustering patterns of monarch butterflies (Le. pects of their outbreaks into North America and Eu. pidoptera: Danaidae) at two California central coast overwintering sites. Ann. Entorno!. Soco Am. 85: 148-153.. Nielssen, V. & J. Monge-Nájera. 1991'. A comparison of. Ooulson-D; J. Ollerton & C. Sluman. 1997. Foraging stra tegies in the small skipper butterfly, Thymelicus fla vus: When to switch? Anim. Behav. 53: 1009-1016 Outiérrez, D. 1997. Importance of historical factors on species richness and composition of butterfly assem blages (Lepidoptera: Rhopalocera) in a northern Ibe rian mountain range. J. Biogeogr. 24: 77-88. Hansen, M.D.D. 1997 Observations on the migrations of the Painted Lady Vanessa cardui (L.) in Denmark in 1996: (Lepidoptera: Nymphalidae). Ent.Meddelelser 65: 165-173.. rope. Can. Field-Nat.99: 147-155.. four methods to evaluate butterfly abundance, using a tropical community. J. Lepid. SOCo 45: 241-243. Neve, O., L. Mousson & M. Baguette. 1997. Adult disper sal and genetic structure of butterfly populations in a fragmented landscape. Acta Oecol. 17: 621-626. Nylin, S. 1997. Seasonal adaptations in butterflies: A story on plasticity. Entom. Tidskrlft 117: 1-10 O'Carroll, D.C., NJ. Bidwell, S.B. Laughlin & E.J. Wa rrant. 1996 Insect motion detectors matched to visual ecology. Nature 382: 63-66 Pollard, E; & J-N. Oreatorex. 1997. Flight-periods of the. Harrison, S., D.D.Murphy & P.R. Ehrlich. 1988. Distribu tion of the Bay Checkerspot butterfly, Euphydryas editha bayensis: evidence for a metapopulation mo de!. Amer. Nat. 132: 360-382.. small heath butterfly, Coenonympha pamphilus (Lin. Kitahara M. & K. Fuji. 1997. An island biogeographic ap. Palmqvist, 0. 1998. Remarkable records ofMacrolepidop. proach to the analysis of butterfly community pat terns in newly designed parks. Res. Popo Eco!. (Kyo to) 39: 23-35.. tera in Sweden 1997. Entomol Tidskrlft 119: 13-27.. naeus) (Lepidoptera: Nymphalidae, Satyrinae) on chalk downs and in woodland in southern England. Entom. Oaz. 48: 3-7. Pasteur, O. 1984. Les migrations du monarque, Lepidopte re Danaide. C.R. Soco Biogéogr. 60: 84.. Kozlov, M.V., A.L. Lvovsky & K. Mikkola. 1997. Abun danCe of day-flying Lepidoptera along an air pollu tion gradient in the northern boreal forest zone. Ent. Fennica 7: 137-144. Peterson, M.A. 1997. Host plant phenology and butterfly dispersal: Causes and consequences of uphill move ment. Ecology 78: 167-180..
(9) INTERNATIONAL JOURNAL OF TROPICAL BIOLOGY AND CONSERVATION. 613. Stefanescu, C., M. Lockwood & M. Cebolla. 1996. The. Wickman, P.O. 1992a. Mating system of Coenonympha. butterfly assemblages of northeastern Spanish we. butterflies in relatíon lo longevity. Anim. Behav. 44:. tlands. Nota Lepid. 19: 165-184. 141-148.. Steiner, A. 1991. Unusual flight times of butterflies in. Wickman, P.O. 1992b. The influence of temperature on. 1989 and 1990: Results of the global c1imatic chan ge? Atalanta 22: 237-244.. small heath butterfly, Coenonympha pamphilus (1. ). Singer, M.C. & C.D. Thomas, 1997. Evolutionary respon ses of a butterfly metapopulation to human- and cli mate-caused environmental variatíon. Am. Nat. 148 (Suppl.): S9-S39 Srygley, R.B., E.G. Oliveira & R. Dudley. 1997. Wind drift compensation, flyways, and conservation of diurnal, migrant Neotropical Lepidoptera. Proc. Roy. Soco Lond. Bio\. Sci. 263: 1351-1357 StekoJnikov, A.A. 1992. Dynamics of sorne Lepidoptera (Hesperioidea, Papilionoidea, Sphingoidea, etc.) fau na of the protected oak fores! "forest of the Vorskla" during 50 years of observation. Vestn. St-Peterburg Univ. Ser. 3 Biol. (2): 28-36. Swanson, H.E 1998.20 years of butterfly revelations. E.O. Painter, Winter Park, Florida. 102 p. (Library of Con. the territorial and mate locating behaviour of the (Lepidoptera: Saturidae). Behav. Eco\. Sociobiol. 16: 233-238. Wickman, P.O., C. Wiklund & B. Karlsson. 1990. Compa rative phenology of four satyrine butterflies inhabi ting dry grasslands in Sweden. Rolarctic Ecol. 13: 238-346. Wiltshire, E.P. 1997a. Territorial red admirals: Beha viour of Vanessa atalanta (Lepidoptera, Nymphali dae) in a Berkshire garden during the 1995 heatwa ve, discussed and compared. Entomologist 116: 5865. Wiltshire, E.P. 1997b. Observations of butterfly behavior in 1996 in Berkshire: Interplay between Vanessa ata. lanta (L.), Cynthia cardui (L.) (Lep.: Nymphalidae) and one gay cat, an (?) alluring predator. Ent. Rec. J. Variat. 109: 255-260.. gress LC No. 98090034). Van-Dyck, H. & E. Matthysen. 1998. Thermoregulatory Thomas, J .A. 1984. The conservation of butterflies in tem. differences between phenotypes in the speckled. perate countries: past efforts and lessons for the futu. wood butterfly: Hot perchers and cold patrollers?. re, p. 333-353. In R.L Vane-Wright & P.R. Ackery. Oecologia 114: 326-334.. (eds.). The biology of butterflies. Royal Entomologi cal Society, London.. Van-Dyck, R., E. Matthysen & A.A. Dhondl. 1997a. The ef feet of wing colour on maJe behavioural strategies in. Thompson, J.N. 1988. Variation in preference and specifi. the speckled wood butterfly. Anim. Behav. 53: 39-51.. city in monophagous and oJigophagous swallowtail butterflies. Evolution 42: 118-128.. Van-Dyck, H., E. Matthysen & A.A. Dhondt. 1997b. Ma te-locating strategies are reJated lo relative body. Tuberville, TD., P.G. Dudley & AJ. Pollard. 1996. Res ponses of invertebrate herbivores to stinging tricho. length and wing colour in the speckJed wood but terfly Paral-ge aegeria. Ecol. En!. 22: 116-120.. mes of Urtica dioica and Laportea canadensis. Oi kos 75: 83-88.. Young, A.M. 1983. Egg placement by Phoebis (Pieridae) on Cassia (Leguminosae) "anticipates" the tropical. Vane-Wright, R.L & P.R. Ackery (eds.). 1984. The biology. rainy season. J. Lepid. Soco 37: 313-317.. of butterflies. Royal Entomological Society, London. 429 p. Editor: José A. Vargas Received 2-VI-1999. Corrected 25-liI-2000. Accepted 27-IV-2000..
(10)
Figure


Documento similar
A revision of the methodology for the study of attitudes toward mathematics for the correct validation of the instrument is carried out in this research, and an estimation of
This exploratory study has provided us with information and particulars that not only enable us to have a better understanding of the behavior of our research population – the
In this paper, we study the asymptotic (large time) behavior of a selection-mutation-competition model for a population structured with respect to a phenotypic trait when the rate
No obstante, como esta enfermedad afecta a cada persona de manera diferente, no todas las opciones de cuidado y tratamiento pueden ser apropiadas para cada individuo.. La forma
– Spherical Mexican hat wavelet on the sphere software (developed by the Observational Cosmology and Instrumentation Group at
Assessment of small bowel mucosal healing by video capsule endoscopy for the prediction of short‐term and long‐term risk of Crohn's disease flare: a prospective cohort study..
In this prospective study of patients in a major depressive episode (MDE) and meeting criteria for major depressive disorder (MDD) or bipolar disorder (BD), we
In this study, the Surface Energy Balance Algorithm for Land (SEBAL) and meteorological data from the Global Land Data Assimilation System (GLDAS) were used to estimate long-term
