i
UNIVERSIDAD TÉCNICA DE AMBATO
FACULTAD DE CIENCIA E INGENIERÍA EN ALIMENTOSCARRERA DE INGENIERÍA BIOQUÍMICA
Evaluación de la capacidad citotóxica y proapoptótica del látex de Brosimum utile (Sande) sobre células HeLa como posible alternativa terapéutica para el
tratamiento del cáncer de cuello uterino.
Trabajo de Titulación, modalidad Proyecto de Investigación, previa la obtención del Título de Ingeniero Bioquímico, otorgado por la Universidad Técnica de Ambato, a través de la Facultad de Ciencia e Ingeniería en Alimentos.
Autor: Acosta Ganán Michael Andrés Tutor: Carrero Castillo Yenddy Nayghit, PhD. Co-tutor: López Hernández Orestes Darío, PhD.
Ambato - Ecuador Agosto 2018 P O R T A D A P O R T A D A P O R T A D A P
ii
APROBACIÓN DEL TUTOR Y CO-TUTOR
Carrero Castillo Yenddy Nayghit, PhD. López Hernández Orestes Darío, PhD.
CERTIFICAN:
Que el presente trabajo de titulación ha sido minuciosamente revisado. Por lo tanto, autorizamos la presentación de este Trabajo de Titulación modalidad Proyecto de Investigación, el mismo que responde a las normas establecidas en el reglamento de Títulos y Grados de la Facultad.
Ambato, 25 de julio de 2018
Yenddy Carrero Castillo, PhD. Orestes López Hernández, PhD.
C.I.175740457-7 C.I. 175478486-4
iii
DECLARACIÓN DE AUNTETICIDAD
Yo, Acosta Ganán Michael Andrés, manifiesto que los resultados obtenidos en el presente Proyecto de Investigación, previo a la obtención del título de Ingeniero Bioquímico son absolutamente originales, auténticos y personales, a excepción de las citas.
Sr. Michael Andrés Acosta Ganán C.I. 125003608-2
iv
APROBACIÓN DE LOS MIEMBROS DE TRIBUNAL DE GRADO
Los suscritos profesores calificadores, aprueban el presente trabajo de Titulación modalidad Proyecto de Investigación, el mismo que ha sido elaborado de conformidad con las disposiciones emitidas por la Facultad de Ciencia e Ingeniería en Alimentos de la Universidad Técnica de Ambato.
Para constancia firman:
Presidente del Tribunal
Mg. Yunys Pérez Betancourt C.I. 175647174-0
PhD. Alcides Bustillos Ortiz C.I. 180400106-1
v
DERECHOS DE AUTOR
Autorizo a la Universidad Técnica de Ambato, para que haga de este Proyecto de Investigación o parte de él, un documento disponible para su lectura, consulta y procesos de investigación, según las normas de la institución.
Cedo los derechos en línea patrimoniales de mi Proyecto, con fines de difusión pública, además apruebo la reproducción de este Proyecto dentro de las regulaciones de la Universidad, siempre y cuando esta reproducción no suponga ganancia económica y se realice respetando mis derechos de autor.
Sr. Michael Andrés Acosta Ganán C.I. 125003608-2
vi
DEDICATORIA
A mis padres, por haberme brindado su apoyo incondicional en las decisiones que he tomado en mi vida, las cuales me han permitido aprender y crecer como persona; por enseñarme a distinguir lo bueno y lo malo y que pese a las adversidades que se presenten en la vida nunca hay que darse por vencido.
A mis hermanos de sangre y corazón, en especial al más pequeño del hogar, a mi “Marianito”, por estar siempre a mi lado, ya sea, riendo, jugando, discutiendo, pero lo más importante, por ser mi fuente de motivación para alcanzar mis metas y seguir siempre hacia adelante.
Al resto de mi gran familia, a mis tías y a mis tíos, por habernos brindado su apoyo en los momentos más difíciles, por enseñarme que la solidaridad es una de las cualidades más hermosas que puede tener un ser humano.
De corazón, muchas gracias a todos.
vii
AGRADECIMIENTOS
Primeramente, agradezco a mi Alma mater, la Universidad Técnica de Ambato, Facultad de Ciencia e Ingeniería en Alimentos, por abrirme las puertas de su seno científico para así poder estudiar mi carrera, así como también a los diferentes docentes que impartieron sus conocimientos, experiencias y su apoyo durante los diversos ciclos académicos. Mi más profundo y sincero agradecimiento a la Dra. Yenddy Carrero PhD. por haberme brindado la oportunidad de formar parte de su grupo de trabajo, por su gran amistad y confianza depositada, agradezco sus consejos, su compresión y su paciencia durante todo el desarrollo de mi tesis.
También quiero agradecer al Dr. Orestes López PhD. por su comprensión y ayuda fundamental en la presentación del presente proyecto.
A mi gran amiga y compañera tesista, Chenny, por ser un gran apoyo en todo el sentido de la palabra, así como también a Dianita y Belita, por su gran amistad y confianza; de corazón muchas gracias.
Para finalizar, mi agradecimiento al viejo Manja, por sus enseñanzas imprescindibles, su paciencia y su gran amistad. Asimismo, a los demás amigos del laboratorio, gracias por convertirlo en una estancia con un ambiente de trabajo enriquecedor y divertido.
viii
ÍNDICE GENERAL DE CONTENIDOS
PORTADA ... i
APROBACIÓN DEL TUTOR Y CO-TUTOR...ii
DECLARACIÓN DE AUNTETICIDAD ... iii
APROBACIÓN DE LOS MIEMBROS DE TRIBUNAL DE GRADO ... iv
DERECHOS DE AUTOR... v
DEDICATORIA ... vi
AGRADECIMIENTOS ...vii
ÍNDICE GENERAL DE CONTENIDOS ... viii
ÍNDICE DE FIGURAS ... x RESUMEN ... xi ABSTRACT ...xii INTRODUCCIÓN ...xii CAPÍTULO I EL PROBLEMA ... 2 1.1 TEMA ... 2 1.2 JUSTIFICACIÓN ... 2 1.3 OBJETIVOS ... 5 1.3.1 Objetivo General ... 5 1.3.2 Objetivos Específicos ... 5 CAPÍTULO II MARCO TEÓRICO ... 6 2.1 ANTECEDENTES INVESTIGATIVOS ... 6 2.1.1 Cáncer ... 6
2.1.2 Cáncer de cuello uterino... 7
2.1.3 Apoptosis y el cáncer ... 8
ix
2.1.5 Agentes quimioterapéuticos y la medicina tradicional ... 12
2.1.6 Látex de Brosimum utile ... 15
2.2 HIPÓTESIS ... 17
2.2.1 Actividad citotóxica in vitro del látex de Sande ... 17
2.2.2 Actividad pro apoptótica in vitro del látex de Sande ... 17
2.3 SEÑALAMIENTO DE LAS VARIABLES DE LA HIPÓTESIS ... 17
2.3.1 Variable Dependiente ... 17
2.3.2 Variable Independiente ... 17
CAPÍTULO III MATERIALES Y MÉTODOS ... 18
3.1 MATERIALES Y MÉTODOS... 18
3.1.1 Obtención del látex de Sande ... 18
3.1.2 Cuantificación de proteínas presentes en la muestra de látex ... 18
3.1.3 Cultivo Celular ... 19
3.1.4 Ensayo de citotoxicidad celular ... 19
3.1.5 Ensayos inmunocitoquímicos... 19
3.1.6 Ensayo de TUNEL ... 20
3.2 DISEÑO EXPERIMENTAL ... 21
CAPÍTULO IV RESULTADOS Y DISCUSIÓN... 22
4.1 ANÁLISIS Y DISCUSIÓN DE LOS RESULTADOS ... 22
4.1.1 Evaluación in vitro de la capacidad citotóxica del látex ... 22
4.1.2 Evaluación in vitro de la capacidad proapoptótica del látex ... 24
4.2 VERIFICACIÓN DE HIPÓTESIS... 35
4.2.1 Actividad citotóxica in vitro del látex de Sande ... 35
4.2.2 Actividad proapoptótica in vitro del látex de Sande ... 35
CAPÍTULO V CONCLUSIONES Y RECOMENDACIONES ... 36
5.1 CONCLUSIONES ... 36
x
REFERENCIAS BIBLIOGRÁFICAS ... 38 ANEXOS... 43
ÍNDICE DE FIGURAS
Figura 1. Vías de activación de la apoptosis. ... 10 Figura 2. Agentes quimioterapéuticos empleados para el tratamiento de diversos tipos de cáncer. ... 14 Figura 3. Actividad citotóxica de látex de Sande en células HeLa tratadas por 24 h.. .... 22 Figura 4. Efectos del látex de Sande en la expresión de la proteína proapoptótica BAX en células HeLa tratadas por 24 h.. ... 24 Figura 5. Expresión de la proteína BAX en células HeLa tratadas por 24 h.. ... 25 Figura 6. Efectos del látex de Sande en la expresión de la proteína antiapoptótica BCL-2 en células HeLa tratadas por 24 h. ... 26 Figura 7. Expresión de la proteína BCL-2 en células HeLa tratadas por 24 h.. ... 26 Figura 8. Razón entre las intensidades de expresión de las proteínas apoptóticas BCL-2 y BAX en células HeLa tratadas por 24 h. ... 27 Figura 9. Efectos del látex de Sande en la expresión de Caspasas 3/7 en células HeLa tratadas por 24 h. ... 29 Figura 10. Expresión de Caspasas 3/7 en células HeLa tratadas por 24 h. ... 30 Figura 11. Porcentaje de células apoptóticas (positivas) tras un periodo de tratamiento con látex de Sande de 24 h. ... 32 Figura 12. Detección de apoptosis por APO-BrdU™ TUNEL en células Hela. ... 32 Figura 13. Correlación de parámetros estudiados de acuerdo al tratamiento y control. .. 34
xi
RESUMEN
El cáncer es una de las principales causas de muerte a nivel mundial, generando una carga para la sociedad tanto en los países más desarrollados como lo menos desarrollados económicamente. En la actualidad existen diversos métodos terapéuticos para combatir el cáncer, no obstante, en su mayoría resultan agresivos e incapaces de combatir en su totalidad este padecimiento. En los últimos años se ha puesto hincapié en la investigación de productos naturales a fin de identificar y desarrollar nuevas estrategias terapéuticas efectivas contra el cáncer. En este contexto, la presente investigación se fundamentó en evaluar el potencial citotóxico y proapoptótico del látex procedente del árbol Brosimum
utile sobre la línea celular de cáncer cervical HeLa. Se realizaron análisis de viabilidad
celular (MTT) y ensayos inmunocitoquímicos por microscopia de fluorescencia de proteínas involucradas en la vía de apoptosis. El látex mostró una potencial actividad citotóxica a altas concentraciones de proteínas, que está relacionada con la capacidad de promover muerte celular programada al inducir la expresión de BAX y caspasas 3/7 y silenciar la expresión de BCL-2. Los resultados obtenidos indican que el látex de
Brosimum utile proporciona una fuente de principios activos prometedores para el
desarrollo de nuevos agentes quimioterapéuticos contra el cáncer de cuello uterino y posiblemente para varios tipos de neoplasias.
Palabras clave: Cáncer cervical, células HeLa, látex, apoptosis, actividad citotóxica,
xii
ABSTRACT
Cancer is one of the leading causes of death worldwide, generating a burden on society in both the most developed and economically less developed countries. At present, there are several therapeutic methods to fight cancer; however, most are aggressive and unable to combat this condition in its entirety. In recent years, emphasis has been placed on the research of natural products in order to identify and develop new effective therapeutic strategies against cancer. In this context, the present investigation was based on evaluating the cytotoxic and proapoptotic potential of the latex from the Brosimum utile tree on the HeLa cervical cancer cell line. Cell viability analysis (MTT) and immunocytochemical assays were performed by fluorescence microscopy of proteins involved in the apoptosis pathway. The latex showed a potential cytotoxic activity at high concentrations of proteins, which is related to the ability to promote programmed cell death by inducing the expression of BAX and caspases 3/7 and silencing the expression of BCL-2. The results obtained indicate that Brosimum utile latex provides a source of promising active ingredients for the development of new chemotherapeutic agents against cervical cancer and possibly for several types of neoplasms.
Keywords: Cervical cancer, HeLa cells, latex, apoptosis, cytotoxic activity, proapoptotic
1
INTRODUCCIÓN
Los últimos reportes estadísticos de incidencia y mortalidad del cáncer a nivel mundial son abrumadores. Según la base de datos de GLOBOCAN, se produjeron cerca de 14.1 millones de nuevos casos de cáncer y 8.2 millones de muertes por este padecimiento en el año 2012. En cuanto al cáncer de cuello uterino, se registraron cerca de 527600 nuevos casos y 265700 muertes (Torre et al., 2015).
El cáncer es un término empleado para describir un amplio grupo de enfermedades que se caracterizan por un desequilibro en la regulación del crecimiento y/o muerte celular con tendencia a la proliferación descontrolada y autónoma de las células, las cuales tienen la capacidad de invadir localmente y a distancia otros tejidos (Acuña et al., 2014; De la Garza & Juárez, 2014). En este sentido la muerte celular programada ha sido blanco de investigación para el desarrollo de agentes quimioterapéuticos destinados a la activación de esta vía de muerte en células tumorales, no obstante, se ha registrado la capacidad de estas células para inhibir las señales de activación de la apoptosis por diversos mecanismos. Adicionalmente y por desgracia, la efectividad de estos agentes se ve obstaculizada por su pobre farmacocinética y su distribución inespecífica en el organismo que a menudo generan toxicidad y efectos secundarios (Fiandalo & Kyprianou, 2012; Shan-Shan, Jia-Hui, Hao-Han, & Shu-Qin, 2017).
En la actualidad, existe un gran interés tanto científico como comercial, por parte de la industria farmacéutica, en continuar el descubrimiento de nuevos agentes con capacidad anticancerígena a partir de productos naturales con la estrategia de que los protocolos quimioterapéuticos eliminen las células malignas sin causar efectos tóxicos en el paciente (Elufioye, Abdul, & Moody, 2017). Motivo por el cual, diversas investigaciones se han enfocado en la medicina tradicional, la cual les ha resultado beneficiosa a lo largo de la historia, permitiéndoles descubrir nuevas moléculas anticancerígenas, en su mayoría de origen vegetal, para el tratamiento de diversos tipos de neoplasias.
2
CAPÍTULO I
EL PROBLEMA
1.1 TEMA
Evaluación de la capacidad citotóxica y proapoptótica del látex de Brosimum utile (Sande) sobre células HeLa como posible alternativa terapéutica para el tratamiento del cáncer de cuello uterino.
1.2 JUSTIFICACIÓN
El cáncer de cuello uterino se constituye como el segundo tipo de cáncer más comúnmente diagnosticado a nivel mundial y la tercera causa de muerte por cáncer en las mujeres en los países menos desarrollados. A nivel mundial, las tasas de incidencia son más altas en África subsahariana, Melanesia, América Latina y el Caribe, mientras que las más bajas se registran en Asia occidental, Australia/Nueva Zelanda y América del Norte (Torre et al., 2015).
En Ecuador, el cáncer de cérvix es la segunda causa de muerte en la población femenina después del cáncer de mama. Según las últimas estadísticas realizadas en el país por el Instituto Nacional de Estadística y Censos (INEC), cada año se diagnostican alrededor de 1600 casos nuevos de cáncer de cuello uterino, con un registro de 650 pacientes fallecidas en el año 2014 a causa de esta patología (Solca Quito, 2017).
Actualmente existe una amplia gama de tratamientos empleados para el cáncer de cuello uterino, entre los más comunes se encuentran la cirugía, radioterapia, quimioterapia y terapia molecular o dirigida, e incluso combinaciones de las mismas. Sin embargo, estos tratamientos no son capaces de controlar en su totalidad este padecimiento, tomando en cuenta que se han documentado una gran cantidad de efectos adversos en las pacientes
3
como consecuencia de estos procedimientos (American Cancer Society, 2016c). Así, diversas investigaciones se han enfocado en la búsqueda de nuevas estrategias terapéuticas en pro y beneficio de la humanidad y de su entorno, dando cabida a la investigación de opciones alternativas como la medicina natural y ancestral, consideradas para muchos, prácticas médicas sin validez científica por no responder a los cánones de la medicina convencional pero que merecen ser valoradas y estudiadas críticamente, ya que podrían tener un impacto sustantivo sobre la salud pública (Rojas, Silva, Sansó, & Alonso, 2013). El látex procedente de Brosimun utile, conocido coloquialmente como “látex de Sande”, es utilizado en la medicina ancestral por parte de las comunidades indígenas en donde se encuentra distribuida la especie. En el norte de Ecuador y en el sur de Colombia se reporta el uso del látex de Sande en el tratamiento de la gastritis, e inclusive ha sido utilizado por una comunidad indígena del Putumayo-Colombia (comunidad Camëntsa) en el tratamiento de diversos tipos de cáncer, de los cuales se han obtenido resultados aparentemente satisfactorios que mejoran la calidad de vida del paciente (Chindoy, 2013). El Ecuador, al ser un país multiétnico y pluricultural que alberga en sus territorios un sinnúmero de especies vegetales con fines medicinales, las cuales sientan las bases de una serie de conocimientos terapéuticos para varias enfermedades por parte de las comunidades nativas. Estos conocimientos médicos empíricos son alternativas a pacientes a quienes la medicina convencional no las ha podido ayudar y por el contrario ha incidido en su calidad de vida, así mismo para aquellos que no gozan de los recursos económicos para su tratamiento, pues en contraste la medicina tradicional es cómodamente asequible. En varias comunidades indígenas del país, en su mayoría pertenecientes a la región amazónica, se ha utilizado el látex del árbol Brosimum utile, conocido coloquialmente como Sande, para el tratamiento de varias enfermedades, especialmente la gastritis. Sin embargo, se ha reportado que en el país contiguo Colombia, una comunidad indígena del Putumayo (comunidad Camëntsa) utiliza el látex de Sande en el tratamiento de diversos tipos de cáncer, de los cuales se han obtenido resultados aparentemente satisfactorios que
4
mejoran la calidad de vida de los pacientes al disminuir los efectos secundarios de la quimioterapia a la cual han sido sometidos por medicina convencional (Chindoy, 2013). Este conocimiento empírico sobre el uso del látex de Sande hasta el momento no ha sido contundentemente validado desde el punto de vista científico, ya que no cuenta con la suficiente información que lo corrobore. Aunque se recalca la investigación realizada por Chindoy en el año 2013, en la cual aisló una fracción acuosa proteica del látex de este árbol de la cual reporta actividad citotóxica sobre la línea celular K562 (leucemia mieloide crónica). Sin embargo, la investigación no proporciona mucha información acerca de las proteínas de látex y los mecanismos por los cuales suprimen el crecimiento celular de la línea cancerígena. Por otro lado, Rivera-Parada (2012) demuestra que el látex de Sande presenta en su composición diversos compuestos químicos aromáticos que le confieren propiedades antioxidantes al látex, los cuales impiden el daño causado por radicales libres que han sido asociados con la presencia de cáncer.
Debido a la evidencia científica existente de las propiedades del látex de Sande, el estudio de la capacidad citotóxica y proapoptótica in vitro aportará mayor información que contribuirá en la validación del conocimiento tradicional en beneficio de la comunidad. De forma complementaria el estudio sentará las bases para la investigación de moléculas potenciales presentes en el látex como posibles agentes quimioterapéuticos para el tratamiento de varios tipos de neoplasias, especialmente el cáncer de cuello uterino teniendo en cuenta su alta incidencia en el país.
5
1.3 OBJETIVOS
1.3.1 Objetivo General
Evaluar la capacidad citotóxica y proapoptótica del látex de Sande (Brosimum utile) en la línea celular de cáncer cervical HeLa.
1.3.2 Objetivos Específicos
1. Determinar la actividad citotóxica del látex in vitro.
2. Evaluar la capacidad proapoptótica in vitro del látex mediante el análisis de la expresión de BAX, BCL-2, Caspasas 3/7 y apoptosis tardía.
3. Establecer la correlación de los parámetros estudiados con el efecto in vitro del látex.
6
CAPÍTULO II
MARCO TEÓRICO
2.1 ANTECEDENTES INVESTIGATIVOS
2.1.1 Cáncer
El cáncer es una condición patológica compleja provocada por un grupo de células transformadas que evolucionan progresivamente hasta alcanzar un estado neoplásico (mutaciones del ADN hereditarias que se transmiten a las nuevas generaciones de las células). A medida que el proceso avanza las células normales adquieren una sucesión de capacidades distintivas que en conjunto les brindan características propias de una célula cancerígena, las cuales durante el proceso de progresión tumoral les confieren la capacidad de formar tumores, que pueden progresar a la malignidad. Se han identificado ocho capacidades biológicas que las células cancerígenas necesitan para dar inicio y continuidad al proceso neoplásico, entre las que se destacan el mantenimiento de la señalización proliferativa, la resistencia a la muerte celular, la inducción de angiogénesis y la activación de la invasión y metástasis (Hanahan & Weinberg, 2011; Sirica, 1989; Thakur & Mishra, 2016).
El conjunto o aglomeración de células que han adquirido algunas capacidades distintivas por mutaciones somáticas en su material genético se denominan neoplasias o tumores, que se definen como masas anormales de tejido, cuyo crecimiento además de ser relativamente autónomo excede al de los tejidos normales, caracterizándose por tener rasgos funcionales y morfológicos diferentes a los de sus precursoras, con persistencia de proliferación incluso tras el cese del estímulo desencadenante de la transformación (Sirica, 1989; Soimout, 2008).
7
De acuerdo al comportamiento clínico y la caracterización histopatológica que posean las neoplasias se distinguen en benignas y malignas. Las primeras se caracterizan por ser de crecimiento lento e incapaces de generar metástasis, además se encuentra en una zona bien localizada y su tratamiento consiste en una extirpación quirúrgica. Por el contrario, los tumores malignos son de crecimiento rápido, se infiltran en estructuras adyacentes y se propagan a otros tejidos u órganos por vía sanguínea y linfática y se denominan según su tipo de origen, el órgano, la extensión de la enfermedad alcanzada y las características propias de cada tumor (Acuña et al., 2014; De la Garza & Juárez, 2014; Soimout, 2008). El cáncer es una enfermedad que data desde tiempos inmemoriales y es considerada en la actualidad una de las principales causas de muerte a nivel mundial, tanto en los países desarrollados como en los menos desarrollados económicamente. Según la Agencia Internacional para la investigación del Cáncer (IARC) se estima que en el año 2012 se produjeron 14.1 millones de nuevos casos de cáncer y 8.2 millones de muertes por este padecimiento. Sin lugar a duda, el cáncer es un problema de salud pública global y a pesar de los esfuerzos realizados hasta el momento, continúa afectando y causando la muerte de un gran número de personas sin distinción alguna (Salaverry, 2013; Torre et al., 2015).
2.1.2 Cáncer de cuello uterino
Entre los tipos de cáncer más comúnmente diagnosticados en mujeres se destaca el cáncer cervicouterino, una enfermedad neoplásica maligna que se origina en el epitelio del cérvix. Inicialmente, este proceso preinvasivo se limita al epitelio cervical y se conoce de forma variable como lesión intraepitelial escamosa (LIE). Aproximadamente el 90% de los cánceres de cuello uterino son carcinomas de células escamosas y se desarrollan a partir de células del exocérvix cercanas a la zona de transformación, y el 10% restante son adenocarcinomas que se originan en las células glandulares productoras de moco del endocérvix, cerca del cuerpo del útero (American Cancer Society, 2016a; Duarte-Franco & Franco, 2004).
8
Según los últimos reportes estadísticos de GLOBOCAN, realizados por la IARC, el cáncer cervicouterino se ha constituido como el segundo tipo de cáncer más comúnmente diagnosticado y la tercera causa de muerte por cáncer en las mujeres en los países en vía de desarrollo, con un estimado mundial de 527600 nuevos casos de cáncer de cuello uterino y 265700 muertes por este padecimiento en el año 2012 (Torre et al, 2015).
2.1.3 Apoptosis y el cáncer
En las últimas décadas, se han descrito diversos mecanismos involucrados en el proceso de carcinogénesis, así como el estudio de la respuesta inmunitaria del individuo y los mecanismos efectores que contribuyen con la erradicación de células tumorales y su progresión. En este sentido, la apoptosis, un metabolismo autodestructivo de acuerdo con la señal de muerte celular codificada genéticamente, ha jugado un papel fundamental en la respuesta intrínseca contra el cáncer, así como en su tratamiento, participando durante la regulación del crecimiento celular y el desarrollo de un tejido; de tal forma que la supresión de esta vía de muerte celular conduce a la iniciación, crecimiento y progresión de las neoplasias; motivo por el cual el estudio y comprensión de los mecanismos apoptóticos pueden contribuir al desarrollo de estrategias terapéuticas efectivas contra el cáncer (Elufioye, Abdul, & Moody, 2017; Fiandalo & Kyprianou, 2012).
Cuando una célula normal sufre daños extremos e irreparables a nivel del ADN u otros componentes, ya sea por diversas causas, experimenta una muerte celular programada (apoptosis) caracterizada por una serie de cambios morfológicos, los cuales incluyen: fragmentación nuclear, condensación de la cromatina, formación de pliegues en la membrana plasmática, desensamble del citoesqueleto y contracción celular; culminando el proceso con la formación de pequeños sacos que albergan todo el material celular denominados cuerpos apoptóticos, que posteriormente son eliminados por fagocitosis, sin generar un proceso inflamatorio (Elufioye et al., 2017; Muñoz & Cuca, 2016).
9
Existen diversas vías que desencadenan la ejecución de la apoptosis, pero cada una es activada por diferentes factores desencadenantes como señales mitocondriales (vía intrínseca), ligandos de muerte (vía extrínseca) y desprendimiento celular (anoikis), siendo este último, un mecanismo diseñado específicamente para protegerse contra la metástasis celular (Fiandalo & Kyprianou, 2012).
La vía intrínseca se inicia por estímulos internos (daño en el ADN genómico, estrés oxidativo, etc.) a través de señales mitocondriales que desencadenan en la activación de proteínas pertenecientes a la familia de las caspasas (caspasas 2, 4, 6, 8, 9, 10 y 12). Esta vía se encuentra regulada fuertemente por varios tipos de moléculas que se pueden diferenciar en dos clases principales: las proteínas antiapoptóticas o pro supervivencia como las IAP (inhibidores de proteínas de la apoptosis) y algunas proteínas de la familia BCL-2 (BCL-2, BCL-xL, BCL-W, MCL-1, BFL-1, BCL-B y A1); y las proteínas proapoptóticas que incluyen miembros de la misma familia BCL-2 (BAX, BAK, BID, BOK, SMAC y DIABLO), las cuales se activan a través de eventos de señalización que conducen a la permeabilización de la membrana mitocondrial externa, desencadenando la liberación del citocromo C, mismo que se une con APAF-1 y caspasa 9 para formar un complejo proteico denominado apoptosoma. Tras la formación de apoptosomas, la caspasa 9 se vuelve catalíticamente activa y actúa sobre dianas posteriores incluyendo las caspasas 3, 6 y 7 (caspasas efectoras), conduciendo finalmente a la degradación irreversible del ADN y la proteólisis de la célula.
Por otra parte, la vía extrínseca o también conocida como la vía del receptor de la muerte hace referencia a la inducción apoptótica por señales extracelulares de estrés que implican la activación de receptores de muerte a través de la unión con sus respectivos ligandos, tales como TNF-α, FASL y TRAIL. Tanto FASL como TRAIL inducen estrictamente la muerte apoptótica, ambos ligandos son muy similares y convergen en la liberación de la forma activa de caspasa 8 que desencadena la activación de las caspasas efectoras. Por otro lado, TNF-α posee la capacidad de inducir apoptosis al igual que FASL y TRAIL, pero también es capaz de activar la vía prosupervivencia mediante la liberación del complejo proteico NF-κB, el cual una vez activado se transloca al núcleo y promueve la
10
expresión de genes involucrados en la supervivencia celular, por ejemplo, miembros antiapoptóticos de la familia BCL-2, inhibidor de caspasa 8 (c-FLIP), IAP, entre otros. (Caballero, 2016; Fiandalo & Kyprianou, 2012; Ramírez-García, Márquez-González, Barranco-Lampón, & López-Aguilar, 2014).
Figura 1. Vías de activación de la apoptosis. La vía extrínseca (izquierda) activada por la unión del ligando
de muerte FASL con su receptor específico FAS, localizado en la membrana celular. Se induce la activación de caspasa 8 y consecuente la activación de la caspasa 3. La vía intrínseca o vía mitocondrial (derecha) activada por estrés celular conduce a una baja regulación de los miembros de la familia BCL-2 antiapoptóticos, lo que permite a los miembros proapoptóticos (BAX, BAK) permeabilizar la membrana mitocondrial y provocar la liberación del citocromo c (Bellance, Lestienne, & Rossignol, 2009).
11
Además, durante la apoptosis se producen alteraciones en la membrana plasmática como la translocación del lípido fosfatidilserina hacia la cara externa de la bicapa lipídica (superficie celular), evento que facilita el reconocimiento de las células apoptóticas por parte de los fagocitos (macrófagos, células dendríticas y epiteliales) para de esta manera ser eliminadas eficientemente por fagocitosis, evitando así la inflamación en dicha zona (Porras & Marzo, 2010; Taylor, Cullen, & Martin, 2008).
2.1.4 Breve historia de los tratamientos contra el cáncer
Los mecanismos de la apoptosis han sido blanco de numerosas investigaciones, las cuales han dado como resultado tratamientos efectivos para combatir el cáncer, los cuales son consecuencia directa de una larga data de ensayos y aplicaciones de otros tratamientos menos exitosos, tomando en cuenta que un mismo tratamiento no es efectivo para todos los individuos debido a la respuesta inmunitaria, estadios y agresividad del tumor. Los primeros tratamientos ortodoxos contra el cáncer datan a mediados del siglo XIX, tiempo en el que se realizan los primeros ensayos para la resección completa de los tumores, describiéndose desde ese momento las técnicas estándar de la cirugía oncológica. Actualmente, la cirugía es uno de los tres tratamientos, junto con la radioterapia y la quimioterapia, más aceptados para combatir el cáncer y se puede utilizar para el diagnóstico del mismo, determinar su localidad, saber si ha diseminado o si afecta las funciones de otros órganos, extirpar el tumor de manera parcial o total, entre otras (De la Garza & Juárez, 2014; Granados, Beltrán, Soto, & León, 2011).
La radioterapia data a inicios del siglo XX en el tratamiento de enfermedades como el lupus y los epiteliomas, orientándose más tarde al tratamiento de los tumores malignos. Su principio consiste en la aplicación controlada de radiaciones ionizantes (rayos X) sobre el tejido afectado a fin de evitar la proliferación de las células transformadas; sin embargo, las células normales adyacentes también pueden resultar afectadas, aunque tienden a recuperarse y continuar con sus funciones normales (Acuña et al., 2014; American Cancer Society, 2017; Graña, 2015).
12
La primera opción como tratamiento para varios tipos de cáncer es la quimioterapia, cuyo inicio data a mediados del siglo XX con el empleo del gas mostaza en el tratamiento de linfomas (De la Garza & Juárez, 2014; Graña, 2015). El tratamiento consiste en la administración de agentes quimioterapéuticos que suprimen la proliferación de las células cancerígenas de la neoplasia a través de la inducción de las vías de muerte celular. No obstante, la quimioterapia surte su efecto a través de todo el organismo, lo cual difiere de la cirugía y la radioterapia que actúan en una localización especifica (Elufioye et al., 2017; Shan-Shan, Jia-Hui, Hao-Han, & Shu-Qin, 2017). Además, existen varios tratamientos modernos para combatir el cáncer, en su mayoría tratamientos derivados de la quimioterapia, tales como, la terapia hormonal, la inmunoterapia y más recientemente la terapia molecular o terapia dirigida (De la Garza & Juárez, 2014).
2.1.5 Agentes quimioterapéuticos y la medicina tradicional
En la actualidad, se conoce un gran número de agentes quimioterapéuticos o fármacos destinados al tratamiento contra el cáncer, ya sea por si solos o en combinación con otros medicamentos, según su mecanismo de acción se clasifican en agentes citostáticos y citotóxicos. Los primeros evitan la división celular o el crecimiento de la neoplasia a diferencia de los segundos que inciden en la muerte celular. Los agentes quimioterapéuticos tradicionales que tienen la capacidad de activar el sistema de señalización de apoptosis e interferir en la mitosis celular siguen siendo los gold estándar de los fármacos citotóxicos contra el cáncer en la aplicación clínica. Se incluyen los agentes alquilantes (bulsufán, altretamina, ciclofosfamida, temozolomida. etc.), antimicrotubulares (vincristina, vinblastina, paclitaxel, etc.), antimetabolitos (floxiridina, hidroxiurea, metotrexato, etc.), inhibidores de topoisomerasa (topotecán, etopósido, tenipósido, etc.), antibióticos citotóxicos (daunorubicina, doxorrubicina, epirubicina, etc.), entre otros (American Cancer Society, 2016b; De la Garza & Juárez, 2014; Shan-Shan et al., 2017).
13
Sin embargo, a pesar de las ventajas que ofrece la quimioterapia, la eficacia de la mayoría de los agentes quimioterapéuticos se ve obstaculizada por su pobre farmacocinética y su distribución inespecífica en el organismo, además de que su efectividad a menudo está limitada por su toxicidad y efectos secundarios en los tejidos y órganos normales. La toxicidad y los efectos secundarios más habituales incluyen: disfunción hematopoyética de la médula ósea, dolores de cabeza, pérdida del cabello, fatiga, náuseas y vómitos; efectos que generalmente están relacionados con el mecanismo de los agentes quimioterapéuticos (Sak, 2012; Shan-Shan et al., 2017).
Hoy en día, existe un considerable interés científico y comercial en continuar el descubrimiento de nuevos agentes anticancerígenos a partir de productos naturales (incluido plantas, animales, microorganismos y minerales) con la estrategia de que los protocolos quimioterapéuticos eliminen las células cancerosas sin causar efectos tóxicos en el huésped (Elufioye et al., 2017). En las últimas décadas se han reportado productos naturales con utilidad como agentes quimioterapéuticos directos, mientras que otros poseen la capacidad de disminuir la resistencia de las células cancerosas a los fármacos quimioterapéuticos, mejorar la eficacia de los mismos y reducir los efectos secundarios producidos por la quimioterapia (Elufioye et al., 2017; Sak, 2012).
En general, los metabolitos secundarios de las plantas han sido la fuente más vasta y exitosa de potenciales agentes quimioterapéuticos, e incluso la mayoría de las drogas modernas utilizadas en clínicas son producidas a partir de estos compuestos o de sus derivados, por ejemplo los ácidos procedentes del ácido cafeoilquínico (clorogénico, dicafeoilquínico y tricafeoilquínico), aislados de los tubérculos de Ipomoea batatas, poseen un efecto quimioprotector contra el cáncer, o el descubrimiento de uno de los agentes anticancerígenos más relevantes de la historia, el paclitaxel, aislado de la corteza del tejo (Taxus brevifolia), actúa como un antagonista en el desensamble de los microtúbulos durante la mitosis, inhibiendo así la replicación de las células (Dias, Urban, & Roessner, 2012; Pandey & Madhuri, 2009; Schaepfer & Mendoza-Espinoza, 2010). De manera complementaria, la medicina tradicional ha desempeñado un rol importante en el descubrimiento de la mayoría de los agentes quimioterapéuticos, pues gracias a los
14
conocimientos empíricos de las culturas ancestrales se han descubierto de plantas medicinales un gran número de estructuras químicas que sientan las bases para el desarrollo de nuevos fármacos; como es el caso de la planta Catharantas roseus, utilizada por varias culturas para el tratamiento de la diabetes, que permitió el aislamiento de los alcaloides de la vinca (vinblastina y vincristina), los cuales tienen la capacidad de prevenir la formación de microtúbulos y por tanto la inhibición de la mitosis, y que han sido ampliamente utilizados para combatir varios tipos de cáncer (Cragg, Kingston, & Newman, 2012; Laza, Rodríguez, & Sardiña, 2003; Schaepfer & Mendoza-Espinoza, 2010; Shan-Shan et al., 2017).
Figura 2. Agentes quimioterapéuticos empleados para el tratamiento de diversos tipos de cáncer.
Moléculas aisladas de fuentes de origen vegetal (vinblastina, vincristina y paclitaxel) y análogos sintéticos (topotecán y metotrexato).
15
Debido al rol que ha desempeñado la medicina tradicional en el descubrimiento de principios activos a lo largo de la historia, la Organización Mundial de la Salud (OMS, 2013) la ha definido como “la suma total de los conocimientos, capacidades y prácticas basados en las teorías, creencias y experiencias propias de diferentes culturas, bien sean explicables o no, utilizadas para mantener la salud y prevenir, diagnosticar, mejorar o tratar enfermedades físicas y mentales”. Pero, a pesar de los aportes de la medicina tradicional, sus conocimientos siguen siendo infravalorados por no responder a los cánones de la medicina convencional, sin embargo, dichas prácticas médicas deberían ser investigadas ampliamente debido a que podrían tener un impacto sustantivo sobre la salud pública (Rojas et al., 2013).
2.1.6 Látex de Brosimum utile
En la actualidad, y debido a la amplia gama de plantas medicinales en todas partes del mundo, existe un sinnúmero de especies que aún no han sido rigurosamente estudiadas, tal es el caso del árbol Brosimum utile, una especie perteneciente a la familia Moraceae, de aproximadamente 35 m de altura y 1.5 m de diámetro, de corteza delgada que al hacer incisiones sobre éste fluye un látex de apariencia blanquecina y pegajosa; crece en bosques húmedos tropicales en zonas de terrazas bajas o suelos habitualmente inundados y se distribuye geográficamente desde Costa Rica hasta Perú (Chindoy, 2013).
En Ecuador, el árbol se localiza en la región costa y amazónica, y generalmente es utilizado por la industria maderera y artesanal en la fabricación de muebles y contrachapados (Jorgensen & León-Yánez, 1999; Palacios, 2016). Aunque, por otro lado, se reporta el uso del látex con fines medicinales por parte de comunidades indígenas en el tratamiento de la gastritis e incluso en varios tipos de cáncer, como la leucemia, con resultados “satisfactorios” por su mejora en la calidad de vida de los pacientes al disminuir los efectos secundarios de la quimioterapia a la cual han sido sometidos por medicina convencional(Chindoy, 2013; Palacios, 2016).
16
Se estima que tales resultados pueden ser atribuibles a la variedad de compuestos químicos presentes en el látex, ya que según Rivera-Parada (2012) en su composición el látex de Sande presenta taninos, flavonoides, alcaloides, esteroles y tripertenoides, que le confieren actividad antioxidante por la presencia de anillos aromáticos en las moléculas. Por otro lado, se reporta una alta actividad citotóxica de la fracción clorofórmica obtenida del látex sobre el organismo modelo Artemia salina, evidenciándose así que el látex posee entre sus componentes de baja o nula polaridad un grupo de metabolitos con actividad citotóxica. Asimismo, Chindoy en el año 2013 reporta una alta actividad citotóxica de una fracción proteica acuosa obtenida del látex de Sande sobre la línea celular K562 (Leucemia mieloide crónica), también menciona una posible actividad citotóxica de compuestos de bajo peso molecular por debajo de los 7000 Daltons.
Adicionalmente, diversas investigaciones reportan actividad citotóxica y antitumoral por parte de proteínas laticíferas de varias especies, tales como, Calotropis procera y
Himatanthus drasticus (Mart) Plumel, pertenecientes a la familia Asclepiadaceae y
Apocynaceae respectivamente. C. procera mostró una actividad citotóxica in vitro considerable sobre las líneas celulares HL-60, HCT-8, SF295 y MDA-MB-435, e incluso inhibición de crecimiento in vivo del sarcoma 180. Por otra parte, H. drasticus mostró actividad in vivo contra los tumores sarcoma 180 y walker 256; evidenciándose de esta manera el alto potencial citotóxico y antitumoral de las proteínas laticíferas de las plantas medicinales (Mousinho et al., 2011; Oliveira et al., 2007, 2010).
Por lo anteriormente expuesto y tomando en consideración el potencial del látex del
Brosimum utile para el tratamiento del cáncer, se plantea analizar la actividad citotóxica
y proapoptótica in vitro del látex de Sande, a fin de aportar mayor información científica que contribuirá en la validación del conocimiento tradicional en beneficio de la comunidad y además sentar las bases para la investigación de estructuras químicas potenciales presentes en el látex encaminadas al desarrollo de agentes quimioterapéuticos para el tratamiento de varios tipos de neoplasias como el cáncer de cuello uterino.
17
2.2 HIPÓTESIS
2.2.1 Actividad citotóxica in vitro del látex de Sande
- Hipótesis Nula: El látex de Sande no presenta actividad citotóxica sobre células HeLa.
- Hipótesis alternativa: El látex de Sande presenta actividad citotóxica sobre células HeLa.
2.2.2 Actividad pro apoptótica in vitro del látex de Sande
- Hipótesis Nula: El látex de Sande no presenta actividad proapoptótica sobre células HeLa.
- Hipótesis alternativa: El látex de Sande presenta actividad proapoptótica sobre células HeLa.
2.3 SEÑALAMIENTO DE LAS VARIABLES DE LA HIPÓTESIS
2.3.1 Variable Dependiente
- Látex de Sande.
2.3.2 Variable Independiente
- Actividad citotóxica in vitro. - Actividad proapoptótica in vitro.
18
CAPÍTULO III
MATERIALES Y MÉTODOS
3.1 MATERIALES Y MÉTODOS
3.1.1 Obtención del látex de Sande
El látex se adquirió de la corteza de Brosimum utile, ubicado en la comunidad Pitirishca perteneciente a la provincia de Pastaza. Para el proceso de extracción del látex se realizaron incisiones en la corteza del tallo con un machete de acero inoxidable, mismo que se recolectó en un frasco de vidrio y se mezcló suavemente con buffer fosfato salino (PBS) a razón 1:1 (v/v), e inmediatamente se almacenó a 4 ºC.
3.1.2 Cuantificación de proteínas presentes en la muestra de látex
Para la cuantificación del contenido proteico del látex se empleó el ensayo colorímetro de Bradford y como patrón para la curva de calibración se utilizó albúmina de suero bovino (BSA) a 0.25, 0.50, 0.75, 1.00, 1.25, 1.50 y 1.75 mg/ml respectivamente. En una placa de 96 pocillos se colocaron 5 µl de la muestra y del patrón por triplicado. Seguidamente se añadieron 150 µl del reactivo Bradford (colorante) y se dejó reposar 5 min a temperatura ambiente. La lectura de las absorbancias se realizó a 595 nm en un espectrofotómetro de microplacas (PerkinElmer Víctor X3).
19
3.1.3 Cultivo Celular
Las células HeLa, cedidas por la Dra. Mileidys Pérez Alea del Grupo de Investigación de Modelos Animales y Cáncer de la Fundación Hospital Universitario Vall d'Hebron - Instituto de Investigación (Barcelona, España), se cultivaron en frascos T75 en medio DMEM suplementado con 10% suero fetal bovino (FBS) y 1% penicilina/estreptomicina y se incubaron a 37 ºC en una atmosfera al 5% CO2.
3.1.4 Ensayo de citotoxicidad celular
La actividad citotóxica del látex se determinó mediante el método colorimétrico MTT (Bromuro de 3-(4,5-dimetiltiazol-2-il)-2,5-difeniltetrazolio) de acuerdo al protocolo de la casa comercial (Thermo Fisher Scientific). Las células se sembraron en una placa de 96 pocillos a una densidad de 10000 células/pozo y se incubaron por 24 h. Posteriormente, se aplicó el tratamiento desde una concentración de 20 µg/ml y se diluyó 6 veces en escala logarítmica de base 10. Al cabo de 24 h, las células se lavaron con PBS 1X y consecutivamente se añadió el reactivo MTT a una concentración final de 0.5 mg/ml con un periodo de incubación de 4 h. Inmediatamente, se adicionaron 100 µl de SDS-HCl 0.01 M y se incubaron nuevamente las células por 4 h. Las absorbancias se determinaron a 570 nm en un espectrofotómetro de microplacas (PerkinElmer Víctor X3). El valor de la concentración inhibitoria media (IC50) se determinó utilizando el programa GraphPad
Prism V7.0.
3.1.5 Ensayos inmunocitoquímicos
Las células se sembraron en una placa de 6 pocillos sobre la superficie de un cubreobjeto estéril a una densidad de 300000 células/pozo y se incubaron por 48 h. A continuación, se adicionaron los respectivos tratamientos en base al valor del IC50. Después de 24 h las
células se lavaron con PBS 1X y se fijaron con paraformaldehído al 4% por 20 min y posteriormente se trataron con buffer de permeabilización (20 mM glicina 0.5 % tritón) por 15 min con lavados intermedios de PBS. Posteriormente se adicionaron 500 µl/pozo del anticuerpo a ensayar (BCL-2 y BAX en dilución 1:500 y Caspasas 3/7 a concentración
20
5 µM) y se incubaron a 37 ºC en agitación constante por 1 h. En el caso de los anticuerpos primarios no conjugados (BCL-2, BAX) se lavaron nuevamente con sus respectivos buffers de lavado y se añadieron 500 µl/pozo del anticuerpo secundario (fluoresceína o ficoeritrina en dilución 1:2000) y se incubaron a 37 ºC en agitación constante por 30 min. Una vez culminada la incubación se lavaron nuevamente las células y se retiraron los cubreobjetos de la placa, luego se dejaron secar a temperatura ambiente, protegidos de la luz. Al cabo de 24 h se añadió medio de montaje con DAPI (4',6-diamidino-2-fenilindol) y para las lecturas de las muestras se utilizó un microscopio invertido de fluorescencia (Leica DMi8).
3.1.6 Ensayo de TUNEL
Las células se sembraron en una placa de 6 pocillos sobre la superficie de un cubreobjeto estéril a una densidad de 1000000 células/pozo y se incubaron por 24 h de acuerdo al protocolo de la casa comercial (Thermo Fisher Scientific). A continuación, se adicionó el respectivo tratamiento en base al valor del IC50 y al cabo de 24 h las células se lavaron
con PBS y se fijaron en frío con paraformaldehído al 1% por 15 min. Posteriormente las células se trataron con etanol frío al 70% durante 18 h a -20 °C. Transcurrido este periodo, se lavaron nuevamente con buffer de lavado por 5 min y se añadieron 50 µl de solución de marcaje de ADN (DNA-labeling solution) por 1 h a 37 °C. Seguidamente, se colocó buffer de enjuague por 5 min y a continuación se añadieron 100 µl del anticuerpo (Anti-BrdU mouse monoclonal antibody PRB-1, Alexa Fluor 488 conjugate) en dilución 1:20 por 30 min a temperatura ambiente. Seguidamente se adicionaron 500 µl de buffer de tinción (Propidium iodide/RNase A staining buffer) por 30 min a temperatura ambiente. Una vez culminada la incubación se retiraron los cubreobjetos de la placa y se dejaron secar a temperatura ambiente, protegidos de la luz. Al cabo de 24 h se añadió medio de montaje sin DAPI y para las lecturas de las muestras se utilizó igualmente un microscopio invertido de fluorescencia.
21
3.2 DISEÑO EXPERIMENTAL
Todos los ensayos planteados en el presente trabajo de investigación se realizaron por triplicado. En el caso de las muestras analizadas en el microscopio de fluorescencia se cuantificaron las intensidades fluorescentes por campo 630X (10 campos aprox.), a través de los filtros DAPI (489-513 nm), GFP (475-509 nm) y Texa Red (595-615nm), de las proteínas involucradas en la apoptosis y el número de células positivas para el caso del ensayo de TUNEL. Los resultados obtenidos se procesaron y analizaron en el programa GraphPad Prism V7.0 y se presentan como promedio ±1 desviación estándar. Para los análisis comparativos se utilizó ANOVA de un solo factor más post test de Bonferroni y se consideraron significativos los valores de p ≤ 0.05.
22
CAPÍTULO IV
RESULTADOS Y DISCUSIÓN
4.1 ANÁLISIS Y DISCUSIÓN DE LOS RESULTADOS
4.1.1 Evaluación in vitro de la capacidad citotóxica del látex
El ensayo de citotoxicidad celular se trabajó con la muestra látex-PBS 1:1 en un rango de concentración de 20.38 µg/mL de proteína, diluido consecutivamente en una escala logarítmica de base 10 hasta el orden 10-6. En la figura 3 se presentan los resultados de la evaluación citotóxica del tratamiento con látex en células HeLa en un periodo de 24 h, cabe indicar que como control positivo se incluyó el agente quimioterapéutico sintético metotrexato (Mtx), un potente agente inductor de apoptosis y tratamiento de elección en muchos tipos de neoplasias.
-4 . 6 9 -3 . 6 9 -2 . 6 9 -1 . 6 9 -0 . 6 9 0 . 3 1 1 . 3 1 0 2 0 4 0 6 0 8 0 1 0 0 % V ia b il id a d c e lu la r L S M tx lo g1 0 [ µ g /m l] lo g2 [ m M ] - 4 .0 0 - 3 .0 0 - 2 .0 0 - 1 .0 0 0 1 . 0 0 2 . 0 0 F i g u r a 1 . A c t i v i d a d c i t o t ó x i c a d e l á t e x d e S a n d e ( L S ) y m e t o t r e x a t o ( M t x I C 5 0 ) e n c é l u l a s H e L a t r a t a d a s d u r a n t e 2 4 h . L o s v a l o r e s d e I C 5 0 f u e r o n c a l c u l a d o s p o r r e g r e s i ó n n o l i n e a l u t i l i z a n d o G r a p h P a d P r i s m V 7 . 0 I C5 0: 2 .4 7 µ g /m l I C5 0: 0 .4 9 m M
Figura 3. Actividad citotóxica de látex de Sande en células HeLa tratadas por 24 h. Se
utilizó Mtx como control positivo. Los valores de IC50 fueron calculados por regresión no lineal utilizando GraphPad Prism V7.0.
23
La actividad citotóxica del látex indica una supresión notoria en el crecimiento celular in
vitro a concentraciones proteicas altas de 20.380 y 2.038 µg/ml, a partir de esta última la
supresión en el crecimiento celular es intrascendente. Cabe recalcar que cuanto más bajo sea el valor de IC50 mayor será la actividad citotóxica de la muestra ensayada sobre la
línea celular y viceversa.
Al comparar el IC50 obtenido (2.47 µg/ml) con los resultados de Chindoy (2013) en la
línea celular K562 (leucemia mieloide crónica humana), IC50 1.49 µg/ml, se observa una
similitud en ambos casos a pesar de las respectivas diferencias como la línea celular y la duración del tratamiento que en el caso de Chindoy fue de 2 días. Por otro lado y aunque un poco lejano si se habla en términos de especie, los resultados obtenidos también reflejan similitud con la investigación realizada por Oliveira et al., (2007), en la cual una fracción proteica del látex de Calotropis procera presenta actividad citotóxica en las líneas celulares HL-60 (leucemia promielocítica humana), HCT-8 (carcinoma de colon humano), SF-295 (carcinoma de cerebro humano) y MDA-MD-435 (carcinoma mamario humano) con valores para IC50 de 0.86, 1.12, 0.42 y 1.36 µg/ml, respectivamente.
Es importante señalar que el IC50 obtenido fue calculado en base al contenido proteico de
la muestra látex-PBS, la cual no recibió ningún proceso específico de aislamiento y/o purificación de proteínas, por lo que, no se descarta de ninguna manera la posible actividad citotóxica de los demás compuestos químicos presentes en el látex, tales como, flavonoides, alcaloides y tripertenoides, de los cuales se ha demostrado actividad antioxidante y anticancerígena (Elufioye et al., 2017; Rivera-Parada, 2012).
Con la finalidad de evaluar si la capacidad citotóxica del látex contra las células HeLa está relacionada con la muerte celular programada se procedieron a realizar una sucesión de ensayos inmucitoquímicos de proteínas cruciales involucradas en los mecanismos apoptóticos (BAX, BCL-2, Caspasas 3/7) y además un análisis de apoptosis tardía (ensayo de TUNEL).
24
4.1.2 Evaluación in vitro de la capacidad proapoptótica del látex
La evaluación de la capacidad proapoptótica de la muestra de látex se realizó mediante ensayos inmucitoquímicos a dos concentraciones diferentes en base al intervalo de confianza del 95% del IC50 (0.51-11.82 µg/ml), las cuales fueron 2 y 5 µg/ml, codificados
como LS 2 y LS 5 respectivamente. De igual manera, se adicionó Mtx como control positivo de apoptosis celular.
4.1.2.1 Efecto del látex en la de expresión de las proteínas BAX y BCL-2
Al igual que el ensayo de citotoxicidad celular la duración del tratamiento con látex en todos los ensayos inmunocitoquímicos fueron de 24 h. Una vez finalizado el tratamiento se realizaron los respectivos procesos para las tinciones de los anticuerpos monoclonales para de esta manera visualizar y cuantificar las proteínas involucradas en la apoptosis celular por microscopia de fluorescencia.
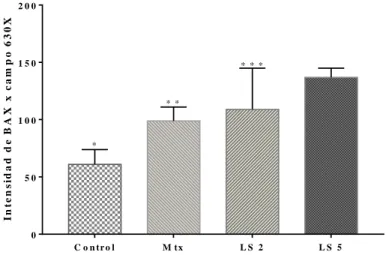
C o n tr o l M tx L S 2 L S 5 0 5 0 1 0 0 1 5 0 2 0 0 I n te n s id a d d e B A X x c a m p o 6 3 0 X * * * * * * F ig u r a X . E f e c to s d e l lá te x d e S a n d e e n la e x p r e s ió n d e la p r o te ín a p r o a p o p to tic a B a x e n c é lu la s H e L a tr a ta d a s d u r a n te 2 4 h . * p < 0 .0 0 1 v s . L S 2 y L S 5 . * * p < 0 .0 1 v s . c o n tr o l y L S 5 . * * * p < 0 .0 5 v s . L S 5 .
Figura 4. Efectos del látex de Sande en la expresión de la proteína
proapoptótica BAX en células HeLa tratadas por 24 h. *p<0.001 vs. LS 2
y LS 5. **p<0.01 vs. control y LS 5. ***p<0.05 vs. LS 5.
En la figura 4 se puede apreciar un evidente incremento en la expresión de la proteína proapoptótica BAX en ambos tratamientos con látex (p< 0.001) y de igual manera con Mtx (p<0.01) en relación al control. También se observó diferencia significativa entre los tratamientos LS 5 y Mtx (p<0.001).
25
Adicionalmente, en la figura 5 se puede observar la expresión de la proteína BAX (color verde) en células HeLa, la cual se localiza tanto en el citoplasma como en la membrana mitocondrial externa y de color azul los núcleos teñidos con DAPI.
Figura 5. Expresión de la proteína BAX en células HeLa tratadas por 24 h. (A)
Control (B) Mtx (C) LS 2 (D) LS 5. Magnificación 630X.
En contraste, en la figura 6 se observa un leve decrecimiento en la expresión de la proteína prosupervivencia BCL-2 en el tratamiento LS 2 (p<0.01) pero contundente en LS 5 y Mtx (p<0.001) con respecto al control. Al comparar el fármaco estándar Mtx con los tratamientos laticíferos no se observa diferencia significativa con LS 5 pero sí con LS 2 (p<0.001). Al igual que BAX, BCL-2 (color rojo) también se localiza en la membrana mitocondrial externa y en el retículo endoplasmático, así como en la membrana nuclear (figura 7).
26 C o n tr o l M tx L S 2 L S 5 0 5 0 1 0 0 1 5 0 I n te n s id a d d e B C L -2 x c a m p o 6 3 0 X * * * * * F ig u r a X . E f e c to s d e l lá t e x d e S a n d e e n la e x p r e s i ó n d e la p r o te ín a a n tia p o p tó ti c a B c l- 2 e n c é l u l a s H e L a t r a t a d a s d u r a n t e 2 4 h . * p < 0 .0 0 1 v s . L S 2 y L S 5 . * * p < 0 .0 0 1 v s . L S 2 . * * * p < 0 .0 1 v s . c o n tro l. *
Figura 6. Efectos del látex de Sande en la expresión de la proteína antiapoptótica
BCL-2 en células HeLa tratadas por 24 h. *p<0.001 vs. Mtx y LS 5. **p<0.001
vs. LS 2. ***p<0.01 vs. control.
Figura 7. Expresión de la proteína BCL-2 en células HeLa tratadas por 24 h. (A) Control (B) Mtx (C) LS 2 (D) LS 5. Magnificación 630X.
27
Según la literatura, ambas proteínas se encuentran estrechamente relacionadas, debido a que pertenecen a la misma familia de proteínas y comparten una extensa homología en sus secuencias aminoacídicas, y aunque desempeñan funciones relativamente opuestas sus roles durante la regulación de la apoptosis resulta ser sumamente importante, a tal manera que la sobreexpresión o el silenciamiento de éstas se encuentra íntimamente relacionada con el desarrollo y la agresividad de las neoplasias (Fernald & Kurokawa, 2013; Ramírez-García et al., 2014). Las dos proteínas interactúan a nivel mitocondrial, en la cual BCL-2 modula la inactivación de BAX mediante heterodimerización, lo que impide la permeabilización de la membrana mitocondrial externa. Según el modelo del reóstato cuando BCL-2 se encuentra en exceso forma homodímeros BCL-2:BCL-2 y heterodímeros BCL-2:BAX, evento que protege a la célula de la apoptosis. En contraste, cuando BAX está en exceso, dominan los homodímeros BAX:BAX y la célula es susceptible de muerte celular programada. En consecuencia, la proteína más abundante de las dos decidirá la supervivencia o la muerte de la célula (Kale, Osterlund, & Andrews, 2017; Korsmeyer, Shutter, Veis, Merry, & Oltvai, 1993).
C o n tr o l M tx L S 2 L S 5 0 . 0 0 . 5 1 . 0 1 . 5 2 . 0 2 . 5 R e la c ió n B C L -2 /B A X * * F ig u r a 4 . R a z ó n e n tr e la s in te n s id a d e s d e e x p r e s ió n d e la s p r o te ín a s B C L - 2 y B A X e n c é lu la s H e L a tr a ta d a s d u r a n te 2 4 h . * p < 0 .0 0 1 v s . M tx , L S 2 y L S 5 . * * p < 0 .0 1 v s . L S 2 . * * * p < 0 .0 0 1 v s . L S 5 . * * * *
Figura 8. Razón entre las intensidades de expresión de las proteínas apoptóticas
BCL-2 y BAX en células HeLa tratadas por 24 h. *p<0.001 vs. Mtx, LS 2 y LS
28
En la figura 8 se presentan los resultados de la relación entre las dos proteínas (BAX y BCL-2) involucradas en la muerte apoptótica, en la cual se aprecia un decrecimiento neto en la tasa BCL-2/BAX en los tres tratamientos ensayados (p<0.001) con respecto al control; e igualmente diferencia significativa entre LS 2 y LS 5 (p<0.001) y entre LS 2 y Mtx (p<0.01). Además, se puede observar que el control presenta una tasa de BCL-2/BAX muy por encima de 1, lo que indica la supremacía de la proteína prosupervivencia sobre la proteína proapoptótica, resultado que no es de extrañar pues diversas investigaciones han demostrado la sobreexpresión de BCL-2 en células cancerígenas e igualmente la supresión de la subfamilia de proteínas proapoptóticas como BAX (Ramírez-García et al., 2014). En LS 2, se observa una tasa ligeramente inferior a 1, valor que demuestra un equilibro parcial entre ambas proteínas. Por otro parte, LS 5 y Mtx presentan una tasa relativamente inferior a 1, que indica la supremacía de BAX sobre BCL-2 y según el modelo del reóstato la eminente muerte de la célula maligna por apoptosis.
29
4.1.2.2 Efecto del látex en la expresión de Caspasas 3/7
En la figura 9 se muestra los resultados de la expresión de las caspasas efectoras 3/7 tras la administración de los respectivos tratamientos (Mtx, LS 2 y LS 5), siendo LS 5 el más contundente en cuanto a expresión de caspasas se trata. Mtx y LS 2 de igual manera ejercen un incremento en la expresión de ambas proteínas, pero con menor intensidad. Asimismo, se observa diferencia significativa entre Mtx y LS 2 (p<0.01) y entre Mtx y LS 5 (p<0.001). Adicionalmente, en la figura 10 se puede apreciar la expresión de caspasas 3/7 de color verde, las cuales se localizan tanto en el citoplasma como en el núcleo celular. C o n tr o l M tx L S 2 L S 5 0 1 0 0 2 0 0 3 0 0 4 0 0 5 0 0 I n te n s id a d d e C a s p a s a s 3 /7 x c a m p o 6 3 0 X * * * * * * F ig u r a X . E f e c to s d e l lá te x d e S a n d e e n la e x p r e s ió n d e la s p r o te ín a s c a s p a s a s 3 /7 e n c é lu la s H e L a tr a ta d a s d u r a n te 2 4 h . * p < 0 .0 0 1 v s . c o n tr o l, L S 2 y L S 5 . * * p < 0 .0 1 v s . L S 2 . * * * p < 0 .0 0 1 v s M tx .
Figura 9. Efectos del látex de Sande en la expresión de Caspasas 3/7 en células
HeLa tratadas por 24 h. *p<0.001 vs. Control, LS 2 y LS 5. **p<0.01 vs. LS 2.
30
Figura 10. Expresión de Caspasas 3/7 en células HeLa tratadas por 24 h. (A, B) Control (C, D) Mtx (E, F) LS 2 (G, H) LS 5. Magnificación 630X.
31
Como se ha mencionado en anteriores apartados, la muerte celular programada posee diversas vías de activación, sin embargo, todas convergen en la liberación de la forma activa de las caspasas efectoras (caspasas 3, 6 y 7), quienes se encargan de ejecutar en sí la muerte de la célula, pues tras su activación se inicia rápidamente la degradación celular, hidrolizando diversos sustratos citoplasmáticos importantes para la viabilidad de la célula, como diversos inhibidores de apoptosis, proteínas del citoesqueleto, nucleares, del retículo endoplasmático y del aparato de Golgi, proteínas que intervienen en la regulación del ciclo celular, reparación, replicación y transcripción del ADN, entre muchas otras. Además, poseen como sustratos a las caspasas iniciadoras (caspasas 8, 9 y 10) lo que genera una retroalimentación en las señales de muerte (Caballero, 2016; Taylor et al., 2008). En consecuencia, el análisis de la expresión de las caspasas 3/7 además de indicar la expresión de las mismas también funcionaría como un método de identificación de células apoptóticas, pues tras su activación la ejecución de la apoptosis es irreversible. Como se puede observar en la figura 9, LS 5 presenta mayor intensidad en la expresión de las caspasas 3/7 en relación al control, lo que sugiere un incremento en la cantidad de células tumorales que sufrieron apoptosis producto de la acción del látex; le siguen los tratamientos LS 2 y Mtx respectivamente. No obstante, fue necesario analizar la cantidad de células apoptóticas por un método más específico.
4.1.2.3 Inducción de apoptosis por látex
El ensayo de TUNEL, al igual que los anteriores ensayos inmunocitoquímicos, se trabajó en un periodo de tratamiento de 24 h. Es importante señalar, que para este ensayo únicamente se trabajó con la muestra látex-PBS a concentración 5 µg/ml (LS 5), debido a que fue el tratamiento laticífero en el que se obtuvieron mejores efectos in vitro.
En la figura 11 se presentan los resultados del ensayo de TUNEL en porcentaje de células positivas o células apoptóticas tras los respectivos tratamientos con LS 5 y Mtx, en los cuales se observa diferencia significativa (p<0.001) en relación al control y de igual forma entre ambos tratamientos (p<0.001). Asimismo, se pueden apreciar los fragmentos de cadena de ADN de color verde intenso en el núcleo celular (figura 12).
32 C o n tr o l M tx L S 5 0 2 0 4 0 6 0 8 0 1 0 0 % C é lu la s p o s it iv a s x c a m p o 6 3 0 X * * *
Figura 11. Porcentaje de células apoptóticas (positivas) tras un periodo de
tratamiento con látex de Sande de 24 h. *p<0.001 vs. Mtx y LS 5. **p<0.001 vs.
LS 5.
Figura 12. Detección de apoptosis por APO-BrdU™ TUNEL en células Hela. (A)
Control, (B) Mtx, (C y D) LS 5. Las flechas indican la fragmentación del ADN detectado con un anticuerpo anti-BrdU marcado con Alexa Flúor® 488. Magnificación 630X.
33
Cabe mencionar que el ensayo de TUNEL es un método que se caracteriza por el análisis de la apoptosis en sus últimas etapas, enfocándose en los cambios de la morfología nuclear, específicamente en la degradación del ADN en una escalera de fragmentos, producto de la escisión de la cromatina mediada por endonucleasas, considerándose bajo estas circunstancias una célula positiva (Taylor et al., 2008). Como se puede observar en la figura 11, LS 5 incrementa la cantidad de células positivas al igual que Mtx en comparación al control, resultados que indican que el tratamiento con látex induce la muerte celular programada en células HeLa.
Además, los resultados del estudio demuestran una relación in vitro entre la expresión de las caspasas efectoras y la cantidad de células positivas en sus últimas etapas de la apoptosis, pues el incremento en la expresión de caspasas 3/7 reincide en un aumento de células apoptóticas, debido a que una vez activadas las caspasas efectoras la muerte de la célula es inminente. Igualmente se establece una correlación entre la expresión de las proteínas apoptóticas BCL-2/BAX y la expresión de las caspasas efectoras, en la cual, los tratamientos que presentaron supremacía de la proteína proapoptótica BAX sobre la proteína prosupervivencia BCL-2 inciden en un incremento en la expresión de las caspasas 3/7 (figura 13), de tal manera que dicha supremacía inclina la balanza hacia la muerte celular programada mediante la liberación de la forma activa de las caspasas efectoras (Caballero, 2016; Kale et al., 2017). En este sentido, los resultados obtenidos ponen en evidencia la potencial capacidad citotóxica y proapoptótica de los componentes del látex de Sande y prometen una base para el desarrollo de agentes quimioterapéuticos.
34 C o n tr o l M tx L S 2 L S 5 0 1 0 0 2 0 0 3 0 0 4 0 0 5 0 0 I n te n s id a d x c a m p o 6 3 0 X B A X B C L -2 C a s p a s a s 3 /7
Figura 13. Correlación de parámetros estudiados de acuerdo al tratamiento y control. Se observa incremento en la expresión de Caspasas 3/7 en los tratamientos que presentan supremacía de BAX sobre BCL-2.
Es importante rememorar que diversas investigaciones han demostrado que la proteína prosupervivencia BCL-2 desempeña un rol importante en la apoptosis y en el desarrollo de las neoplasias y por ende su influencia en los tratamientos contra el cáncer. Así, las grandes compañías farmacéuticas han dado prioridad al desarrollo de pequeñas moléculas moduladoras de la familia BCL-2; generando fármacos miméticos que inhiben a miembros de la familia antiapoptótica, tales como ABT-737, ABT-263, ABT-199, sin embargo, no son agentes selectivos e inhiben a miembros de la subfamilia proapoptótica, lo que disminuye su potencial en la destrucción de las células malignas, resultando poco efectivo, pues el objetivo primordial es disminuir tanto la resistencia frente a los agentes quimioterapéuticos como la supervivencia de la célula neoplásica (Kale et al., 2017; Ramírez-García et al., 2014).
Sin duda alguna los hallazgos científicos obtenidos en la presente investigación dirigen la atención al látex de B. utile como una fuente de moléculas con capacidad citotóxica, proapoptótica y por lo tanto anticancerígena en células HeLa, que podrían sentar las bases para el desarrollo de nuevos agentes quimioterapéuticos enfocados al tratamiento del cáncer de cuello uterino y posiblemente de varios tipos de neoplasias.