Spatial patterns of barnacle settlement in central Chile: Persistence at daily to inter annual scales relative to the spatial signature of physical variability
9
0
0
Texto completo
(2) 152. F.J. Tapia, S.A. Navarrete / Journal of Experimental Marine Biology and Ecology 392 (2010) 151–159. 2008). Inferring transport mechanisms from a single site is therefore problematic even with high-frequency observations. Increasing spatial coverage while keeping the temporal resolution is then a logical next step in the quest for a set of observations that can provide clues as to the physical processes that drive the transport and onshore delivery of larvae to intertidal habitats. Physical variability in the nearshore is largely determined by the interaction between mesoscale forcing and local topography. Thus, at local scale some physical variables such as temperature are expected to have a characteristic spatial signature (e.g. Narváez et al., 2004; Tapia et al., 2009). If the arrival of larvae to intertidal habitats is mediated by a similar set of physical forces, the deterministic component of larval settlement should bear a similar spatial signature (e.g. Wing et al., 1995). Of course, in many cases the stochastic ‘noise’ around this process might make it impossible to detect temporal or spatial trends in daily settlement. A recent study on barnacle recruitment patterns collected every 15 days at sites spread over 120 km in central Chile showed that the spatial structure of recruitment in chthamaloid barnacles (but not balanoids) was consistent with spatial–temporal variability of coastal upwelling (Lagos et al., 2005, 2007), implying that over these spatial scales and with time-integrated settlement (i.e. recruitment) data, it is possible to recover predictable spatial signals in larval arrival. Although the coarse temporal resolution of that study did not allow to infer potential mechanisms of larval transport to shore, as clearly illustrated by Shanks (2009a,b) at a single location, the spatial structure of mesoscale processes was apparent in the recruitment pattern. Here, we present a set of settlement observations on three barnacle species gathered within the same region as the bi-weekly recruitment observations by Lagos et al. (2005, 2007) in central Chile, and with an unprecedented combination of temporal resolution (daily) and spatial coverage (1–10 km). Conducted over three consecutive settlement seasons, these observations allowed us to assess the relative persistence of settlement patterns over a range of temporal scales and across species. We focus here on temporal changes in spatial patterns within and among seasons, rather than the variability of settlement time series at individual sites. We attempt to answer three questions: (1) whether spatial patterns of barnacle settlement monitored along the same stretch of coastline are persistent within a settlement season (synoptically) and among different seasons (interannually), (2) whether the observed persistence is similar among species, and (3) whether similar levels and temporal scales of persistence can be observed in the spatial patterns of environmental conditions. We reasoned that spatially-persistent patterns in the arrival of larvae (i.e. ordering of sites is maintained over time irrespective of changes in larval abundance) can be produced by: a) spatially invariant features of the environment such as site topography, which can produce persistent hydrodynamic structure (e.g. flows, turbulence) that can in turn modulate larval delivery, b) spatiallypersistent hydrodynamic processes that either directly transport larvae to shore or modulate those transport mechanisms, and c) the interaction of local topography and oceanographic processes. 2. Materials and methods 2.1. Study area and species The bay of Cartagena is a SE-facing open embayment in central Chile (33°31.2′S, 71°37.1′W) with an approximate length of 6 km. Coastal topography is abrupt, wave-exposed, and dominated by rocky substrate at both ends of the bay, and by a ∼ 5 km sandy beach at its center (Fig. 1). The region is characterized by a narrow continental shelf, and nearshore hydrography is dominated by wind-induced upwelling during austral spring and early summer (Strub et al., 1998). However, although the bay lies within a region influenced by strong. upwelling at Punta Toro and Matanzas (Wieters et al., 2003; Tapia et al., 2009), the local-scale interaction of coastal winds and shoreline topography generates small-scale structure in sea surface temperature (SST), and often promotes the retention of warm surface water near the northern tip of the bay in the form of an upwelling shadow (Kaplan et al., 2003; Narváez et al., 2004). The main sessile invertebrate species in the rocky-intertidal communities are mussels and balanomorph barnacles (Navarrete and Castilla, 1990). Among the latter, three species are numerically dominant: the chthamaloids Jehlius cirratus and Notochthamalus scabrosus that occupy the upper portion of the shore, and the balanoid Notobalanus flosculus that is mostly found in the low intertidal zone. Previous studies along central Chile (Lagos et al., 2005; Navarrete et al., 2008) have shown that these three species recruit at high rates in the bay of Cartagena, and that spatial scales of recruitment synchrony appear to be modulated by mesoscale physical variability induced by coastal upwelling (Lagos et al., 2007). It must be noted that daily settlement observations at one site near the northern tip of the bay showed that larval delivery to shore is not directly correlated with the upwelling-relaxation dynamics (Narváez et al., 2006), suggesting that the spatial upwelling signal is not a simple consequence of variation in upwelling fronts and cross-shore transport. Development of planktonic larvae in J. cirratus and N. scabrosus takes 22–29 d at 18–20 °C, and 39–41 d at 15–18 °C (Venegas et al., 2000). Although information about larval duration in N. flosculus has not been published, laboratory experiments suggest that it is similar or slightly longer than in the 2 chthamaloid species, with a duration of ca. 39 d at 18 to 20 °C (A. Olguin & S. A. Navarrete unpubl. data). 2.2. Data collection 2.2.1. Barnacle settlement Daily surveys of barnacle settlement at 5–6 sites around the bay (Fig. 1) were conducted on three consecutive seasons (2006–2008) in late spring to early summer (Table 1), spanning the months where the highest rates of barnacle settlement have been recorded for this region (Lagos et al., 2005; Navarrete et al., 2008). Since outside this time window settlement is nearly zero (Navarrete et al., 2008), data are representative of yearly barnacle settlement in the study area. Settlement was monitored using 10 × 10 cm Plexiglas plates covered with a rubbery standard surface (Safety-walk, 3 M — Menge, 2000), and attached to the substrate by means of stainless steel screws. Four plates were deployed at each site in the low intertidal zone (20 to 40 cm above Mean Lower Low Water), and spread over 10 to 30 m long transects parallel to the coastline of rocky platforms. Plates from all sites were recovered daily during the same low tide, at which time fresh plates were deployed. Depending on weather conditions, plate retrieval from all sites around the bay took between 2.5 and 3 h. As lunar cycles progressed during our surveys, it was necessary to switch collection times from evening to morning low tides. Hence, 12 h instead of 24 h deployments were made every 14 days. A separate analysis of settlement variability derived from this subset of night time deployments will be presented elsewhere. Upon recovery, collectors were placed in separate plastic bags and brought into the laboratory for immediate counting under the microscope. With the exception of 5 days in November 2008 – when over 1000 settlers of N. flosculus were found on each plate at CA, PC and EC – there was no sub-sampling of plates. All settlers on each plate were counted and individually removed. Plates were later brushed, rinsed repeatedly with freshwater, and dried in the sun for at least 24 h prior to their next deployment. 2.2.2. Environmental data Predictions of tidal height for the study area were obtained from the Servicio Hidrográfico y Oceanográfico de la Armada de Chile (www.shoa.cl). Tidal range during these periods varied between 0.7.
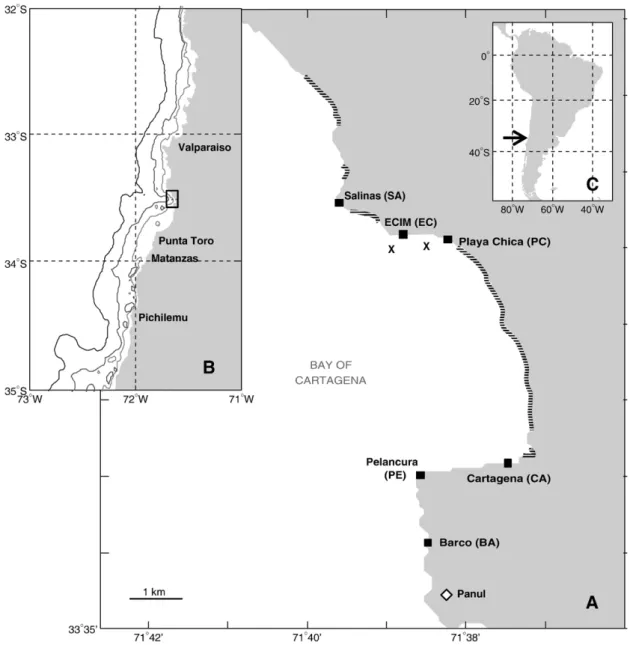
(3) F.J. Tapia, S.A. Navarrete / Journal of Experimental Marine Biology and Ecology 392 (2010) 151–159. 153. Fig. 1. (A) Geographic location of the study area, and spatial distribution of the sites where settlement was monitored over 3 consecutive seasons (squares). Hatched lines indicate areas covered by sandy beaches. Crosses near ECIM show the location of thermistor lines moored at ∼20 m depth during each season. The white diamond labeled Panul shows the location of a meteorological station where hourly wind records were gathered throughout the study. (B) Regional context for study area (rectangle) in central Chile and (C) its approximate location along the coast of South America. Contours in (B) correspond to the 1000, 200, and 100 m isobaths.. and 1.7 m (Table 1). Hourly records of wind speed and direction were gathered from a meteorological station installed at Pt. Panul (Fig. 1), at 30 m above sea level, and provided by the Servicio Meteorológico de la Armada de Chile. Sea surface temperature (SST) was recorded continuously in the shallow subtidal at 5 of the 6 sites (SA through PE, Fig. 1) using Stowaway TidBit loggers (Onset Computer Corp.), which were programmed to take synchronous point measurements at 5 min intervals. Loggers were installed at ∼ 1 m below MLLW in October. Table 1 Basic information on dates and duration of each settlement surveys, along with the range of tidal heights recorded during each period.. Number of sites Sampling dates Number of observations Min/max tidal range (m). 2006. 2007. 2008. 5 19 Oct–6 Dec 48 0.77–1.55. 6 20 Oct–8 Jan 71 0.69–1.62. 6 22 Oct–21 Dec 53 0.68–1.68. 2006, and have been serviced and/or replaced since. Similar loggers were attached to lines moored near ECIM (Fig. 1), at depths of 16 m (East mooring) and 18 m (West mooring), and used to monitor innershelf variability in thermal stratification. Temperature at 6 different depths (1, 4, 7, 10, 13, and 16 m above the bottom) was recorded every 5 min for the duration of each settlement survey. 2.3. Data processing and analysis 2.3.1. Settlement data Series of daily settlement rates for each species and site were produced by averaging settler counts across plates, and dividing by the number of hours that plates were deployed at each site. Although cyprids and spat were enumerated separately for each species, and a greater percentage of cyprids was consistently found for N. flosculus (N60%) versus N. scabrosus (40–60%) and J. cirratus (b40%), total numbers of settlers are used here to calculate settlement rates. An analysis of temporal and spatial variabilities in the relative numbers of cyprid versus spat will be presented elsewhere..
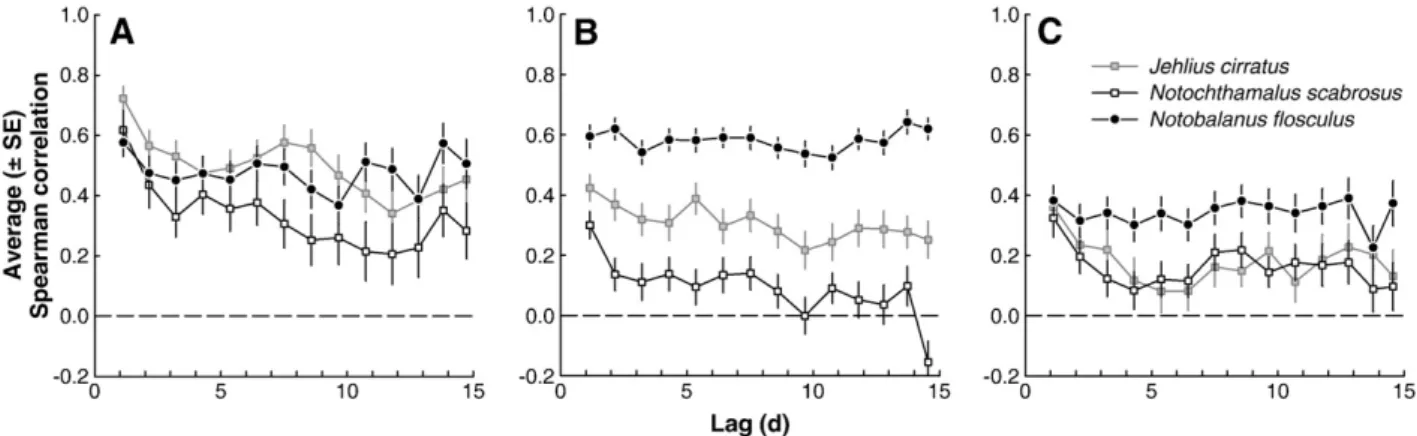
(4) 154. F.J. Tapia, S.A. Navarrete / Journal of Experimental Marine Biology and Ecology 392 (2010) 151–159. Among-site coherence in settlement was assessed for each species through Kendall correlation coefficients computed for each pair of settlement time series within each season. Average Kendall correlations were then computed across all pairs of sites. Given that each settlement survey encompassed at least 1 full lunar cycle (3, 5 and 4 fortnights in 2006, 2007 and 2008), the association between phase of the lunar cycle and mean settlement rates was examined for each species. Across-bay mean settlement rates were computed for each one of the 172 sets of observations, and then classified into one of 29 categories that corresponded approximately to days in the lunar cycle, with day 1 being the day after new moon (Pineda, 1995). Thus, for each lunar day this procedure yielded between 4 and 7 observations that were averaged to produce a mean lunar month for the settlement of each species. The analysis of pattern persistence was based on time series of ranks, rather than settlement rates, assigned to sites on each day with settlement observations. In this way, emphasis was placed on the order of sites in terms of settlement (from highest to lowest), and the correlation coefficient was not influenced by a few unusually high settlement events over a given season. Ranks from 1 up to 6 (or up to 5 in 2006) were assigned to the lowest and highest settlement rates, respectively. Days with missing observations for 1 or more sites (2 in 2006 and 1 in 2008), or with zero settlement across all sites in the bay (6, 8, and 15 for J. cirratus, N. scabrosus, and N. flosculus), were not included in this analysis. Persistence is defined here as the consistency between ranks assigned to each site at different times during a given season. If the spatial pattern of settlement is persistent, a site will be ranked similarly throughout the season regardless of bay-scale changes in mean settlement rates. Finally, to examine temporal scales at which settlement patterns were persistent, rank correlations were calculated for all pairs of around-the-bay settlement observations gathered with increasing time lags up to 15 days. In this way, a Spearman rank correlation near 1 at time lag 1 meant that the ranking of sites remains unaltered from one day to the next (see Navarrete et al., 2008). 2.3.2. Environmental data Hourly series of eastward and northward wind velocities were produced for each season from the original speed and direction records, and used to determine the main axis of wind variability for each season. The magnitude and angle of this axis were obtained from the largest eigenvalue and the corresponding eigenvector of the eastward versus northward covariance matrix. Angles calculated for the main axis of variability were corrected for magnetic declination at the study area, which fluctuated between 3.95 and 4.1 degrees from true north (°TN) between October 2006 and December 2008 (http:// www.ngdc.noaa.gov). Temperature records gathered at the sites since October 2006 were quality controlled to eliminate out-of-the-water measurements taken during logger servicing (up to 15 min) and averaged to produce hourly SST series, which were low-pass filtered using a Lanczos cosine filter with a cutoff period of 40 h to remove temperature oscillations induced by semi-diurnal tides and diurnal heating. Daily SST series were then produced by averaging over the filtered hourly series. In addition, to examine spatial patterns in high-frequency SST variability, periodic fluctuations with cycles that ranged between 6 h and 365 days (i.e. from semi-diurnal tides to the annual cycle in temperature) were removed from raw hourly series. Harmonic analyses were performed on hourly series of SST gathered over 954 days (October 2006 through May 2009) using t_tide for MATLAB (Pawlowicz et al., 2002). Estimated harmonics accounted for 63–74% of total variability in the raw hourly records, and were subtracted from the full series to produce what we henceforth will refer to as ‘high-frequency SST’ (i.e. fluctuations b6 h). Temperature records gathered at the two moorings near ECIM (Fig. 1) were processed in a similar way. Series of hourly means from the top and bottom loggers. were low-pass filtered, and used to calculate daily surface-to-bottom gradients in temperature as a proxy for nearshore stratification (see Vargas et al., 2004). 3. Results Settlement surveys in 2006, 2007 and 2008 spanned 1.5, 2.5, and 2 lunar cycles, respectively (Fig. 2), with equal numbers (3, 5, and 4, respectively) of spring and neap tides in each season. For all three species, and within each site, inter-annual differences were observed both in the rates and timing of larval settlement (Table 2, Fig. 2). Although within each species there were large among-site differences in settlement rates (Fig. 2), spatially coherent settlement pulses were observed at different times during all three seasons. For instance, in 2006 there were coherent pulses of N. scabrosus settlement at the southern end of the bay (CA and PE, Fig. 2D,E), whereas settlement of N. flosculus was coherent across the bay (i.e. SA through CA) in 2007 (Fig. 2F–I) and 2008 (Fig. 2L–O). Overall, among-site coherence in the temporal patterns of settlement increased for all three species from 2006 to 2008, as shown by mean Kendall correlations computed for daily settlement rates (Table 3). At the scale of the entire bay, there was an apparent association between the timing of settlement of N. flosculus and the phase of the lunar cycle (Fig. 3C), whereas no such association was found for chthamaloids (Fig. 3A,B). Despite inter-annual changes in settlement (Fig. 2), around-thebay distributions of mean ranks for each species (Fig. 4) indicated that spatial patterns of settlement were consistent from year to year. For both chthamaloids, highest and lowest settlement ranks were always found at the southern and northern tips of the bay (PE versus EC or SA), respectively (Fig. 4A,B). A contrasting but still persistent pattern was observed for N. flosculus (Fig. 4C), with the highest settlement ranks found at “interior” sites (i.e. CA, PC, and EC) and lower ranks at the “exterior” sites (i.e. PE, SA). Spearman correlations computed for settlement rates over time lags of 1 to 15 days (Fig. 5) unveiled striking inter-specific and interannual differences in the degree and temporal scale of pattern persistence. Settlement patterns of N. flosculus were spatially structured and persistent over scales of at least 10 days in 2006 and 2007, and slightly less structured in 2008 (Fig. 5A,C). There was no evidence of “drift” or loss of the spatial structure with time in this species, which means that the “good” and “bad” larval delivery sites for this species remained roughly the same day after day within a given season. Spatial structure in settlement of N. flosculus was particularly strong and persistent in 2007, as indicated by the small variability associated to rank correlations over the entire range of time lags (see error bars in Fig. 5B). Persistence of settlement patterns in chthamaloids, on the other hand, was high over scales of 2–5 days in 2006 – though more so for J. cirratus than for N. scabrosus – and substantially lower in 2007 and 2008, when site ranks fluctuated through the season (Fig. 5A–C). This inter-annual drop in mean persistence was gradual from 2006 to 2008 for J. cirratus and more abrupt from 2006 to 2007 for N. scabrosus (Fig. 5B). A consistent decline in persistence as a function of time lag observed for both chthamaloids in 2008 (Fig. 5C) indicated a loss of spatial structure in larval arrival over scales of 2–5 days. Inter-annual differences in mean persistence, or lack thereof, were not correlated with changes in the mean rates of settlement (p N 0.60 for J. cirratus, p N 0.16 for N. scabrosus, and p N 0.34 for N. flosculus). Spatial patterns of synoptic-scale variability in sea surface temperature (SST) around the bay were persistent for all three years and over the entire range of time lags examined, with a slight increase in mean persistence from 2006 to 2007 and 2008 (Fig. 6A). In contrast, patterns of high-frequency SST variability (i.e. hours), showed no persistence among sites (Fig. 6B). A common feature of the high-frequency SST patterns for all three years was a sharp drop in persistence after only 12 h (Fig. 6B), after which increases in.
(5) F.J. Tapia, S.A. Navarrete / Journal of Experimental Marine Biology and Ecology 392 (2010) 151–159. 155. Fig. 2. Daily rates of larval settlement for 3 species of intertidal barnacles recorded around the bay of Cartagena, central Chile, over 3 consecutive seasons. Left y-axes correspond to the chthamaloids Jehlius cirratus (thick grey line) and Notochthamalus scabrosus (thin black line), whereas the right y axes correspond to the balanoid Notobalanus flosculus (thick black line). Note the changes in y-axis scales from year to year. Dates corresponding to the new and full moon (black and white circles, respectively) are shown at the top.. persistence were observed at regular intervals of 1 day lags for 2007 and 2008 (Fig. 6B). This suggests that there is a daily process that tends to moderately structure the spatial pattern of high-frequency SST variability around the bay. A shift in coastal wind patterns was observed between 2006 and 2008. Although there was no substantial change in the average speed of coastal winds (Fig. 7A, grey bars), the ratio of alongshore to crossshore winds changed between seasons (Fig. 7A, squares and dashed line), owing mostly to a weakening of alongshore winds from 2006 to 2007, and to a subsequent weakening of cross-shore winds in 2008 (data not shown). This change in the anisotropy of coastal winds was accompanied by a 10° clockwise rotation of the main axis of wind variability (Fig. 7A, circles and solid line). Hence, winds became increasingly more perpendicular to the shoreline (Fig. 1). Concurrent. with this change in the regime of wind forcing, mean daily temperatures measured in the shallow subtidal at 5 sites around the bay increased by 1.4 °C between 2006 and 2008 (Fig. 7B). A similar increase was detected in the mean surface-to-bottom temperature gradient measured at two moorings near ECIM (Fig. 7C). 4. Discussion Our results show that despite order-of-magnitude changes in barnacle settlement rates, the spatial pattern of daily larval arrival to intertidal habitats may persist over synoptic scales and, in some cases, from year to year. Such was the case of the balanoid N. flosculus, whose spatial patterns of settlement were highly persistent in 2006 and 2007, and slightly less so in 2008. In contrast, settlement of. Table 2 Mean settlement rates (individuals per plate/h) recorded for each species on each of the seasons and sites. Standard deviations are shown in parentheses. Site. SA EC PC CA PE BA. Jehlius cirratus. Notobalanus flosculus. Notochthamalus scabrosus. 2006. 2007. 2008. 2006. 2007. 2008. 2006. 2007. 2008. 0.134 (0.148) 0.052 (0.061) 0.278 (0.358) 0.219 (0.234) 0.858 (0.838) – –. 0.107 (0.219) 0.105 (0.285) 0.201 (0.440) 0.249 (0.662) 0.336 (0.627) 0.275 (0.619). 0.229 (0.335) 0.174 (0.247) 0.852 (1.556) 0.569 (0.747) 1.119 (1.963) 1.210 (2.301). 0.615 (0.688) 0.328 (0.412) 1.069 (1.497) 0.678 (0.829) 1.399 (1.191) – –. 0.211 (0.319) 0.276 (0.820) 0.244 (0.327) 0.232 (0.365) 0.230 (0.293) 0.341 (0.396). 0.128 (0.251) 0.114 (0.172) 0.341 (0.635) 0.146 (0.187) 0.304 (0.729) 0.498 (1.552). 0.009 (0.018) 0.056 (0.108) 0.067 (0.162) 0.204 (0.265) 0.011 (0.015) – –. 0.164 (0.504) 0.468 (1.087) 0.153 (0.517) 0.463 (0.681) 0.023 (0.041) 0.014 (0.040). 1.132 (3.456) 1.987 (5.032) 1.609 (5.649) 3.930 (12.491) 0.437 (1.224) 0.184 (0.437).
(6) 156. F.J. Tapia, S.A. Navarrete / Journal of Experimental Marine Biology and Ecology 392 (2010) 151–159. Table 3 Mean Kendall correlations calculated for all pairs of daily settlement series across the bay. Season. J. cirratus. N. scabrosus. N. flosculus. 2006 2007 2008. 0.211 0.562 0.625. 0.293 0.379 0.656. 0.310 0.416 0.716. chthamaloids was spatially structured and persistent during the first year, but became less persistent in the following years. These differences in the persistence of settlement patterns among species may stem from differences in the way time-invariant factors modulating larval delivery around the bay of Cartagena (e.g. local bathymetry, shoreline orientation, site topography) affect larvae of these species. A similar role has been ascribed to local topography by Rilov et al. (2008) to explain pattern persistence in the settlement of intertidal mussels from Oregon and New Zealand. Below we discuss potentially driving factors behind the spatial patterns of barnacle settlement in central Chile, and highlight the importance of conducting settlement observations over multiple sites and, ideally, over different years. Bi-weekly observations conducted over 6 months in a previous study that spanned 120 km and encompassed the bay of Cartagena (Lagos et al., 2005, 2007) had indicated that spatial recruitment patterns in J. cirratus and N. scabrosus resemble those of mesoscale SST variability induced by coastal upwelling (Lagos et al., 2007), whereas greater small-scale variability in the recruitment of N. flosculus appeared to result from local factors such as site topography and larval behavior (Lagos et al., 2005). Our observations, gathered over a much smaller region but with higher temporal resolution, agree well with the previously detected differences in settlement patterns between chthamaloid and balanoid species. The persistence of N. flosculus settlement patterns over each season, and the slight interannual changes we observed, may result from the relatively invariant interaction of site-scale topographic features and processes that force synoptic-scale SST variability (see below), or from a greater importance of small-scale features over synoptic-scale variability. Conversely, the lack of such persistence in spatial patterns of chthamaloid settlement in 2 out of 3 seasons suggests that invariant features such as local topography are less important for these species, at least over the coastal region encompassed by this study. We then asked whether there were physical processes that exhibited changes in their spatial structure that resembled variation in settlement patterns of chthamaloids. Such process or processes could underlie spatial structure in barnacle settlement onshore and could then be used to predict spatial patterns of larval arrival. This would be an. Fig. 4. Inter-annual variability of spatial structure in the settlement of 3 species of intertidal barnacles: (A) Jehlius cirratus, (B) Notochthamalus scabrosus, and (C) Notobalanus flosculus. Ranks were assigned to daily settlement rates and averaged over each season for each species. Only the 5 sites that were monitored in all three years were included in this analysis. See Fig. 1 for site name codes.. important finding for intertidal ecologists since a spatial pattern of larval arrival that persists over temporal scales of an entire settlement season, or even from year to year, can have important implications for spatial variation in population dynamics. It must be noted that a lack of correspondence in the spatial structure of physical variables and settlement does not necessarily prove that a given physical process is not involved in larval delivery to shore; it only suggests that such process is not likely to produce persistent spatial structure in settlement within the study region. This would be case when the. Fig. 3. Lunar-month distribution of bay-scale rates of settlement for (A) Jehlius cirratus, (B) Notochthamalus scabrosus, and (C) Notobalanus flosculus. All 172 sets of daily settlement observations were used to compute average rates across sites, and then grouped according to their relative position in the lunar cycle (day 1 corresponds to the day after new moon)..
(7) F.J. Tapia, S.A. Navarrete / Journal of Experimental Marine Biology and Ecology 392 (2010) 151–159. 157. Fig. 5. Change in the persistence of spatial patterns in barnacle settlement as a function of time lag between observations in (A) 2006, (B) 2007, and (C) 2008. Correlations were computed on sets of ranks assigned each day to settlement observations made across the bay. See Materials and methods for more details.. transport process and/or larval availability are spatially stochastic over the study region. Conversely, physical processes can leave characteristic spatial signals on settlement or recruitment without being directly involved in larval transport to shore, for instance by modulating other transport processes or affecting advection/diffusion in the larval pool (see Largier, 2003). Our analysis of pattern persistence in synoptic-scale and highfrequency SST variability was aimed at capturing spatial structure that could be used as a proxy for processes inducing larval transport around the bay. Although the transport of water and suspended particles to and from the shoreline could be inferred more directly from long-term records of current velocities at multiple points and depths, obtaining such records is logistically challenging and often financially unfeasible. Alternatively, continuous measurements of temperature variability are a relatively inexpensive way to obtain high-resolution records that, given certain assumptions as to solar radiation and heat fluxes, may provide a good proxy for water transport in the coastal ocean (e.g. Pineda, 1991; Leichter et al., 1996; Pineda, 1999; Kaplan et al., 2003; Narváez et al., 2006; Shanks, 2006). Typically, synoptic-scale SST variability in this region has been associated to cycles of upwelling/relaxation (e.g. Narváez et al., 2004, 2006; Lagos et al., 2007), whereas high-frequency SST variability has been often monitored at single sites and used as a proxy for the occurrence of internal motions (e.g. Vargas et al., 2004; Ladah et al., 2005). Our results show that high-frequency oscillations in SST do not have a persistent spatial structure around the bay of Cartagena in any of the 3 settlement seasons, and it is therefore unlikely that they could by themselves establish spatially-persistent settlement patterns as those observed for all species in year 2006 and for N. flosculus in 2006 and 2007.. In contrast to high-frequency oscillations, synoptic-scale variability in SST did have a remarkably persistent spatial structure among sites, with only slight variation among years. It is not completely understood what produces such a persistent SST pattern among sites separated by only a few kilometers, and why it would change from year to year, but recent observational and modeling studies (Aiken et al., 2008; Fewings et al., 2008; Tapia et al., 2009) have documented the dynamical interaction between coastal topography, bathymetry, and changes in the orientation of prevailing winds, as well as the interaction between mesoscale winds and upwelling plumes generated upstream (see Wieters et al., 2003). Whichever the physical mechanisms, the spatially-persistent structure of synoptic-scale variability might underlie spatial structure in barnacle settlement. This would have certainly been our final conclusion had we only collected data in 2006, when there was good agreement between synoptic SST structure and spatial persistence of all three barnacle species. But on successive years, a clear loss of spatial structure in chthamalid settlement had no parallel in SST spatial structure, which remained highly persistent. Clearly, and unfortunately from the point of view of intertidal ecologists, synoptic-scale SST is not sufficient to explain spatial settlement patterns, at least not over the scales examined in this study. More complex interactions must therefore be considered if we are to understand and predict spatial variation in settlement. Coastal winds are a key source of forcing on the surface ocean that, upon interaction with shoreline topography, may induce persistent hydrodynamic patterns in the inner shelf (Graham and Largier, 1997; Wieters et al., 2003; Nielsen and Navarrete, 2004; Broitman et al., 2005; Lagos et al., 2005; Tapia et al., 2009). In addition to this synoptic-scale forcing of nearshore hydrodynamics, diurnal onshore. Fig. 6. Persistence of spatial patterns in sea surface temperature (SST) measured at 5 sites around the bay of Cartagena over 3 consecutive seasons. Calculations of serial Spearman correlations were performed using time series of (A) daily means of low-pass filtered SST, and (B) hourly means of the high-pass filtered SST series obtained after removal of harmonics with periods ranging from 6 h to 365 d (see Materials and methods)..
(8) 158. F.J. Tapia, S.A. Navarrete / Journal of Experimental Marine Biology and Ecology 392 (2010) 151–159. Fig. 7. Inter-annual differences in (A) the intensity (grey bars) and geometry (symbols and lines) of coastal winds. Squares correspond to the ratio of alongshore versus cross-shore winds (first right axis), and circles show the angle of maximum wind variability θv (second right axis). (B) Mean daily surface temperature measured at 5 sites around the bay, and (C) mean surface-to-bottom gradients in temperature measured by thermistors moored at depths of 18 m (West) and 16 m (East) near ECIM (see Fig. 1, crosses). Mean temperatures were calculated from daily series produced for each record (see Materials and methods).. winds associated with the summertime sea breeze promote shorterterm variability in the cross-shelf transport of surface material, including the larvae of intertidal barnacles in the area of our study (Vargas et al., 2006), nearshore accumulation of warm surface waters (Kaplan et al., 2003; Tapia et al., 2004), and the onshore propagation of internal features such as tidal bores (Pineda and López, 2002; Shanks, 2006). Changes in the orientation of prevailing winds relative to the shoreline and local bathymetry, as well as in the relative intensity of synoptic-scale versus diurnal wind forcing, can induce drastic changes in the stratification and transport of inner-shelf waters. For example, whereas synoptic equatorward winds force the retention of warm surface waters at the northern tip of Monterey Bay (Graham and Largier, 1997), summertime diurnal sea breezes blowing over the same area may induce upwelling at local scales (Woodson et al., 2007). Thus, changes in the geometry of wind forcing at a given site may alter the balance of meso- and small-scale forcing of the surface ocean, and produce drastically different patterns of onshore transport, accumulation, and retention of surface water. Consequently, patterns of larval delivery to intertidal habitats may change drastically as well. Over the three seasons spanned by our study, we detected a sustained clockwise rotation of the main axis of wind variability, which went from laying at ca. 40° east of true north (TN) in 2006 to angles close to 50° TN in 2007 and 2008. This increase in the angle of prevailing winds relative to the bay's main axis (340° TN) may have driven the slight increase in persistence of SST patterns observed between 2006 and the other two seasons (Fig. 6). Persistence of patterns in high-frequency SST variability increased at regular lag increments of ca. 1-day in 2007 (Fig. 6B), which might be an indication of the importance of sea-breezes versus mesoscale forcing in structuring high-frequency SST variability around the bay (Kaplan et al., 2003). In all likelihood, inter-annual changes in patterns of wind variability modified the balance of synoptic (i.e. coastal upwelling) and diurnal (i.e. sea breeze) forcing on the surface ocean, determining the observed changes in SST patterns around the bay. We speculate that this chain of events, i.e. change in wind patterns potentially followed by a change in patterns of onshore transport and retention, which may be perceived as a change in SST structure around the bay,. may be responsible for the inter-annual changes in persistence of settlement patterns observed for chthamaloid barnacles. The larvae of balanoids, on the other hand, may in addition be responding to local features that induce spatial structure, such as site topography or the abundance of conspecifics, or perhaps be retained much closer to the shoreline during their entire planktonic development. Whenever observations of larval settlement and environmental variability are used to make inferences regarding physical processes that mediate larval transport, two main caveats must be taken into consideration. First, the temporal resolution at which settlement is monitored sets a limit to the signals that can be detected, and may obscure the connection between certain processes and the arrival of larvae to intertidal habitats (Shanks, 2009a,b). Second, patterns of larval settlement do not necessarily reflect patterns of larval supply to a site (Pineda, 2000), as biological interactions and substrate availability may decouple supply and settlement. Conversely, the occurrence of a physical process that is critical for the cross-shelf transport of larvae will not necessarily translate into a settlement signal, since competent larvae may not be available in the plankton. It is thus desirable to complement observations such as those presented here with the monitoring of nearshore availability of planktonic larvae at appropriate spatial–temporal scales (e.g. Rilov et al., 2008). In this contribution we have only focused on spatial patterns of settlement, leaving a detailed analysis of transport processes involved in larval delivery to our study sites for a later contribution. Studying patterns of temporal variability in larval supply in concert with the spatial patterns of settlement and their scales of persistence is necessary to identify larval transport processes and their relative importance, and to achieve a more mechanistic understanding of planktonic–benthic coupling in the coastal ocean. Acknowledgments This study was made possible by the generous support and encouragement of many people at the Estación Costera de Investigaciones Marinas, Pontificia Universidad Católica de Chile, and by the financial support of FONDECYT-Chile through a post-doctoral grant (3070014) awarded to FT in 2007. Additional funding was provided.
(9) F.J. Tapia, S.A. Navarrete / Journal of Experimental Marine Biology and Ecology 392 (2010) 151–159. by the Andrew W. Mellon Foundation and FONDAP–FONDECYT 15001-001 to SAN. Logistic support and advice from Fredy Veliz were essential to setup and launch this study. Randy Finke and Manuel Castillo provided critical support with field measurements, data collection, and processing. Jenna Shinen provided insightful discussion and help in the field and laboratory. Special thanks are due to J. Escobar, M. Parragué, G. Maguelonne, F. Villenas, E. Ramos, M. Aguilera, J. Pantoja, S. Dudas, J. Tyburczy, R. Millston-Clemens, J. Schaefers, and G. Rilov for the countless hours they spent swapping settlement plates in the field, and counting larvae in the laboratory. [SS] References Aiken, C.M., Castillo, M.I., Navarrete, S.A., 2008. A simulation of the Chilean Coastal Current and associated topographic upwelling near Valparaíso, Chile. Continental Shelf Research 28, 2371–2381. Broitman, B.R., Blanchette, C.A., Gaines, S.D., 2005. Recruitment of intertidal invertebrates and oceanographic variability at Santa Cruz Island, California. Limnology and Oceanography 50, 1473–1479. Connolly, S.R., Roughgarden, J., 1998. A latitudinal gradient in northeast Pacific intertidal community structure: evidence for an oceanographically based synthesis of marine community theory. American Naturalist 151, 311–326. Fewings, M., Lentz, S.J., Fredericks, J., 2008. Observations of cross-shelf flow driven by cross-shelf winds on the inner continental shelf. Journal of Physical Oceanography 38, 2358–2378. Graham, W.M., Largier, J.L., 1997. Upwelling shadows as nearshore retention sites: the example of northern Monterey Bay. Continental Shelf Research 17, 509–532. Hatton, H., 1938. Essais de bionomie explicative sur quelques espèces intercotidales d'algues et d'animaux. Annales de l'Institut Océanographique 17, 241–348. Kaplan, D.M., Largier, J.L., Navarrete, S., Guiñez, R., Castilla, J.C., 2003. Large diurnal temperature fluctuations in the nearshore water column. Estuarine, Coastal and Shelf Science 57, 385–398. Ladah, L.B., Tapia, F.J., Pineda, J., López, M., 2005. Spatially heterogeneous, synchronous settlement of Chthamalus spp. larvae in Northern Baja California. Marine Ecology Progress Series 302, 177–185. Lagos, N.A., Navarrete, S.A., Veliz, F., Masuero, A., Castilla, J.C., 2005. Meso-scale spatial variation in settlement and recruitment of intertidal barnacles along the coast of central Chile. Marine Ecology Progress Series 290, 165–178. Lagos, N.A., Tapia, F.J., Navarrete, S.A., Castilla, J.C., 2007. Spatial synchrony in the recruitment of intertidal invertebrates along the coast of central Chile. Marine Ecology Progress Series 350, 29–39. Largier, J.L., 2003. Considerations in estimating larval dispersal distances from oceanographic data. Ecological Applications 13, S71–S89. Leichter, J.J., Wing, S.R., Miller, S.L., Denny, M.W., 1996. Pulsed delivery of subthermocline water to Conch Reef (Florida Keys) by internal tidal bores. Limnology and Oceanography 41, 1490–1501. Menge, B.A., 2000. Recruitment vs. postrecruitment processes as determinants of barnacle population abundance. Ecological Monographs 70, 265–288. Mitarai, S., Siegel, D.A., Winters, K.B., 2008. A numerical study of stochastic larval settlement in the California Current system. Journal of Marine Systems 69, 295–309. Narváez, D., Poulin, E., Leiva, G., Hernandez, E., Castilla, J.C., Navarrete, S.A., 2004. Seasonal and spatial variation of nearshore hydrographic conditions in central Chile. Continental Shelf Research 24, 279–292. Narváez, D.A., Navarrete, S.A., Largier, J., Vargas, C.A., 2006. Onshore advection of warm water, larval invertebrate settlement, and relaxation of upwelling off central Chile. Marine Ecology Progress Series 309, 159–173. Navarrete, S.A., Castilla, J.C., 1990. Barnacle walls as mediators of intertidal mussel recruitment: effects of patch size on the utilization of space. Marine Ecology Progress Series 68, 113–119.. 159. Navarrete, S.A., Broitman, B.R., Menge, B., 2008. Interhemispheric comparison of recruitment to intertidal communities: pattern persistence and scales of variation. Ecology 89, 1308–1322. Nielsen, K.G., Navarrete, S.A., 2004. Mesoscale regulation comes from the bottom-up: intertidal interactions between consumers and upwelling. Ecology Letters 7, 31–41. Pawlowicz, R., Beardsley, B., Lentz, S., 2002. Classical tidal harmonic analysis including error estimates in MATLAB using T_TIDE. Computers & Geosciences 28, 929–937. Pineda, J., 1991. Predictable upwelling and the shoreward transport of planktonic larvae by internal tidal bores. Science 253, 548–551. Pineda, J., 1994. Spatial and temporal patterns in barnacle settlement rate along a southern California rocky shore. Marine Ecology Progress Series 107, 125–138. Pineda, J., 1995. An internal tidal bore regime at nearshore stations along western U.S.A.: predictable upwelling within the lunar cycle. Continental Shelf Research 15, 1023–1041. Pineda, J., 1999. Circulation and larval distribution in internal tidal bore warm fronts. Limnology and Oceanography 44, 1400–1414. Pineda, J., 2000. Linking larval settlement to larval transport: assumptions, potentials, and pitfalls. In: Färber-Lorda, J. (Ed.), Oceanography of the Eastern Pacific, 1. CICESE, Ensenada, pp. 84–105. Pineda, J., López, M., 2002. Temperature, stratification and barnacle larval settlement in two Californian sites. Continental Shelf Research 22, 1183–1198. Rilov, G., Dudas, S.E., Menge, B.A., Grantham, B.A., Lubchenco, J., Schiel, D.R., 2008. The surf zone: a semi-permeable barrier to onshore recruitment of invertebrate larvae? Journal of Experimental Marine Biology and Ecology 361, 59–74. Shanks, A.L., 2006. Mechanisms of cross-shelf transport of crab megalopae inferred from a time series of daily abundance. Marine Biology 148, 1383–1398. Shanks, A.L., 2009a. Barnacle settlement versus recruitment as indicators of larval delivery. I. Effects of post-settlement mortality and recruit density. Marine Ecology Progress Series 385, 205–216. Shanks, A.L., 2009b. Barnacle settlement versus recruitment as indicators of larval delivery. II. Time-series analysis and hypothesized delivery mechanisms. Marine Ecology Progress Series 385, 217–226. Strub, P.T., Mesías, J.M., Montecino, V., Rutlant, J., Salinas, S., 1998. Coastal ocean circulation off western South America. In: Robinson, A.R., Brink, K.H. (Eds.), The Sea, 11. John Wiley & Sons, New York, pp. 273–313. Tapia, F., Navarrete, S.A., Castillo, M., Menge, B.A., Castilla, J.C., Largier, J., Wieters, E.A., Broitman, B.L., 2009. Thermal indices of upwelling effects on inner-shelf habitats. Progress in Oceanography 83, 278–287. Tapia, F.J., Pineda, J., Ocampo-Torres, F.J., Fuchs, H.L., Parnell, P.E., Montero, P., Ramos, S., 2004. High-frequency observations of wind-forced onshore transport at a coastal site in Baja California. Continental Shelf Research 24, 1573–1585. Vargas, C.A., Narváez, D.A., Piñones, A., Venegas, R.M., Navarrete, S.A., 2004. Internal tidal bore warm fronts and settlement of invertebrates in central Chile. Estuarine Coastal and Shelf Science 61, 603–612. Vargas, C.A., Narváez, D.A., Piñones, A., Navarrete, S.A., Lagos, N.A., 2006. River plume dynamics influence transport of barnacle larvae in the inner shelf off central Chile. Journal of the Marine Biological Association of the United Kingdom 86, 1057–1065. Venegas, R.M., Ortiz, V., Olguin, A., Navarrete, S.A., 2000. Larval development of the intertidal barnacles Jehlius cirratus and Notochthamalus scabrosus (Cirripedia: Chthamalidae) under laboratory conditions. Journal of Crustacean Biology 20, 495–504. Wieters, E.A., Kaplan, D.M., Navarrete, S.A., Sotomayor, A., Largier, J., Nielsen, K.J., Veliz, F., 2003. Alongshore and temporal variability in chlorophyll a concentration in Chilean nearshore waters. Marine Ecology Progress Series 249, 93–105. Wing, S.R., Botsford, L.W., Largier, J.L., Morgan, L.E., 1995. Spatial structure of relaxation events and crab settlement in the northern California upwelling system. Marine Ecology Progress Series 128, 199–211. Woodson, C.B., Eerkes-Medrano, D.I., Flores-Morales, A., Foley, M.M., Henkel, S.K., Hessing-Lewis, M., Jacinto, D., Needles, L., Nishizaki, M.T., O'Leary, J., Ostrander, C.E., Pespeni, M., Schwager, K.B., Tyburczy, J.A., Weersing, K.A., Kirincich, A.R., Barth, J.A., McManus, M.A., Washburn, L., 2007. Local diurnal upwelling driven by sea breezes in northern Monterey Bay. Continental Shelf Research 27, 2289–2302..
(10)
Figure

Documento similar