Wnt signaling in skeletal muscle dynamics: Myogenesis, neuromuscular synapse and fibrosis
16
0
0
Texto completo
(2) Mol Neurobiol (2014) 49:574–589. like cells. All of these factors lead to tissue disorganization and to a progressive failure in muscle regeneration that culminates with a decrease in muscle activity [13, 14]. Recent studies suggest that the activation of Wnt signaling promotes muscle fibrosis [13, 14]. Particularly, the so-called canonical Wnt/βcatenin pathway has been described as profibrotic as it relates with changes in the expression of some ECM components. However, based on the fact that muscle fibers rely in calcium for most of their functions, we will also consider the potential and still unexplored role of noncanonical Wnt/Ca+2 signaling in this process [13–16]. In addition, the suggested profibrotic effect of Wnt signaling could be explained by its interaction with other pathways previously involved in the onset of fibrosis. For instance, the connective tissue growth factor (CTGF) and the transforming growth factor β (TGF-β) signaling cascades have been described in several organs as profibrotic including liver, kidney and skeletal muscle [17–19]. However, the molecular effects of Wnt signaling pathway on the activity of TGF-β and CTGF, as well as the effect of these interactions on the onset of muscle fibrosis are not well understood. The aim of this review is to describe the role of the Wnt pathway on skeletal muscle dynamics including myogenesis, NMJ formation, and muscle fibrosis.. Wnt Signaling Pathways Wnt ligands belong to a conserved family of cysteine rich glycoproteins, which are essential to a variety of biological processes. In humans, 19 Wnt genes have been described, each one with a different expression pattern and function [20–22]. The Wnt signaling can be divided into two types: canonical or βcatenin-dependent and noncanonical or β-catenin-independent pathways (Fig. 1) [21, 23]. Canonical or β-Catenin-Dependent Wnt Signaling Pathway The canonical Wnt pathway begins with the binding of the Wnt ligand to the widely expressed seven-transmembrane Frizzled (Fzd) receptors, of which ten members have been described in vertebrates [24]. The Wnt–Fzd interaction requires the LDL-receptor-related proteins 5/6 (LRP5/6), which act as co-receptors of Fzd [25]. Intracellulary, the canonical Wnt pathway requires the intracellular protein β-catenin. In the absence of Wnt ligands, β-catenin levels remain relatively low by the action of a so-called “destruction complex” formed by the scaffold protein Axin, adenomatous polyposis coli (APC) and the enzyme glycogen synthase kinase 3β (GSK3 β), that phosphorylates β-catenin stimulating its destruction by the proteosomal pathway [25]. Upon Wnt–Fzd interaction, the scaffold protein Dishevelled (Dvl) is recruited, causing the dissociation of the β-catenin destruction complex by a series of phosphorylations that inhibit GSK-3β. Under these conditions,. 575. β-catenin is accumulated in the cytoplasm and then translocated to the nucleus where interacts with the T-cell specific transcription factor (TCF) and the lymphoid enhancer-binding factor (LEF), inducing the expression of Wnt target genes (Fig. 1a) [21, 26, 27]. Noncanonical or β-Catenin-Independent Wnt Signaling Pathway At least two noncanonical pathways are also activated by Wnt ligands. In the planar cell polarity pathway (Wnt/PCP), the Wnt–Fzd dependent recruitment of Dvl leads to activation of small GTPases proteins, such as Rho and Rac, which subsequently activate the c-Jun N-terminal kinase. This protein can either signal to the nucleus and it also can modify the cytoskeleton stability, as it affects the phosphorylation of microtubule associated proteins and is also able to interact with actinregulator proteins (Fig. 1b) [28]. In turn, in the Wnt/Ca+2 pathway, signaling downstream of Dvl stimulates trimeric G proteins and the enzyme phospholipase C, which increase the production of inositol triphosphate (IP3), thus triggering an increase in intracellular Ca+2. As a consequence Ca+2-dependent proteins, such as protein kinase C (PKC), calciumcalmodulin-dependent protein kinase II (CaMKII), and the phosphatase calcineurin are activated. Some of these enzymes regulate the transcription factor NF-AT, promoting the expression of specific target genes (Fig. 1c) [20, 29, 30].. Wnt Signaling in Myogenesis Skeletal muscle tissue in vertebrates originates from the first germination plate of the mesoderm [31, 32]. Normal muscle differentiation involves the expression of several MRFs, such as Myf5, MyoD, and Pax3/7, which participates in the early stages of the patterning of mesoderm-derived multipotential cells [33–35]. The relationship between the Wnt signaling and the myogenesis is supported by in vivo studies using knockout mutants for Fzd receptors as well as for some Wnt ligands (for a comprehensive review, see von Maltzahn et al. [36, 37]). In these studies, the lack of some crucial Wnt effectors led to pronounced tissue damage, and poor muscle development leading to mice death, suggesting that the Wnt pathway is critical for prenatal myogenic development [36]. For instance, in studies using mutant Wnt10b−/−, the activation of canonical Wnt signaling by GSK-3β inhibition or the overexpression of Wnt10b in myoblasts, accelerates myoblast differentiation and promotes muscle development [38, 39]. In prenatal myogenesis, activation of Wnt pathways generates diverse effects. For instance, in chicken embryos, the somite myogenesis in presegmented paraxial mesoderm can be stimulated by activators of Shh pathway together with some Wnt ligands, including Wnt1, Wnt3, and Wnt4. Indeed,.
(3) 576. Mol Neurobiol (2014) 49:574–589. Fig. 1 Wnt signaling pathways. a In the absence of Wnt, GSK-3β protein promotes the phosphorylation and subsequent degradation of β-catenin. In the canonical Wnt signaling, Fzd and LRP 5/6 activation by Wnt ligand binding leads to the accumulation and migration of β-catenin to the nucleus where it could interact with transcription factors LEF/ TCF family and they potentiate transcription of Wnt target genes including Bcl2, axin-2 and ECM components. b Wnt/PCP signaling pathway, the Wnt ligand binding lead the activation of Fzd and Dvl and this complex activated smalls protein G such us Rho and Rac leading to activation of the JNK pathway and its interaction with the cytoskeleton. c Wnt/Ca+2 signaling, the activation of Fzd-Dvl complex lead to the generation of IP3 that activates the IP3 receptor and releases of intracellular Ca+2 , this activate several proteins including CamKII and calcineurin that regulate the gene expression via the transcription factors NF-AT. at early stages of differentiation, the expression of MyoD is dependent on the simultaneous activation of the Wnt and Shh pathways; in later stages of development, however, Wnt activity alone is able to stimulate the expression of MyoD,. suggesting that Wnt signaling is a key factor for late prenatal differentiation [40]. The distribution pattern of Fzd and Wnts ligands in different regions of chick embryo support a crucial role for Wnt signaling in early myogenesis, by showing a.
(4) Mol Neurobiol (2014) 49:574–589. direct correlation between the expression of some receptors and ligands; however, there is a clear overlapping expression pattern in an embryonic region that could activate both canonical and noncanonical Wnts pathways to regulate myogenesis. This includes somites, segmental plate mesoderm, and neural folds [41, 42]. In this regard, even though a crucial role has been well documented for the Wnt/β-catenin pathway during different steps of myogenesis, cumulative evidence suggest that other noncanonical pathways, such as the PCP or an alternative mTOR-dependent pathway, also play a role in the process [5]. Nevertheless, the precise mechanisms by which different Wnt ligands and pathways could interplay to regulate myogenesis require further studies.. Wnt Signaling in the Function of the Neuromuscular Junction The Neuromuscular Junction The vertebrate NMJ is a cholinergic synapse that controls skeletal muscle contraction. Unveiling how this peripheral synapse forms does not only represent a potential benefit to recover movement after pathological or traumatic conditions but also provides valuable information to understand central synapses; indeed, the NMJ has been widely used as an archetypical model to identify signaling molecules secreted at both sides of the synapse that potentially play positive or negative roles to shape functional synapses (for reviews, see [43–45]). An early hallmark of postsynaptic differentiation at the NMJ is the aggregation of several postsynaptic proteins, including the acetylcholine receptors (AChRs), in discrete domains of the sarcolemma [44]. Even though it was originally believed that only nerve-derived molecules induced AChR aggregation, the existence of an early prepattern of AChR clusters before inervation has been demonstrated to be crucial for the subsequent positioning of motor axons for NMJ assembly [46–48]. Later on, upon nerve–muscle contact, most of these prepatterned AChR clusters are disassembled by the inhibitory effect of acetylcholine (ACh), which signals through a Cdk5dependent pathway [49–51]. In turn, those aggregates located in close apposition with the motor axon are stabilized by the anti-inhibitory role of agrin, a motor neuron-derived proteoglycan originally believed to induce the aggregation of postsynaptic proteins [50–52]. Agrin signals through the musclespecific tyrosine kinase receptor MuSK [53–55] forming a membrane complex with a low density lipoprotein receptorrelated protein 4 (Lrp4) [56–58] and the cytosolic proteins rapsyn, which binds to AChRs, as well as to the MuSKbinding proteins Dok-7 and Tid1 [59–61]. In support of their crucial role, mice deficient in MuSK, Lrp4, rapsyn, Dok-7, or Tid1 show no signs of postsynaptic differentiation [53, 56,. 577. 58–61], whereas agrin null mice fail to maintain AChR clusters [47, 51]. In contrast, and according to its inhibitory role, mice rendered unable to synthesize ACh, display more and wider postsynaptic densities than wild-type littermates [50, 51]. The identity and function of proteins regulating the presynaptic differentiation of motor neurons have not been extensively described; however, genetic in vivo studies have identified important regulators of the process. For instance, transgenic mice for members of the ephrin family of bidirectional signaling molecules revealed an essential in vivo role for these axonal guidance regulators on the formation of properly positioned NMJs [62]. Regarding neuromuscular synaptogenesis, it has been shown that whereas FGF signaling induces early presynaptic differentiation, laminin-2 affects the maturation and function of active zones, while collagen IV plays a later maintenance role [63]. Remarkably, mice null for Lrp4 or MuSK, key regulators of postsynaptic assembly, also display defects on presynaptic differentiation [53, 57]. According to this observation, Lrp4 has been recently shown to bind motor axons to induce key features of presynaptic differentiation, such as the clustering of presynaptic and active zone components, through a mechanism independent of MuSK and agrin [64]. Together, the confluence of multiple signals originated at both sides of the synapse refine the building of functional NMJs. Wnt Signaling on Postsynaptic Development at the Vertebrate NMJ Several lines of evidence have shown that Wnt pathways affect postsynaptic differentiation at the vertebrate NMJ in vivo. For instance, chick muscles exposed to the Wnt-binding inhibitor secreted Frizzled related protein (Sfrp1) display impaired AChR clustering [65], suggesting that endogenous Wnt ligands regulate postsynaptic assembly. In support of this notion, mutant mice for Dvl, a common mediator of several Wnt pathways, display abnormal postsynaptic development at the NMJ [65], a phenotype consistent with previous in vitro data showing that Dvl regulated the function of MuSK to induce AChR clustering [66]. In zebrafish, the Wnt11r ligand, expressed by tissues adjacent to the newly formed muscle fibers, interacts with the ligand-binding domain of MuSK to induce the prepatterning of AChR clusters and the guidance of motor axons [46] (Fig. 2a). Accordingly, mutant fish for Wnt11r or MuSK display similar severe defects in AChR prepatterning and axonal branching [46, 67]. Consistent with these findings, the mouse muscle-derived ligand Wnt4 binds to and phosphorylates MuSK [68]. Wnt4 null mice have less AChR than control littermates just before inervation, whereas cultured muscle cells exposed to Wnt4 display increased AChR clustering [68]. Similarly, the mouse-derived ligands Wnt9a and Wnt11 induce AChR clustering in cultured myotubes through a mechanism dependent on MuSK and.
(5) 578. Mol Neurobiol (2014) 49:574–589. Fig. 2 Wnt signaling in the vertebrate neuromuscular synaptogenesis. a Wnts induce aneural AChR clustering. Several muscle-derived Wnt ligands activate MuSK-dependent AChR clustering in cultured myotubes. In zebrafish, Wnt11r and Wnt4a induce the internalization of MuSK to endosomes located in the middle region of the myofiber. MuSK assemble a complex with the scaffolding proteins diversin, Daam1 and RhoA to position aneural AChR clusters in a central muscle band (gray stripe), which will guide the incoming motor axons for subsequent NMJ assembly. b. Wnts are positive signals for postsynaptic differentiation. Wnt3 and agrin released from the presynaptic terminal collaborate to promote the formation of AChR clusters. Wnt3 induces the formation of AChR microclusters via Rac1, which are aggregated into fullsize clusters by the Rhodependent effect of agrin. c Wnt/ β-catenin pathways inhibit AChR clustering but promote presynaptic behavior. Wnt3a, secreted by muscle cells at the stages of NMJ formation, activates a β-catenin pathway that induces the dispersal of AChR clusters through the inhibition of rapsyn expression. Specific ablation or stabilization of βcatenin in muscles, but not in motor neurons, result in presynaptic defects, suggesting that muscle β-catenin induces the expression of a still unknown retrograde signal for presynaptic differentiation. Lrp4 [69]. Remarkably, recent findings obtained in zebrafish show that Wnt11r and Wnt4a induce a localized endocytosis of MuSK to recycling endosomes which, in turn, accumulates AChRs at the sites where motor axons will be guided to assemble functional NMJs [70] (Fig. 2a). Together, these. findings reveal a novel key MuSK-dependent mechanism by which Wnt ligands induce the aneural clustering of AChRs on newly formed muscle cells. Wnt ligands can also regulate neuromuscular synaptogenesis [12, 71, 72]. On the one hand, transplantation of chick wings.
(6) Mol Neurobiol (2014) 49:574–589. with cells secreting the Wnt3 ligand, which is expressed by motor neurons at the time of NMJ formation [73], led to increased AChRs clustering [65]. In cultured myotubes, Wnt3 activates the small GTPase Rac1 to induce the formation of AChR microclusters, which only coalesce into bigger clusters after agrin-dependent activation of Rho [65] (Fig. 2b). These findings reveal a potential neural-dependent cross-talk of Wntand agrin-mediated pathways to induce postsynaptic differentiation at the vertebrate NMJ. On the other hand, the highly identical ligand Wnt3a impairs agrin-induced AChR clustering and disassemble preformed aggregates [74, 75]. Wnt3a is expressed by developing skeletal muscles and mediates its dispersal activity by down-regulating rapsyn expression via a β-catenin-dependent, but TCF-independent, pathway [75] (Fig. 2c). Even though these findings reveal an inhibitory effect for β-catenin on AChR clustering, the in vivo role of this crucial effector of the Wnt canonical pathway at the NMJ is rather complex. Thus, even though specific ablation of βcatenin in skeletal muscles, but not in motor neurons, gives rise to enlarged AChR clusters, this effect is primarily related to presynaptic defects in axonal branching and neurotransmission [74, 76]. In turn, specific stabilization of β-catenin in muscle but not in neuronal cells, resulted in excessive nerve branching and defasciculation, while NMJ formation or function were unaffected [77] (Fig. 2c). Taken together, these findings reveal that activation of different Wnt pathways control opposite but complementary roles on the assembly of postsynaptic densities at nascent neuromuscular synapses. Wnt Signaling on Presynaptic Differentiation Even though our current knowledge regarding the potential role of Wnt signaling on presynaptic differentiation of vertebrate motor neurons is virtually null, the function of diverse Wnt pathways on several steps of presynaptic behavior in central synapses, from the establishment of neuronal cell polarity to synaptogenesis, has been well documented. In primary cultures of hippocampal neurons, it has been described that Wnt5a actives axonal differentiation in a βcatenin-independent manner via the activation of the protein complex Par6-Par3-atypical protein kinase C (aPKC) [78, 79]. Wnt5a signaling is also involved in regulating polarity via aPKC activation in a Dvl-dependent manner; however, in Drosophila , aPKC mutants do not show failures in neuronal polarity [80]. Other studies indicate that gradients of Wnt1 and Wnt5a expressed in the dorsal spinal cord in an anterior– posterior direction attracts ascending somatosensory axons projecting from the spinal cord to the brain, while the same gradient repels descending corticospinal tracts in the opposite direction, from the brain to the spinal cord [81, 82]. In addition, the repulsive signal of Wnt/Ca+2 signaling has been reported in mutants for CaMKII; indeed, this mutation causes severe failures in axonal guidance given the lack of a repulsive. 579. Wnt mediated signal [83]. Together, these data support the participation of Wnt signaling in the axonal guidance with repulsive and attractive effects depending of the Wnt ligand (Table 1). However, the molecular mechanism of the effect and the interaction networks of Wnt signaling are still to be fully underscored. Regarding presynaptic differention, Wnt7a stimulates the maturation of neuronal connections and the presence of the Wnt antagonist sFRP-1 blocks the growth of axons in the cerebellum [84–86]. Similarly, in hippocampal neurons, Wnt7a stimulates the clustering of the presynaptic protein synaptophysin and increases the mEPSC frequency [87]. Altogether, these data expose the critical role of Wnt signaling on presynaptic assembly at the central nervous system, modulating several processes, which lead to the correct formation of synapses (Table 2). However, the advance in the central synapse is not proportional to our knowledge of the differentiation of the presynaptic component at the vertebrate NMJ.. Wnt Signaling in Muscle Fibrosis Fibrosis in Skeletal Muscle Fibrosis is characterized by the aberrant deposition of ECM components including fibronectin and collagen type I or III. This accumulation of ECM components leads to the tissue disorganization and to the loss of muscle activity and eventually to death [15, 88, 89]. Several organs are susceptible to fibrosis including lung, kidney, liver, and skeletal muscle; however, all fibrotic reactions share common underlying cellular and molecular mechanisms, including tissue degeneration, macrophage infiltration, inflammation, and proliferation of fibroblast-like cells [15, 89–93]. The loss of organ architecture and the activation of leukocytes and fibroblasts increase the production of several molecules, such as growth factors, proteolytic enzymes, angiogenic factors, and fibrogenic cytokines. Together, these molecules lead to the accumulation of ECM components [16, 94, 95]. Table 1 Effect of Wnt ligands in axonal guidance Type of signal. Wnt1 Wnt3 Wnt4 Wnt5 Wnt5a Wnt7b Wnt8b Wnt11r. References. Attractive. Repulsive. + + + +. +. + + +. + +. [81, 155] [49] [54] [56, 155] [83] [76] [61] [46].
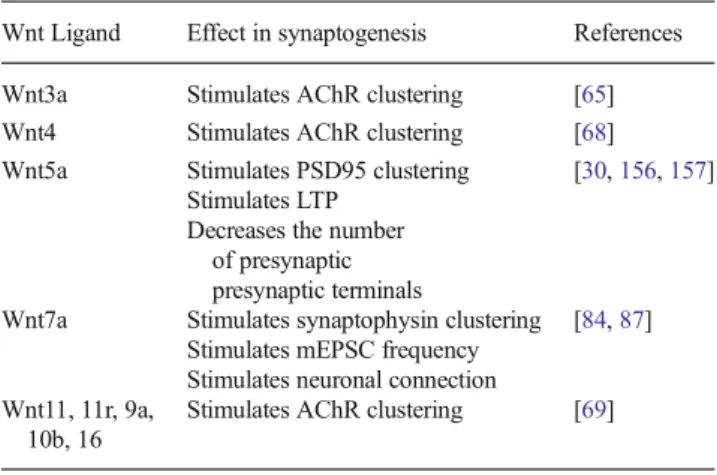
(7) 580. Mol Neurobiol (2014) 49:574–589. Table 2 Effect of Wnt ligands in synaptogenesis Wnt Ligand. Effect in synaptogenesis. References. Wnt3a Wnt4 Wnt5a. Stimulates AChR clustering Stimulates AChR clustering Stimulates PSD95 clustering Stimulates LTP Decreases the number of presynaptic presynaptic terminals Stimulates synaptophysin clustering Stimulates mEPSC frequency Stimulates neuronal connection Stimulates AChR clustering. [65] [68] [30, 156, 157]. Wnt7a. Wnt11, 11r, 9a, 10b, 16. [84, 87]. [69]. Several models have been described to study the regenerative capacity of the skeletal muscle, almost all the models use acute injury induced by injection of a toxin or myofiber destruction. These studies have allowed the description of all stages of the regenerative process including the inflammatory response, which follows an ordered pattern [96–98]. Early after injury, the damaged area is infiltrated by inflammatory cells. In the mdx mice, the model animal for Duchenne's muscle dystrophy (DMD, see below), these inflammatory cells are critical to promote the survival and proliferation of myogenic precursor cells, promoting the repair of the skeletal muscle [16, 95, 99, 100]. The innate immune response is activated by the release to the extracellular space of several factors from the damaged fiber. These factors lead to the infiltration of the damage area by monocytes and neutrophils [99, 101]. The neutrophils play a key role in repair facilitating phagocytosis and the recruitment of monocytes by the release of cytokines [102]. Proinflammatory macrophages, observed experimentally in the context of muscle repair, are phenotypically similar to classically activated M1 macrophages and are usually found at early stages after muscle injury. These cells also release proinflammatory cytokines, such as interleukin-1 beta (IL-1β) and tumor necrosis factor alpha (TNF-α). In a chronic muscle damage, the M1 macrophages release several ECM remodelers like urokinase-type plasminogen activator (uPA), plasminogen activator inhibitor-1 (PAI-1), plasmin, matrix metalloproteinases and tissue inhibitors of metalloproteinases (TIMPs). The role of these molecules is to modulate the proliferation of myoblasts and to promote the excessive ECM production/accumulation and the replacement of muscle fiber with fibrotic tissue [103, 104]. After the activation of M1 macrophages, it has been described the activation of the M2c phenotype, so-called because of their role in deactivating M1 macrophages. These cells release antiinflammatory factors including the interleukin-10 (IL-10). The activity of these cells is important to regulate the end of the inflammatory response [105]. In skeletal muscle, fibrosis is commonly related with muscle dystrophies, a molecularly heterogeneous group of diseases,. characterized by skeletal muscle wasting, weakness, decrease in the fiber size, and muscle necrosis that compromises patient mobility, leading to death [106–108]. One of these diseases is the DMD [15, 91, 109]. The clinical manifestation of DMD includes an onset during childhood with progressive weakness and death in early adulthood [91, 108]. The DMD is caused by a failure in the gene coding for the cytoskeletal protein dystrophin, which is required for the proper interaction between the plasma membrane and the ECM. The absence of dystrophin leads to the loss of function in the normal muscle regeneration cycle, leading to an increase in the production of ECM components [107, 108, 110]. In normal conditions, the skeletal muscle have a great capacity to repair itself after injury; however, under chronic injury conditions such as DMD, the skeletal muscle progressively loses its capacity to regenerate, likely by the decrease in the number of satellite cells, the skeletal muscle stem cells, which are unable to repair the damaged tissue. As a consequence, the muscle tissue is progressively replaced by adipose and especially fibrotic tissue [4, 95, 111, 112]. Currently, no effective clinical treatment to combat or attenuate muscle fibrosis in DMD patients is available; however, recent studies using the mdx mouse have focused more attention on elucidating the cellular and molecular mechanisms underlying skeletal muscle fibrosis associated with dystrophin deficiency. These studies have tested several pharmacological agents that target muscle fibrosis and strongly suggest that combating fibrosis could ameliorate DMD progression and increase the success of new cell- and gene-based therapies [113, 114]. In fibrotic skeletal muscle, several pathways are turning on by different cells such as fibroblasts, macrophages and leukocytes. Despite the complex mixture of signaling implicated in the onset of skeletal muscle fibrosis until now two key molecules have been described as critical in the development of the disease: the TGF-β and the CTGF [15, 17, 110, 115–117]. TGF-β is a potent inducer of CTGF, and most models postulate that CTGF acts as a downstream mediator of TGF-β activity; by contrast, other studies support the independent action of each molecule [19, 118–121]. TGF-β Signaling in Fibrosis and its Possible Cross-Talk with the Wnt Signaling There are three TGF-β isoforms, TGF-β1, TGF-β2, and TGF-β3. The activation of these signaling relies on the binding of the ligand to a heterodimeric receptor complex which includes one TGF-β type I receptor molecule, termed activin linked kinase 5 (ALK5) [122–124]. In the canonical TGF-β signaling, ligand binding leads ALK5 to phosphorylate Smad2/3, which in turn, activates Smad4 and this protein complex translocates into the nucleus to activate transcription factors and lead to the expression of target genes including fibronectin, CTGF and PAI-1 (Fig. 3) [122, 123]..
(8) Mol Neurobiol (2014) 49:574–589. 581. has been studied, it is possible a therapeutic approach, as the inhibition of this signaling has been reported to decrease the development of fibrosis. The relationship between canonical Wnt and TGF-β pathways has been described in several fibrotic models, and the evidence suggests a collaboration of both pathways in the fibrotic disease; indeed, the inhibition of both pathways prevents the development of fibrosis (Fig. 5) [131, 132]. Recent studies in vivo have shown that activation of the canonical Wnt pathway is required for the action of TGF-β, since the presence of dickkopf 1 (DKK1) (a Wnt signaling inhibitor) decreases the activity of TGF-β in a model of muscle fibrosis (Table 3) [123]. In this regard, in studies using chondrocytes as models, it has been reported that the TGF-β signaling inhibits the expression of axin-2, and this leads to an increases in the levels of β-catenin. This could be a negative feedback for the TGF-β signaling through the inhibition of Smad3 protein [133]. These data suggest a direct relationship between the TGF-β and Wnt/ β-catenin pathways in the establishment of the fibrotic disease, and that the effects of TGF-β needs the activation of the canonical Wnt signaling in the progress of the disease. Cross-Talk Between Wnt Signaling and CTGF in a Model of Muscle Disease Fig. 3 TGF-β signaling. Active TGF-β binds to its receptor type I/II this leads to the activation, via phosphorylation of Smad2/3 pathways, which regulates the transcription of several target genes involved in several cellular functions like CTGF and ECM proteins. TGF-β can also activate other signaling in Smad-independent manner. The expression of TGF-β has been described in normal skeletal muscle, mdx models and patients with DMD. Also, it has been described an over-expression of these ligands after muscle injury [125]. In skeletal muscle cells, TGF-β acts as a strong myogenic inhibitor. It is also known that TGF-β can inhibit myoblast differentiation in vitro, affecting the expression of muscle proteins, such as myosin heavy chain and creatine kinase [126, 127]. The profibrotic effect of TGF-β could be explained by the stimulation of fibroblasts to produce ECM proteins such as collagen and fibronectin. Furthermore, it has been described that the activation of TGF-β signaling leads to a decrease in the expression of metalloprotease like collagenase and to an increase in the expression of TIMPs. Together these effects could lead to the accumulation of ECM [121, 123, 124, 128, 129]. In fact, the injection of TGF-β in vivo in skeletal muscle stimulates the production of ECM in the injected area and the overexpression of TGF-β in myoblasts promotes the formation of myofibroblast cells, suggesting a critical role of the TGF-β in the onset of the fibrotic condition in the skeletal muscle [110, 130]. Since the critical role of these pathways. CTGF is a cysteine rich 36–38 kDa secreted protein, which belongs to CCN family (acronym for connective tissue growth factor, cysteine-rich-protein, and nephroblastoma-overexpressed). CTGF is expressed in various cell types including fibroblasts, chondrocytes, and leukocytes and is poorly expressed in the central nervous system [18, 134, 135]. The CTGF gene contains five exons. Exon 1 encodes a signal peptide (SP) for secretion, while exons 2–5 encode for modules 1–4 of the secreted CTGF protein which are homologous to other proteins including insulin-like growth factor binding protein (IGFBP) domain (module 1), a von Willebrand factor type C (VWC) domain (module 2), a thrombospondin type 1 (TSP1) domain (module 3), and a C-terminal (CT) module (Fig. 4) [134–136]. The modular structure of CTGF allows the interaction of CTGF with multiple molecules suggesting a role in several signaling downstream, including Notch1, LRPs, and growth factors such as BMP, TGF-β, and vascular endothelial growth factor. The interaction of CTGF and the receptors described previously allows the activation of several pathways downstream including PKC, MEK/ERK, and protein kinase B (PKB) [19, 137–140]. In physiological conditions, CTGF is involved in angiogenesis and cellular differentiation, but in pathological conditions, it has been described as a profibrotic molecule, stimulating the proliferation of fibroblasts, their differentiation towards myofibroblasts and ECM production [110, 115, 121, 134, 141]. In fact, in vivo studies have showed that the CTGF.
(9) 582. Mol Neurobiol (2014) 49:574–589. Table 3 Effect of different signaling components on fibrosis. Effect over fibrosis Decrease. Tissue. References. Liver/muscle/kidney/lung Liver/muscle/kidney/lung Muscle/kidney Muscle. [18, 125, 129] [19, 134] [17, 142, 143] [14, 147]. Kidney/liver Mammary epithelial cells Heart Chondrocytes. [158, 159] [160] [161] [144]. Dermal fibrosis Dermal fibrosis. [162] [163]. Increase. Treatment or drug TGF-β CTGF TGF-β+ CTGF Wnt3a Classical Wnt/β-catenin inhibitors DKK1 + sFRP1 + sFRP2 + Wif-1 + GSK-3β inhibitors PKF118-310 XAV-939. + + ++ ++. + +. injection stimulates the fibrosis and the coinjection of CTGF and TGF-β resulted in a persistent fibrosis [142, 143]. Together, this has led to the notion that CTGF plays an important role in tissue response to injury and fibrosis (Fig. 5). In chondrocytes, CTGF binds the Wnt inhibitory factor 1 (WIF-1), an antagonist of the Wnt pathway, through the CT domain of CTGF. It has been also described a direct relationship in the expression patterns of CTGF and WIF-1, and the presence of WIF-1 inhibits the expression of CTGF; however, CTGF is unable of interfering with the effect of WIF-1 on the canonical Wnt signaling [144]. Furthermore, it has been described that CTGF can also interact with the CT domain of LRP6, so CTGF is also able to affect the interaction of Wnt with their coreceptor [145, 146]. Other studies in human mesangial cells have been described suggesting that CTGF stimulates the canonical Wnt pathway in an LRP6-dependent manner, and this effect is inhibited in the presence of DKK1 (Table 3) [145]. Remarkably, studies in a fibroblast cell line show that the treatment with the Wnt3a ligand, which signals through the Wnt/β-catenin pathway, increases the expression of CTGF and TGF-β mRNAs, reinforcing the view that these signaling pathways may crosstalk in the context of fibrosis [147].. Fig. 4 CTGF signaling. The CTGF protein contains a signal peptide (SP) as well as four modules: IGFBP, VWC, TSP1, and CT. Through these domains CTGF can interact with several molecules including BMP, TGFβ, and possibly some Wnt ligands. CTGF interacts with several membrane proteins, including LRP proteins, integrins, TrkA, BMP, and Notch 1; these interactions trigger various pathways downstream including PKC, MEK, p38, and PKB JNK, and the activation of these signaling lead to several effects like proliferation, fibroblast differentiation, and production of ECM components, leading to the onset of fibrotic condition. Proposed Model for the Interaction Between Wnt/β-catenin, CTGF, and TGF-β Signaling in the Development of Skeletal Muscle Fibrosis The data presented above support an important role for the Wnt/β-catenin signaling pathway during the onset and development of the fibrotic condition in skeletal muscle. The effect of the Wnt pathway could be explained at least in part by the relationship between this signaling cascade with the signaling mediated by TGF-β and CTGF [17, 128, 144]. In addition,.
(10) Mol Neurobiol (2014) 49:574–589. 583. Fig. 5 Cross-talk between Wnt and TGF-β signaling. In fibrotic condition, the levels of TGF-β are increased, and this promotes the stimulation of the target genes of this pathway, like CTGF and ECM components such as type I collagen; this promote the accumulation of connective tissue in the extracellular space, the transcription effect of the TGF−β signaling, leading a down-regulation of axin-2 a protein part of the. complex destruction of β-catenin, and the activation of TGF-β leads to an increase in cytoplasmic β-catenin and an activation of canonical Wnt signaling and an increase in their target genes including fibronectin, leading to a major accumulation of ECM components promoting the development of muscle fibrosis. Wnt signaling could modulate early processes related to the regeneration of skeletal muscles after injury, and, in this way, it could trigger a deregulation of the regenerative capacity of the muscle, promoting the onset of fibrosis [14, 148]. Therefore, the activation of the Wnt/β-catenin signaling could stimulate the formation and activation of fibroblasts into myofibroblast derived from the satellite cells activated by injury. The activated fibroblasts could increase the expression and release of TGF-β to the environment. In turn, this ligand, in a autocrine form, could increase the expression of CTGF and together these ligands could activate downstream signaling leading to the over-expression of ECM components like fibronectin [149–152]. Furthermore, the activation of Wnt/β-catenin could itself stimulate the expression of fibronectin, and also there are. some reports suggesting that the CTGF could be a possible target gene of the canonical Wnt signaling [153]. Therefore, the Wnt/β-catenin signaling pathway could have a key role in triggering the activation of TGF-β and CTGF in activated fibroblasts and maybe in other cells of the fibrotic environment. The presence of several Wnt/β-catenin inhibitors, such as DKK1 and Sfrp1 and 2 inhibit the signaling of TGF-β and CTGF, leading to an inhibition of the fibrosis (Table 3 and Fig. 6). This new role of the Wnt/β-catenin signaling as a profibrotic factor makes this signaling pathway an interesting candidate for therapeutic interventions. In fact, there is evidence describing that inhibition of the Wnt/β-catenin pathway decreases the progression of the fibrosis [154]. Certainly, further studies are.
(11) 584. Mol Neurobiol (2014) 49:574–589. Fig. 6 Proposed model for the role of Wnt signaling in Fibrosis. After acute muscle injury, the damage area is infiltrated by macrophages and neutrophils; these cells release several cytokines and lead to the activation of the inflammation process. In normal postinjury muscle regeneration, these inflammation is acute and important for the correct replacement of the muscle fiber. In a second step, the satellite cells are activated and star the differentiation/proliferation to myoblast, and finally, these cells star the fusion process to form a new muscle fiber; in these normal cycle of regeneration has been described a low expression of Wnt ligands and a low activity of the Wnt/β-catenin pathway. By contrast, in a chronic muscle injury like DMD, the inflammation process is deregulated, and these lead to the deregulation of the regeneration processes stimulating. the differentiation of the fibroblast into myofibroblast. The activated myofibroblast cells increase the production of ECM components like fibronectin and collagen and also release profibrotic molecules like CTGF and TGF-β. In these chronic damage condition has been described an increase in the expression of Wnt ligands, and these lead to the activation of the Wnt/β-catenin pathway in the myofibroblast increasing the production of the ECM components and the release of profibrotic molecules; together, this lead to aberrant regeneration. Several inhibitors of Wnt/βcatenin signaling like DKK1 and Wif-1 block the TGF-β and CTGF profibrotic effect, suggesting the central role of the Wnt signaling in the progress of fibrosis. necessary to better define the effect of Wnt signaling inhibition on skeletal muscle fibrosis.. formation of the motoneuron-muscle synapse. In myogenesis, the effect of Wnt signaling leads to the progression of the differentation at early developmental stages and inhibition of this signaling leads to a poor skeletal muscle formation. In the maturation of the NMJ, the Wnt signaling modulates the localization of several proteins critical for the patterning of the synapse and for the precise control of the muscle contraction. In muscle fibrosis, apparently the Wnt signaling plays a critical. General Conclusions The Wnt signaling pathway plays a critical role in several processes including the development of muscle tissue and the.
(12) Mol Neurobiol (2014) 49:574–589. role, as it is able to functionally interact with other profibrotic molecules, such as CTGF and TGF-β. Understanding these interactions could be critical in the discovery of future treatments for this pathological condition. Acknowledgments This work was supported by grants from the Basal Center of Excellence in Aging and Regeneration (CONICYT-PFB 12/ 2007) and FONDECYT (no. 1120156 to N.C.I. and no. 1130321 to J.P.H.). PC was a postdoctoral fellow from the PFB (12/2007) grant to NCI and EB. Graphic work was carried out by Graphique-Science (http:// graphique-science.blogspot.com). Conflicts of Interest There is no conflict of interest.. References 1. Burden SJ (1998) The formation of neuromuscular synapses. Genes Dev 12(2):133–148 2. Fallon JR, Gelfman CE (1989) Agrin-related molecules are concentrated at acetylcholine receptor clusters in normal and aneural developing muscle. J Cell Biol 108(4):1527–1535 3. Polesskaya A, Seale P, Rudnicki MA (2003) Wnt signaling induces the myogenic specification of resident CD45+ adult stem cells during muscle regeneration. Cell 113(7):841–852 4. Rudnicki MA, Le Grand F, McKinnell I, Kuang S (2008) The molecular regulation of muscle stem cell function. Cold Spring Harb Symp Quant Biol 73:323–331 5. von Maltzahn J, Chang NC, Bentzinger CF, Rudnicki MA (2012) Wnt signaling in myogenesis. Trends Cell Biol 22(11):602–609 6. Moody-Corbett F (1986) Formation of the vertebrate neuromuscular junction. Dev Biol 2:605–635, N Y 1985 7. Daniels MP (1997) Intercellular communication that mediates formation of the neuromuscular junction. Mol Neurobiol 14(3):143– 170 8. Hughes BW, Kusner LL, Kaminski HJ (2006) Molecular architecture of the neuromuscular junction. Muscle Nerve 33(4):445–461 9. Aldunate R, Casar JC, Brandan E, Inestrosa NC (2004) Structural and functional organization of synaptic acetylcholinesterase. Brain Res Brain Res Rev 47(1–3):96–104 10. Park M, Shen K (2012) WNTs in synapse formation and neuronal circuitry. EMBO J 31(12):2697–2704 11. Budnik V, Salinas PC (2011) Wnt signaling during synaptic development and plasticity. Curr Opin Neurobiol 21(1):151–159 12. Henriquez JP, Salinas PC (2011) Dual roles for Wnt signalling during the formation of the vertebrate neuromuscular junction. Acta Physiol (Oxf). doi:10.1111/j.1748-1716.2011.02295.x 13. Brack AS, Conboy IM, Conboy MJ, Shen J, Rando TA (2008) A temporal switch from notch to Wnt signaling in muscle stem cells is necessary for normal adult myogenesis. Cell Stem Cell 2(1):50–59 14. Brack AS, Conboy MJ, Roy S, Lee M, Kuo CJ, Keller C, Rando TA (2007) Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science 317(5839):807–810 15. Bitterman PB, Henke CA (1991) Fibroproliferative disorders. Chest 99(3 Suppl):81S–84S 16. Serrano AL, Munoz-Canoves P (2010) Regulation and dysregulation of fibrosis in skeletal muscle. Exp Cell Res 316(18):3050–3058 17. Wang Q, Usinger W, Nichols B, Gray J, Xu L, Seeley TW, Brenner M, Guo G, Zhang W, Oliver N, Lin A, Yeowell D (2011) Cooperative interaction of CTGF and TGF-beta in animal models of fibrotic disease. Fibrogenesis Tissue Repair 4(1):4 18. Leask A, Abraham DJ (2004) TGF-beta signaling and the fibrotic response. FASEB J 18(7):816–827. 585 19. Abreu JG, Ketpura NI, Reversade B, De Robertis EM (2002) Connective-tissue growth factor (CTGF) modulates cell signalling by BMP and TGF-beta. Nat Cell Biol 4(8):599–604 20. Nusse R, Varmus H (2012) Three decades of Wnts: a personal perspective on how a scientific field developed. EMBO J 31(12): 2670–2684 21. Clevers H, Nusse R (2012) Wnt/beta-catenin signaling and disease. Cell 149(6):1192–1205 22. Gordon MD, Nusse R (2006) Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors. J Biol Chem 281(32):22429–22433 23. Toledo EM, Colombres M, Inestrosa NC (2008) Wnt signaling in neuroprotection and stem cell differentiation. Prog Neurobiol 86(3): 281–296 24. Wang HY, Liu T, Malbon CC (2006) Structure–function analysis of Frizzleds. Cell Signal 18(7):934–941 25. Niehrs C (2012) The complex world of WNT receptor signalling. Nat Rev Mol Cell Biol 13(12):767–779 26. Arrazola MS, Varela-Nallar L, Colombres M, Toledo EM, Cruzat F, Pavez L, Assar R, Aravena A, Gonzalez M, Montecino M, Maass A, Martinez S, Inestrosa NC (2009) Calcium/calmodulin-dependent protein kinase type IV is a target gene of the Wnt/beta-catenin signaling pathway. J Cell Physiol 221(3):658–667 27. Hodar C, Assar R, Colombres M, Aravena A, Pavez L, Gonzalez M, Martinez S, Inestrosa NC, Maass A (2010) Genome-wide identification of new Wnt/beta-catenin target genes in the human genome using CART method. BMC genomics 11:348 28. Rosso SB, Sussman D, Wynshaw-Boris A, Salinas PC (2005) Wnt signaling through Dishevelled, Rac and JNK regulates dendritic development. Nat Neurosci 8(1):34–42 29. Inestrosa NC, Montecinos-Oliva C, Fuenzalida M (2012) Wnt signaling: role in Alzheimer disease and schizophrenia. J Neuroimmune Pharmacol 7(4):788–807 30. Varela-Nallar L, Alfaro IE, Serrano FG, Parodi J, Inestrosa NC (2010) Wingless-type family member 5A (Wnt-5a) stimulates synaptic differentiation and function of glutamatergic synapses. Proc Natl Acad Sci U S A 107(49):21164–21169 31. Manceau M, Marcelle C, Gros J (2005) A common somitic origin for embryonic muscle progenitors. Med Sci (Paris) 21(11):915–917 32. Gros J, Manceau M, Thome V, Marcelle C (2005) A common somitic origin for embryonic muscle progenitors and satellite cells. Nature 435(7044):954–958 33. Zammit PS, Partridge TA, Yablonka-Reuveni Z (2006) The skeletal muscle satellite cell: the stem cell that came in from the cold. J Histochem Cytochem 54(11):1177–1191 34. Olguin HC, Pisconti A (2012) Marking the tempo for myogenesis: Pax7 and the regulation of muscle stem cell fate decisions. J Cell Mol Med 16(5):1013–1025 35. Tsivitse S (2010) Notch and Wnt signaling, physiological stimuli and postnatal myogenesis. Int J Biol Sci 6(3):268–281 36. van Amerongen R, Berns A (2006) Knockout mouse models to study Wnt signal transduction. Trends Genet 22(12):678–689 37. Chien AJ, Conrad WH, Moon RT (2009) A Wnt survival guide: from flies to human disease. J Invest Dermatol 129(7):1614–1627 38. Vertino AM, Taylor-Jones JM, Longo KA, Bearden ED, Lane TF, McGehee RE Jr, MacDougald OA, Peterson CA (2005) Wnt10b deficiency promotes coexpression of myogenic and adipogenic programs in myoblasts. Mol Biol Cell 16(4):2039–2048 39. Miyoshi K, Rosner A, Nozawa M, Byrd C, Morgan F, LandesmanBollag E, Xu X, Seldin DC, Schmidt EV, Taketo MM, Robinson GW, Cardiff RD, Hennighausen L (2002) Activation of different Wnt/beta-catenin signaling components in mammary epithelium induces transdifferentiation and the formation of pilar tumors. Oncogene 21(36):5548–5556 40. Munsterberg AE, Kitajewski J, Bumcrot DA, McMahon AP, Lassar AB (1995) Combinatorial signaling by Sonic hedgehog and Wnt.
(13) 586 family members induces myogenic bHLH gene expression in the somite. Genes Dev 9(23):2911–2922 41. Borello U, Buffa V, Sonnino C, Melchionna R, Vivarelli E, Cossu G (1999) Differential expression of the Wnt putative receptors Frizzled during mouse somitogenesis. Mech Dev 89(1–2):173–177 42. Cauthen CA, Berdougo E, Sandler J, Burrus LW (2001) Comparative analysis of the expression patterns of Wnts and Frizzleds during early myogenesis in chick embryos. Mech Dev 104(1–2):133–138 43. Kummer TT, Misgeld T, Sanes JR (2006) Assembly of the postsynaptic membrane at the neuromuscular junction: paradigm lost. Curr Opin Neurobiol 16(1):74–82 44. Sanes JR, Lichtman JW (2001) Induction, assembly, maturation and maintenance of a postsynaptic apparatus. Nat Rev Neurosci 2(11): 791–805 45. Wu H, Xiong WC, Mei L (2010) To build a synapse: signaling pathways in neuromuscular junction assembly. Development 137(7):1017–1033 46. Jing L, Lefebvre JL, Gordon LR, Granato M (2009) Wnt signals organize synaptic prepattern and axon guidance through the zebrafish unplugged/MuSK receptor. Neuron 61(5):721–733 47. Lin W, Burgess RW, Dominguez B, Pfaff SL, Sanes JR, Lee KF (2001) Distinct roles of nerve and muscle in postsynaptic differentiation of the neuromuscular synapse. Nature 410(6832):1057–1064 48. Vock VM, Ponomareva ON, Rimer M (2008) Evidence for muscledependent neuromuscular synaptic site determination in mammals. J Neurosci 28(12):3123–3130 49. An MC, Lin W, Yang J, Dominguez B, Padgett D, Sugiura Y, Aryal P, Gould TW, Oppenheim RW, Hester ME, Kaspar BK, Ko CP, Lee KF (2010) Acetylcholine negatively regulates development of the neuromuscular junction through distinct cellular mechanisms. Proc Natl Acad Sci U S A 107(23):10702–10707 50. Lin W, Dominguez B, Yang J, Aryal P, Brandon EP, Gage FH, Lee KF (2005) Neurotransmitter acetylcholine negatively regulates neuromuscular synapse formation by a Cdk5-dependent mechanism. Neuron 46(4):569–579 51. Misgeld T, Kummer TT, Lichtman JW, Sanes JR (2005) Agrin promotes synaptic differentiation by counteracting an inhibitory effect of neurotransmitter. Proc Natl Acad Sci U S A 102(31): 11088–11093 52. Gautam M, Noakes PG, Moscoso L, Rupp F, Scheller RH, Merlie JP, Sanes JR (1996) Defective neuromuscular synaptogenesis in agrin-deficient mutant mice. Cell 85(4):525–535 53. DeChiara TM, Bowen DC, Valenzuela DM, Simmons MV, Poueymirou WT, Thomas S, Kinetz E, Compton DL, Rojas E, Park JS, Smith C, DiStefano PS, Glass DJ, Burden SJ, Yancopoulos GD (1996) The receptor tyrosine kinase MuSK is required for neuromuscular junction formation in vivo. Cell 85(4): 501–512 54. Glass DJ, Bowen DC, Stitt TN, Radziejewski C, Bruno J, Ryan TE, Gies DR, Shah S, Mattsson K, Burden SJ, DiStefano PS, Valenzuela DM, DeChiara TM, Yancopoulos GD (1996) Agrin acts via a MuSK receptor complex. Cell 85(4):513–523 55. Valenzuela DM, Stitt TN, DiStefano PS, Rojas E, Mattsson K, Compton DL, Nunez L, Park JS, Stark JL, Gies DR et al (1995) Receptor tyrosine kinase specific for the skeletal muscle lineage: expression in embryonic muscle, at the neuromuscular junction, and after injury. Neuron 15(3):573–584 56. Kim N, Stiegler AL, Cameron TO, Hallock PT, Gomez AM, Huang JH, Hubbard SR, Dustin ML, Burden SJ (2008) Lrp4 is a receptor for Agrin and forms a complex with MuSK. Cell 135(2):334–342 57. Weatherbee SD, Anderson KV, Niswander LA (2006) LDLreceptor-related protein 4 is crucial for formation of the neuromuscular junction. Development 133(24):4993–5000 58. Zhang B, Luo S, Wang Q, Suzuki T, Xiong WC, Mei L (2008) LRP4 serves as a coreceptor of agrin. Neuron 60(2):285–297. Mol Neurobiol (2014) 49:574–589 59. Linnoila J, Wang Y, Yao Y, Wang ZZ (2008) A mammalian homolog of Drosophila tumorous imaginal discs, Tid1, mediates agrin signaling at the neuromuscular junction. Neuron 60(4):625–641 60. Noakes PG, Phillips WD, Hanley TA, Sanes JR, Merlie JP (1993) 43K protein and acetylcholine receptors colocalize during the initial stages of neuromuscular synapse formation in vivo. Dev Biol 155(1): 275–280 61. Okada K, Inoue A, Okada M, Murata Y, Kakuta S, Jigami T, Kubo S, Shiraishi H, Eguchi K, Motomura M, Akiyama T, Iwakura Y, Higuchi O, Yamanashi Y (2006) The muscle protein Dok-7 is essential for neuromuscular synaptogenesis. Science 312(5781): 1802–1805 62. Feng G, Laskowski MB, Feldheim DA, Wang H, Lewis R, Frisen J, Flanagan JG, Sanes JR (2000) Roles for ephrins in positionally selective synaptogenesis between motor neurons and muscle fibers. Neuron 25(2):295–306 63. Fox MA, Sanes JR, Borza DB, Eswarakumar VP, Fassler R, Hudson BG, John SW, Ninomiya Y, Pedchenko V, Pfaff SL, Rheault MN, Sado Y, Segal Y, Werle MJ, Umemori H (2007) Distinct target-derived signals organize formation, maturation, and maintenance of motor nerve terminals. Cell 129(1):179–193 64. Yumoto N, Kim N, Burden SJ (2012) Lrp4 is a retrograde signal for presynaptic differentiation at neuromuscular synapses. Nature 489(7416):438–442 65. Henriquez JP, Webb A, Bence M, Bildsoe H, Sahores M, Hughes SM, Salinas PC (2008) Wnt signaling promotes AChR aggregation at the neuromuscular synapse in collaboration with agrin. Proc Natl Acad Sci U S A 105(48):18812–18817 66. Luo ZG, Wang Q, Zhou JZ, Wang J, Luo Z, Liu M, He X, Wynshaw-Boris A, Xiong WC, Lu B, Mei L (2002) Regulation of AChR clustering by Dishevelled interacting with MuSK and PAK1. Neuron 35(3):489–505 67. Zhang J, Lefebvre JL, Zhao S, Granato M (2004) Zebrafish unplugged reveals a role for muscle-specific kinase homologs in axonal pathway choice. Nat Neurosci 7(12):1303–1309 68. Strochlic L, Falk J, Goillot E, Sigoillot S, Bourgeois F, Delers P, Rouviere J, Swain A, Castellani V, Schaeffer L, Legay C (2012) Wnt4 participates in the formation of vertebrate neuromuscular junction. PLoS One 7(1):e29976 69. Zhang B, Liang C, Bates R, Yin Y, Xiong WC, Mei L (2012) Wnt proteins regulate acetylcholine receptor clustering in muscle cells. Mol Brain 5:7 70. Gordon LR, Gribble KD, Syrett CM, Granato M (2012) Initiation of synapse formation by Wnt-induced MuSK endocytosis. Development 139(5):1023–1033 71. Henriquez JP, Krull CE, Osses N (2011) The Wnt and BMP families of signaling morphogens at the vertebrate neuromuscular junction. Int J Mol Sci 12(12):8924–8946 72. Koles K, Budnik V (2012) Wnt signaling in neuromuscular junction development. Cold Spring Harb Perspect Biol 4(6) 73. Krylova O, Herreros J, Cleverley KE, Ehler E, Henriquez JP, Hughes SM, Salinas PC (2002) WNT-3, expressed by motoneurons, regulates terminal arborization of neurotrophin-3-responsive spinal sensory neurons. Neuron 35(6):1043–1056 74. Wang J, Luo ZG (2008) The role of Wnt/beta-catenin signaling in postsynaptic differentiation. Commun Integr Biol 1(2):158–160 75. Wang J, Ruan NJ, Qian L, Lei WL, Chen F, Luo ZG (2008) Wnt/ beta-catenin signaling suppresses Rapsyn expression and inhibits acetylcholine receptor clustering at the neuromuscular junction. J Biol Chem 283(31):21668–21675 76. Li XM, Dong XP, Luo SW, Zhang B, Lee DH, Ting AK, Neiswender H, Kim CH, Carpenter-Hyland E, Gao TM, Xiong WC, Mei L (2008) Retrograde regulation of motoneuron differentiation by muscle beta-catenin. Nat Neurosci 11(3):262–268 77. Liu Y, Sugiura Y, Wu F, Mi W, Taketo MM, Cannon S, Carroll T, Lin W (2012) beta-Catenin stabilization in skeletal muscles, but not.
(14) Mol Neurobiol (2014) 49:574–589. 78.. 79.. 80.. 81. 82.. 83.. 84.. 85.. 86.. 87.. 88. 89. 90. 91. 92.. 93. 94. 95.. 96.. 97.. 98.. 99.. in motor neurons, leads to aberrant motor innervation of the muscle during neuromuscular development in mice. Dev Biol 366(2):255– 267 Nishimura T, Kato K, Yamaguchi T, Fukata Y, Ohno S, Kaibuchi K (2004) Role of the PAR-3-KIF3 complex in the establishment of neuronal polarity. Nat Cell Biol 6(4):328–334 Shi SH, Jan LY, Jan YN (2003) Hippocampal neuronal polarity specified by spatially localized mPar3/mPar6 and PI 3-kinase activity. Cell 112(1):63–75 Rolls MM, Doe CQ (2004) Baz, Par-6 and aPKC are not required for axon or dendrite specification in Drosophila. Nat Neurosci 7(12):1293–1295 Dickson BJ (2005) Wnts send axons up and down the spinal cord. Nat Neurosci 8(9):1130–1132 Liu Y, Shi J, Lu CC, Wang ZB, Lyuksyutova AI, Song XJ, Zou Y (2005) Ryk-mediated Wnt repulsion regulates posterior-directed growth of corticospinal tract. Nat Neurosci 8(9):1151–1159 Hutchins BI, Li L, Kalil K (2012) Wnt-induced calcium signaling mediates axon growth and guidance in the developing corpus callosum. Sci Signal 5(206):pt1 Hall AC, Lucas FR, Salinas PC (2000) Axonal remodeling and synaptic differentiation in the cerebellum is regulated by WNT-7a signaling. Cell 100(5):525–535 Ciani L, Salinas PC (2005) WNTs in the vertebrate nervous system: from patterning to neuronal connectivity. Nat Rev Neurosci 6(5): 351–362 Ahmad-Annuar A, Ciani L, Simeonidis I, Herreros J, Fredj NB, Rosso SB, Hall A, Brickley S, Salinas PC (2006) Signaling across the synapse: a role for Wnt and Dishevelled in presynaptic assembly and neurotransmitter release. J Cell Biol 174(1):127–139 Cerpa W, Godoy JA, Alfaro I, Farias GG, Metcalfe MJ, Fuentealba R, Bonansco C, Inestrosa NC (2008) Wnt-7a modulates the synaptic vesicle cycle and synaptic transmission in hippocampal neurons. J Biol Chem 283(9):5918–5927 Minino AM, Arias E, Kochanek KD, Murphy SL, Smith BL (2002) Deaths: final data for 2000. Natl Vital Stat Rep 50(15):1–119 Wynn TA (2008) Cellular and molecular mechanisms of fibrosis. J Pathol 214(2):199–210 Wynn TA (2011) Integrating mechanisms of pulmonary fibrosis. J Exp Med 208(7):1339–1350 Zhou L, Lu H (2010) Targeting fibrosis in Duchenne muscular dystrophy. J Neuropathol Exp Neurol 69(8):771–776 Liu L, Carron B, Yee HT, Yie TA, Hajjou M, Rom W (2009) Wnt pathway in pulmonary fibrosis in the bleomycin mouse model. J Environ Pathol Toxicol Oncol 28(2):99–108 Peng H, Cheng ML (2008) The relationship of metallothionein and liver fibrosis. Zhonghua Gan Zang Bing Za Zhi 16(1):78–79 Huang C, Ogawa R (2012) Fibroproliferative disorders and their mechanobiology. Connect Tissue Res 53(3):187–196 Serrano AL, Mann CJ, Vidal B, Ardite E, Perdiguero E, MunozCanoves P (2011) Cellular and molecular mechanisms regulating fibrosis in skeletal muscle repair and disease. Curr Top Dev Biol 96: 167–201 Lagrota-Candido J, Canella I, Pinheiro DF, Santos-Silva LP, Ferreira RS, Guimaraes-Joca FJ, Lannes-Vieira J, Quirico-Santos T (2010) Characteristic pattern of skeletal muscle remodelling in different mouse strains. Int J Exp Pathol 91(6):522–529 Soehnlein O, Lindbom L (2010) Phagocyte partnership during the onset and resolution of inflammation. Nat Rev Immunol 10(6):427– 439 Kharraz Y, Guerra J, Mann CJ, Serrano AL, Munoz-Canoves P (2013) Macrophage plasticity and the role of inflammation in skeletal muscle repair. Mediat Inflamm 2013:491497 Tidball JG, Villalta SA (2010) Regulatory interactions between muscle and the immune system during muscle regeneration. Am J Physiol Regul Integr Comp Physiol 298(5):R1173–R1187. 587 100. Lesault PF, Theret M, Magnan M, Cuvellier S, Niu Y, Gherardi RK, Tremblay JP, Hittinger L, Chazaud B (2012) Macrophages improve survival, proliferation and migration of engrafted myogenic precursor cells into MDX skeletal muscle. PLoS One 7(10): e46698 101. Chazaud B, Brigitte M, Yacoub-Youssef H, Arnold L, Gherardi R, Sonnet C, Lafuste P, Chretien F (2009) Dual and beneficial roles of macrophages during skeletal muscle regeneration. Exerc Sport Sci Rev 37(1):18–22 102. Lockhart NC, Brooks SV (2008) Neutrophil accumulation following passive stretches contributes to adaptations that reduce contraction-induced skeletal muscle injury in mice. J Appl Physiol 104(4):1109–1115 103. Moyer AL, Wagner KR (2011) Regeneration versus fibrosis in skeletal muscle. Curr Opin Rheumatol 23(6):568–573 104. Davalos D, Akassoglou K (2012) Fibrinogen as a key regulator of inflammation in disease. Semin Immunopathol 34(1):43–62 105. Arnold L, Henry A, Poron F, Baba-Amer Y, van Rooijen N, Plonquet A, Gherardi RK, Chazaud B (2007) Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. J Exp Med 204(5):1057–1069 106. Porter JD, Merriam AP, Leahy P, Gong B, Feuerman J, Cheng G, Khanna S (2004) Temporal gene expression profiling of dystrophindeficient (mdx) mouse diaphragm identifies conserved and muscle group-specific mechanisms in the pathogenesis of muscular dystrophy. Hum Mol Genet 13(3):257–269 107. Emery AE (2002) Muscular dystrophy into the new millennium. Neuromuscul Disord 12(4):343–349 108. O'Brien KF, Kunkel LM (2001) Dystrophin and muscular dystrophy: past, present, and future. Mol Genet Metab 74(1–2):75–88 109. Shin J, Tajrishi MM, Ogura Y, Kumar A (2013) Wasting mechanisms in muscular dystrophy. Int J Biochem Cell Biol 45(10):2266–2279. doi:10.1016/j.biocel.2013.05.001 110. Brandan E, Cabello-Verrugio C, Vial C (2008) Novel regulatory mechanisms for the proteoglycans decorin and biglycan during muscle formation and muscular dystrophy. Matrix biology : J Int Soc Matrix Biol 27(8):700–708 111. Karalaki M, Fili S, Philippou A, Koutsilieris M (2009) Muscle regeneration: cellular and molecular events. In Vivo 23 (5):779–796 112. Le Grand F, Rudnicki M (2007) Satellite and stem cells in muscle growth and repair. Development 134(22):3953–3957 113. Scime A, Rudnicki MA (2008) Molecular-targeted therapy for Duchenne muscular dystrophy: Progress and potential. Mol Diagn Ther 12(2):99–108 114. von Maltzahn J, Renaud JM, Parise G, Rudnicki MA (2012) Wnt7a treatment ameliorates muscular dystrophy. Proc Natl Acad Sci U S A 109(50):20614–20619 115. Morales MG, Cabello-Verrugio C, Santander C, Cabrera D, Goldschmeding R, Brandan E (2011) CTGF/CCN-2 overexpression can directly induce features of skeletal muscle dystrophy. J Pathol 225(4):490–501 116. Vial C, Gutierrez J, Santander C, Cabrera D, Brandan E (2011) Decorin interacts with connective tissue growth factor (CTGF)/ CCN2 by LRR12 inhibiting its biological activity. J Biol Chem 286(27):24242–24252 117. Cabello-Verrugio C, Santander C, Cofre C, Acuna MJ, Melo F, Brandan E (2012) The internal region leucine-rich repeat 6 of decorin interacts with low density lipoprotein receptor-related protein-1, modulates transforming growth factor (TGF)-beta-dependent signaling, and inhibits TGF-beta-dependent fibrotic response in skeletal muscles. J Biol Chem 287(9):6773–6787 118. Qi W, Chen X, Twigg S, Zhang Y, Gilbert RE, Kelly DJ, Pollock CA (2007) The differential regulation of Smad7 in kidney tubule cells by connective tissue growth factor and transforming growth factor-beta1. Nephrology (Carlton) 12(3):267–274.
(15) 588 119. Wahab NA, Weston BS, Mason RM (2005) Modulation of the TGFbeta/Smad signaling pathway in mesangial cells by CTGF/ CCN2. Exp Cell Res 307(2):305–314 120. Cabello-Verrugio C, Cordova G, Vial C, Zuniga LM, Brandan E (2011) Connective tissue growth factor induction by lysophosphatidic acid requires transactivation of transforming growth factor type beta receptors and the JNK pathway. Cell Signal 23(2):449–457 121. Vial C, Zuniga LM, Cabello-Verrugio C, Canon P, Fadic R, Brandan E (2008) Skeletal muscle cells express the profibrotic cytokine connective tissue growth factor (CTGF/CCN2), which induces their dedifferentiation. J Cell Physiol 215(2):410–421 122. Massague J (2012) TGFbeta signalling in context. Nat Rev Mol Cell Biol 13(10):616–630 123. Akhurst RJ, Hata A (2012) Targeting the TGFbeta signalling pathway in disease. Nat Rev Drug Discov 11(10):790–811 124. Massague J (1998) TGF-beta signal transduction. Annu Rev Biochem 67:753–791 125. Zhou L, Porter JD, Cheng G, Gong B, Hatala DA, Merriam AP, Zhou X, Rafael JA, Kaminski HJ (2006) Temporal and spatial mRNA expression patterns of TGF-beta1, 2, 3 and TbetaRI, II, III in skeletal muscles of mdx mice. Neuromuscul Disord 16(1):32–38 126. Massague J, Cheifetz S, Endo T, Nadal-Ginard B (1986) Type beta transforming growth factor is an inhibitor of myogenic differentiation. Proc Natl Acad Sci U S A 83(21):8206–8210 127. Like B, Massague J (1986) The antiproliferative effect of type beta transforming growth factor occurs at a level distal from receptors for growth-activating factors. J Biol Chem 261(29):13426–13429 128. Attisano L, Labbe E (2004) TGFbeta and Wnt pathway cross-talk. Cancer Metastasis Rev 23(1–2):53–61 129. Mezzano V, Cabrera D, Vial C, Brandan E (2007) Constitutively activated dystrophic muscle fibroblasts show a paradoxical response to TGF-beta and CTGF/CCN2. J Cell Commun Signal 1(3–4):205–217 130. Zhu J, Li Y, Shen W, Qiao C, Ambrosio F, Lavasani M, Nozaki M, Branca MF, Huard J (2007) Relationships between transforming growth factor-beta1, myostatin, and decorin: implications for skeletal muscle fibrosis. J Biol Chem 282(35):25852–25863 131. Cheon SS, Wei Q, Gurung A, Youn A, Bright T, Poon R, Whetstone H, Guha A, Alman BA (2006) Beta-catenin regulates wound size and mediates the effect of TGF-beta in cutaneous healing. FASEB J 20(6):692–701 132. Eger A, Stockinger A, Park J, Langkopf E, Mikula M, Gotzmann J, Mikulits W, Beug H, Foisner R (2004) beta-Catenin and TGFbeta signalling cooperate to maintain a mesenchymal phenotype after FosER-induced epithelial to mesenchymal transition. Oncogene 23(15):2672–2680 133. Dao DY, Yang X, Chen D, Zuscik M, O'Keefe RJ (2007) Axin1 and Axin2 are regulated by TGF- and mediate cross-talk between TGFand Wnt signaling pathways. Ann N Y Acad Sci 1116:82–99 134. Shi-Wen X, Leask A, Abraham D (2008) Regulation and function of connective tissue growth factor/CCN2 in tissue repair, scarring and fibrosis. Cytokine Growth Factor Rev 19(2):133–144 135. Brigstock DR (2002) Regulation of angiogenesis and endothelial cell function by connective tissue growth factor (CTGF) and cysteine-rich 61 (CYR61). Angiogenesis 5(3):153–165 136. Bork P (1993) The modular architecture of a new family of growth regulators related to connective tissue growth factor. FEBS Lett 327(2):125–130 137. Chen CC, Lau LF (2009) Functions and mechanisms of action of CCN matricellular proteins. International J Biochem Cell biol 41(4):771–783 138. Yoshida K, Munakata H (2007) Connective tissue growth factor binds to fibronectin through the type I repeat modules and enhances the affinity of fibronectin to fibrin. Biochimica et biophysica acta 1770(4):672–680 139. Wahab NA, Weston BS, Mason RM (2005) Connective tissue growth factor CCN2 interacts with and activates the tyrosine kinase receptor TrkA. J Am Soc Nephrol : JASN 16(2):340–351. Mol Neurobiol (2014) 49:574–589 140. Minamizato T, Sakamoto K, Liu T, Kokubo H, Katsube K, Perbal B, Nakamura S, Yamaguchi A (2007) CCN3/NOV inhibits BMP-2induced osteoblast differentiation by interacting with BMP and Notch signaling pathways. Biochem Biophys Res Commun 354(2):567–573 141. Passerini L, Bernasconi P, Baggi F, Confalonieri P, Cozzi F, Cornelio F, Mantegazza R (2002) Fibrogenic cytokines and extent of fibrosis in muscle of dogs with X-linked golden retriever muscular dystrophy. Neuromuscul Disord 12(9):828–835 142. Mori T, Kawara S, Shinozaki M, Hayashi N, Kakinuma T, Igarashi A, Takigawa M, Nakanishi T, Takehara K (1999) Role and interaction of connective tissue growth factor with transforming growth factor-beta in persistent fibrosis: A mouse fibrosis model. J Cell Physiol 181(1):153–159 143. Frazier K, Williams S, Kothapalli D, Klapper H, Grotendorst GR (1996) Stimulation of fibroblast cell growth, matrix production, and granulation tissue formation by connective tissue growth factor. J Invest Dermatol 107(3):404–411 144. Surmann-Schmitt C, Sasaki T, Hattori T, Eitzinger N, Schett G, von der Mark K, Stock M (2012) The Wnt antagonist Wif-1 interacts with CTGF and inhibits CTGF activity. J Cell Physiol 227(5):2207– 2216 145. Rooney B, O'Donovan H, Gaffney A, Browne M, Faherty N, Curran SP, Sadlier D, Godson C, Brazil DP, Crean J (2011) CTGF/CCN2 activates canonical Wnt signalling in mesangial cells through LRP6: Implications for the pathogenesis of diabetic nephropathy. FEBS Lett 585(3):531–538 146. Mercurio S, Latinkic B, Itasaki N, Krumlauf R, Smith JC (2004) Connective-tissue growth factor modulates WNT signalling and interacts with the WNT receptor complex. Development 131(9): 2137–2147 147. Chen S, McLean S, Carter DE, Leask A (2007) The gene expression profile induced by Wnt 3a in NIH 3T3 fibroblasts. J Cell Commun Signal 1(3–4):175–183 148. Bentzinger CF, von Maltzahn J, Rudnicki MA (2010) Extrinsic regulation of satellite cell specification. Stem Cell Res Ther 1(3):27 149. Wynn TA, Ramalingam TR (2012) Mechanisms of fibrosis: therapeutic translation for fibrotic disease. Nat Med 18(7):1028–1040 150. Sampson N, Berger P, Zenzmaier C (2012) Therapeutic targeting of redox signaling in myofibroblast differentiation and age-related fibrotic disease. Oxidative Med Cell Longev 2012:458276 151. Hu B, Phan SH (2013) Myofibroblasts. Curr Opin Rheumatol 25(1): 71–77 152. Phan SH (2008) Biology of fibroblasts and myofibroblasts. Proc Am Thorac Soc 5(3):334–337 153. Si W, Kang Q, Luu HH, Park JK, Luo Q, Song WX, Jiang W, Luo X, Li X, Yin H, Montag AG, Haydon RC, He TC (2006) CCN1/ Cyr61 is regulated by the canonical Wnt signal and plays an important role in Wnt3A-induced osteoblast differentiation of mesenchymal stem cells. Mol Cell Biol 26(8):2955–2964 154. Henderson WR Jr, Chi EY, Ye X, Nguyen C, Tien YT, Zhou B, Borok Z, Knight DA, Kahn M (2010) Inhibition of Wnt/betacatenin/CREB binding protein (CBP) signaling reverses pulmonary fibrosis. Proc Natl Acad Sci U S A 107(32):14309–14314 155. Masiakowski P, Yancopoulos GD (1998) The Wnt receptor CRD domain is also found in MuSK and related orphan receptor tyrosine kinases. Curr Biol 8(12):R407 156. Cerpa W, Gambrill A, Inestrosa NC, Barria A (2011) Regulation of NMDA-receptor synaptic transmission by Wnt signaling. J Neurosci 31(26):9466–9471 157. Farias GG, Alfaro IE, Cerpa W, Grabowski CP, Godoy JA, Bonansco C, Inestrosa NC (2009) Wnt-5a/JNK signaling promotes the clustering of PSD-95 in hippocampal neurons. J Biol Chem 284(23):15857–15866 158. Ren S, Johnson BG, Kida Y, Ip C, Davidson KC, Lin SL, Kobayashi A, Lang RA, Hadjantonakis AK, Moon RT, Duffield JS (2013).
(16) Mol Neurobiol (2014) 49:574–589 LRP-6 is a coreceptor for multiple fibrogenic signaling pathways in pericytes and myofibroblasts that are inhibited by DKK-1. Proc Natl Acad Sci U S A 110(4):1440–1445 159. Cheng JH, She H, Han YP, Wang J, Xiong S, Asahina K, Tsukamoto H (2008) Wnt antagonism inhibits hepatic stellate cell activation and liver fibrosis. Am J Physiol Gastrointest Liver Physiol 294(1):G39– G49 160. Gauger KJ, Chenausky KL, Murray ME, Schneider SS (2011) SFRP1 reduction results in an increased sensitivity to TGF-beta signaling. BMC cancer 11:59 161. He W, Zhang L, Ni A, Zhang Z, Mirotsou M, Mao L, Pratt RE, Dzau VJ (2010) Exogenously administered secreted frizzled related. 589 protein 2 (Sfrp2) reduces fibrosis and improves cardiac function in a rat model of myocardial infarction. Proc Natl Acad Sci U S A 107(49):21110–21115 162. Beyer C, Reichert H, Akan H, Mallano T, Schramm A, Dees C, Palumbo-Zerr K, Lin NY, Distler A, Gelse K, Varga J, Distler O, Schett G, Distler JH (2013) Blockade of canonical Wnt signalling ameliorates experimental dermal fibrosis. Ann Rheum Dis 72(7): 1255–1258 163. Distler A, Deloch L, Huang J, Dees C, Lin NY, Palumbo-Zerr K, Beyer C, Weidemann A, Distler O, Schett G, Distler JH (2013) Inactivation of tankyrases reduces experimental fibrosis by inhibiting canonical Wnt signalling. Ann Rheum Dis 72:1575–1580.
(17)
Figure

Documento similar
The expansionary monetary policy measures have had a negative impact on net interest margins both via the reduction in interest rates and –less powerfully- the flattening of the
Jointly estimate this entry game with several outcome equations (fees/rates, credit limits) for bank accounts, credit cards and lines of credit. Use simulation methods to
In our sample, 2890 deals were issued by less reputable underwriters (i.e. a weighted syndication underwriting reputation share below the share of the 7 th largest underwriter
1. S., III, 52, 1-3: Examinadas estas cosas por nosotros, sería apropiado a los lugares antes citados tratar lo contado en la historia sobre las Amazonas que había antiguamente
Whereas both the use of the chemical TGF-β type I receptors inhibitor EW-7197 and the silencing of key components of the canonical TGF-β pathway (SMAD2 and SMAD3) provoke a
Since such powers frequently exist outside the institutional framework, and/or exercise their influence through channels exempt (or simply out of reach) from any political
In the previous sections we have shown how astronomical alignments and solar hierophanies – with a common interest in the solstices − were substantiated in the
In this review we will briefly summarize: (i) the sites of production, mechanism of action and signaling pathways that are activated by ROS and (ii) the role of the mitochondrial H
