Regulation of pannexin and connexin channels and their functional role in skeletal muscles
7
0
0
Texto completo
(2) 2930. myogenin, which transform these cells into proliferative myoblasts and, subsequently, drive their continued differentiation [1]. A critical step that triggers the expression of these transcription factors is the increase in intracellular free calcium concentration that promotes activation of a great variety of protein kinases and phosphatases, including the calcium-dependent protein phosphatase calcineurin, which promotes the expression of myogenic factor 5 [2]. Therefore, identification of the mechanisms that lead to increases in intracellular free calcium concentration is a relevant issue to be solved and will be discussed in the present review. In this respect, the discovery of pannexin1 hemichannels (Panx1 HCs) in myotubes and adult myofibers opens new avenues to further understand several physiologic responses. In addition, new reports have demonstrated that denervated fast skeletal muscles express several nonselective membrane channels that might explain a crucial step between the lack of nerve continuity and muscle atrophy. These two findings are also described and discussed in this review. There are several other review papers describing the roles of Panx1 and connexin (Cx) proteins in skeletal muscle biology and pathology [3–6]. Here, we focus on the role of these channels in the commitment of myogenic precursors to the myogenic lineage, to their differentiation, fusion into myotubes and ultimate formation of mature skeletal muscle fibers. The function of these channels in muscle physiology and pathology, including recent advances linking such channels to calcium release through dihydropyridine and ryanodine receptors, is also discussed. Finally, advances regarding the participation of these channels in the rapid atrophy that follows paralysis by nerve transection will be reviewed.. Pannexin hemichannels in striate muscle ontogeny The increase in intracellular free calcium concentration required for initiating the expression of transcription factors involved in muscle differentiation could result from calcium diffusion from the extracellular milieu as well as from calcium release from intracellular stores. The former could be triggered by activation of store operated calcium entry channels, including calcium sensor stromal interaction molecule 1 and 2 channels and calcium releaseactivated calcium channel protein 1 channels [7, 8], and/or activation of purinergic P2 receptors (P2R) by adenosine diphosphate (ADP) and adenosine triphosphate (ATP) (Fig. 1). Calcium release from intracellular stores is mainly induced by inositol triphosphate, since ryanodine receptors are either inactive or absent [9]. Activation of P2XR2, P2XR4 and P2XR7 has also been shown to increase cellular membrane permeability to small molecules, such as Lucifer yellow, ethidium bromide and. 123. J. C. Sáez et al.. YO-PRO-1, in different cell types, including myoblasts [3, 10]. In macrophages, the increased permeability of the cell membrane to dyes induced by P2X7 receptor activation is abrogated by knockdown of Panx1 expression [11], indicating that Panx1 HCs mediate the dye uptake induced by P2X7 receptor activation. Nevertheless, recent arguments have been presented that inhibition of dye uptake by probenecid is independent of Panx HCs, suggesting that dye uptake induced by extracellular ATP acting on P2X7 receptor is independent of Panx1 HCs [12]. Thus, further studies are required to clarify this apparent controversy. Of interest, activation of Panx1 HCs has also been associated with activation of P2YRs by ATP [13]. Panxs are expressed by vertebrate cells and are orthologues of innexins found in invertebrates [14]. Panxs are membrane glycoproteins that form nonselective membrane HCs [15] and are encoded by three gene members, namely, Panx1, Panx2 and Panx3. Panx1 is the most ubiquitously expressed member of this family. Current available information indicates that Panx2 expression is rather restricted to few cell types and tissues, including brain [16] and T-cells [17]. Panx3 is preferentially expressed in bone and skin [16]. Several investigators have proposed that Panx1 does not form gap junction channels (GJC) because the glycosylation moiety would hinder stabilization of GJCs [18]. A few reports have demonstrated the formation of Panx1 GJCs in exogenous expression systems [19, 20]. Further studies are required to determine whether such GJCs participate in physiological cell-cell communication. Here, Panx1 channels will be referred to as Panx1 HCs. HCs connect the intracellular and extracellular compartments, allowing the diffusional passage of ions and small molecules, including ATP and glucose [21, 22]. Although numerous studies indicate that Panx1 HCs are permeable to ATP, these findings have been challenged by Ramanov and group, who described that Panx1 transfected in three different cell lines yields outward rectifying anion channels with negligible permeability to anions exceeding 250 Da, thus excluding ATP. Again, this apparent controversy could be clarified with more studies on permeability properties of Panx1 HCs [23]. Several lines of evidence indicate that Panx1 HCs function to switch myogenic progenitors from proliferation to myogenic differentiation as well as in subsequent fusion of such progenitors to form myotubes and, eventually, muscle fibers. Early studies in L6 cells, a cell line derived from rat myoblasts, treated with b-glycyrrhetinic acid, a blocker of Cx HCs, GJCs and Panx1 HCs [24], showed inhibition of myogenin and myogenic regulatory factor 4 expression as well as inhibition of the cell fusion process that leads to myotube formation [25]. In contrast, treatment with octanol, a blocker of Cx HCs, GJCs, but not of Panx1 HCs [11, 19], did not alter myogenesis [25]. These findings.
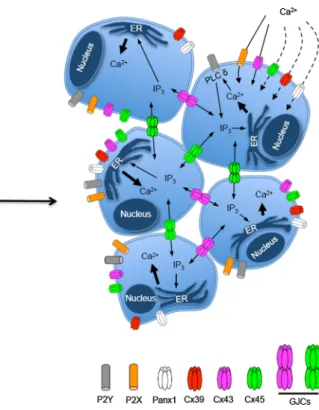
(3) Regulation of pannexin and connexin channels and their functional role in skeletal muscles. 2931. Fig. 1 Scheme of early myogenic event where Panx-based and Cxbased channels play a relevant role. In cells without acquisition of muscle commitment, Panx1 HCs and ionotropic and metabotropic purinergic P2X and P2Y receptors play a critical role controlling the increase in intracellular free calcium levels required for activation of intracellular signaling pathways that regulate the expression of transcription factors of the muscle differentiation process, such as myogenic differentiation 1 protein (MyoD). Activation of P2YRs by extracellular ATP released via Panx1 HCs leads to activation of phospholipase C d (PLC d) that generate inositol triphosphate (IP3),. which induce calcium from intracellular stores located in the endoplasmic reticulum (ER). Then, GJCs and Cx HCs located at cell interfaces and cells surface, respectively, participate in coordinating different metabolic responses that trigger differentiation. Since GJCs are permeable to IP3, they coordinate diverse metabolic responses among coupled cells. P2X7 receptor and Cx HCs are permeable to calcium and when activated allow calcium influx. The permeability to calcium of Cx39-based HCs and Cx45-based HCs as well as Panx1 HCs remains unknown. strongly suggest that Cx HCs and GJCs do not participate in the process of myoblast commitment and fusion during myogenesis. In line with this conclusion, Panx1 HCs activated by extracellular ATP were detected in C2C12 cells before commitment of acquisition to myogenesis [26]. Moreover, Panx1 HCs play a critical role in skeletal muscles progenitor differentiation (Fig. 1), since inhibition or knockdown of Panx1HCs drastically reduces the differentiation process, whereas overexpression of Panx1 enhances myogenic differentiation and inhibits proliferation [26, 27]. However, it remains puzzling that embryonic stem cell deletions of Panx1 exhibit no defects in myogenesis. Possibly, they express other channels that take over the role of Panx1 HC in the early steps of muscle differentiation. Myogenic differentiation is a multistep process that starts with commitment of cells of the myogenic lineage to myogenic fate characterized by expression of myogenin and myogenic differentiation 1 protein, and ends with cell fusion and innervation-induced changes. In order to specifically dissect the step in which Panx1 HCs play a. critical role in differentiation of striated muscles, undifferentiated C2C12 cells that are not yet committed to the myogenic lineage and that are negative for myogenic differentiation 1 protein, called reserve cells, were treated with Panx1 HC blockers or transfected with small interfering RNA directed against Panx1 mRNA, which completely prevented subsequent differentiation of these cells [26]. Results of these 2 experimental approaches and the fact that reserve cells do not express activatable Cx HCs when exposed to divalent cation-free solution, known to increase the open probability of Cx HCs [26], support the interpretation that functional Panx1 HCs and not Cx HCs are required for commitment of early myogenic precursor cells to the myogenic fate. At late stages of myogenesis, myotubes also express Panx1 HCs that allow ATP release induced by repetitive electrical stimulation [28]. In these cells, Panx1 seems to be closely associated to dihidropyridine and P2Y2 receptors, since they coprecipitate and this molecular triad seems to play an important role in modulating the intracellular calcium homeostasis [28].. 123.
(4) 2932. Connexin-based channels in myogenesis In addition to Panxs, myoblast express Cxs, which, at least after commitment to myogenic differentiation, have been shown to form GJCs that allow communication between myoblasts during differentiation [3]. Myoblasts also appear to express Cx HCs as has been found in most vertebrate cells studied to date [29]. The importance of Cx HCs in myogenesis has not been dissected yet. However, it is likely that GJCs are not required for acquisition of myogenic commitment, since it can occur in pluripotent cells without physical contact between them [26] (Fig. 1). Over the past four decades, the possible role of GJCs in myogenesis has been studied. Ultrastructural analyses revealed the presence of GJCs between myoblasts and between myoblasts and myotubes [30–32]. In addition, functional gap junctions have been found in developing myoblasts [31, 33]. More recently, Cx43 mRNA, but not Cx32 mRNA, was detected at high levels in cycling L6 myoblasts and Cx43 expression was shown to drastically decrease in these cells following fusion [25, 34]. Moreover, Cx43 has also been detected in prefusional C2C12 myoblasts [35] and transient expression of Cx40 was found in axial skeletal muscles of mouse embryos during myoblast fusion [36]. In addition, the application of 18b-glycyrrhetinic acid or octanol to L6 cells myoblasts undergoing differentiation blocked the expression of myogenin and myotube formation [25]. However, these compounds are not selective inhibitors of GJCs or HCs and 18b-glycyrrhetinic acid blocks Cx and Panx1 HCs. Thus, these findings should thus be interpreted with caution. Several recent studies have demonstrated that during myogenesis in development or regeneration, myoblasts or satellite cells express at least 3 other Cx species in addition to Cx40, including Cx39, Cx43 and Cx45 [36–41]. It has been possible to study the consequences of a lack of each of these Cxs during myogenesis. The absence of either Cx43 or Cx45 was found to delay the differentiation process in vitro or in vivo [37, 38]. However, Cx39 is expressed in murine myotubes, but not in myoblast cells or adult myofibers [41], suggesting that this Cx is involved in skeletal muscle differentiation, but not in the process of commitment to myogenic differentiation. In addition, the lack of Cx39 accelerates skeletal muscle differentiation [40]. Moreover, the reduced expression of myogenin, creatine kinase activity and the decrease in cell fusion found in myoblasts after induced deletion of Cx43 suggests a role of Cx43 in myoblast terminal differentiation [37]. Collectively, these studies provide relevant information on the role of each protein, but it is important to keep in mind that the absence of a particular Cx protein abrogates the formation of both GJCs and HCs, leaving open the question of. 123. J. C. Sáez et al.. whether both Cx-based channels or just one of them participate in myogenesis. Relevant to this issue, Cx39 does not form GJCs in exogenous expression systems [39], suggesting that for this channel, it is less likely that GJCs participate in the influence of Cx39 on myogenesis. It still remains to be determined if Cx39 forms functional HCs. In addition, the role of each protein might be independent of their capacity to form membrane channels, as has been shown in other systems [42].. Pannexin-based and connexin-based channels in adult skeletal muscles Cx expression and GJC formation is progressively lost during the late steps of myogenesis. After innervation, myofibers of most skeletal muscles do not contain GJCs [5]. Similarly, Cx HCs are also absent in adult innervated skeletal muscles of rodents [43]. However, the demonstration of Cx39.9 expression in slow twitch muscles of teleosts [44] suggests that lack of skeletal muscle Cx expression is not a general feature of all vertebrates. Normal adult muscles express Panx1, but not Panx2 or Panx3 [5, 22, 45]. Panx1 has been localized in transverse T-tubules, where it resides next to dihidropyridine receptors [5] (Fig. 2). Repetitive electrical stimulation induces Panx1 HC opening and subsequent ATP release. ATP released in this manner is vital to potentiation of the skeletal muscle contraction response [22]. It has been suggested that another function of ATP released via Panx1 HCs could be in skeletal muscle plasticity [45]. In support to these proposed roles of Panx1 HC-mediated ATP release, it has been shown that Panx1-/- muscles or muscles treated with Panx1 HC blockers do not show potentiation of the contraction response promoted by repetitive electrical stimulation [22]. In addition, the application of exogenous ATP to Panx1-/- muscles promotes potentiation of the contraction response [46]. Another important function of Panx1 HCs is that they are permeable to glucose and account for about 50 % of glucose uptake induced by repetitive electrical stimulation, while the other 50 % is taken over by glucose transporter type 4 [22]. Opening of Panx1 HCs can be induced by extracellular ATP or repetitive electrical stimulation, but the molecular mechanisms involved remain to be identified. One interesting finding is that Panx1 has several putative phosphorylation sites for different protein kinases, including protein kinase A, protein kinase C and calmodulindependent protein kinase type II, in its carboxyl-terminal tail. Accordingly, Panx1 was found to be a phosphoprotein and phosphorylation of serine and threonine residues is.
(5) Regulation of pannexin and connexin channels and their functional role in skeletal muscles. 2933. Fig. 2 Scheme showing the putative location of Panx1 HCs in the transverse T-tubule of adult skeletal muscles. Panx1 HCs are in close association with dihidropyridine receptors (DhpR), which in an alternate way are in close contact with the ryanodine receptor (RyR). a View in the bilipid membrane and b view in a monolipid membrane. c Perpendicular view of C (90° rotation). The putative localization of Panx1 HCs needs to be confirmed experimentally. enhanced during potentiation of muscle contraction induced by repetitive electrical stimulation [22]. Whether the increase in the phosphorylation status is the cause of activation of Panx1 HCs remains unknown. A recent unexpected finding was that denervated rodent skeletal muscles show de novo expression of several nonselective membrane channels, including Cx HCs, P2X7 receptors and transient receptor potential type V2 channels, and upregulation of Panx1 [43]. Deletion of only Cx43 and Cx45 was sufficient to drastically reduce muscle atrophy and to completely prevent activation of the transcription factor nuclear factor kB and to increase of mRNA levels of proinflammatory cytokines [43]. In addition, the upregulated expression of P2X7 receptor was almost completely blocked, suggesting that activation of its expression is downstream from Cx43 and Cx45 expression. Since several of these membrane channels, including Cx43 HCs, P2X7 receptors and transient receptor potential type V2 channels, are permeable to calcium, it has been proposed that they serve as membrane pathways for calcium influx that would activate intracellular metabolic pathways previously shown to participate in muscle wasting [43]. In contrast, denervation-induced muscle atrophy was not prevented by the lack of Panx1 expression in Panx1-/mice [43]. An additional interesting finding is that in spite of the Cx expression in denervated muscle, these cells do not form functional GJCs [43]. Other factors that promote the expression of Cx HCs in skeletal muscles are the proinflammatory cytokines tumor. necrosis factor a and interleukin 1b [5], which might be relevant in diverse conditions associated with inflammation that lead to muscle atrophy. Moreover, skeletal muscles of mice express Panx3 in response to high-fat diet, and the palmitate-induced expression and activity of Panx3 HCS in myotubes has been proposed to be mediated by Toll-like receptor 4 and nuclear factor kB [47]. Accordingly, Tolllike receptor 4 and nuclear factor kB inhibition or Panx3 knockdown prevents monocyte chemoattraction [47].. Conclusions and perspectives The accumulated evidence strongly suggests that Panx1 HCs are essential for myogenic fate commitment and that Cx-based channels play critical roles in the coordination and progression of subsequent steps required for myogenic differentiation and myogenesis. Additional work is needed to understand the relative importance of Cx GJCs and Cx HCs in these processes. Also, further studies are required to identify the phosphorylated amino acid residues of Panx1 as well as the protein kinases responsible for this posttranslational modification. It is clear that either repetitive electrical stimulation or extracellular ATP leads to rapid opening of Panx HCs in the sarcolemma. However, several questions remain to be answered, among which (1) is the increase in phosphorylation state of Panx1 HCs responsible for the enhanced activity of this channel in the sarcolemma? (2) Is the. 123.
(6) 2934. increase in Panx1 HC activity seen during potentiation of muscle contraction due to increase in opening probability of available channels in the cell membrane or is due to recruitment of more Panx1 HCs to the sarcolemma? (3) What mechanisms restore the activity of Panx1 HCs to the basal state? Similarly, the atrophy observed in adult denervated or inflamed skeletal muscles can be largely prevented in muscles that do not express Cx43 and Cx45. Is this a mechanism that obeys a hierarchical expression of calcium-permeable channels or, instead, might all of them be involved such that each contribute to reach a critical increase in intracellular free calcium concentration? In general, the field of Cx-based and Panx-based channel research is rather limited, but offers numerous relevant questions to be answered to further increase our knowledge in skeletal muscle physiology and pathophysiology. Moreover, Cx HCs might be potential molecular targets to reduce muscle atrophy in diverse circumstances. The mechanisms by which expression of sarolemmal Cx HCs is repressed in innervated and noninflamed muscles remains unknown. In addition, the development of potent and selective Cx HC blockers might be useful to reduce muscle degeneration induced by spinal cord injury and many other pathological conditions in which the integrity of nerve muscle contact and functional regulation has been altered. Acknowledgments This work was partially funded by FONDECYT grants 1150291 (to Juan C. Sáez), ICM-Economı́a P09-022-F Centro Interdisciplinario de Neurociencias de Valparaı́so (to Juan C. Sáez) and the Department of Veterans Affairs Rehabilitation Research and Development Service (B9212C) and the James J. Peters VA Medical Center. Bruno A. Cisterna and Anibal Vargas acknowledge the support of a CONICYT fellowship. Conflict of interest The authors declare that the research was conducted in the absence of any commercial or financial relationship that could be construed as a potential conflict of interest.. References 1. Charge SB, Rudnicki MA (2004) Cellular and molecular regulation of muscle regeneration. Physiol Rev 84(1):209–238. doi:10.1152/physrev.00019.2003 2. Friday BB, Pavlath GK (2001) A calcineurin- and NFAT-dependent pathway regulates Myf5 gene expression in skeletal muscle reserve cells. J Cell Sci 114(Pt 2):303–310 3. Araya R, Riquelme MA, Brandan E, Saez JC (2004) The formation of skeletal muscle myotubes requires functional membrane receptors activated by extracellular ATP. Brain Res Brain Res Rev 47(1–3):174–188. doi:10.1016/j.brainresrev.2004. 06.003 4. Casas M, Buvinic S, Jaimovich E (2014) ATP signaling in skeletal muscle: from fiber plasticity to regulation of metabolism. Exerc Sport Sci Rev 42(3):110–116. doi:10.1249/JES.000000 0000000017 5. Cea LA, Riquelme MA, Cisterna BA, Puebla C, Vega JL, Rovegno M, Saez JC (2012) Connexin- and pannexin-based. 123. J. C. Sáez et al.. 6.. 7.. 8.. 9.. 10.. 11.. 12.. 13.. 14.. 15.. 16.. 17.. 18.. 19.. 20.. channels in normal skeletal muscles and their possible role in muscle atrophy. J Membr Biol 245(8):423–436. doi:10.1007/ s00232-012-9485-8 Cisterna BA, Cardozo C, Saez JC (2014) Neuronal involvement in muscular atrophy. Front Cell Neurosci 8:405. doi:10.3389/ fncel.2014.00405 Darbellay B, Arnaudeau S, Ceroni D, Bader CR, Konig S, Bernheim L (2010) Human muscle economy myoblast differentiation and excitation-contraction coupling use the same molecular partners, STIM1 and STIM2. J Biol Chem 285(29):22437–22447. doi:10.1074/jbc.M110.118984 Darbellay B, Arnaudeau S, Konig S, Jousset H, Bader C, Demaurex N, Bernheim L (2009) STIM1- and Orai1-dependent store-operated calcium entry regulates human myoblast differentiation. J Biol Chem 284(8):5370–5380. doi:10.1074/jbc. M806726200 Antigny F, Konig S, Bernheim L, Frieden M (2014) Inositol 1,4,5 trisphosphate receptor 1 is a key player of human myoblast differentiation. Cell Calcium 56(6):513–521. doi:10.1016/j.ceca. 2014.10.014 Ryten M, Dunn PM, Neary JT, Burnstock G (2002) ATP regulates the differentiation of mammalian skeletal muscle by activation of a P2X5 receptor on satellite cells. J Cell Biol 158(2):345–355. doi:10.1083/jcb.200202025 Pelegrin P, Surprenant A (2006) Pannexin-1 mediates large pore formation and interleukin-1beta release by the ATP-gated P2X7 receptor. EMBO J 25(21):5071–5082. doi:10.1038/sj.emboj. 7601378 Bhaskaracharya A, Dao-Ung P, Jalilian I, Spildrejorde M, Skarratt KK, Fuller SJ, Sluyter R, Stokes L (2014) Probenecid blocks human P2X7 receptor-induced dye uptake via a pannexin-1 independent mechanism. PLoS One 9(3):e93058. doi:10.1371/ journal.pone.0093058 Locovei S, Wang J, Dahl G (2006) Activation of pannexin 1 channels by ATP through P2Y receptors and by cytoplasmic calcium. FEBS Lett 580(1):239–244. doi:10.1016/j.febslet.2005. 12.004 Ambrosi C, Gassmann O, Pranskevich JN, Boassa D, Smock A, Wang J, Dahl G, Steinem C, Sosinsky GE (2010) Pannexin1 and Pannexin2 channels show quaternary similarities to connexons and different oligomerization numbers from each other. J Biol Chem 285(32):24420–24431. doi:10.1074/jbc.M110.115444 Bruzzone R, Barbe MT, Jakob NJ, Monyer H (2005) Pharmacological properties of homomeric and heteromeric pannexin hemichannels expressed in Xenopus oocytes. J Neurochem 92(5):1033–1043. doi:10.1111/j.1471-4159.2004.02947.x Baranova A, Ivanov D, Petrash N, Pestova A, Skoblov M, Kelmanson I, Shagin D, Nazarenko S, Geraymovych E, Litvin O, Tiunova A, Born TL, Usman N, Staroverov D, Lukyanov S, Panchin Y (2004) The mammalian pannexin family is homologous to the invertebrate innexin gap junction proteins. Genomics 83(4):706–716. doi:10.1016/j.ygeno.2003.09.025 Shoji KF, Saez PJ, Harcha PA, Aguila HL, Saez JC (2014) Pannexin1 channels act downstream of P2X 7 receptors in ATPinduced murine T-cell death. Channels 8(2):142–156. doi:10. 4161/chan.28122 Sosinsky GE, Boassa D, Dermietzel R, Duffy HS, Laird DW, MacVicar B, Naus CC, Penuela S, Scemes E, Spray DC, Thompson RJ, Zhao HB, Dahl G (2011) Pannexin channels are not gap junction hemichannels. Channels 5(3):193–197 Bruzzone R, Hormuzdi SG, Barbe MT, Herb A, Monyer H (2003) Pannexins, a family of gap junction proteins expressed in brain. Proc Natl Acad Sci USA 100(23):13644–13649. doi:10.1073/ pnas.2233464100 Sahu G, Sukumaran S, Bera AK (2014) Pannexins form gap junctions with electrophysiological and pharmacological.
(7) Regulation of pannexin and connexin channels and their functional role in skeletal muscles. 21.. 22.. 23.. 24.. 25.. 26.. 27.. 28.. 29. 30.. 31. 32.. 33. 34.. properties distinct from connexins. Scientific reports 4:4955. doi:10.1038/srep04955 Locovei S, Bao L, Dahl G (2006) Pannexin 1 in erythrocytes: function without a gap. Proc Natl Acad Sci USA 103(20):7655–7659. doi:10.1073/pnas.0601037103 Riquelme MA, Cea LA, Vega JL, Boric MP, Monyer H, Bennett MV, Frank M, Willecke K, Saez JC (2013) The ATP required for potentiation of skeletal muscle contraction is released via pannexin hemichannels. Neuropharmacology 75:594–603. doi:10. 1016/j.neuropharm.2013.03.022 Romanov RA, Bystrova MF, Rogachevskaya OA, Sadovnikov VB, Shestopalov VI, Kolesnikov SS (2012) The ATP permeability of pannexin 1 channels in a heterologous system and in mammalian taste cells is dispensable. J Cell Sci 125(Pt 22):5514–5523. doi:10.1242/jcs.111062 Schalper KA, Palacios-Prado N, Orellana JA, Saez JC (2008) Currently used methods for identification and characterization of hemichannels. Cell Commun Adhes 15(1):207–218. doi:10.1080/ 15419060802014198 Proulx AA, Lin ZX, Naus CC (1997) Transfection of rhabdomyosarcoma cells with connexin43 induces myogenic differentiation. Cell Growth Differ 8(5):533–540 Riquelme MA, Cea LA, Vega JL, Puebla C, Vargas AA, Shoji KF, Subiabre MJCS (2015) Pannexin channels mediate the acquisition of myogenic commitment in C2C12 reserve cells promoted by P2 receptor activation. Front Cell Dev Biol 3(25):1–11. doi:10.3389/fcell.2015.00025 Langlois S, Xiang X, Young K, Cowan BJ, Penuela S, Cowan KN (2014) Pannexin 1 and pannexin 3 channels regulate skeletal muscle myoblast proliferation and differentiation. J Biol Chem 289(44):30717–30731. doi:10.1074/jbc.M114.572131 Buvinic S, Almarza G, Bustamante M, Casas M, Lopez J, Riquelme M, Saez JC, Huidobro-Toro JP, Jaimovich E (2009) ATP released by electrical stimuli elicits calcium transients and gene expression in skeletal muscle. J Biol Chem 284(50): 34490–34505. doi:10.1074/jbc.M109.057315 Saez JC, Leybaert L (2014) Hunting for connexin hemichannels. FEBS Lett 588(8):1205–1211. doi:10.1016/j.febslet.2014.03.004 Duxson MJ, Usson Y, Harris AJ (1989) The origin of secondary myotubes in mammalian skeletal muscles: ultrastructural studies. Development 107(4):743–750 Kalderon N, Epstein ML, Gilula NB (1977) Cell-to-cell communication and myogenesis. Journal Cell Biol 75(3):788–806 Rash JE, Staehelin LA (1974) Freeze-cleave demonstration of gap junctions between skeletal myogenic cells in vivo. Dev Biol 36(2):455–461 Schmalbruch H (1982) Skeletal muscle fibers of newborn rats are coupled by gap junctions. Dev Biol 91(2):485–490 Balogh S, Naus CC, Merrifield PA (1993) Expression of gap junctions in cultured rat L6 cells during myogenesis. Dev Biol 155(2):351–360. doi:10.1006/dbio.1993.1034. 2935. 35. Constantin B, Cronier L (2000) Involvement of gap junctional communication in myogenesis. Int Rev Cytol 196:1–65 36. Dahl E, Winterhager E, Traub O, Willecke K (1995) Expression of gap junction genes, connexin40 and connexin43, during fetal mouse development. Anat Embryol (Berl) 191(3):267–278 37. Araya R, Eckardt D, Maxeiner S, Kruger O, Theis M, Willecke K, Saez JC (2005) Expression of connexins during differentiation and regeneration of skeletal muscle: functional relevance of connexin43. J Cell Sci 118(Pt 1):27–37. doi:10.1242/jcs.01553 38. Araya R, Eckardt D, Riquelme MA, Willecke K, Saez JC (2003) Presence and importance of connexin43 during myogenesis. Cell Commun Adhes 10(4–6):451–456 39. von Maltzahn J, Euwens C, Willecke K, Sohl G (2004) The novel mouse connexin39 gene is expressed in developing striated muscle fibers. J Cell Sci 117(Pt 22):5381–5392. doi:10.1242/jcs. 01413 40. von Maltzahn J, Wulf V, Matern G, Willecke K (2011) Connexin39 deficient mice display accelerated myogenesis and regeneration of skeletal muscle. Exp Cell Res 317(8):1169–1178. doi:10.1016/j.yexcr.2011.01.017 41. von Maltzahn J, Wulf V, Willecke K (2006) Spatiotemporal expression of connexin 39 and -43 during myoblast differentiation in cultured cells and in the mouse embryo. Cell Commun Adhes 13(1–2):55–60. doi:10.1080/15419060600631508 42. Zhou JZ, Jiang JX (2014) Gap junction and hemichannel-independent actions of connexins on cell and tissue functions—an update. FEBS Lett 588(8):1186–1192. doi:10.1016/j.febslet.2014. 01.001 43. Cea LA, Cisterna BA, Puebla C, Frank M, Figueroa XF, Cardozo C, Willecke K, Latorre R, Saez JC (2013) De novo expression of connexin hemichannels in denervated fast skeletal muscles leads to atrophy. Proc Natl Acad Sci USA 110(40):16229–16234. doi:10.1073/pnas.1312331110 44. Hirata H, Wen H, Kawakami Y, Naganawa Y, Ogino K, Yamada K, Saint-Amant L, Low SE, Cui WW, Zhou W, Sprague SM, Asakawa K, Muto A, Kawakami K, Kuwada JY (2012) Connexin 39.9 protein is necessary for coordinated activation of slowtwitch muscle and normal behavior in zebrafish. J Biol Chem 287(2):1080–1089. doi:10.1074/jbc.M111.308205 45. Jorquera G, Altamirano F, Contreras-Ferrat A, Almarza G, Buvinic S, Jacquemond V, Jaimovich E, Casas M (2013) Cav11 controls frequency-dependent events regulating adult skeletal muscle plasticity. J Cell Sci 126(5):1189–1198. doi:10.1242/jcs. 116855 46. Cea LA, Riquelme MA, Vargas AA, Urrutia C, Saez JC (2014) Pannexin 1 channels in skeletal muscles. Front Physiol 5:139. doi:10.3389/fphys.2014.00139 47. Pillon NJ, Li YE, Fink LN, Brozinick JT, Nikolayev A, Kuo MS, Bilan PJ, Klip A (2014) Nucleotides released from palmitatechallenged muscle cells through pannexin-3 attract monocytes. Diabetes 63(11):3815–3826. doi:10.2337/db14-0150. 123.
(8)
Figure
Documento similar