Recent rodent models for Alzheimer's disease: clinical implications and basic research
23
0
0
Texto completo
(2) 174. derived from amyloid precursor protein (APP) (Masters et al. 1985). Essentially, APP is cleaved by b-secretase via the endosomal–lysosomal pathway to generate the amino terminus of Ab. The fragment is further processed by c-secretase at positions 40, 42, and 43 to generate the Ab peptide (Vassar et al. 1999). Several different N-terminally truncated Ab have been detected in post-mortem AD brain tissue (Hartig et al. 2010; Saido et al. 1995, 1996). Indeed, AbpE3 (N-terminally truncated Ab starting with pryoglutamate) has been found to represent a dominant fraction of Ab in the AD brain (Saido et al. 1995, 1996). Ab1–42 is the most neurotoxic of the amyloid species, due in part to its tendency to form insoluble fibrils (Edbauer et al. 2003). However, a growing number of studies have recently shown that soluble amyloid oligomers may represent the primary pathological structure rather than mature amyloid fibrils (Glabe 2006; Hardy and Selkoe 2002; Harper et al. 1997). These oligomers can permeabilise cellular membranes and lipid bilayers which can initiate a cascade of downstream effects such as calcium dyshomeostasis (Mina et al. 2004) and free radical damage (Schubert et al. 1995), mitochondrial dysfunction (Hashimoto et al. 2003), and impaired autophagy (Liberski et al. 2004), leading to cellular loss observed in AD (Glabe 2006). A majority of cases of AD occur over the age of 50 years, with a progressive increase in incidence with increasing age (Ferri et al. 2006). Although most cases are sporadic, it has been estimated that up to 10% of patients demonstrate a family history of dementia (Ferri et al. 2006). While the exact aetiology remains unknown, several genes, proteins, toxins and metabolic abnormalities have been linked to the development of AD (Suh and Checler 2002). Recently, the identification of presenilin 1 (PS1), presenilin 2 (PS2) and the ApoE4 allele has significantly modified our understanding of the disease (Suh and Checler 2002). In particular, presenilins form an integral part of the c-secretase pathway, and mutations in these proteins are associated with increased ratios of the Ab1–42 versus Ab1–40 (Golde and Younkin 2001). While neuronal dysfunction and cell death are the most obvious substrates for dementia, it is clear that a variety of complex mechanisms may contribute to the clinical signs of progressively decline in cognition. With the aging of the world population, AD has become an increasing and major public health problem in the industrialised world. New approaches aimed at prevention, delay and therapy are necessary to lessen the burden. In this respect, animal models have become vital tools for modelling disease states to test hypotheses leading to the development of potential treatment therapies.. 123. N. Braidy et al.. Non-transgenic mammalian animal models for AD Primates Human neurodegenerative diseases are complex multifactorial disorders with both neuropathological and psychiatric alterations. Non-human primates (i.e. various species of monkeys) are genetically and physiologically the closest animals to humans. The non-human primate brain shares similar structural and biochemical changes that occur parallel to several changes that have been observed in the aged human brain (Bons et al. 2006). For example, the ageassociated neurodegeneration reported in primates is associated with brain atrophy, abundant amyloid plaques, tau pathology and a loss of cholinergic neurons (Voytko 1998). Amyloid deposition has been investigated in several mammalian species (Walker 1991, 1997). In monkeys, several groups have shown that the amyloid precursor protein (APP) shares the same cellular localisation in humans (Martin et al. 1991). Moreover, the 695-Ab peptide in macaque monkey is similar to that of the human (Podlisny et al. 1991). The frequency of amyloid pathology in these primate models represents interesting animal models to study human neurodegenerative diseases (Bons et al. 1991, 1994; Gearing et al. 1994, 1996, 1997). They are likely to be the most successful models to identify markers for diagnosis and for the development of safe and efficient treatment for human brain disorders (Chan 2004). Electronmicroscopic analyses of senile plaques obtained from aged macaque monkeys are analogous to those observed in AD, with a similar dense amyloid core, but lack the characteristic twisted tubules present in monkeys (Wisniewski et al. 1973). While similar age-related change abnormalities have been reported between aged monkeys and in patients with AD, it is important to note that vascular Ab deposits in macaque monkeys stain strongly against Ab42, whereas both Ab40 and Ab42 are present in senile plaques, contrary to humans (Nakumura et al. 1995; Kanemaru et al. 1996). Aged rhesus monkeys also demonstrate a significantly greater ratio of Ab40 to Ab42 compared to patients with AD, suggesting that primates exhibit differences in amyloid processing and deposition during the aging process (Gearing et al. 1996a, b). Despite these differences in amyloid deposition in nonhuman primates, apolipoprotein E, a1-antichymotrypsin and additional complement factors C1q and C3c in macaques are homologous to the isoforms identified in human AD (Podlisny et al. 1991; Abraham et al. 1989; Poduri et al. 1994; Hartig et al. 1997). This indicates an overall similarity in the cascade of molecular events that trigger the development of senile plaques in monkeys and humans..
(3) Recent rodent models for Alzheimer’s disease. While age-associated brain amyloid deposition has been studied in lower monkeys, no formal quantitative studies, to our knowledge have been performed in great apes due to the scarcity of brain specimens from aged animals, and their cost and maintenance is very expensive compared to rodents (Gearing et al. 1994, 1996a, 1997). Although cortical neuronal numbers remain preserved in aged macaque monkeys several alterations in cholinergic and aminergic neurons have been reported in the subcortical nuclei (Rapp and Amaral 1992; Siddique and Peters 1999). Progress loss of cholinergic neurons has been implicated in AD and extensive cholinergic deficits have been implicated in age-related memory loss and late stages of AD (Rapp and Amaral 1992). Therefore, cognitive deficits related to normal aging in older primates may be due to specific alterations to specific systems that are not necessarily visible in a light microscope. Age-related changes in both monoaminergic and cholinergic circuits have been observed in aged monkeys suggesting the possibility for alterations in specific receptor molecules, although the extent to which this may be true has not been examined (Goldman-Rakic and Brown 1981). Moreover, the relationship between cognitive function and age-related changes in neurochemical signalling has not been investigated. Fibrillary neuronal and neuritic changes have been reported in aged baboons, blue guenons, brown lemurs and rhesus monkeys (Schultz et al. 1998, 2000a, b; Hartig et al. 2000). These lesions are not only found in neuronal cells, but also in glial cells as well, and extend beyond AD pathology, since tau-containing glial lesions are also found in progressive supranuclear lesions and corticobasal degeneration (Buee et al. 2000). It is also important to note that fibrillary lesions in aged grey mouse lemurs are identical to the neurofibrillary tangles observed in the human AD brain (Kiatipattanasakul et al. 2000). However, tau proteins in these species are capable of undergoing agerelated modifications, which is unique to lemurs. In contrast to humans, the neocortical areas appear to be mostly affected in young mouse lemurs, while the subiculum and entorhinal cortex are only occasionally affected in animals aged 8 years (Kiatipattanasakul et al. 2000). Together, these data collectively suggest that tau-related pathology in certain primate species can demonstrate specific regional and cellular distribution that is different in human AD. Dogs Dogs appear to be an exceptional model to study human aging and AD (Speakman 2005). Domestic dogs have larger body sizes compared to other mammals, and have an elaborate social life being closely associated with humans, allowing advanced cognitive and behavioural (Hare et al.. 175. 2002) studies. The canids are highly suitable to model AD owing to the close homology of several APP processing genes, ApoE and presenilin between dogs and humans. Interestingly, the amino acid sequence of Ab42 is identical between dogs and humans, but differs by three amino acids in rats and mice (Johnstone et al. 1991). Moreover, the mass spectrophotometric profile of canine Ab peptides is identical to humans, although marked differences are observed compared to rats and mice (Du et al. 2007). Findings in dogs and early human brain amyloidosis suggest that dogs represent a relatively appropriate model to study amyloid deposition in the aging brain (Rofina et al. 2000; Satou et al. 1997; Pugliese et al. 2006; Russell et al. 1992). Like human AD, deposition of Ab in canines begins in the prefrontal cortex, including the gyrus proreus and the hippocampus (Rofina et al. 2000; Satou et al. 1997). However, unlike humans, dogs do not form dense neuritic plaques and neurofibrillary tangles (Satou et al. 1997; Pugliese et al. 2006). Also, Ab plaques do not occur in all aged dogs, suggesting that they are not part of normal physiological aging, but are formed as a result of numerous endogenous/exogenous factors making certain canine individuals vulnerable towards amyloidosis (Russell et al. 1992). Aged dogs have also been shown to exhibit a wide variety of cognitive abilities similar to AD. A pitfall of the extended use of dogs for modelling AD is the lack of accurate testing of visuospatial memory appropriate for the study of these domestic animals. Salvin et al. (2011a, b) recently developed a canine sand maze to test age-related changes in spatial memory in dogs. In this test, a 4.5-m diameter circular pool was filled with both sand and powdered food. Subjects are given 4 habituation and 16 learning trials leading to a food reward being half buried for control trials or completely buried for acquisition trials at a fixed locations. Younger dogs crossed the annulus zone surrounding the buried reward more frequently (2.6 times) than older dogs (1.5 times) (Salvin et al. 2011a). The same group has also developed a canine cognitive dysfunction rating scale (CCDR) for the assessment and longitudinal tracking of cognitive changes in community-based dogs (Salvin et al. 2011b). Dolphins Apart from primates and dogs, certain cetacean species such as dolphins have extended life spans that might be useful in modelling AD (Sarasa and Gallego 2006). Extensive Ab deposition has been recently reported throughout the brain of stranded dolphins, including the cerebellum and medulla oblongata. Ab depositions in the latter brain regions have only been observed in the most severe cases of human AD. This has lead to the hypothesis. 123.
(4) 176. that coastal breaching by these animals may occur as a response to significant cognitive deficit of aged individuals, leading younger animals. It is not surprising that the amino acid sequence of Ab42 for at least three dolphin species (Grampus griseus, Stenella coeruleoalba, and Tursiops truncatus) is identical between dogs and humans (Sarasa and Gallego 2006). Rabbits Rabbits, similar to guinea pigs, share the same Ab peptide sequence to humans, but do not develop AD pathology spontaneously (Johnstone et al. 1991). However, Sparks and Schreurs (2003) demonstrated that the addition of reactive copper to a group of cholesterol-fed rabbits can induce amyloid deposition and cognitive impairment in these animals (Sparks and Schreurs 2003). These findings suggest that redox active metals such as copper, but not aluminium or zinc can promote Ab aggregation (Sparks et al. 2006). As such, rabbits have become useful for modelling AD when validating the neuroprotective effects of metal chelators (Ghribi et al. 2006; Woodruff-Pak et al. 2007). Chicken embryo The chick embryo is a useful model to study AD because it is very accessible and easy to work with (Carrodeguas et al. 2005). Moreover, primary neuronal cultures derived from freshly dissected chick embryos are easy to grow and have been used to identify several assembly mechanisms associated with pathological lesions in AD (Carrodeguas et al. 2005). Abnormal aggregates of actin-associated protein cofilin are widespread in the AD brain and form punctuate and rod-like linear arrays through the neuropil (Minamide et al. 2000). Our group has recently demonstrated that mitochondrial inhibition in primary chick embryonic neurons can activate actin-depolymerising (ADF)/cofilin sequesters into rods that recruit tau and mimic neuropil threads (Whiteman et al. 2009). Chick neuronal cultures are therefore useful to study the effect of oxidative stress and mitochondrial dysfunction at the cellular level where access to human neuronal tissue is unavailable. Furthermore, the chick APP gene is identical to humans, and the Ab sequence is the same (Carrodeguas et al. 2005). Similarly, the chick embryo also appears to contain the necessary machinery required for the production of Ab protein during prenatal development, including BACE-1. BACE-2, PS1 and PS2, niastrin, and neprilysin, which is required for the degradation and clearance of the Ab peptide (Carrodeguas et al. 2005). Therefore, the chick embryo represents a valuable model to investigate the pathways regulating amyloidosis.. 123. N. Braidy et al.. Despite the importance of the chick embryo, the crucial question is whether adult-aged chicken represents an appropriate model for AD. So far, data that demonstrates evidence for cerebral amyloid plaques in aged chickens are not available, and it remains unclear whether older chickens suffer from any age-related neurobehavioral syndrome similar to dementia (Dani 1997). Current information arises from brain specimens obtained from 10-year-old chickens. As the approximate lifespan of chicken is about 30 years of age, these subjects are too young to develop amyloid deposition, and are not significantly different from the brains of chicken that are 2 and 14 months old (Dani 1997). Rats While rat remains the most commonly used experimental model to study neurodegenerative diseases, the use of transgenic mice models to study AD is increasing significantly. The rat has been used to study the effects of cortical Ab accumulation on the cholinergic, noradrenergic and serotonergic systems (Gonzalo-Ruiz et al. 2003). However, a major confounding factor hindering the use of rats to model AD is the significant variation in both the primary structure and complete profile of the Ab peptide (Du et al. 2007). Although transgenic models for AD have been shown to mimic some of the pathology associated with AD, these models are induced by genetic manipulation and therefore are unlikely to represent a true model for sporadic AD. Recently, evidence indicating impaired brain insulin signalling in AD (Hoyer 2004; De La Monte and Wands 2005) has led to the development of an experimental rat model using the drug streptozotocin (STZ). These animals typically exhibit reduced choline acetyltransferase activity within the first week of injection and the effect remains present for up to 3 weeks post-injection (Prickaerts et al. 1999; Ardiles et al. 2011). These changes are accompanied with significantly high levels of oxidative stress and nitrative stress that is common in several human neurodegenerative diseases (Sharma and Gupta 2002; Pathan et al. 2006). STV-icv administration can also lead to AD-related morphological changes as early as 1 week following a single drug dose. Glial fibrillary acidic protein (GFAP), a marker for astrogliosis, together with several inflammatory processes and markers for axonal neurotoxicity have been reported to increase in brain homogenates (Prickaerts et al. 1999). Despite these similarities to human AD, some morphological differences have been observed between human sporadic AD and the STZ-icv rat model which may be attributed directly to STZ actions. For instance, a significant enlargement in trans-Golgi segment have been observed 3 weeks after STZ injection which does not.
(5) Recent rodent models for Alzheimer’s disease. resemble the Golgi atrophy found in sporadic AD patients. The authors have suggested that this may be due to a marked desensitisation of the insulin receptor that predisposes cells to Ab deposition (Grieb et al. 2004). Changes in brain insulin and tau/Ab homeostasis have also been reported following STZ injection (Grunblatt et al. 2006; Salkovic-Petrisic et al. 2006). However, limited information is available regarding the immunohistochemical analysis of both tau protein and Ab plaques in these animals, making it difficult to interpret these findings (Chu and Qian 2005). STZ-icv treated rats also demonstrate deficits in learning, memory and cognitive behaviour as early as 2 weeks after injection and is maintained up to 12 weeks post-treatment (Prickaerts et al. 1999; Sharma and Gupta 2002; Grunblatt et al. 2006; Salkovic-Petrisic et al. 2006). While the exact mechanism for the loss in cognition in these rats remains unclear, reduced mitochondrial energy production, cholinergic deafferentiation, increased oxidative and nitrative stress, may play a key role in reduced memory and learning capacities (Weinstock and Shoham 2004). Also, a large amount of data has been collected to further our understanding of the pathological process in AD, including studies related to experimental lesioning as a model for cholinergic neuronal loss and changes to the complement (C) system in the brain during AD. The complement (C) system is an important humoral defence system containing at least 25 different proteins involved in the inflammation (Morgan 1990). Increased C mRNA has been reported in the cortex of the AD brain and occurs in part associated with microglia. Increased C mRNA has also been observed in the entorhinal cortex perforant path transfection in the rat brain (Johnson et al. 1992). Therefore, the cholinergic neuronal loss and reduced synaptic plasticity observed in AD may occur as a function of changes to C protein. Whilst extensive data are available to support the pathological role of Ab in AD, the amyloid hypothesis remains controversial, mainly because the nature of several Ab species and their effect on synaptic function remains unclear in vivo. Work by Walsh et al. (2002) and Glabe (2006) have shown that natural Ab oligomers with different C-terminal endings are formed within specific intracellular vesicles and are subsequently secreted from the cell (Glabe 2001, 2006; Walsh et al. 2002). Cerebral microinjection of a vehicle containing these oligomers and Ab monomers but no amyloid fibrils inhibited hippocampal long-term potentiation (LTP) in rats (Walsh et al. 2002). Most studies in transgenic mice over-expressing human APP fail to show any significant change in LTP. This may be due to the altered assembly forms of naturally occurring human Ab at physiological concentrations (Walsh et al. 2002).. 177. Mammalian hibernation as an attractive model for tau phosphorylation As previously mentioned, abnormal hyperphosphorylation of tau is a major pathological hallmark for AD. Up to 30 different phosphorylation sites have been identified in human tau protein, and have been implicated in numerous physiological and pathological processes (Stieler et al. 2011). Arendt et al. (2003) and others have hypothesised that tau phosphorylation initiates as a neuroprotective process that becomes cytotoxic during the course of the disease (Arendt et al. 2003). While this represents an important step in the pathogenesis of AD, the regulation of tau phosphorylation remains unclear, mainly due to a lack of appropriate animal models of tau phosphorylation. Stieler et al. (2008) recently showed that tau phosphorylation occurs in hibernating mammals, and this has been verified by others (Stieler et al. 2008). In addition to hibernation, increased tau phosphorylation has been observed following starvation, anaesthesia and cold water stress (Stieler et al. 2011). Under conditions of lowered metabolic activity, brain specimens from hibernating animals demonstrate increased levels of tau phosphorylation, which can be reversed following arousal. This correlates strongly with altered synaptic connectivity and impaired cognitive function and memory decline observed in AD (Stieler et al. 2011). Tau phosphorylation is temperaturespecific in the arctic ground squirrel (Spermaphilus parryii), Syrian hamsters (Mesocricetus auratus) and black bears (Ursus americanus), with more intense phospho-tau levels reported in torpid animals compared to euthermic animals (Stieler et al. 2011). Moreover, tau phosphosites are highly phosphorylated in early torpor and the degree of phosphorylation does not change as torpor progresses. The same study also reported a differential pattern of activity for tau kinase glycogen synthase kinase 3 beta (GSK3-beta) cyclin-dependent kinase 5 (cdk5), stressactivated protein kinase/Jun-amino-terminal kinase (SAPK/JNK) and mitogen-activated protein kinases/ extracellular regulated protein kinase (MAPK/ERK) in these species (Stieler et al. 2011). Although cdk5 and ERK1 are positively regulated in torpid animals, GSK3beta, SAPK/JNK and ERK2 are negatively regulated, and are analogous in both assessed species. It is highly likely that cdk5 and ERK1 are actively involved in tau phosphorylation under physiological conditions whilst activation of GSK3-beta and SAPK/JNK may indicate a pathophysiological mechanism as observed in AD (Stieler et al. 2011). Hibernating animals therefore have shown great importance to study the pathways regulating tau phosphorylation, and may play a vital role to test new therapeutics for the treatment of AD.. 123.
(6) 178. N. Braidy et al.. Non-transgenic non-mammalian animal models for AD Several non-mammalian species have also been used as experimental models to study the neuropathogenesis of AD. These include insects such as the fruit fly Drosophila melanogaster (Gunawardena and Goldstein 2001), roundworm such as the nematode Caenorhabditis elegans (Kraemer et al. 2003), and fishes such as sea lamprey Petromyzon marinus (Hall et al. 2001). While these lower organisms offer some unique experimental advantages, their brain structure and genetic composition is very different from humans, making any comparison in terms of neurodegenerative diseases relatively difficult (Table 1). Familial genetic forms of AD have led to the development of transgenic (Tg) animal models for several specific mutated genes (Spires and Hyman 2005). Such models have provided critical insights into the understanding of molecular mechanisms leading to neurodegeneration (Spires and Hyman 2005). Most transgenic rodent models are based on the over-expression of three genes: (1) amyloid precursor. protein (APP) and/or (2) presenilins (PS1 and 2), and/or (3) tau. Over-production and accumulation of the derived proteins lead, respectively to formation of senile plaques (SP), tau hyperphosphorylation and formation of neurofibrillary tangles (NFT) (Spires and Hyman 2005). These Tg mice can display neuropathological and behavioural features similar as those found in AD patients including amyloid deposits, neuritic plaques, astrogliosis, synaptic alteration, neuronal loss and memory impairment (Spires and Hyman 2005; German and Eisch 2004). Knockout models for APP, PS and Tau have also been developed to study the native functions of these genes (Table 2), and to examine whether AD pathogenesis is associated with a decrease or an increase in function of these pathways.. APP transgenic mice A predominant view of the cause of AD is the amyloid-b cascade hypothesis, i.e. Ab accumulation is the essential. Table 1 Advantages, limitations and modelled aspects of AD in experimental species Species. Advantages. Limitations. Identified aspects of AD. M. musculus (Sarasa and Pesini 2009). Comparable brain anatomy to humans Ab plaques and NFT reproducible. Crossbreeding/microinjection of transgenic lines expensive and time consuming. Significance of Ab plaques and NFT in AD pathology but significant neuronal loss. Learning and memory performance assessable using behavioural tests. Strain difference between transgenic lines. Role of familial AD in aetiology of AD Behavioural changes associated with region-specific pathology. Therapeutic benefits through examination of histopathology and behavioural tests D. melanogaster (Gunawardena and Goldstein 2001). Reproduction of biochemical changes in AD most fruitful Low cost. Neurodegeneration is less profound. Small size. Behavioural studies difficult to address. Easy to breed. Brain anatomy different from humans. Interaction of APP and tau may converge on similar cellular processes. Short lifespan Useful for high-throughput drug screening (HTS) for therapeutics C. elegans (Kraemer et al. 2003). First organism with fully characterised genomic sequence Easy to identify gene knockouts using RNA interference Low cost. Physiological role of APP as a receptor for axonal transport Over-expression of human tau can induce adult-onset progressive neurodegeneration. Neurodegeneration is less profound Brain anatomy different from humans Behavioural studies difficult to address. Isolation and characterisation of genes associated with AD Synaptic and pathological abnormalities associated with tau over-expression. Small size Easy to breed Short lifespan Useful for HTS for therapeutics P. marinus (Hall et al. 2001). 123. Central nervous system readily accessible. Neurodegeneration is less profound Brain anatomy different from humans Behavioural studies difficult to address. Stereotyped degenerative changes in AD over time.
(7) Recent rodent models for Alzheimer’s disease. 179. Table 2 Transgenes for the most commonly used transgenic mice mentioned in this review Line. Mouse strain. Mutations. Promoter used. Pathology. Cognitive impairment. Neural plasticity impaired. APP23. C57BL/6-DBA/2. K670N/M671L. Murine Thy-1. Ab deposits; congophilic senile plaques that are immunoreactive for hyperphosphorylated tau (Van Dam et al. 2003). Yes. No. PS2APP. B6D2F1 (i.e. C57BL/6J _ DBA/2 cross), B6CBAF1, and C57BL/6J. K670N/M671L. Murine Thy-1.2. Severe cerebral amyloidosis (Richards et al. 2003). Yes. Yes. TgCRND8. C57BL/6-CH3. K670N/M671L and V717F. Hamster PrP. Ab deposits, dense-cored plaques; and neuritic pathology (Chishti et al. 2001). Yes. No. PrP promoterdriven P301L. Swiss Webster-B6D2F1. P301. Murine PrP. Gene dose-dependent development of neurofibrillary tangles (Lewis et al. 2000). Yes. No. mThy1.2 promoterdriven P301L. Swiss Webster-B6D2F1. P301. Murine Thy-1.2. Neuritic tangles (Lewis et al. 2000). Yes. Yes. R406W. C57BL6/C3H. R406W. No. B6SJL. V337M. Filamentous intraneuronal tau aggregates (Tatebayashi et al. 2002) Accumulation of hyperphosphorylated tau (Tanemura et al. 2002).. Yes. V337M. Murine PrP PDGFpromoter. Yes. Yes. 39Tg-AD. C57bl6. PS-1 M146V APP K670M/ N671L Tau P301. Murine Thy1.2. Development of plaques and tangles (Hare et al. 2002).. Yes. Yes. Tg2576. C57B6J-SJLF hybrid mice. KM670/671NL. Hamster PrP. Ab deposits. Yes. Yes. event leading to neurodegeneration in AD (Hardy and Higgins 1992). This hypothesis is supported by research on transgenic mice expressing familial mutations of the human amyloid precursor protein (APP) and presenilins (Vardy et al. 2005; Hardy 2006). These animals reproduce some of the key aspects of the disease, including amyloid plaques, deficits in cognitive tasks, and abnormalities in the mechanisms of synaptic plasticity responsible for learning and memory (Higgins and Jacobsen 2003; Hsiao et al. 1996). However, they seldom develop neurofibrillary tangles and exhibit little synaptic and neuronal loss, which are pathological hallmarks of AD (reviewed in Gotz et al. 2004). This might relate to the fact that wild-type mice never develop AD-like pathologies (Gotz et al. 2006). In addition, information gleaned from models of familial genetic forms of AD may not translate to sporadic (nonfamilial) forms of the disease, the latter of which comprises more than 90% of the cases (Mattson 2004; Hoyer 2000). It remains unclear whether learning and memory deficits observed in AD patients and animal models is a direct consequence of Ab deposits in hippocampal and neocortical structures (reviewed in Gotz et al. 2004). The aim of. this part of this review is to correlate the histopathology, cognitive performances in learning tasks, and the changes in the synaptic plasticity after the apparition of Ab plaque deposition in the double-transgenic mice APP23, PS2APP and TgCRND8 (Table 2). Histopathology It has been shown that functional changes, preceding the Ab plaque deposits can underlie the initial cognitive deficits reported in both AD patients and animals models (Kamenetz et al. 2003). Histological examination was unable to demonstrate any Ab plaques before the age of 6 months in APP23 mice and 5 months in PS2APP mice. Additionally, Ab peptides were found in the cortex and the hippocampus from the age of 5 months in both models. TgCRND8 mice manifested early and progressive Ab plaques formation in their brains from 12 weeks of age (Chishti et al. 2001). The role of NFT in the progression of neurodegeneration is strongly debated since they also occur in a number of different neuropathological conditions such as amyotrophic. 123.
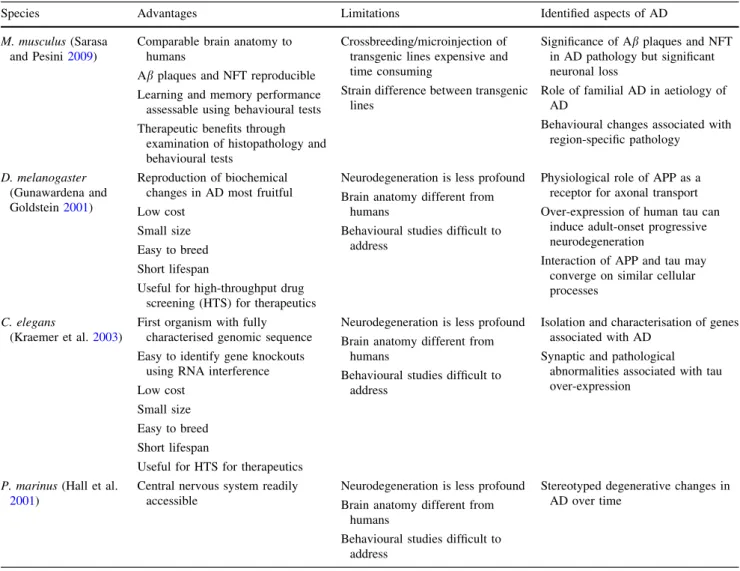
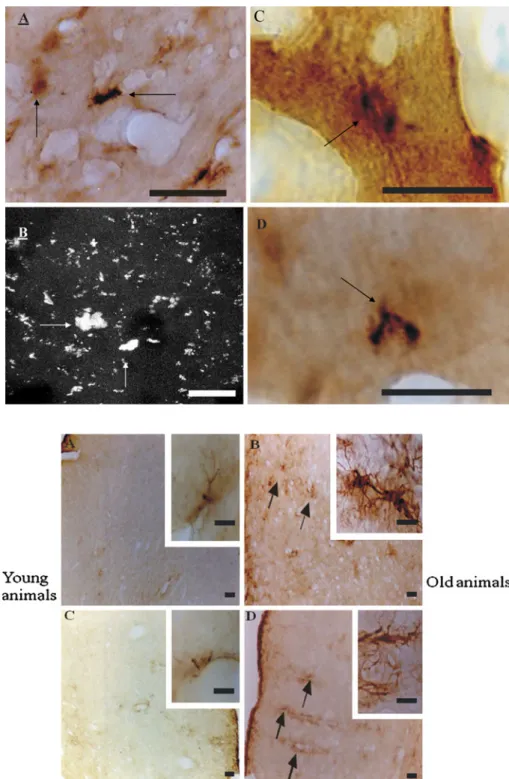
(8) 180. N. Braidy et al.. lateral sclerosis, progressive nuclear palsy and frontotemporal dementia. None of the transgenic mice containing APP alone showed signs of NFT (reviewed in Howlett and Richardson 2009). Extensive data are available indicating that inflammation is a key feature of AD pathology. Heightened inflammation and cortical atrophy are consistent and prominent features of both the genetic and sporadic forms of AD (reviewed in Howlett and Richardson 2009). Our group has previously demonstrated that activated microglia and astrocytes encase Ab deposits and infiltrating macrophages and other blood cells are present within the plaques in AD tissue. PS2APP mice exhibit plaque pathology as early as 3 months of age, and these plaques were strongly associated with activated glial cells (Howlett and Richardson 2009). Behavioural studies The Morris water maze (MWM) is a widely used behavioural test to evaluate spatial memory performance in rodents (Table 3). Memory acquisition is reflected as the performance to find the platform (escape latency) and path length, whereas a probe trial is used to assess storage and retrieval (D’Hooge and De Deyn 2001). At the age of 6–8 weeks, APP23 mice performed slightly lower compared to wild type only during the first two acquisition trials of the hidden-platform. After additional trials, both genotypes learned the position of the platform in a similar manner, which is consistent with learning as a gradual process that requires reinforcing (Van Dam et al. 2003). At the age of 3 months, APP23 mice swam a significantly greater distance and needed a significantly longer escape latency to reach the platform when compared to wild-type mice. Deviating acquisitions curves, both in path length and escapes latency, were also observed at the age of 6 months (Van Dam et al. 2003). However, no significant difference was observed between the PS2APP mice and wild type at 5 months (Van Dam et al. 2003). Table 3 Advantages and disadvantages of using Octodon degu compared to the current transgenic mice Advantages. Disadvantages. Complete physiological phenotype. Longer lifespan. Inherent age-related decline in cognition. Difficult to breeding. All pathology hallmarks of AD, including Ab deposits and tau hyperphosphorylation Extensive neuronal loss occurring with age Other pathological markers such as inflammation and gliosis Impaired neural transmission. 123. Both APP transgenic mice and control groups showed a trial-dependent decrease in latency between 8 and 12 months of age, but the transgenic group was slower overall, whereas no group differences were observed on cued learning in MWM. At 12 months, no group differences were found on cued learning in Morris water maze, although a significant main effect of genotype was revealed on place learning (Richards et al. 2003). In both the spatial and cued versions of the MWM, there were no significant differences between 6- and 8-week-old TgCRND8 and non-TgCRND8 in escape latency or path length. No differences were observed between 19- and 22-week-old TgCRND8 and non-TgCRND8 mice on the cued version, whereas, TgCRND8 mice took longer than non-TgCRND8 mice to locate the hidden escape platform in the spatial version. By contrast, both TgCRND8 and non-TgCRND8 at 39- to 40-week-old mice showed poor performances (Hydea et al. 2005). In a single trial avoidance paradigm, mice are allowed to cross from an illuminated chamber into a dark chamber. As soon they entered the dark compartment, the animal received a punishing electrical shock creating a stable memory trace. Memory was then measured as the tendency for the animals to avoid the dark side in subsequent trials. In this experiment, the latency to re-enter the dark compartment was not affected in the APP23 mice at 6–8 weeks, 3 months and 6 months (Van Dam et al. 2003). These results indicate that the retention latency was higher than training latency, indicative of memory formation, in all age groups except the 25-month-old transgenic. While no marked effect on retention latency was observed with age in wild type, APP23 mice demonstrated markedly short retention latency, indicative of impaired memory retention in this transgenic model. Moreover, the retention latency in the 25-month APP23 group was significantly less than that in same age wild-type mice and even lower than that in the same-genotype 3-month-old mice. Thus, only the oldest APP23 mice were impaired (Kelly et al. 2003). No significant effect was observed in avoidance behaviour in PS2APP mice compared to controls at 4 months of age. However, slower acquisition of the avoidance behaviour was observed in the transgenic group between 8 and 12 months of age (Richards et al. 2003). At 12 months of age, main effect of genotype was recorded on percentage avoidance, suggesting that with extended training the transgenic group would reach the final avoidance level of controls. At 16 months, the transgenic group reflected severe learning impairment in the PS2APP mice (Richards et al. 2003). Long-term potentiation (LTP) is a form of plasticity that may be associated with learning and memory deficits. Although an effect related to age was observed, it was not.
(9) Recent rodent models for Alzheimer’s disease. possible to detect age-dependent changes for basal synaptic transmission, and the magnitude of the LTP was not significantly different in the hemizygous APP23 mice compared with the wild-type animals (Kelly et al. 2003). At the Schaffer collateral-CA1 hippocampal pathway from PS2APP mice, an effect related to the age was not observed in short-term plasticity, relative to paired-pulse facilitation or long-term potentiation. Only subtle attenuation on the post-tetanic potentiation (PTP) was observed in transgenic mice in the 9-month age group (Richards et al. 2003). There was no significant difference in LTP in 9-weekold TgCRND8 mice and wild-type littermates. In contrast, there was a significant increase in LTP in 20-week-old TgCRND8 mice when compared with wild-type mice. These differences cannot be explained by changes in the basal synaptic transmission, because, normalised input– output curves generated in slices from transgenic mice were not significantly different from those generated in age-matched wild-type controls (Jolas et al. 2002). Taken together, APP transgenic mice have highlighted the importance of Ab in plaque formation and the development of AD-like behavioural symptoms. Double-transgenic models containing the c-secretase component PS2 have further replicated the disease-enhancing role of presenilins in AD. However, while these models have contributed significantly to our understanding of the pathogenesis of AD, they fail to develop NFT. As previously mentioned NFT are the second-most important histopathological hallmarks of AD and have been addressed in tau single transgenic mice models.. 181. mention in this review are the PrP and mThy1.2 promoterdriven P301L, V337M and R406W mice. Histopathology When a human tau isoform where the two amino-terminal inserts are absent is expressed using the PrP promoter, P301L tau levels were significantly higher in neurons found in the somatosensory cortex and motor neurons of the spinal cord compared to mThy1.2-driven P301L mice (Gotz and Nitsch 2001). AD is not a disease of motor neurons, suggesting that P301L mice containing the PrP promoter may have developed an unusual phenotype that is not representative of AD (Lewis et al. 2000; Gotz and Nitsch 2001). Extensive NFT were identified in PrP-driven P301L mice using Gallyas silver staining and thioflavin-S fluorescence microscopy (Gotz and Nitsch 2001). These pathological lesions are reported as early as 4.5 months of age in homozygous mice, and around 6.5 months of age in hemizygous tau mice. NFT developed mainly in the amygdala of mThy1.2 promoter-driven P301L mice (Gotz et al. 2001). Cortical brain areas contained both tau-positive astrocytes and neurons. These models are not ideally suited because glial tau inclusions are not commonly observed in AD pathology (Gotz et al. 2001). The V337M tau mutant mice only express tau in the hippocampus (Tanemura et al. 2002). Likewise, R406W transgenic mice express tau in the hippocampal region and to a lesser extent, in other cortical and subcortical regions, with relatively little tau expression in the amygdala, even in 16- to 23-month-old animals (Tatebayashi et al. 2002).. Tau transgenic mice Behavioural studies Tau belongs to a group of neuronal microtubule-associated proteins that forms an essential component of the paired helical filaments that make up NFT in the AD brain (Alonso et al. 2001). Tau is a key regulator of microtubule dynamics, axonal transport and neurite outgrowth, all of which are modulated through site-specific phosphorylation (Alonso Adel et al. 2004). Abnormal hyperphosphorylation has been found to occur in neurodegenerative disorders including AD, but the contribution of tau to neuronal and synaptic loss remains controversial (Alonso Adel et al. 2006; Alonso et al. 1996). This is largely because mice models for AD over-expressing APP alone or in conjunction with PS develop Ab deposits but indicate no signs of NFT or hyperphosphorylation of tau (Howlett and Richardson 2009). To elucidate the role of tau in AD pathology, transgenic mouse models have been developed by expressing the human tau protein using several different promoters. The main tau transgenic mice models worthy of. Over-expression of human tau has demonstrated evidence to mimic behavioural disturbances observed in AD. The PrP promoter-driven mice showed a poor ambulatory response within 2 weeks of phenotype onset (Lewis et al. 2000). Dystonic posturing accompanied by severe fatigue in the extremities were observed within 3–4 weeks of phenotype onset and these mice became dilapidated. These deficits in balance correspond to the most severe human cases of AD symptomatology (Lewis et al. 2000). These mice were excluded from memory and learning performance tests, e.g. MWM, due to severity of symptoms (Lewis et al. 2000). The open-field test and elevated plus maze are common behavioural tests used to assess anxiety-related behaviour. The V337M tau mutant mice displayed hyperactivity and enhanced locomotion in the open-field and elevated plus maze. This indicated that V337M mutants show an. 123.
(10) 182. anxiogenic phenotype that is confirmed in the elevated plus maze (Tanemura et al. 2002). Additionally, these mice presented with memory deficiency in the elevated plus maze suggestive of AD-like symptoms. On the contrary, R406W mice showed a slight reduction in locomotor activity compared to V337M mice. However, cognitive deficit in R406W mice was detected using the contextual fear conditioning (CFC) test (Tatebayashi et al. 2002). Since NFT occur more commonly in the amygdala of mThy1.2 promoter-driven P301L mice, these animals were assessed using amygdala-specific tests (LeDoux 2000). The amygdala is thought to be a key regulator of basic emotions such as fear, anger and craving. Amygdala atrophy is observed early in AD leading to impaired learning and memory, and emotional deficit (Tatebayashi et al. 2002). These symptoms can be assessed in animal models using the CFC and conditioned-taste aversion (CTA) tests. Surprisingly, P301L and aged APP mutant mice demonstrated a lack of deficit in fear conditioning similar to wild-type mice (Pennanen et al. 2004). This is unexpected, since both mice models demonstrate either increased NFT and tau aggregation, or Ab plaques in the hippocampus and amygdala, respectively. This observation may be explained by the lack of neurodegeneration detected in animal models for AD and negative results reported for the CFC and CTA tests which contradict the impaired feared conditioning reported in human AD patients (Hamann et al. 2002). CTA is a well-established learning and memory-based assessment task in which subjects are taught how to link a novel taste with nausea. Consequently, these subjects avoid consumption of this specific taste at the next presentation (Pennanen et al. 2004). P301L transgenic mice showed no significant impairment in the acquisition and consolidation of CTA memory. However, P301L mice extinguished the CTA more rapidly than wild-type mice. This suggests that acquisition of CTA is impaired in the presence of tau aggregation in the amygdala and is sufficient to cause ADlike behavioural deficits even in the absence of NFT (Pennanen et al. 2004). The choice of promoter used to drive tau over-expression in tau transgenic mice can potentially influence the pattern of tau pathology leading to the development of vastly dissimilar behavioural phenotypes.. Bigenic APP and tau transgenic mice As previously mentioned, AD pathology is characterised by neuritic plaques containing extracellular Ab deposits, and NFT containing intracellular bundles of hyperphosphorylated tau protein (reviewed in Suh and Checler 2002). How these pathologies are associated with the progressive loss of cholinergic neurons in AD remains unknown. The. 123. N. Braidy et al.. major problem regarding the development of mouse models for AD is the ability to develop animals that contain both pathological hallmarks of AD. For instance, homozygous transgenic mice expressing human wild-type tau were unable to develop NFT in the presence of Ab1–42 even at 12 months of age (Gotz et al. 2001). In contrast, mThy1.2 promoter-driven P301L mice injected with Ab1–42 developed an up to fivefold increase in NFT formation 18 days following Ab1–42 injections in the amygdala (Gotz et al. 2001). These studies further reinforce a potential interaction between Ab and tau protein. To further investigate the interaction of Ab and tau, Lewis et al. (2000) crossed APP Tg2576 transgenic mice with mThy1.2 promoter-driven P301L tau mutant mice. These mice showed an almost sevenfold increase in NFT formation in the olfactory bulb and amygdala compared to single tau transgenic mice alone. However, no significant difference in the amount of tau expression was observed between single- and double-transgenic mice (Lewis et al. 2001). The enhanced NFT pathology was therefore thought to be associated with region-specific Ab deposition in bigenic mice and reflects the neuropathological correlates of the AD brain (Lewis et al. 2001).. Triple transgenic mice As mentioned several transgenic mouse models have proven to be invaluable tools for increasing our understanding of the pathogenesis of AD. However, it is clear that no single animal model can accurately represent the neuropathological hallmarks of AD. APP transgenic mice have successfully replicated Ab plaques, and addition of a mutant PS2 allele can potentiate disease progression similar to AD, but they do not develop NFT (Howlett and Richardson 2009). Similarly, while tau transgenic mice mirror the observed tauopathy in AD they do not develop Ab plaques, which are fundamental lesions in AD (Tanemura et al. 2002). Simultaneous formation of Ab plaques and NFT have required the insertion of several transgenes into the same mouse by either microinjecting the pathological protein into the brains of monogenic mice or crossing several independent transgenic lines. In one study, PS1 M1461 knocking mice were co-injected with constructs obtained from mutant (APPswe) and mThy1.2 promoter-driven P301L tau transgenes (Lewis et al. 2001). These models were more efficient than crossbreeding in terms of speed and did not alter the genotype of these animals. The triple transgenic mouse (39Tg-AD) (Oddo et al. 2003) harbors three mutant genes: Ab precursor protein (APPswe), presenilin-1 (PS1M146V), and tauP301L. This model was developed by inclusion of two additional.
(11) Recent rodent models for Alzheimer’s disease. transgenes into the germline of a single transgenic mouse model. The 39Tg-AD mice is the first model, to our knowledge to progressively develop Ab, NFT and tau pathology, with a temporal- and regional-specific profile that closely mimics their development in the human AD brain (Oddo et al. 2003). These mice display most of the neuropathological and behavioural features of AD including Ab deposits, neuritic plaques, gliosis, synaptic damages, neurodegeneration and memory impairment (Dodart et al. 2002). Histopathology As with single APP transgenic mice, intraneuronal Ab immunoreactivity is observed in neocortical regions between 3 and 4 months of age in 39Tg-AD. Similarly, extracellular Ab deposits were first observed in 6-monthold mice in the frontal cortex and are prevalent in the hippocampus by 12 months of age (Dodart et al. 2002). However, it is unclear whether the anti-Ab antibodies used in this study can distinguish between the free Ab peptide and the same Ab epitope within the intraneuronal APP. Using 39Tg-AD mice, Winton et al. (2011) showed that intraneuronal Ab share epitopes with APP but not free Ab. While APP and tau were both expressed at comparable levels in the same brain region, the amount of tau in 39TgAD was significantly higher than previously reported for P301L mice (Dodart et al. 2002). The tau pathology was first observed by 12 months in 39Tg-AD within pyramidal neurons of the CA1 subfield (Dodart et al. 2002). Therefore, while both Ab and tau expression is comparable in triple transgenic mice, Ab pathology occurs much earlier than tau pathology, which is consistent with the amyloid cascade hypothesis. Using 39Tg-AD mice deficient of b-secretase (BACE), the protease necessary for Ab production, Winton et al. (2011) further showed that tau lesions were not different between 39Tg-AD and 39Tg-AD/ BACE-/-, suggesting that tau pathology forms independently of amyloidosis in this mouse model. Behavioural studies Another important manifestation of AD is neuronal and synaptic dysfunction. Synaptic dysfunction strongly correlates with memory and cognitive deficits observed in AD. To assess basal synaptic transmission, Oddo et al. (2003) measured field-excitatory postsynaptic potentials (fEPSPs) elicited in CA1 following stimulation of the Schaffer collaterals at increasing stimulus intensities. Although no significant difference was observed between 1 month of age and gender-matched non-Tg animals, smaller fEPSP slopes and amplitudes were reported at all stimulus intensities tested between 39Tg-AD and non-Tg mice (Dodart. 183. et al. 2002). However, no significant difference was reported in 39Tg-AD, and PS1-Kl mice, which contain a line expressing a mutant form of human PS1 (M146V). This suggests that basal synaptic impairment is an important pathology in both PS1-Kl and 39Tg-AD after 6 months of age (Dodart et al. 2002). Short-term plasticity was modelled in these mice using paired-pulse facilitation (PPF). No significant difference in PPF was observed between 39Tg-AD, PS1-Kl and non-Tg mice (Dodart et al. 2002). However, PPF was significantly elevated in PS1-Kl compared to non-Tg mice. PPF are thought to be regulated by a presynaptic mechanism involving residual Ca2? in the nerve terminal during the first stimulus, with a subsequent increase in neurotransmitter release following the second stimulus (Zucker and Regehr 2002). The elevated PPF in PS1-Kl mice may be due to modification of intracellular Ca2? levels, which may be eliminated by over-expression of APP in triple transgenic mice. LTP was also assessed in 39Tg-AD mice in the CA1 hippocampal region (Dodart et al. 2002). While no significant difference in LTP was observed between 39TgAD, PS1-Kl, and non-Tg mice at 1 month of age, LTP was severely impaired in 6-month-old 39Tg-AD mice compared to non-Tg mice (Dodart et al. 2002). The finding that synaptic dysfunction and LTP deficits in 39TgAD mice develops prior to the formation of Ab plaques and NFT suggests that impaired synaptic transmission is an early symptom of AD that Ab deposition is not the underlying cause of synaptic deficit (Dodart et al. 2002). This observation reinforces previous studies using other transgenic mice models where synaptic transmission is compromised in the absence of plaque formation. Triple transgenic mice have therefore increased our appreciation of the important role of synaptic dysfunction in early stages of AD.. Five familial AD (FAD) mutations While triple transgenic mice, which express APP and PS1, overproduce Ab42 with similar histopathology observed in AD, these models develop plaques very slowly. Recently, transgenic mice with five familial AD mutations have been developed to accelerate plaque development (Oakley et al. 2006). These models co-express a total of five FAD mutations [APP K670N/M671L (Swedish) ? I716V (Florida) ? V717I (London) and PSq M146L ? L286V] to rapidly drive Ab42 plaque formation at 2 months of age. These ‘‘59FAD’’ transgenic mice also accumulate higher amounts of Ab42, marked gliosis, extensive neuronal loss, reduced synaptic markers, and significant loss in memory and cognitive behaviour (Oakley et al. 2006).. 123.
(12) 184. Histopathology The 5XFAD mice produce excessive amounts of cerebral Ab42, exceeding that of Ab40. Therefore, FAD mutations, when combined together can generate an additive effect to markedly increase Ab42 production in vivo, and develop AD-like amyloid pathology at a very young age (2 months) (Oakley et al. 2006). Moreover, these models show extensive accumulation of intraneuronal Ab42, as determined by thioflavin-S staining, prior to plaque formation, and occur within neuronal soma and neurites. Intraneuronal Ab42 has been reported in the AD brain. Indeed, amyloid plaques have been thought to originate from neuronal cell bodies. The current data from 59FAD mice suggest that intraneuronal Ab aggregates are involved in neurodegeneration and related neuronal loss and amyloid plaque formation (Oakley et al. 2006). Unlike other APP transgenic mice, 59FAD do not display abnormal hyperphosphorylated tau. Although the reason for this phenomenon remains unclear, further analysis of pathological tau epitopes in these rodent models is required (Oakley et al. 2006). In addition, important synaptic markers such as synaptophysin, syntaxin, and postsynaptic density95 are reduced with age in 59FAD mice, and extensive loss of cholinergic neurons in cortical layer 5 and the subiculum are lost, possibly as a result of the Thy1 transgene promoter expression pattern (Oakley et al. 2006). Moreover, cyclindependent kinase 5 (cdk5) and p25 are all elevated at 9 months in 59FAD mice, with an upward trend observed after 3 months, before significant neuronal loss is observed. In AD, calpain cleavage generates p25, which subsequently activates cdk5, and plays a key role in promoting Ab toxicity (Oakley et al. 2006). While these models provide a greater understanding of AD-related pathology, there are several issues that need to be considered when interpreting data obtained from these models and its relevance to human AD. Firstly, no cases of AD have been reported when all five familial mutations are present. Secondly, the ratio of Ab42 to Ab40 is higher in 59FAD mice, therefore suggesting that a much greater degree of toxicity exists in 59FAD mice compared to the human AD brain. While these points of criticism are equally related to other transgenic mice models, they have provided invaluable insight into the pathogenesis of AD and highlighted new therapeutic strategies to treat this debilitating disease. 59FAD mice develop amyloid pathology more rapidly than in human AD and other transgenic models, making it a potentially relevant model to study AD-like amyloidosis. Behavioural studies Unlike in 39Tg-AD mouse, which demonstrate long-term retention memory deficits in the MWM and accumulation. 123. N. Braidy et al.. of intraneuronal Ab at 4 months, 59FAD mice do not show any Y-maze deficit at 2 months of age, although intraneuronal Ab begin to accumulate at 1.5 months. The Y-maze does not measure long-term memory retention, and this can explain the difference in behavioural results between the two models (Oakley et al. 2006). The observed Y-maze deficit in 59FAD mice occurs at 4–5 months of age, and correlates with the decline in whole-brain synaptophysin levels, suggesting that 59FAD-mediated memory dysfunction may be associated with neurodegeneration (Oakley et al. 2006). Recently, Jawhar et al. (2011) showed that overexpression of glutaminyl cyclise (QC), the enzyme responsible for pyroglutamate Ab formation can induce behavioural deficits. In that study, 59FAD mice were crossed with transgenic mice expressing human QC using the Thy1 promoter. These mice demonstrated significantly higher levels of AbpE3 compared to 59FAD mice correlating with significantly greater motor and working memory impairment using the Y- and cross-maze (Jawhar et al. 2011). Similarly, QC knockout was able to rescue the wildtype mice behavioural phenotype in 59FAD mice (Jawhar et al. 2011). This highlights the reduction of QC as a new promising therapeutic target for the treatment of AD.. Implications for Bace1 and Apoe4 The central role of Ab and tau pathology in the development and progression of AD has been well established. However, other factors may also play a crucial role in the pathogenesis of this disease. BACE1, a type 1 transmembrane aspartyl protease has been identified as the b-secretase involved in cleavage of APP to Ab (Vassar et al. 1999). BACE1 knockout mice expressing the APPswe and PS1 mutant do not develop Ab plaques. Moreover, Tg2576 mice which over-express the APP gene (hAPP695) containing the Swedish (K670N) mutation have demonstrated a significant reduction in Ab levels and no significant impairment in memory and learning following BACE1 gene deletion (reviewed in Li 2004). These studies provide a strong rationale for the development of novel anti-amyloidogenic therapeutics, which target BACE1. In addition, while several genetic risk factors have been associated with the onset and development of AD, only the ApoE4 allele has been directly linked to the pathogenesis of AD (reviewed in Rocchi et al. 2003). ApoE is a glycoprotein, which transports cholesterol and other lipids in the bloodstream. While several different isoforms of ApoE are known to exist, ApoE4 has demonstrated the greatest ability to bind Ab and thus influencing Ab aggregation and deposition without affecting Ab synthesis (Brendza et al. 2002). APP transgenic mice (APPswe) lacking the ApoE4.
(13) Recent rodent models for Alzheimer’s disease. gene have shown a reduction in Ab deposition compared to normal APP mice. Additionally, ApoE4 null mice (-/-) expressing human APP (hAPP770) with a FAD mutation (V717F) showed significant improvements in learning and memory deficits compared to ApoE4 ?/? animals (Holtzman et al. 1999, 2000). It has been postulated that the ApoE4 allele may contribute to the impaired Ab metabolism, which can increase the predisposition of late-onset AD. The ability to study similar neuropathological processes in living animals have provided over the years new insights into potential disease mechanisms and more importantly the opportunity to test therapeutic agents.. Transgenic rat models for AD Despite the advancement in research that has been obtained from transgenic mouse models for AD, efforts have also been directed at the development of transgenic rat models which deposit Ab (Spires and Hyman 2005). This is largely because rats are more acquiescent than mice for studies involving canniculation, electrophysiology and neuroimaging. Moreover, rats appear to be a better model than mice for experiments such as behaviour, cognitive function and electrophysiology (Spires and Hyman 2005). Several transgenic rat models have been developed using similar transgenes expressed in transgenic mouse models. The GP56 rat over-expressing the APP695 transgene used in the Tg2576 APP transgenic mouse model expressed very low levels of Ab and Ab deposition was not detected in aged models (Hsiao et al. 1996). Similarly, transgenic rats expressing mutations in human APP751 and M146V mutation in human PS1 did not develop extracellular Ab deposition by 24 months (Lopez et al. 2004). Cephalon (a US biotech company) has also recently developed an APP/PS1 39Tg-AD rat. This rat model is the first to show both amyloid deposits and NFT neuropathology (Flood et al. 2007). Essentially, Ab deposits were either fibrillar or diffuse. Fibrillar deposits were associated with significant glial reactivity and tau hyperphosphorylation similar to that described in mouse models and the AD brain (Flood et al. 2007). Rats are practical species to remodel Ab deposition in the AD brain for research requiring larger rodents. Prior to the development of the transgenic rat models with Ab deposition, it was difficult to determine why such models were not possible to obtain compared to the successes of several mouse models already available (Spires and Hyman 2005). Persistent failures have suggested the possibility that the rat brain might be more resistant to extracellular Ab deposition than mice (Flood et al. 2003). Strain differences in Ab deposition have. 185. been previously reported in mice models containing the same transgene (Flood et al. 2003). Nevertheless, the development of additional transgenic rat lines is essential before the relative resistance of rats to Ab deposition can be evaluated.. Involvement of transgenic rodent models for the development of novel therapeutics for AD At present, there is no known cure for AD. While several drugs have been developed to slow down cognitive decline, these drugs do not stop the progression of the disease. Novel approaches have been recently developed for the treatment of AD. These therapeutic targets are mostly aimed at limiting Ab production, Ab aggregation, metal accumulation and the secondary neuroinflammatory and neurotoxic cascade. Vaccination trials against Ab Several groups have used mouse models over-expressing APP and developing amyloid plaques to test immunisation strategies (Nikolic et al. 2007). Schenk et al. (1999) immunised the PDAPP (platelet-derived growth factor driven APP) mice with Ab1–42 peptide leading to the induction of an active immune response by the host able to recognise and clear the Ab plaques. The results showed that this active immunisation of young animals prevented the development of senile plaques, neuritic dystrophy and astrogliosis, while treatment of older animals reduced the extent and progression of the AD-like neuropathology, suggesting that immunisation with Ab was effective in both preventing and treating AD. However, the clinical trial in humans (AN-1792) was halted as 6% of patients developed meningo-encephalitis (cerebral inflammation) associated with immune activation (Schenk et al. 2005). Therefore, Ab plaque removal does not appear to be sufficient enough to stop the progression of AD, thus challenging the amyloid hypothesis. In May 2007, Elan and Wyeth announced a phase 3 clinical trial of their new antibody AAB-001 (bapineuzumab) for the treatment of patients with mild to moderate AD. Elan and Wyeth expect that AAB-001 will be as efficient as AN-1792 to clear Ab but with little or no adverse reactions and a therefore more tolerable safety profile (reviewed in Gotz and Ittner 2008). Additional antibodies under testing include LY2062430 (solanezumab, Eli Lilly, a phase III study ongoing), PF-04360365 (Pfizer, phase I completed), R1450 (Hoffman-LaRoche, phase I completed), GSK933776A (GlaxoSmithKline, phase I ongoing) and V950 (Merck, phase I ongoing) (Galimberti and Scarpini 2011).. 123.
(14) 186. N. Braidy et al.. Blocking Ab assembly. Metal chelators. Following the discovery that b-sheet breakers can inhibit Ab fibrillogenesis modified peptides have been developed that can reduce Ab deposition in APP V717I and APP V717I/ PS1 A246E transgenic mice (Permanne et al. 2002). However, these drugs require multiple weekly dosing and may therefore be unpractical for long-term treatment of AD.. Another strategy that has been tested on rodent models involves lowering of metal levels such as zinc, iron and copper that form complexes with Ab. Administration of compounds such as clioquinol (PBT1) and hydroxy-quinolinine (PBT2), which chelate zinc and copper, have led to a substantial reduction in brain Ab deposition in Tg mice (Cherny et al. 2001; Dedeoglu et al. 2004). A pilot phase 2 clinical trial using clioquinol has been performed in 36 patients with moderately severe AD showed some promising results. Some of the treated patients displayed a slower cognitive decline and lower plasma levels of Ab1–42. While the drug showed limited side effects, the trial was stopped due to the presence of toxic impurities in the drug formulation (Ritchie et al. 2003).. c-Secretase inhibitors Other groups have shown that administration of various gamma-secretase inhibitors in APP transgenic mouse models can reduce brain levels of Ab (Asberom et al. 2007) often in a dose-dependent manner, and without significant neuroinflammation and toxicity (Prasad et al. 2007). Moreover, the use of c-secretase inhibitors can inhibit synaptotoxic Ab oligomer formation at doses that promote monomer production without disrupting LTP (Walsh et al. 2002). This could have important implications for the treatment of protein-folding disorders. However, c-secretases have additional substrates such as the Notch signalling protein which is required for the differentiation and proliferation of embryonic cells, T cells, gastrointestinal goblet cells, and splenic B cells (Wong et al. 2004). Doses of these drugs have been shown to not only lower Ab concentrations but also interfere with lymphocyte differentiation in a transgenic mice model (Wong et al. 2004). Therefore, safety is important when considering this class of compounds for the treatment of AD. Reduction of tau Recent studies have shown that whilst Ab vaccination trials can reduce Ab levels, they have limited effect on tau pathology (Kulic et al. 2006). However, reduced tau levels and NFT were observed in JNPL3 mice vaccinated with a tau peptide containing PHF1, a phospho-epitope (Asuni et al. 2007). Boutajangout et al. (2010) recently developed a new accelerated tangle mouse model to study tau immunotherapy and prevent cognitive decline by crossing htau mice with a model carrying PS1 M146L mutation. These mice display tau pathology more rapidly than the htau mice, although the distribution is similarly located in the hippocampal and cortical regions, making the model highly suitable to study tau-targeting therapy (Boutajangout et al. 2010). The same group showed that tau immunotherapy can ameliorate cognitive decline using the radial arm maze, closed field symmetrical maze and object recognition learning and memory tests (Boutajangout et al. 2010). This study is the first to indicate that immunotherapy targeting tau is feasible for the treatment of tauopathies such as AD.. 123. Lowering cholesterol While the ApoE4 allele has been implicated in the pathogenesis of AD, it is also a key regulator of lipid metabolism. Subsequent studies have indicated that other lipids, such as cholesterol may be involved in the development of AD (Puglielli et al. 2001). Increased cholesterol levels correlate strongly with elevated Ab deposition in vitro and using animal models of AD, and drugs which inhibit cholesterol synthesis have been shown to reduce Ab in these models (Puglielli et al. 2003). Antioxidants Dietary antioxidants have also been envisaged as potential therapeutics for the treatment of AD. Apart from their antioxidant properties, green tea polyphenols such as Gingko biloba have been shown to reduce Ab production in Tg2576 APP transgenic mice (Rezai-Zadeh et al. 2005). Additionally, a diet consisting largely of omega-3 polyunsaturated fatty acids (FFA) has been shown to prevent the formation of extracellular Ab plaques, although the exact mechanism remains unclear. FFA are thought to suppress Ab aggregation by reducing secretase activity and therefore preventing the cleavage of APP to the neurotoxic Ab1–42 (Lim et al. 2005). Calorie restriction An increasing number of studies have shown a potential therapeutic role for calorie restriction (CR) in the treatment of AD. Calorie restriction has been shown to reduce Ab deposition in APP transgenic mice with a concomitant reduction in astrocyte activation (Kim et al. 2007). Similarly, CR improved memory and learning deficit in 39TgAD mice. However, levels of Ab and tau were not.
(15) Recent rodent models for Alzheimer’s disease. significantly affected, suggesting that CR may exhibit a protective effect downstream if Ab and tau (Halagappa et al. 2007). Wnt signalling activation Previous studies indicated that the activation of the Wnt signalling pathway protects hippocampal neurons from Ab neurotoxicity, similar results plus the reversion of behavioural changes were obtained in a rat model of AD treated with lithium, a GSK-3b inhibitor (Morgan 1990). Recent studies using the double transgenic (APPswe ? PS-1) mice, indicated that both lithium and rosiglitazone, a PPARa agonist, reverse behavioural derangements, decrease Ab oligomers and increase several Wnt components, including, dishevelled, b-catenin and GSK-3b (Johnson et al. 1992). Other targets Several other studies have been recently performed or are currently in progress targeting other mechanisms involved in AD such as amyloid catabolic enzymes including insulin-degrading enzyme (Hussain et al. 2007) and neprilysin (Poirier et al. 2006), neurotrophic factors such as cerebrolysin (Rockenstein et al. 2006), cholesterol metabolism (Li et al. 2006), and protein kinase and protease inhibitors (Hussain et al. 2007).. Limitations of using transgenic rodent models for AD Although transgenic rodent models have continuously improved during the last few years, to date there is still no perfect small animal model able to mimic perfectly both the pathological hallmarks and behaviour observed in AD patients. Despite the similarities described above, there are still crucial differences in neuropathology and behaviour between these rodent models and AD. The two major histopathological hallmarks crucial to the diagnosis of AD are Ab plaques and NFT, and these features have been reproduced in transgenic rodent models. The involvements of presenilins, ApoE4 and secretase activity have also been addressed. However, despite these advancements, no transgenic model to date has been able to accurately demonstrate a progressive loss of cholinergic neurons in the temporal and frontal lobes of the AD brain (Lewis et al. 2000). In the APP transgenic models, up to 14% neuronal loss has been reported in a single model, and neurodegeneration has been confined to the CA1 region of the hippocampus (Calhoun et al. 1998). A subsequent model (Tg2576) developed by Hsiao has recently shown age-related cognitive impairment in addition to Ab deposition (Hsiao et al. 1996). Moreover,. 187. as these models do not demonstrate significant neuronal loss, they are only useful for examining early stage AD. Also, the presentation of AD-related tauopathies has not been accurately modelled in any of the presently available tau transgenic models for AD, except in the motor neurons in the spinal cord (Lewis et al. 2000). As AD does not involve motor neurons, the therapeutic effects of these pathological features in AD are unwarranted. There are several possible reasons for the failure of successful rodent studies to develop neuronal loss in regions of the brain associated with AD. For instance, rodent neurons may be less susceptible to degeneration compared to human neurons. Alternative hypotheses relate the lack of neurodegeneration in transgenic rodent models to the short lifespan of these animals. The size of the rodent brain and the lack of cortex are likely to be one of the major issues in these discrepancies. Implanting human neurons into mouse brain prior to delivery of AD-associated transgenic genes has been suggested as a possible option to resolve this mystery. Moreover, the exact spatial distribution of AD pathology has so far not been recapitulated in any of the current transgenic rodent models for AD. As previously mentioned, the localisation of Ab plaques and NFT are dependent on the choice of promoter and integration site of the selected transgenic constructs in the model genome. This makes it almost impossible to compare different transgenic rodent models, since their genetic makeup strongly influences the onset histopathological features and cognitive decline observed in AD. As mentioned by Benatar in relation to ALS mouse models, the data obtained with these rodent models gets ‘‘lost in translation’’ (Benatar et al. 2006). Also, all the transgenic mice used in the studies mentioned in this review represent only a model of familial AD rather than sporadic AD. It might even be argued that these mice are only representatives of familial AD due to specific mutations in the APP, PS or Tau genes. Since the pathophysiology of AD is only partly understood, it is possible that early initiating neurodegenerative processes may differ between the sporadic and familial forms of AD. Therapeutic actions that may act on one form of the disease may not work on the other form. Another important factor is that, in all these rodent models, treatments are initiated often before any clinical onset of the disease. Of course, this strategy is not applicable to human AD as it is not yet possible to identify which subjects will develop AD. Finally, it should also be considered that drug dosage and bioavailability in rodent tissue cannot really be translated to human body if we consider that a mouse weighs about 4.106 times less than a human. In order to illustrate the problems of extrapolating outcomes in mice to clinical trials in humans, recent problems in vaccination trials. 123.
Figure
Documento similar
For example, in (Ju´lvez, Bemporad, Recalde, & Silva 2004) hybrid control techniques based on Mixed Logic Dynamical systems (Bemporad & Morari, 1999) are applied to
Incidence of dementia and probable Alzheimer´s disease in a general population: The Framingham study.. Hendiré H., Ogunniyi A.,
no obstante, cabe señalar en este sentido que los robots simuladores sí hacen uso de este rol dentro del género en cuanto a su comportamiento discursivo, magnificado en el texto
Our genome-wide transcriptome analyses revealed significant changes in transcriptional pathways related to neurotransmission, synaptic plasticity, learning/memory, and
Mice deficient for the telomerase component Terc and mice deficient for the shelterin component Trf1 represent good models to study telomere dysfunction.. Indeed, those mice show
Consequently, in this thesis we have generated two cellular models, a mouse model based on NURR1 overexpression in neural stem cells, and a human model based
‐ EMMAservice: Servicing the European Biomedical Research Community: archiving and dissemination of Mouse Models of Human Disease, European
User-dependent Hidden Markov Models A recent study [10] has proven the benefits of using user-dependent models by specifically setting the num- ber of states and Gaussian mixtures
