Unraveling the immune function of mineralocorticoid receptor (MR) and aldosterone in animal models
144
0
0
Texto completo
(2) PONTIFICIA UNIVERSIDAD CATÓLICA DE CHILE Facultad de Ciencias Biológicas Programa Doctorado en Ciencias Biológicas Mención Genética Molecular y Microbiología DOCTORAL THESIS: UNRAVELING THE IMMUNE FUNCTION OF MINERALOCORTICOID RECEPTOR (MR) AND ALDOSTERONE IN ANIMAL MODELS Tesis entregada a la Pontificia Universidad Católica de Chile en cumplimiento parcial de los requisitos para optar al Grado de Doctor en Ciencias con mención en Genética Molecular y Microbiología. Por NATALIA MUÑOZ DURANGO Tutor: Dr. Alexis M. Kalergis Parra Comité de tesis: Dra. Susan Bueno Dra. María Estela Andrés Dr. Alfredo de Ioannes Dr. Luis Michea Dr. Rodrigo de la Iglesia. SEPTEMBER, 2019. 2.
(3) PONTIFICIA UNIVERSIDAD CATÓLICA DE CHILE Facultad de Ciencias Biológicas Programa Doctorado en Ciencias Biológicas Mención Genética Molecular y Microbiología La defensa final de la tesis Doctoral titulada: UNRAVELING THE IMMUNE FUNCTION OF MINERALOCORTICOID RECEPTOR (MR) AND ALDOSTERONE IN ANIMAL MODELS Tesis entregada a la Pontificia Universidad Católica de Chile en cumplimiento parcial de los requisitos para optar al Grado de Doctor en Ciencias con mención en Genética Molecular y Microbiología. Presentada con fecha de hoy por la candidata a Doctor NATALIA MUÑOZ DURANGO Ha sido aprobada por el Tribunal Examinador, constituido por los profesores abajo firmantes, calificándose el trabajo realizado, el manuscrito sometido y la defensa oral con nota _______. Dr. Alexis Kalergis Parra Director de Tesis Facultad de Ciencias Biológicas PUC. Dra. Susan Bueno Miembro del comité de Tesis Facultad de Ciencias Biológicas PUC. Dra. María Estela Andrés Miembro del comité de Tesis Facultad de Ciencias Biológicas PUC. Dr. Alfredo de Ioannes Miembro del comité de Tesis Facultad de Ciencias Biológicas PUC. Dr. Luis F. Michea Acevedo Miembro del comité de Tesis Facultad de Medicina UdeCh. Dr. Rodrigo de la Iglesia Miembro del comité de Tesis Facultad de Ciencias Biológicas PUC. SEPTEMBER, 2019. 3.
(4) INDEX ACKNOWLEDGMENT ....................................................................................................... 7 ABBREVIATIONS .............................................................................................................. 9 GENERAL ABSTRACT .................................................................................................... 10 GENERAL INTRODUCTION ............................................................................................ 12 1. The immune response and its modulation. ............................................................... 12 2. Biology of aldosterone and Mineralocorticoid Receptor (MR).................................... 14 3. Models to study immunological role of aldosterone and MR ..................................... 17 3.1. Immune cells stimulation by aldosterone............................................................ 18 3.2. Pharmacologic MR antagonism in immune cells ................................................ 19 3.3. MR knockout animal models. ............................................................................. 20 GENERAL HYPOTHESIS ................................................................................................ 22 GENERAL AIMS .............................................................................................................. 22 GENERAL DISCUSSION ................................................................................................. 23 GENERAL PROPOSED MODEL...................................................................................... 25 GENERAL BIBLIOGRAPHY ............................................................................................. 26 Chapter I .......................................................................................................................... 33 The axis aldosterone - Mineralocorticoid Receptor (MR) is involved in the regulation of Tlr4 expression in dendritic cells impacting the in vivo response to LPS. ................................. 33 ABSTRACT .................................................................................................................. 33 INTRODUCTION .......................................................................................................... 34 METODOLOGY ............................................................................................................ 38. 4.
(5) RESULTS ..................................................................................................................... 44 1. Aldosterone-driven activation of MR leads to Tlr4 expression by DCs .................. 44 2. Wild type dendritic cells pre-treated with aldosterone displayed a higher inflammatory response to low doses of LPS................................................................................... 48 3. In vivo D-Aldosterone pretreatment in WT/LM mice increases the peripheral tlr4 expression and enhances the response to sublethal dose of LPS. ........................... 50 4. MyMRKO mice infused with D-Aldosterone displayed the same clinical response than MyMRKO non-infused in response to sublethal dose of LPS than mice. ........... 53 5. Dendritic cells derived from MyMRKO mice displayed different patterns of maturation in response to LPS than WT/LM DCs. ...................................................................... 54 DISCUSSION. .............................................................................................................. 59 SUPPLEMENTARY DATA ........................................................................................... 64 BIBLIOGRAPHY ........................................................................................................... 71 Chapter II ......................................................................................................................... 78 Absence of mineralocorticoid receptor in myeloid cells disturbs CD8+ T cells to prevent liver steatosis in a mouse model of nonalcoholic steatohepatitis .............................................. 78 ABSTRACT .................................................................................................................. 78 INTRODUCTION .......................................................................................................... 79 METHODOLOGY ......................................................................................................... 86 RESULTS ..................................................................................................................... 95 1. MyMRKO mice fed with MCD diet displayed lower liver steatosis than control LM/WT mice. ......................................................................................................................... 95 2. MyMRKO mice and LM/WT mice showed similar levels of fibrosis and inflammatory markers despite of their differences in lipid accumulation. ........................................ 98. 5.
(6) 3. Inflammatory cell infiltration of liver during NASH................................................ 103 4. Under inflammatory conditions, MyMRKO dendritic cells impaired the ability of CD8+ T lymphocytes to express activation markers.......................................................... 114 5. Immune response in spleens was similar to the observed in the livers................ 115 DISCUSSION. ............................................................................................................ 118 SUPPLEMENTARY DATA ......................................................................................... 127 BIBLIOGRAPHY ......................................................................................................... 134. 6.
(7) ACKNOWLEDGMENT This work was sponsored by Millennium Institute of Immunology and Immunotherapy and several FONDECYT and FONDEF projects. Also, I want thanks to Doctoral fellowship CONICYT 21140178 and VRI Excellent fellows.. I appreciate Dr. Alexis Kalergis´s the acceptance in his research group, and the confident that he had in my scientific ideas, some of them projects currently developing in his laboratory. To stayed in this place led me the opportunity to actively participate in a broad spectrum of collaborative works with Dra. Claudia Riedel and Dr. Felipe Simon of Universidad Andrés Bello; Dr. Leandro Carreño of Universidad de Chile, and Dra. Susan Bueno, Dr. Marco Arresse, Dr. Daniel Cabrera, Dr. Juan Carlos Roa, Dr. Cesar Sánchez, Dra. Alejandra Pérez, Dra. Andrea Vecchiola, Dr Carlos Fardella and Dra. Katia Abarca of Pontificia Universidad Católica de Chile. Also, I want to tanks to all secretaries and support personal that helped me always that I needed: Isabel Capitán, Claudia Castro, Elizabeth Ly, Maria Teresa Pino, Don José and Don Enrique. All of my thankfulness of members of current Dr. Kalergis´s Lab and Dra. Buenos´s Lab.. In a personal and professional field, I had the great opportunity to meet people that I keep in my hearth, not only for their friendship, also for their inspiring intelligence such Sebastián Riquelme, Pablo Céspedes, Janyra Espinoza, Catalina Pardo, Roberto Gómez, Maria Olga Bargsted and Juan Pablo Mackern. Also, I appreciate the shared moments with Bruno Ramírez, Hernán Correa, Rodrigo Díaz, Daniela Becerra, Claudia Rivera and Evelyn Jara. I had the honor to work with people like Javier Valdés, Emma Rey, Yaneisi Vásquez, Nicolás Gálvez and Jorge Soto in the Phase I Clinical trial of hRSV vaccine. With this group, we built a friendship based in the collaborative work and supporting each to other in the. 7.
(8) challenging labor that we did. I have to do a special mention of a group of friends who shared with me the weakness of eating French fries, do Yoga or just having a good time, such, Geraldyne Salazar, Omar Vallejos, Hernán Peñaloza, Janyra Espinoza, Franciaco Salazar and Valentina Sebastián.. Because I had the blessing to find a family in Chile, I wanted to thank to Doña Cecilia, Gordito, Checha, Cristián, Anita y Sofi for your love an always be there. A special mention to Ale who is my lovely friend or in better words is my Chilean sister, I love you Ale. Also, I had the luck to meet Juan Carlos Fernández and Derly Andrade, a couple of friends that stolen my heart forever.. The most deeply feeling of love to my Family (Aminta, Anibal, Kata, Alejo, Rafa, Mónica, Juancho, Martina y Elena), who are the blame of strongest and happiest feel that ever I would have. You are my north and inspiration, I so blessed to be a part of you. Finally, but in the most important part of my heart, I would like to thanks my husband Jacobo Martínez for be himself by my side. We are growing together, alone and far of home, but building our home together with Lucky and Felina. I appreciate not only your patience, love, companion and comprehension, also be my sponsor when I needed, you are the best partner of any person could have.. 8.
(9) ABBREVIATIONS DCs: Dendritic cells LM: Litter mate WT: Wild type TLR: Toll Like Receptors LPS: Lipopolysaccharide Th: T helper APC: Antigen Presenting Cells MR: Mineralocorticoid receptor GR: Glucocorticoid receptor RAAS: Renin-angiotensin-aldosterone system MyMRKO: Myeloid MR knockout PRR: Patter Receptor Recognition NFkB: Nuclear Factor k B MAPKs: Mitogen-activated protein kinases PAMPs: Pathogen associated molecular patterns DAMPs: Damage associated molecular patterns HRE: Hormonal response elements GRE: Glucorticoid receptor elements MRE: Mineralocorticoid receptor elements PBMCs: Peripheral blood mononuclear cells NASH: Non-alcoholic steatohepatitis MCD: Methionine-Choline deficient diet NALFD: Non-Alcoholic Liver Fatty Disease HSC: Hepatic stellated cells. 9.
(10) GENERAL ABSTRACT Inflammation is a physiological response that could be triggered by endogenous and exogenous stimulus. During inflammation the response is auto limited and regulated to keep homeostasis, because uncontrolled response could lead to death. Inflammatory response is complex and involves many systems of the body which crosstalk each with other. In this line, hormones and its cognate receptors have been described as modulator of immune response, for example glucocorticoids and its knowing immunosuppressive role have been very useful to treat cancer and autoimmune diseases. Aldosterone is another adrenal hormone, that under non-pathological condition controls renal excretion of water and electrolytes to keep blood pressure. This hormone conventionally acts through mineralocorticoid receptor (MR) which is a ligand dependent transcription factor mainly expressed in juxtaglomerular epithelial cells in kidneys. However, MR is also expressed in cells of the immune system, adipose tissue, liver and brain, indicating that could be paying a role. Up to date, it has been described that high levels of aldosterone in vivo and in vitro are related with the induction of inflammatory phenotype in cardiac diseases, atherosclerosis, obesity, insulin resistance and autoimmunity.. Aimed to describe the immunological role in which MR and aldosterone are involved, we used two models to study it. The first model was designed to understand whether MRaldosterone was related with TLR4 expression. According to our findings we described that aldosterone via MR induces the expression of tlr4 in wild type (WT) bone marrow-derived dendritic cells (DCs). Oppositely, the trl4 expression in DCs derived from myeloid MR conditional knockout mice (MyMRKO) did not show to be modulated by aldosterone, indeed these cells displayed impaired capacity to response to LPS stimulation. Finally, in a model of sub-lethal endotoxemia, we found that mice pre-treated with aldosterone succumbs after LPS challenge in contrast to placebo treated mice, mainly due to a multi-organic failure. In. 10.
(11) conclusion our results suggest that aldosterone-MR axis is involved in the regulation of tlr4 expression, and in consequently modulating the DCs response to TLR4 agonist, LPS.. Because the first model was focused in septic inflammation, the second model that we used was focused in aseptic chronic inflammation in which MR and renin-angiotensinaldosterone system (RAAS) played a role. We induced non-alcoholic steatohepatitis (NASH) in LM/WT and MyMRKO mice with methionine-choline deficient diet (MCD). We found that MyMRKO mice fed with MCD diet presented lower lipid accumulation in livers than controls. This finding was also related with lower number of CD8+ T lymphocytes infiltration in livers of MyMRKO. This cells also displayed lower expression CD25 activation receptor, indicating that antigen presenting cells (APC) derived from MyMRKO could be having impairments promoting the immune response. In vitro cocultures performed with CD4+ or CD8+ T lymphocytes plus DCs derived from MyMRKO mice displayed that only CD8+ response was impaired and is related with the phenotype seen in mice.. Summarizing the results, we described that aldosterone through MR are involved in tlr4 expression, because the loss of MR in myeloid DCs impacted in the levels of tlr4 expressed, indeed at basal condition. This result also affected the capacity of MyMRKO DCs to sense and responds to LPS, which is a maturation stimulus, but in these cells failed to induced a proinflammatory prone phenotype. These results are in accordance with the lowest capacity of DCs to drive immune response of CD8+ T lymphocytes impacting in lower steatosis in NASH model. In conclusion, all abovementioned results indicated that loss of MR in myeloid cells impacts in the adaptive CD8+ immune response. However, further analysis about how MR modulates antigen presenting cells to impair CD8+ T lymphocytes function is needed.. 11.
(12) GENERAL INTRODUCTION 1. The immune response and its modulation. Mammal immune system is as complex biological network directed to stablish a multistep and regulate response against environmental injuries (1). This system includes cells, soluble molecules and different body compartments, such, skin, mucosa, blood and lymph nodes (1,2). Inflammation is the classic manifestation of immune response and it is triggered by the recognition of potentially harmful endogenous or exogenous molecules (1,3). This recognition is mainly mediated by a family of receptors able to sense and induce inflammatory response, known as pathogen recognition receptors (PRRs) (4). According to PRR´s role, they are mainly expressed extra and intracellularly in antigen presenting cells (APC), such as, dendritic cells (DCs), monocytes/macrophages and B Lymphocytes (4). One of the most studied family of PRRs are Toll Like Receptors (TLRs) (5), which are type I transmembrane proteins with ectodomains containing interspersed leucine-rich repeats (LRR) motifs that are involved in the recognition of molecular patterns on ligands (6). The cytoplasmic domain is characterized by a Toll/IL-1 receptor (TIR) motif, that is a conserved module necessary for the interactions with adapter proteins to induce signaling transduction by mitogen-activated protein kinases (MAPKs) (6) leading to the activation of nuclear factorkappa B (NF-κB) and production of pro-inflammatory cytokines (7,8). Up to date, there are described at least 13 TLRs which recognizes different molecules derived from pathogens (known as pathogen associated molecular patterns -PAMPs-) (5,9) and from cellular components (known as damage associated molecular patterns -DAMPs-) (10). As result, TLRs have the ability to promotes adaptive immune responses in several ways, including antibody responses, T helper 1 (Th1) and Th17 CD4+ T cell responses, and CD8+ T cell responses (11).. 12.
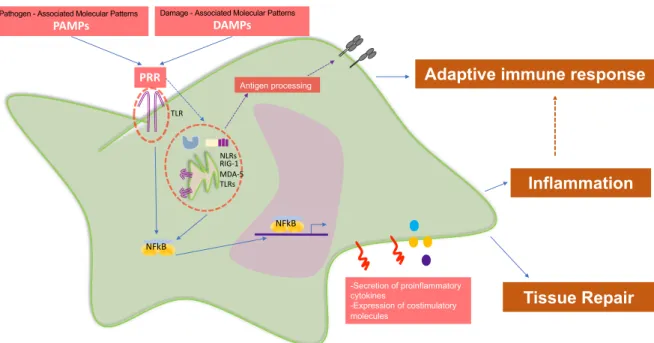
(13) Pathogen - Associated Molecular Patterns. Damage - Associated Molecular Patterns. DAMPs. PAMPs. PRR. Antigen processing. Adaptive immune response. TLR. NLRs RIG-1 MDA-5 TLRs. Inflammation NFkB. NFkB. -Secretion of proinflammatory cytokines -Expression of costimulatory molecules. Tissue Repair. Figure 1: Antigen presenting cells (APCs) express different Patter Receptor Recognition (PRR) to sense very diverse environmental injuries. Some PRRs are: Toll Like Receptors (TLRs) which are located intra an extracellularly; Nucleotide-binding oligomerization domain-like receptors (NLRs) which are located intracellularly; Retinoid Acid Inducible Gene (RIG-1) which are located intracellularly; etc. Each group of PRR sense different type of molecules to induce signaling pathways leading cytokine secretion, costimulatory molecules expression and antigen processing to develop adaptive immune response. APCs stimulation then can result in inflammation induction, tissue repair and adaptive immune response polarization (12).. In this line, it is important to highlight that DCs are the most potent APCs known up to date (13). These cells are characterized by their unique ability to recognize and respond against many stimuli to induce adaptive immune response, serving as a key component of innate immunity and bridge innate and adaptive immune response (13). For that reason, DCs and other immune cells not only express receptors that induce its activation, also express many other receptors to keep tolerance and regulation (14). After activation, DCs start to express negative costimulatory molecules such cytotoxic T-lymphocyte–associated. 13.
(14) antigen 4 (CTLA4) and programed cell death ligand 1 (PDL1) (15,16), suggesting that these inhibitory signals are important for dampening activation in the context of inflammatory signals (14).. Rather than immune specific receptors, DCs also constitutively express “nontraditionally immune receptors” which helps to control immune response, such as glucocorticoid receptor (GR). GR is a nuclear receptor that binds to steroidal glucocorticoid hormone that have a physiologic key role in glucose metabolism, however it is also known as the most potent immunosuppressor because its ability to impair gene expression of proinflammatory cytokines (17,18), CD4+T cells function by limiting IL-2 production and inducing lymphopenia via cell death (19). Mechanistically, it has been reported that cytokine promoters contain hormones response elements (HREs), which can be target by glucocorticoids and potentially by other corticoid hormone, such mineralocorticoids (20). It has been reported that simple binding of ligand-receptor to HRE is enough to induce or repress gene expression, due its nature of nuclear receptor (21). Another nuclear receptor expressed in DCs is mineralocorticoid receptor (MR), but oppositely than described to GR, MR have been closely related with pro-inflammatory condition (22).. 2. Biology of aldosterone and Mineralocorticoid Receptor (MR) Aldosterone is a mineralocorticoid hormone that plays a key role maintaining sodium levels and water homeostasis. The main regulator of aldosterone secretion is the reninangiotensin system (RAAS) (23,24). Angiotensin II induces the synthesis of aldosterone by the zona glomerulosa of the adrenal cortex, but also high potassium levels and the adrenocorticotropic hormone can regulate aldosterone synthesis (25). The result of aldosterone production is the increase of sodium reabsorption at the expense of potassium. 14.
(15) by activating the apical epithelial sodium channel and the basolateral Na+-K+ ATPase in the distal nephron (26). Therefore, this regulation axis leads to an increase of blood volume and bloodstream pressure.. In the abovementioned regulatory axis, aldosterone is known to signal through the mineralocorticoid receptor (MR) expressed in epithelial cells. However, recent studies have described MR-independent effects for aldosterone, also referred to as non-genomic effects (27). MR is a member of the steroid nuclear receptor family and a ligand-inducible transcription factor (28,29). The inactive form of MR is found in the cell cytoplasm forming heterocomplexes with the heat shock protein 90 (hsp90) chaperone (30). Upon ligand binding, MR undergoes conformational changes that lead to the dissociation of the hsp90, and to the formation of homodimers and heterodimers with GR-glucocorticoid complex, which can translocate into the nucleus acting as a transcription factor (31,32). At the nucleus, these homo/heterodimers bind to Glucocorticoid Response Elements (GRE) to repress or induce the expression of target genes mainly involved in water homeostasis, such as, epithelial sodium channel, Na+–K+–ATPase, serum/glucocorticoid regulated kinase 1 (SGK1) (33,34), CHIF and the epidermal growth factor receptor (EGFR) (35). Up to date, there has been described only one specific Mineralocorticoid Response Element (MRE) located in human EGFR promoter (35). However, to explain the specific responses mediated for MR-aldosterone joined to GREs, it has been proposed the recruitment of selective coactivators or corepressors, such as eleven-nineteen lysine-rich leukemia (ELL), Fasassociated factor 1 (FAF-1), nuclear receptor co-repressor (NCoR) l and the silencing mediator of retinoid and thyroid hormone receptor (SMRT) (36). Additionally, the translocation to the nucleus of MR can also be promoted by glucocorticoids, such cortisol, because these molecules bind with to MR with an affinity equivalent to aldosterone (37). But, in epithelial tissues the enzyme 11β hydroxysteroid dehydrogenase type 2 (11βHSD-2). 15.
(16) promotes the selective activation of MR by aldosterone, because this enzyme catalyzes the conversion of cortisol to its inactive form, cortisone (38). Although cortisone is still able to occupy MR, it is not able to induce MR activation (38).. Aldosterone effects on sodium transport in epithelial cells are usually divided into an “early” and a “late” phase (33). The early response to aldosterone takes a place about 0.5 to 3 hours after hormone addition, and is related with the enhancement activity of the preexisting sodium channels (33). For example, the early expression of sgk-1 kinase, which is a serine/threonine kinase involved in increasing epithelial sodium channel activity are recognize as one of this aldosterone early products (33). On the other hand, the rapid nongenomic effects mediated by aldosterone (measured in much shorter times), are thought to be mediated by the stimulation of cellular mechanisms, such as second-messenger systems and signaling pathways including the phosphorylation of MAPKs.. 16.
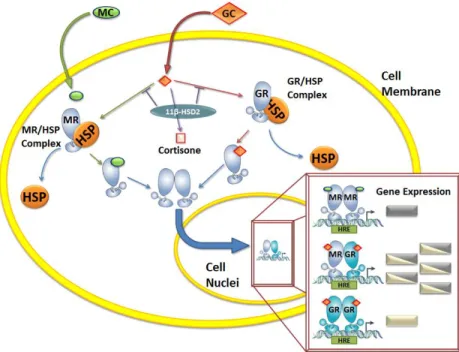
(17) Figure 2: Corticoid signal transduction. Circulating corticoids diffuse across the cell membrane. While mineralocorticoids (MC) almost exclusively bind to the mineralocorticoid receptor (MR), glucocorticoids (GC) can bind either to MR or to the glucocorticoid receptor (GR) creating a diverse gene induction (extracted from Muñoz-Durango, et al 2015).. 3. Models to study immunological role of aldosterone and MR The study of MR stimulation and antagonism have been focused in hypertension and cardiovascular system, due that its mainly role is to keep blood pressure and electrolyte homeostasis. However, recent studies have reported that not only kidney and colonic epithelial cells express MR, also several non-epithelial tissues, such as cardiomyocytes (39), blood vessels (40), cells of the nervous system (41), adipose tissue (42), liver (43) and immune cells (44,45) express MR and respond to aldosterone, supporting the inflammatory role of aldosterone and its receptor (22). In the context of the immune response, it has been consistently reported that aldosterone stimulation promotes proinflammatory responses in. 17.
(18) various tissues (44,46). In human leucocytes, MR expression has been reported in CD34+ hematopoietic progenitor, also in peripheral blood T and B lymphocytes, monocytes, and neutrophils (47). Further, clinical studies have demonstrated that MR antagonism in cardiovascular diseases can generate a beneficial outcome in patients, mainly due to the prevention of inflammatory damage (48).. 3.1. Immune cells stimulation by aldosterone. There are some In vitro studies directed to understand the mechanism by which aldosterone induces an inflammatory phenotype. Peritoneal macrophages isolated from wild type mice and treated with aldosterone displayed a “classical” activation profile, in which Tnf𝛼, Rantes, Mcp1, and Il12 expression was induced (45). This phenotype was prevented after eplerenone pretreatment, indicating that proinflammatory cytokine expression was via MR [25]. In addition, we reported that murine dendritic cells express MR and respond to aldosterone stimulation by inducing the secretion of IL-6 and TGF-𝛽 via mitogen-activated protein kinases (MAPKs) p38 y ERK1/2 (44). Both cytokines induced T cell polarization towards Th17 phenotype and enhance autoimmune damage in an in vivo model of experimental autoimmune encephalomyelitis (EAE) (44). The importance of the above findings is because both, macrophages and DCs, are key cells for the development of adaptive immunity, and their function could be modulated by aldosterone. In this line, it was reported that balance between Th17 pathogenic and T regulatory cells during hypertension mediated by DOCA-salt in rats is altered, resulting in lower ratio of Th17/Treg in PBMCs, kidney, and heart of hypertensive rats (49). However, the Th17/Treg ratio was restored in those animals treated with MR antagonism (spironolactone) (49).. 18.
(19) 3.2. Pharmacologic MR antagonism in immune cells. Two generations of MR antagonists have been developed. The first generation included spironolactone and canrenone, two potent steroidal compounds that are also androgen receptor antagonists and progesterone receptor agonists, both of them producing several active metabolites (50). More recently, a new steroidal MR antagonist was developed: eplerenone, which is less potent than the first generation and has a shorter halflife but is more selective than the previous compounds for the MR and generates no other active metabolites (50).. In vitro studies performed in human peripheral blood mononuclear cells (PBMCs) stimulated with spironolactone showed an immune-modulatory effect per se, in which proinflammatory cytokines related with T helper 1 (Th1) immune response, such TNF-𝛼 and IFN-𝛾, were decreased in contrast toTh2 and anti-inflammatory cytokine expression and production (51,52). Another study showed that spironolactone can modulate the function of transcription factors that control the immune response, such as NF𝜅 B, CEBP𝛽, and MYC (53). Along these lines, experiments made in human monocytes treated with eplerenone showed that MR antagonism helps to modulate the type of macrophage activation, turning those cells into an alternative activation phenotype (54). Although these studies clearly associate MR antagonism with anti-inflammatory effects, neither of them showed changes in IL-1𝛽 expression, suggesting that MR antagonism does not inhibit the inflammasome pathway (52,53), or the production of cytokines related with alternative macrophage polarization, including IL-4 or the anti-inflammatory cytokines TGF-𝛽 and IL-10 (45,52). In contrast, it has been described that treatment of peritoneal macrophages with low doses of corticosterone (10 nM) increases the expression of Tnf𝛼 and this proinflammatory cytokine only returns to basal levels when GR but not MR is blocked (55).. 19.
(20) In addition, studies performed in hypertensive animal models treated with spironolactone or eplerenone have revealed that these drugs protect against inflammation and end-organ damage, rather than preventing the occurrence of hypertension (56,57). For example, it was reported that eplerenone antagonism prevents vascular wall inflammation and fibrosis (58,59) and reduced the gene expression of proinflammatory molecules, such Cox2, Mcp1, and osteopontin (Opn) (57).. 3.3. MR knockout animal models. The development of myeloid conditional knockout MR (MyMRKO) mice has allowed to study the impact of MR in immune cells. As expected, first reports using MyMRKO mice was in animal models of hypertension (45,60), then in chronic inflammatory diseases such EAE animal models (61). All abovementioned models agree with MR is closely related with proinflammatory genes expression inducing disease, because in the absence of MR in all diseases displayed improvements at different levels.. MyMRKO is a double mutant mouse build by Cre/loxP recombination system (62). This animal contains loxP sites flanking the exon 3 of the Nr3c2 gene encoding the MR, resulting in “floxed” MR or MRloxP/loxP (63). The addition of this loxP sites are necessary to further induction of conditionally deletion, because loxP sites are recognized by Cre recombinase enzyme. Cre is also genetically introduced under a lineage specific or cell-type promoter to be conditionally expressed. Because it was attempted to develop myeloid conditional MR knockout mice, MRloxP/loxP mice was crossed with mice expressing Cre recombinase enzyme under the control of the myeloid lineage-specific promoter lysozyme M (LysM-Cre) (64). Then, only mice that simultaneously express Cre recombinase and. 20.
(21) presented loxP sequences will display MyMRKO phenotype. But, whether mice only presented loxP sequences are considered litter mate (LM) (45,63) (Figure 3).. Because our aim was studying the role of aldosterone-MR as modulators of immune response, we used MyMRKO mice to described its immunologic profile in response to TLR4 agonist (LPS) and in response to chronic inflammatory aseptic disease: Non-Alcoholic Steatohepatitis (NASH).. Single mutant mice: Floxed. Double mutant mice: null. 1. Homozygous for MRLoxP/LoxP. 1. Homozygous for MRLoxP/LoxP 2. Hemizygous for LysMCre. Parents Clausen, B. 1999 Usher, M. J. of Clin Invest. 2010. Berger, S. PNAS. 2006. LysM promotor. Lysozyme M. LysM promotor. Lysozyme M. X. LysM promotor LysM promotor. Lysozyme M. MR MR. MR MR. ~ 50% LM. F1. Cre. LysM promotor LysM promotor. Lysozyme M. ~ 50% MyMRKO. LysM promotor LysM promotor. Lysozyme M. MR MR. Cre Lysozyme M. MR. Figure 3: Breading strategy of myeloid MR conditional knockout mice (MyMRKO) and litter Mate (LM) mice.. 21.
(22) GENERAL HYPOTHESIS. Aldosterone and its cognate receptor, mineralocorticoid receptor (MR), are involved in the expression of Pattern Receptor Recognition in dendritic cells, modulating the way of these cells are sensing antigens and affecting its capacity to drive adaptive immune response.. GENERAL AIMS. To characterize in vitro whether dendritic cells derived from conditional myeloid MR knockout mice (MyMRKO) displayed an impaired capacity to responds against inflammatory challenge and drive adaptive immune response.. To evaluate in vivo whether the lack of MR in myeloid cells altered the outcome of MyMRKO in endotoxemia and non-alcoholic steatohepatitis animal models.. 22.
(23) GENERAL DISCUSSION The close relationship between immune and endocrine systems have been explored for years, however, it is remains unknown the precisely mechanisms controlling its complex and diverse interaction. In present study we sought to characterize whether MR-aldosterone axis was involved in the sensing danger capacity of DCs and its impact driving adaptive immune response. Our main proposal is based in the well documented information linking aldosterone via MR with the onset and maintenance of inflammation (46). All of these studies suggest that main mechanism of inflammation is related with oxidative stress, fibrosis and proinflammatory cytokines induction. These features are switched toward anti-inflammatory profile using MR antagonism, silencing or Knockout, indicating that are mediate by aldosterone via MR.. Previous to immune response induction, it is necessary that innate immune cells sense damage signals (65). This sensing is mediated by group of receptors named PRRs which has been broadly described, but little is reported about its regulation. Because MR is a nuclear transcription and has the capacity to induce gene expression, we focused our research to describe whether aldosterone-MR axis was involved in the expression Tlr4 expression on DCs. This link was previously suggested in a model of aldosterone/saltsensitive hypertension, in which experimental hyperaldosteronism upregulated TLR4 expression in kidneys and hearth and inducing systemic inflammation (TNF-a and IL-1b). Cardiac and renal fibrosis and systemic inflammation was prevented using TLR4 signaling antagonist, TAK-242 (66). This cooperative relation between inflammatory cytokines and TLRs expression was also described in TLR2 promoter. This promoter presents binding sites for GRE, NF-κB and STAT5, which are occupied after dexamethasone (glucocorticoids) and TNF-α stimulation controlling the expression of TLR2 (67–69).. 23.
(24) Our results suggest that aldosterone via MR could be involved in Tlr4 expression in LM/WT-DCs after aldosterone stimulation. Opposite effect was obtained from MyMRKODCs because these cells did not increase their levels of Tlr4, indeed displayed lower levels of Tlr4 than basal. More interesting, when DCs were challenged with low doses of LPS showed that MyMRKO-DCs displayed lower capacity to secrete proinflammatory cytokines and express lower costimulatory molecules that LM-DCs. These results fix with the definition of tolerogenic profile of DCs (14), suggesting that MR could be playing a physiologically role supporting the induction of immunogenic immune response, completely opposed to immunosuppressive response induced through GR (17,18).. Because DCs profile directly impacts the polarization of adaptive immune response, we performed coculture experiments with CD4+ and CD8+ T Lymphocytes. According to our results, “tolerogenic” profile of MyMRKO-DCs impacts only CD8+ lymphocytes activation and function. This result also was observed in vivo animal model of NASH, in which lower function of CD8+ T Lymphocytes was associated to phenotype of lower steatosis. CD8+ T Lymphocytes are activated by APCs via major histocompatibility complex – class I (MHC-I) (65,70). Although we did not explore the function of class I antigens processing machinery, we can suggest that MR could be involved, opening an interesting field of research.. Summarizing our results, we proposed that MR under steady state are played a key role maintaining the capacity of DCs to stablish immunogenic immune response, because its lack impacted in the manner of damage is sense, directly affecting the adaptive CD8+ immune response.. 24.
(25) GENERAL PROPOSED MODEL Detrimental effects in MyMRKO mice. Positive effects in MyMRKO mice. MyMRKO DCs. CD8+ T Lymphocytes. 3.. 2.. High plasma aldosterone activated MR. -Lower CD86 -Impaired capacity to activate CD8+. -CD25 -CD69 -Lower migration Liver steatosis. 1. Hepatocytes. Hepatic stellated cells: Fibrosis and inflammation Endothelial cells and hepatocytes: Inflammatory cytokines and immune cells infiltration. -Lower expression of Tlr4 -Impaired capacity to responds TLR4 agonist. Collagen Extracellular matrix. Current work described that MyMRKO DCs presented an altered capacity to sense and responds against inflammatory stimulus, impacting its capacity to develop adaptive immune response. 1. We described that MyMRKO DCs expressed lower levels of Tlr4 that its counterpart LM, for that, its capacity to sense PAMPS and DAMPS could be reduced. This result agrees with its tolerogenic profile, characterized by lower expression of costimulatory molecules and cytokine secretion in response to LPS challenge. 2. Then, same results were found in NASH animal model. Here, rather to describes the lower capacity of liver MyMRKO DCs to express costimulatory molecules, we displayed in vivo and in vitro its impaired capacity to drive effective CD8+ immune response (required to develop lipid storage in hepatocytes). On the other hand, 3. We also observed that NASH occurring with high levels of circulating aldosterone in both, MyMRKO and LM mice. Despite MyMRKO displayed lower liver steatosis and inflammation, higher levels of aldosterone were associated with similar grade of fibrosis. These results suggest that other cells in liver, such as hepatic stellated cells, Kupffer cells, endothelial cells and hepatocytes can responds to aldosterone.. In summary, we described some results which support that myeloid MR could be playing an important role in the way that DCs are recognizing and presenting antigens to CD8+ Lymphocytes.. 25.
(26) GENERAL BIBLIOGRAPHY 1.. Janeway CA. How the immune system works to protect the host from infection: A personal view. Proc Natl Acad Sci [Internet]. 2001 Jun 19;98(13):7461 LP – 7468. Available from: http://www.pnas.org/content/98/13/7461.abstract. 2.. Parkin J, Cohen B. An overview of the immune system. Lancet (London, England) [Internet]. 2001 Jun 2 [cited 2019 Oct 21];357(9270):1777–89. Available from: http://www.ncbi.nlm.nih.gov/pubmed/11403834. 3.. A current view on inflammation. Nat Immunol [Internet]. 2017;18(8):825. Available from: https://doi.org/10.1038/ni.3798. 4.. Janeway CA, Medzhitov R. Innate Immune Recognition. Annu Rev Immunol [Internet]. 2002;20(1):197–216. Available from: https://doi.org/10.1146/annurev.immunol.20.083001.084359. 5.. Hopkins PA, Sriskandan S. Mammalian Toll-like receptors: to immunity and beyond. Clin Exp Immunol [Internet]. 2005 Jun;140(3):395–407. Available from: https://www.ncbi.nlm.nih.gov/pubmed/15932500. 6.. Aderem A, Ulevitch RJ. Toll-like receptors in the induction of the innate immune response. Nature [Internet]. 2000;406(6797):782–7. Available from: https://doi.org/10.1038/35021228. 7.. Laird MHW, Rhee SH, Perkins DJ, Medvedev AE, Piao W, Fenton MJ, et al. TLR4/MyD88/PI3K interactions regulate TLR4 signaling. J Leukoc Biol [Internet]. 2009;85(6):966–77. Available from: https://jlb.onlinelibrary.wiley.com/doi/abs/10.1189/jlb.1208763. 8.. Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol [Internet]. 2010 Apr 20;11:373. Available from: https://doi.org/10.1038/ni.1863. 9.. Bianchi ME. DAMPs, PAMPs and alarmins: all we need to know about danger. J Leukoc Biol [Internet]. 2007;81(1):1–5. Available from: https://jlb.onlinelibrary.wiley.com/doi/abs/10.1189/jlb.0306164. 10.. Chen GY, Nuñez G. Sterile inflammation: sensing and reacting to damage. Nat Rev Immunol [Internet]. 2010;10(12):826–37. Available from: https://doi.org/10.1038/nri2873. 11.. Iwasaki A, Medzhitov R. Regulation of Adaptive Immunity by the Innate Immune System. Science (80- ) [Internet]. 2010 Jan 15;327(5963):291 LP – 295. Available from: http://science.sciencemag.org/content/327/5963/291.abstract. 12.. Kufer TA, Creagh EM, Bryant CE. Guardians of the Cell: Effector-Triggered Immunity Steers Mammalian Immune Defense. Trends Immunol [Internet]. [cited 2019 Oct 21];0(0). Available from: https://www.cell.com/trends/immunology/fulltext/S1471-4906(19)301681?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS1471490 619301681%3Fshowall%3Dtrue#. 26.
(27) 13.. Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature [Internet]. 1998;392(6673):245–52. Available from: https://doi.org/10.1038/32588. 14.. Audiger C, Rahman MJ, Yun TJ, Tarbell K V, Lesage S. The Importance of Dendritic Cells in Maintaining Immune Tolerance. J Immunol [Internet]. 2017 Mar 15;198(6):2223 LP – 2231. Available from: http://www.jimmunol.org/content/198/6/2223.abstract. 15.. Probst HC, McCoy K, Okazaki T, Honjo T, van den Broek M. Resting dendritic cells induce peripheral CD8+ T cell tolerance through PD-1 and CTLA-4. Nat Immunol [Internet]. 2005;6(3):280–6. Available from: https://doi.org/10.1038/ni1165. 16.. Yamazaki T, Akiba H, Iwai H, Matsuda H, Aoki M, Tanno Y, et al. Expression of Programmed Death 1 Ligands by Murine T Cells and APC. J Immunol [Internet]. 2002 Nov 15;169(10):5538 LP – 5545. Available from: http://www.jimmunol.org/content/169/10/5538.abstract. 17.. Waage A, Slupphaug G, Shalaby R. Glucocorticoids inhibit the production of IL 6 from monocytes, endothelial cells and fibroblasts. Eur J Immunol [Internet]. 1990 Nov 1;20(11):2439–43. Available from: https://doi.org/10.1002/eji.1830201112. 18.. Blotta MH, DeKruyff RH, Umetsu DT. Corticosteroids inhibit IL-12 production in human monocytes and enhance their capacity to induce IL-4 synthesis in CD4+ lymphocytes. J Immunol [Internet]. 1997 Jun 15;158(12):5589 LP – 5595. Available from: http://www.jimmunol.org/content/158/12/5589.abstract. 19.. Evans-Storms RB, Cidlowski JA. Regulation of apoptosis by steroid hormones. J Steroid Biochem Mol Biol [Internet]. 1995 Jun 1 [cited 2019 Oct 21];53(1–6):1–8. Available from: https://www.sciencedirect.com/science/article/pii/096007609500034W?via%3Dihub. 20.. Hermoso MA, Cidlowski JA. Putting the brake on inflammatory responses: the role of glucocorticoids. IUBMB Life [Internet]. 2003;55(9):497—504. Available from: https://doi.org/10.1080/15216540310001642072. 21.. Pearce D, Yamamoto KR. Mineralocorticoid and glucocorticoid receptor activities distinguished by nonreceptor factors at a composite response element. Science (80- ) [Internet]. 1993 Feb 19;259(5098):1161 LP – 1165. Available from: http://science.sciencemag.org/content/259/5098/1161.abstract. 22.. Muñoz-Durango, N; Vecchiola, A; Gonzalez-Gomez, L.M; Simon, F; Riedel, C.A; Fardella, C.E. and Kalergis A. Modulation of Immunity and Inflammation by the Mineralocorticoid Receptor and Aldosterone. Biomed Res Int [Internet]. 2015;2015:14. Available from: https://doi.org/10.1155/2015/652738. 23.. Müller-Fielitz H, Lau M, Jöhren O, Stellmacher F, Schwaninger M, Raasch W. Blood pressure response to angiotensin II is enhanced in obese Zucker rats and is attributed to an aldosterone-dependent mechanism. Br J Pharmacol [Internet]. 2012;166(8):2417–29. Available from: https://bpspubs.onlinelibrary.wiley.com/doi/abs/10.1111/j.14765381.2012.01953.x. 27.
(28) 24.. Funder JW. Mineralocorticoids, glucocorticoids, receptors and response elements. Science (80- ) [Internet]. 1993 Feb 19;259(5098):1132 LP – 1133. Available from: http://science.sciencemag.org/content/259/5098/1132.abstract. 25.. Tomaschitz A, Pilz S, Ritz E, Obermayer-Pietsch B, Pieber TR. Aldosterone and arterial hypertension. Nat Rev Endocrinol [Internet]. 2010;6(2):83—93. Available from: https://doi.org/10.1038/nrendo.2009.263. 26.. Shavit L, Lifschitz MD, Epstein M. Aldosterone blockade and the mineralocorticoid receptor in the management of chronic kidney disease: current concepts and emerging treatment paradigms. Kidney Int [Internet]. 2012 May 2;81(10):955–68. Available from: https://doi.org/10.1038/ki.2011.505. 27.. Funder JW. Aldosterone and the Cardiovascular System: Genomic and Nongenomic Effects. Endocrinology [Internet]. 2006 Dec 1;147(12):5564–7. Available from: https://doi.org/10.1210/en.2006-0826. 28.. Viengchareun S, Le Menuet D, Martinerie L, Munier M, Pascual-Le Tallec L, Lombès M. The mineralocorticoid receptor: insights into its molecular and (patho)physiological biology. Nucl Recept Signal [Internet]. 2007 Nov 30;5:e012–e012. Available from: https://www.ncbi.nlm.nih.gov/pubmed/18174920. 29.. Arriza JL, Weinberger C, Cerelli G, Glaser TM, Handelin BL, Housman DE, et al. Cloning of human mineralocorticoid receptor complementary DNA: structural and functional kinship with the glucocorticoid receptor. Science (80- ) [Internet]. 1987 Jul 17;237(4812):268 LP – 275. Available from: http://science.sciencemag.org/content/237/4812/268.abstract. 30.. Gomez-Sanchez CE, de Rodriguez AF, Romero DG, Estess J, Warden MP, GomezSanchez MT, et al. Development of a Panel of Monoclonal Antibodies against the Mineralocorticoid Receptor. Endocrinology [Internet]. 2006 Mar 1;147(3):1343–8. Available from: https://doi.org/10.1210/en.2005-0860. 31.. Ou X-M, Storring JM, Kushwaha N, Albert PR. Heterodimerization of Mineralocorticoid and Glucocorticoid Receptors at a Novel Negative Response Element of the 5-HT1A Receptor Gene. J Biol Chem [Internet]. 2001 Apr 27 [cited 2019 Sep 6];276(17):14299–307. Available from: http://www.jbc.org/cgi/content/short/276/17/14299. 32.. Lombès M, Binart N, Delahaye F, Baulieu EE, Rafestin-Oblin ME. Differential intracellular localization of human mineralocorticosteroid receptor on binding of agonists and antagonists. Biochem J [Internet]. 1994 Aug 15;302(1):191 LP – 197. Available from: http://www.biochemj.org/content/302/1/191.abstract. 33.. Náray-Fejes-Tóth A, Fejes-Tóth G. The <em>sgk</em>, an aldosterone-induced gene in mineralocorticoid target cells, regulates the epithelial sodium channel. Kidney Int [Internet]. 2000 Apr 1;57(4):1290–4. Available from: https://doi.org/10.1046/j.1523-1755.2000.00964.x. 34.. Derfoul A, Robertson NM, Lingrel JB, Hall DJ, Litwack G. Regulation of the human Na/KATPase beta1 gene promoter by mineralocorticoid and glucocorticoid receptors. J Biol. 28.
(29) Chem [Internet]. 1998 Aug 14 [cited 2019 Sep 8];273(33):20702–11. Available from: http://www.ncbi.nlm.nih.gov/pubmed/9694812 35.. Grossmann C, Krug AW, Freudinger R, Mildenberger S, Voelker K, Gekle M. Aldosteroneinduced EGFR expression: interaction between the human mineralocorticoid receptor and the human EGFR promoter. Am J Physiol Metab [Internet]. 2007;292(6):E1790–800. Available from: https://doi.org/10.1152/ajpendo.00708.2006. 36.. Yang J, Young MJ. The mineralocorticoid receptor and its coregulators. J Mol Endocrinol. 2009;43(2).. 37.. Sutanto W, De Kloet ER. Mineralocorticoid receptor ligands: Biochemical, pharmacological, and clinical aspects. Med Res Rev [Internet]. 1991;11(6):617–39. Available from: https://onlinelibrary.wiley.com/doi/abs/10.1002/med.2610110604. 38.. Funder JW, Pearce PT, Smith R, Smith AI. Mineralocorticoid action: target tissue specificity is enzyme, not receptor, mediated. Science (80- ) [Internet]. 1988 Oct 28;242(4878):583 LP – 585. Available from: http://science.sciencemag.org/content/242/4878/583.abstract. 39.. Lombès M, Alfaidy N, Eugene E, Lessana A, Farman N, Bonvalet J-P. Prerequisite for Cardiac Aldosterone Action. Circulation. 1995;92(2):175–82.. 40.. Identification and role of aldosterone receptors in the cardiovascular system. Ann Endocrinol (Paris). 2000 Apr;820(1):5 YP – 85.. 41.. Meijer OC. Coregulator Proteins and Corticosteroid Action in the Brain. J Neuroendocrinol [Internet]. 2002;14(6):499–505. Available from: https://onlinelibrary.wiley.com/doi/abs/10.1046/j.1365-2826.2002.00795.x. 42.. Caprio M, Fève B, Claës A, Viengchareun S, Lombès M, Zennaro M-C. Pivotal role of the mineralocorticoid receptor in corticosteroid-induced adipogenesis. FASEB J [Internet]. 2007;21(9):2185–94. Available from: https://doi.org/10.1096/fj.06-7970com. 43.. Fujisawa G, Muto S, Okada K, Kusano E, Ishibashi S. Mineralocorticoid receptor antagonist spironolactone prevents pig serum-induced hepatic fibrosis in rats. Transl Res [Internet]. 2006 Sep 1;148(3):149–56. Available from: https://doi.org/10.1016/j.trsl.2006.03.007. 44.. Herrada AA, Contreras FJ, Marini NP, Amador CA, González PA, Cortés CM, et al. Aldosterone Promotes Autoimmune Damage by Enhancing Th17-Mediated Immunity. J Immunol [Internet]. 2010 Jan 1;184(1):191 LP – 202. Available from: http://www.jimmunol.org/content/184/1/191.abstract. 45.. Usher MG, Duan SZ, Ivaschenko CY, Frieler RA, Berger S, Schütz G, et al. Myeloid mineralocorticoid receptor controls macrophage polarization and cardiovascular hypertrophy and remodeling in mice. J Clin Invest [Internet]. 2010 Sep 1 [cited 2019 Sep 3];120(9):3350– 64. Available from: https://www.jci.org/articles/view/41080#.XW73xbLCq8I.mendeley. 46.. Munoz-Durango N, Barake MF, Letelier NA, Campino C, Kalergis CEF and AM. Immune System Alterations by Aldosterone During Hypertension: From Clinical Observations to Genomic and Non-Genomic Mechanisms Leading to Vascular Damage [Internet]. Vol. 13,. 29.
(30) Current Molecular Medicine. 2013. p. 1035–46. Available from: http://www.eurekaselect.com/node/111667/article 47.. Grafte-Faure S, Leveque C, Vasse M, Soria C, Norris V, Vannier J-P. Effects of glucocorticoids and mineralocorticoids on proliferation and maturation of human peripheral blood stem cells. Am J Hematol [Internet]. 1999 Oct 1;62(2):65–73. Available from: https://doi.org/10.1002/(SICI)1096-8652(199910)62:2%3C65::AID-AJH1%3E3.0.CO. 48.. Rossignol P, Ménard J, Fay R, Gustafsson F, Pitt B, Zannad F. Eplerenone Survival Benefits in Heart Failure Patients Post-Myocardial Infarction Are Independent From its Diuretic and Potassium-Sparing Effects: Insights From an EPHESUS (Eplerenone Post-Acute Myocardial Infarction Heart Failure Efficacy and Survival Study) Substudy. J Am Coll Cardiol [Internet]. 2011 Nov 1 [cited 2019 Oct 21];58(19):1958–66. Available from: https://www.sciencedirect.com/science/article/pii/S0735109711029883?via%3Dihub. 49.. Amador CA, Barrientos V, Peña J, Herrada AA, González M, Valdés S, et al. Spironolactone Decreases DOCA–Salt–Induced Organ Damage by Blocking the Activation of T Helper 17 and the Downregulation of Regulatory T Lymphocytes. Hypertension. 2014;63(4):797–803.. 50.. Kolkhof P, Borden SA. Molecular pharmacology of the mineralocorticoid receptor: Prospects for novel therapeutics. Mol Cell Endocrinol [Internet]. 2012 Mar 24 [cited 2019 Oct 21];350(2):310–7. Available from: https://www.sciencedirect.com/science/article/pii/S0303720711003583?via%3Dihub. 51.. Hansen PR, Rieneck K, Bendtzen K. Spironolactone inhibits production of proinflammatory cytokines by human mononuclear cells. Immunol Lett [Internet]. 2004 Feb 15 [cited 2019 Oct 21];91(2–3):87–91. Available from: https://www.sciencedirect.com/science/article/pii/S0165247803002761?via%3Dihub. 52.. BENDTZEN K, HANSEN PR, RIENECK K, GROUP THESS. Spironolactone inhibits production of proinflammatory cytokines, including tumour necrosis factor-α and interferon-γ, and has potential in the treatment of arthritis. Clin Exp Immunol [Internet]. 2003 Oct 1;134(1):151–8. Available from: https://doi.org/10.1046/j.1365-2249.2003.02249.x. 53.. Sønder SUS, Mikkelsen M, Rieneck K, Hedegaard CJ, Bendtzen K. Effects of spironolactone on human blood mononuclear cells: mineralocorticoid receptor independent effects on gene expression and late apoptosis induction. Br J Pharmacol [Internet]. 2006 May 1;148(1):46–53. Available from: https://doi.org/10.1038/sj.bjp.0706700. 54.. Łabuzek K, Liber S, Bułdak Ł, Machnik G, Liber J, Okopień B. Eplerenone promotes alternative activation in human monocyte-derived macrophages. Pharmacol Reports [Internet]. 2013;65(1):226–34. Available from: http://www.sciencedirect.com/science/article/pii/S1734114013709836. 55.. Lim H-Y, Müller N, Herold MJ, van den Brandt J, Reichardt HM. Glucocorticoids exert opposing effects on macrophage function dependent on their concentration. Immunology. 30.
(31) [Internet]. 2007/04/23. 2007 Sep;122(1):47–53. Available from: https://www.ncbi.nlm.nih.gov/pubmed/17451463 56.. Ricardo R, N. CP, Kavita K, Andrea Z, T. SC. Mineralocorticoid Blockade Reduces Vascular Injury in Stroke-Prone Hypertensive Rats. Hypertension [Internet]. 1998 Jan 1;31(1):451–8. Available from: https://doi.org/10.1161/01.HYP.31.1.451. 57.. Rocha R, Rudolph AE, Frierdich GE, Nachowiak DA, Kekec BK, Blomme EAG, et al. Aldosterone induces a vascular inflammatory phenotype in the rat heart. Am J Physiol Circ Physiol [Internet]. 2002 Nov 1;283(5):H1802–10. Available from: https://doi.org/10.1152/ajpheart.01096.2001. 58.. Lam EYM, Funder JW, Nikolic-Paterson DJ, Fuller PJ, Young MJ. Mineralocorticoid Receptor Blockade But Not Steroid Withdrawal Reverses Renal Fibrosis in Deoxycorticosterone/Salt Rats. Endocrinology [Internet]. 2006 Jul 1;147(7):3623–9. Available from: https://doi.org/10.1210/en.2005-1527. 59.. Rickard AJ, Funder JW, Morgan J, Fuller PJ, Young MJ. Does Glucocorticoid Receptor Blockade Exacerbate Tissue Damage after Mineralocorticoid/Salt Administration? Endocrinology [Internet]. 2007 Oct 1;148(10):4829–35. Available from: https://doi.org/10.1210/en.2007-0209. 60.. Rickard AJ, Morgan J, Tesch G, Funder JW, Fuller PJ, Young MJ. Deletion of Mineralocorticoid Receptors From Macrophages Protects Against Deoxycorticosterone/SaltInduced Cardiac Fibrosis and Increased Blood Pressure. Hypertension [Internet]. 2009 Sep 1 [cited 2019 Sep 3];54(3):537–43. Available from: https://www.ahajournals.org/doi/10.1161/HYPERTENSIONAHA.109.131110. 61.. Montes-Cobos E, Schweingruber N, Li X, Fischer HJ, Reichardt HM, Lühder F. Deletion of the Mineralocorticoid Receptor in Myeloid Cells Attenuates Central Nervous System Autoimmunity. Front Immunol [Internet]. 2017;8:1319. Available from: https://www.frontiersin.org/article/10.3389/fimmu.2017.01319. 62.. Kuhn R, Schwenk F, Aguet M, Rajewsky K. Inducible gene targeting in mice. Science (80- ) [Internet]. 1995 Sep 8;269(5229):1427 LP – 1429. Available from: http://science.sciencemag.org/content/269/5229/1427.abstract. 63.. Berger S, Wolfer DP, Selbach O, Alter H, Erdmann G, Reichardt HM, et al. Loss of the limbic mineralocorticoid receptor impairs behavioral plasticity. Proc Natl Acad Sci U S A [Internet]. 2006 Jan 3;103(1):195 LP – 200. Available from: http://www.pnas.org/content/103/1/195.abstract. 64.. Clausen BE, Burkhardt C, Reith W, Renkawitz R, Förster I. Conditional gene targeting in macrophages and granulocytes using LysMcre mice. Transgenic Res [Internet]. 1999;8(4):265–77. Available from: https://doi.org/10.1023/A:1008942828960. 65.. Janeway Jr. CA, Medzhitov R. Introduction: The role of innate immunity in the adaptive immune response. Semin Immunol [Internet]. 1998 Oct 1 [cited 2019 Sep 3];10(5):349–50.. 31.
(32) Available from: https://www.sciencedirect.com/science/article/pii/S1044532398901427?via%3Dihub 66.. Zhang Y, Peng W, Ao X, Dai H, Yuan L, Huang X, et al. TAK-242, a Toll-Like Receptor 4 Antagonist, Protects against Aldosterone-Induced Cardiac and Renal Injury. Joles JA, editor. PLoS One [Internet]. 2015 Nov 10 [cited 2019 Sep 4];10(11):e0142456. Available from: https://dx.plos.org/10.1371/journal.pone.0142456. 67.. Hermoso MA, Matsuguchi T, Smoak K, Cidlowski JA. Glucocorticoids and Tumor Necrosis Factor Alpha Cooperatively Regulate Toll-Like Receptor 2 Gene Expression. Mol Cell Biol [Internet]. 2004 Jun 1;24(11):4743 LP – 4756. Available from: http://mcb.asm.org/content/24/11/4743.abstract. 68.. Wang T, Lafuse WP, Zwilling BS. NFκB and Sp1 Elements Are Necessary for Maximal Transcription of Toll-like Receptor 2 Induced by <em>Mycobacterium avium</em> J Immunol [Internet]. 2001 Dec 15;167(12):6924 LP – 6932. Available from: http://www.jimmunol.org/content/167/12/6924.abstract. 69.. Musikacharoen T, Matsuguchi T, Kikuchi T, Yoshikai Y. NF-κB and STAT5 Play Important Roles in the Regulation of Mouse Toll-Like Receptor 2 Gene Expression. J Immunol [Internet]. 2001 Apr 1;166(7):4516 LP – 4524. Available from: http://www.jimmunol.org/content/166/7/4516.abstract. 70.. López de Castro JA, Alvarez-Navarro C, Brito A, Guasp P, Martín-Esteban A, Sanz-Bravo A. Molecular and pathogenic effects of endoplasmic reticulum aminopeptidases ERAP1 and ERAP2 in MHC-I-associated inflammatory disorders: Towards a unifying view. Mol Immunol [Internet]. 2016 Sep 1 [cited 2019 Oct 22];77:193–204. Available from: https://www.sciencedirect.com/science/article/pii/S0161589016301535?via%3Dihub. 32.
(33) Chapter I. The axis aldosterone - Mineralocorticoid Receptor (MR) is involved in the regulation of Tlr4 expression in dendritic cells impacting the in vivo response to LPS.. Natalia Muñoz-Durango1, Andrea Vecchiola2, Felipe Simon3, Carlos E. Fardella2 and Alexis M. Kalergis1,2.. 1. Millennium Institute on Immunology and Immuotherapy, Departamento de Genética. Molecular y Microbiología, Pontificia Universidad Católica de Chile. 2. Millennium Institute on Immunology and Immuotherapy, Departamento de Endocrinología,. Pontificia Universidad Católica de Chile. 3. Instituto Milenio en Inmunología e Inmunoterapia, Departamento de Ciencias Biológicas,. Facultad de Medicina, Universidad Andrés Bello.. ABSTRACT The mineralocorticoid receptor (MR) is a ligand dependent transcription factor. MR has been conventionally related with the control of water and electrolyte homeostasis to keep blood pressure via aldosterone activation. Because MR is also expressed in cells of the immune system, there is a recent interest to studying its role in the modulation of immune response. Up to date, it has been described that high levels of aldosterone in vivo and in. 33.
(34) vitro are related with the induction of inflammatory phenotype in cardiac diseases, atherosclerosis, obesity and autoimmunity. We previously showed that dendritic cells (DCs) express MR and respond to its stimulation with aldosterone by secreting IL-6 and TGFb, leading to Th17 polarization. Otherwise, it has been described that conditional MR knockout in myeloid cells are involved in the tuning control between classical or alternative macrophage polarization. However, the effect of aldosterone excess, MR lacking or antagonism in myeloid cells, only have been explained from inflammatory phenotype resulted, rather than its specific role modulating the expression of molecules that are directly involved in the onset of immune response, such as Toll like receptors. Here we evaluate the role of the aldosterone-MR axis controlling the expression of TLR4. We found that aldosterone via MR induces the expression of Tlr4 in wild type bone marrow-derived DCs. Oppositely, the Trl4 expression in DCs derived from myeloid MR conditional knockout mice (MyMRKO) did not show to be modulated by aldosterone, indeed these cells displayed impaired capacity to response to LPS stimulation. Finally, in a model of sub-lethal endotoxemia, we found that mice pre-treated with aldosterone succumbs after LPS challenge in contrast to placebo treated mice, mainly due to a multi-organic failure. In conclusion our results suggest that aldosterone-MR axis is involved in the regulation of tlr4 expression, and in consequently modulating the DCs response to TLR4 agonist, LPS.. INTRODUCTION Dendritic cells (DCs) are knowing as the most potent antigen presenting cells (APCs) in mammal immune systems (1). These cells are characterized by their unique ability to recognize and respond against many stimuli to induce an adaptive immune response, serving as a key component of innate immunity and bridge innate and adaptive immune response (1). The ability of DCs to recognize molecular patterns derived from pathogens. 34.
(35) (known as pathogen associated molecular patterns -PAMPs-) and from cellular components (known as damage associated molecular patterns -DAMPs-), is because DCs, as other innate cells, express a family of receptors which sense these molecules, named pathogen recognition receptors (PRRs) (2,3). From PRRs, the Toll like receptors (TLRs) are the most characterized family, because they are broadly expressed and they can easily sense and trigger the activation of innate response and also, drive the adaptive immune responses in several ways, including antibody responses (4) and T helper CD4+ and CD8+ cell responses (5,6).. Since the TLRs were described, research has been mainly focused on the discovery of their ligands and the signaling pathways triggered after ligand–TLR interaction, rather that its regulation (7,8). However, some efforts have been made to define regulatory factors involved in the control of TLRs expression in epithelial and immune cells concluding that inflammatory milieu is mandatory for TLRs regulation. Studies performed in type 1 diabetes (T1D) patients who are constantly under pro-inflammatory state, displayed higher levels of TLR4 and TLR2 in peripheral monocytes in contrast with controls (9) indicating that proinflammatory markers such as C reactive protein (CRP) are related with TLRs modulation. Then, authors also demonstrated that patients with T1D with microvascular complications, who presented even higher markers of CRP and IL-1b secretion, showed over-expressed levels of TLR4 and TLR2 in monocytes, as compared with T1D patients and controls (10). Also, Crohn´s Disease patients present higher levels of TLR4 expression in peripheral B cells and monocytes in comparison with healthy controls after cell stimulation with anti-Igµ, anti-CD40 and IL-4 (11). However, the regulation of TLR4 in each cell type uses differentially the transcription factors PU.1, interferon response factor (IRF) 8, IRF4 and cJun, to binds on TLR4 promotor (11). These results agree with previous data that described that TLR4 promoter presents several purine-rich sequences that are recognized. 35.
(36) by PU.1 transcription factor (12) and also sites for the binding of E twenty-six (Ets) and activator protein (AP)-1 transcription factors (13), all of them induced under inflammation. On the other hand, studies performed in the proximal promoter of murine TLR2 have shown that this promotor presents also binding sites for nuclear factor (NF)-κB and STAT5 that are implicated in the rapid induction of TLR2 mRNA expression by pro-inflammatory stimuli in murine macrophages (14,15). Additionally, Hermoso, et.al, demonstrated that TLR2 promoter also presents glucocorticoids response elements (GRE), which regulates positively the TLR2 expression in response to dexamethasone plus inflammatory milieu generated by TNF-α (16). Cumulative evidence indicates that TLRs regulation is closely related with inflammatory signals, due to existence of sites for transcription factors binding in TLR4 y TLR2 promotors that are induced by inflammatory signals.. Along these lines, aldosterone, which is a mineralocorticoid hormone that controls blood pressure through electrolyte homeostasis in renal tissue, also have been linked with inflammatory conditions in cardiovascular diseases (17). For example, Patients with resistant arterial hypertension with aldosterone-producing adenomas, secrete high levels of TNF-𝛼, IL-6 and IL-1𝛽 from monocytes and IL-2, IFN-𝛾 and TNF-𝛼 from lymphocytes. However, after treatment with MR antagonist or adrenalectomization, the levels of all these cytokines decreased significantly as equivalent to essential hypertensive (EH) patients or healthy controls (18). Further, we reported that primary aldosteronism patients displayed increased serum levels of TNF-𝛼 and IL-10, as well as lower serum levels of TGF-𝛽 1 compared to EH patients. Spironolactone treatment of PA patients restored serum levels of all three cytokines (19). Nevertheless, not only deregulated levels of aldosterone have been associated to inflammatory clinical effect during hypertension, also in sepsis and septic shock it has been reported a suprarenal dysfunction with altered levels of both, glucocorticoids and mineralocorticoids (20). Other study made in children with. 36.
(37) meningococcal sepsis showed an impaired adrenocortical function (21). Authors found that aldosterone levels tended to be higher in non-survivor patients of septic shock, as compared to surviving patients. Furthermore, aldosterone levels displayed a significant correlation with both, age and IL-6 levels (21). Another study using rat primary cultures of adrenal zona glomerulosa cells reported that Lipopolysaccharide (LPS) was able to stimulate aldosterone production via TLR2 and TLR4 (22). Finally, a recent clinical study directed to measure the risk to develop sepsis in patients with primary aldosteronism (PA) or aldosterone producing adenoma (APA), showed that both groups displayed same risk to developing sepsis or to dying by septic shock than essential hypertension (EH) controls (23). However, once PA and APA underwent to adrenalectomy, both groups displayed significant lower risk to develop sepsis or septic shock in contrast to EH controls. Authors attributed this drastic switch, in the inflammatory and oxidative detrimental effects related with aldosterone.. Due that aldosterone could be playing an important role in the onset and maintenance of inflammation, and this inflammatory milieu is necessary to control the TLRs expression, in the present study we sought to evaluate whether aldosterone directly modulates the TLR4 expression in DCs. Because the expression of that kind of receptor should be finely regulated, due the probability to TLRs activation in the absence of infection can lead to autoimmune and chronic inflammatory responses, we hypothesized that aldosterone through MR induces a pro-inflammatory state in DCs by promoting overexpression of TLR4. Thus, these changes will render DCs as hyper-sensible to both microbial and cellular agonists. Here, we found that aldosterone-MR are involved in Tlr4 expression in DCs, rendering these cells to be more sensitive to low doses of LPS, the TLR4 ligand. Finally, in a sub-lethal endotoxic animal model we evaluate the biological significances of the inflammatory aldosterone-mediated modulation of DCs.. 37.
(38) METODOLOGY Ethics statements.. All mice were maintained under pathogen-free conditions in the facilities of Pontificia Universidad Católica de Chile (Santiago, Chile). The protocols applied in this work agreed to institutional Animal Health Care Guidelines and were supervised by a veterinarian. The approved institutional protocol number is150519021-1.. Mice breeding and genotyping.. C57BL/6 wild-type mice were obtained from The Jackson Laboratory (Bar Harbor, ME) and were used at 6-8 weeks of age. To dissect the pathway by which aldosterone modulates tlr4 expression in DCs, we used MR knockout conditional mice in myeloid cells (MyMRKO) which was gently donated by donated by Dr. Richard M Mortensen (University of Michigan, Michigan, USA) and from MTA with Dr. Günther Schütz (DKFZ, Im Neuenheimer Feld, Heidelberg, Germany) (24). These conditional knockout mice were breeding by crossing floxed MRflox/flox mice with floxed MRflox/flox/Cre-recombinase mice which express this enzyme under LysM promoter (LysMCre) (24–26).. To determinate whether the genotype of mice corresponded to MyMRKO or its control Litter Mate (LM), all mice was genotyping by PCR from peripheral blood sample prior its uses with the following set of primers: Cre-Fw 5’-AGG TGT AGA GAA GGC ACT CAG C-3’, Cre-Rv 5’-CTA ATC GCC ATC TTC CAG CAG G-3’, the product size was 411bp and should be present only in Cre+ mice. As control gene was used MEH-Fw 5’-AAG TGA GTT TGC ATG GCG CAG C-3’, MEH-Rv 5’-CCC TTT AGC CCC TTC CCT CTG-3’, the product size was. 38.
(39) 341. bp. and. should. be. present. in. all. animals.. Primer. A. (MRflox). 5’-. CTGGAGATCTGAACTCCAGGCT-3’, Primer B (MRflox) 5’-TAG AAA CAC TTC GTA AAG TAG AGC T-3’, Primer C (MRflox) 5’-CCT AGA GTT CCT GAG CTG CTG A-3’, the product sizes were for the WT allele: 285 bp, for the Floxed allele: 335 bp and for the Null allele: 390 bp. The PCR amplification program for Cre and MEH genes were composed by: one cycle of incubation at 95ºC for 5 minutes, followed by 35 cycles of denaturation 95ºC during 30 seconds / annealing 55ºC during 30 seconds / extension 72ºC during 45 seconds and finalizing with 72ºC for 7 minutes. The PCR amplification program for Floxed MR gene was composed by: one cycle of incubation at 95ºC for 5 minutes, followed by 35 cycles of denaturation 95ºC during 30 seconds / annealing 63ºC during 60 seconds / extension 72ºC during 60 seconds and finalizing with 72ºC for 60 seconds.. In vitro and in vivo treatments with aldosterone, spironolactone and eplerenone.. For in vitro tests, aldosterone and its antagonists were prepared as previously described (27). Briefly, stock solutions of aldosterone (Sigma-Aldrich, St. Louis, MO), spironolactone (Sigma-Aldrich, St. Louis, MO) and eplerenone (Tocris, Ellisville, MO) were freshly prepared in ethanol or DMSO according to manufacturer instructions. Then, working solutions were diluted in RPMI 1640 culture media (Thermos Fisher scientific, NY, USA), supplemented with 5% of charcoal stripped fetal bovine serum (Thermos Fisher scientific, NY, USA), 100 I.U. /mL penicillin/ 100 µg/mL streptomycin (Thermos Fisher scientific, NY, USA), 2 mM glutamine (Thermos Fisher scientific, NY), 1 mM non-essential amino acids (Thermos Fisher scientific, NY), 1 mM pyruvate (Thermos Fisher scientific, NY, USA), 1 mM HEPES (Thermos Fisher scientific, NY, USA) and 50 mM β-mercaptoethanol (Winkler, USA). Spironolactone or eplerenone were added to cultures at a final concentration of 5µM, 30 min before aldosterone treatment. Then aldosterone was added to cultures at 10nM or 100nM. 39.
(40) concentration. All cultures included controls without any treatment or vehicle control. To verify that aldosterone stimulation was correctly achieved, it was measured the expression of sgk (28) and tnf-a (24) genes by real time PCR. The primers used were SGK-Fw 5-GAG AAG GAT GGG CCT GAA CGA T-3, SGK-Rv 5-CGG ACC CAG GTT GAT TTG TTG A-3; and, TNF-a-Fw 5-CAG GCG GTG CCT ATG TCT C-3, TNF-a-Rv 5-CGA TCA CCC CGA AGT TCA GTA G-3. The technical specifications for RT-PCR are described below.. In vivo aldosterone infusion was performed by subcutaneous implanting in mouse back pellets of D-Aldosterone which liberates 5µg of D-Aldosterone / day (Innovative Research of America, Sarasota, FL) (29). Placebo and spironolactone (500µg Spironolactone / day) pellets were also implanted as described above.. Primary culture of Dendritic Cells.. DCs were derived from bone marrow of C57BL/6, MyMRKO and LM mice, as previously described with some modifications (30). Briefly, tibias and femurs were aseptically removed from mice and bone marrow was obtained by mechanical rinse out with sterile PBS using a syringe inside of the bone. Then precursor cells were collected and centrifuged for 5 minutes at 300 g at 4°C. The pellet was resuspended in 5 ml of ACK lysis buffer, incubated during 2 minutes at room temperature followed by adding 10 mL of PBS and centrifugation for 5 minutes at 300g at 4°C. Finally, the pellet was washed twice with sterile PBS and resuspended at 1 – 1,5 x106 cells/ml in RPMI 1640 culture media (Thermos Fisher scientific, NY, USA), supplemented with 5% fetal bovine serum (Thermos Fisher scientific, NY, USA), 100 I.U. /mL penicillin/ 100 µg/mL streptomycin (Thermos Fisher scientific, NY, USA), 2 mM glutamine (Thermos Fisher scientific, NY), 1 mM non-essential. 40.
(41) amino acids (Thermos Fisher scientific, NY, USA), 1 mM pyruvate (Thermos Fisher scientific, NY, USA), 1 mM HEPES (Thermos Fisher scientific, NY), 50 mM βmercaptoethanol (Winkler, USA) and 10 ng/ml of recombinant murine GM-CSF (Peprotech, NJ, USA). Then 1 ml of cell suspension were seeded in 24 well plates (SP Life sciences, Korea) and incubated at 37°C with 5% CO2. Culture medium was replaced at 2nd and 4th day post culture, but at 4th day the culture media was supplemented with 5% of charcoal stripped fetal bovine serum (Thermos Fisher scientific, NY) to avoid steroidal hormones conditioning at the time of aldosterone pulse. At 5th day of differentiation DCs were used for all the experiments. The efficiency of DCs differentiation were determined by analyzing the expression of the specific surface marker CD11c mAb (clone HL3, BD Pharmingen) by flow cytometry. All stimuli (aldosterone or pharmacologic antagonist) was added at day 5th of culture. Cells pellet was collected to performed real time (RT)- PCR, flow cytometry, and the supernatants were collected to quantify the cytokine production by ELISA.. Expression of TLR4 by DCs.. The expression analysis of Tlr4 in DCs treated with aldosterone (in both, WT, LM and MyMRKO), will be the firstly measured by quantitative real time RT-PCR. We used the previously described procedure and primers for TLR4-Fw 5-ATG GCA TGG CTT ACA CCA CC-3 and TLR4-Rv 5-GAG GCC AAT TTT GTC TCC ACA-3 (31). Briefly, total RNA was obtained from DCs using TRIZOL reagent (Life Technologies Invitrogen, Carlsbad, CA) according to the manufacturer instructions. Then, 0,5µg of total RNA was reverse transcribed into cDNA using iScriptä Reverse Transcription Supermix for RT-qPCR (BioRad, CA, USA). The cDNA template equivalent to 100 ng of total RNA was used to real-time PCR using Fast SYBR ™ Green qPCR Master Mix (Thermos Fisher scientific, NY, USA) in a StepOnePlus™ thermocycler (Applied Biosystems). Mouse β-actin-Fw 5-AGG CAT CCT. 41.
Figure


+7






Documento similar
The discovery of Siglec-1 expression on myeloid cervical DCs (Figure 19) and the capacity of Siglec-1 + cells to capture viruses in vivo (Figure 25) help to understand how
5 Recently, we described that Chk1 and Atr1, two DNA-damage checkpoint kinases, were activated in response to the formation of b-heterodimer and that this activation resulted in
In this report, we demonstrate that mice treated with different indu- cible models of colon carcinogenesis developed a humoral immune response, which was similar to that observed
To address these issues, we first compared the effect of T3 on the TGF-β transcriptional response in Mv1Lu cells transfected with GFP fusions of the native receptor or a TRβ mutant
The skin phenotype of TR KO mice with defective response to proliferative stimuli, retarded hair growth and impaired wound healing would be compatible with the existence of a defect
In order to characterize whether the ISG15 antiviral activity observed in animal models is cell-type specific and to further characterize its implication in the immune response, we
Valuation of these benefits, in term of willingness to pay to avoid environmental deterioration, is useful to quantify the economic benefits derived from the
Government policy varies between nations and this guidance sets out the need for balanced decision-making about ways of working, and the ongoing safety considerations
