Efecto de la implementación de un sistema de amamantamiento restringido sobre el balance energético y desempeño reproductivo posparto en ovejas de pelo
53
0
0
Texto completo
(2) EFECTO DE LA IMPLEMENTACIÓN DE UN SISTEMA DE AMANTAMIENTO RESTRINGIDO SOBRE EL BALANCE ENERGÉTICO Y DESEMPEÑO REPRODUCTIVO POSPARTO EN OVEJAS DE PELO.. PRESENTADO POR: JEFFREY RODRIGO CARVAJAL BASTO ID: 265871 JOAQUIN HUMBERTO VARGAS MUÑOZ ID: 102042. UNIVERSIDAD COOPERATIVA DE COLOMBIA FACULTAD DE MEDICNA VETERINARIA Y ZOOTECNIA BUCARAMANGA 2016.
(3) CONTENIDO. INTRODUCCIÓN ..................................................................................................... 7 1. OBJETIVO GENERAL....................................................................................... 9 1.1. Objetivos específicos .................................................................................. 9. 2. MARCO TEORICO .......................................................................................... 10 2.1. Regulación del desarrollo folicular y ovulación de la hembra ovina. ......... 10. 2.2. Función de las gonadotropinas. ................................................................ 14. 2.3. Restablecimiento de la actividad ovárica después del parto..................... 18. 2.4. Metabolismo energético de ovejas en el periparto .................................... 22. 2.5. Condición corporal .................................................................................... 25. 2.6. Destete de corderos ................................................................................. 26. 3. METODOLOGÍA .............................................................................................. 31 3.1. Grupos experimentales ............................................................................. 32. 3.1.1 Control (GC): ............................................................................................ 32 3.1.2 Grupo 1 (G1):............................................................................................ 32 3.1.3 Grupo 2 (G2):............................................................................................ 32 3.2. Evaluación de la condición corporal y pesaje de los animales ................. 33. 3.3. Evaluación del peso al destete de los corderos. ....................................... 33. 3.4. Evaluación del balance energético negativo. ............................................ 33. 3.5. Evaluación del intervalo parto-primer celo ................................................ 34. 4. RESULTADOS Y DISCUSIÓN ........................................................................ 36 4.1. Evaluación de la condición corporal y el pesaje de los animales. ............. 36. 4.1.1 Condición Corporal. .................................................................................. 36 4.1.2 Evaluación de peso. ................................................................................. 39 4.2. Peso promedio de las crías. ..................................................................... 41. 4.3. Evaluación de intervalo parto – primer celo. ............................................. 43. 5. CONCLUCIONES ........................................................................................... 46 6. RECOMENDACIÓN ........................................................................................ 47.
(4) 7. BIBLIOGRAFÍA ............................................................................................... 48.
(5) RESUMEN. La finalidad de este estudio fue evaluar el efecto de la implementación de un sistema de amamantamiento restringido sobre el balance energético y desempeño reproductivo post-parto de ovejas de pelo. Se utilizaron 23 ovejas distribuidas en tres grupos, el primer grupo consta de 7 ovejas siendo el GC: amamantamiento sin restricción, el G1 con 8 ovejas restringido la lactancia a partir día 3 post parto, el G2 con 8 ovejas restringido la lactancia partir de los 15 días post parto; las crías que fueron sometidas a los tratamiento. de restricción, permanecían. con las. hembras por dos periodos de 30 minutos al día. Las ovejas se encontraban en un sistema de pastoreo extensivo con pastos nativos de la región Cynodon plectostachium y Brachiaria sp, las crías se les suministro una suplementación con gallinero. Se le dio seguimiento semanal del peso y la condición corporal de las ovejas, las crías fueron pesadas al momento de nacer y el destete (90 días). Se realizaron tomas de muestras de sangre a las hembras ovinas, para pruebas metabólicas una vez por semana, desde la 1ª semana hasta la 5ª semana posparto, y quincenalmente a partir de la 6ª hasta las 12ª semana de lactancia. Durante este proceso a las hembras se les detectó el primer celo post-parto, mediante observación directa en horas de la mañana y tarde, para la cual fueron utilizados tres machos enteros con chaleco marcador, con el objetivo de determinar si el celo detectado en las ovejas fue seguido de ovulación, fueron determinadas las concentraciones de progesterona en el suero sanguíneo. Para.
(6) este propósito se colectarán muestras de sangre a través de venopunción de la vena yugular a cada 3 días desde el día del celo hasta el día 15 posterior a este. Posteriormente se realizó el diagnostico de gestación (45 días post servicio), mediante ultrasonografía transrectal.. Al implementar la restricción de amamantamiento de las ovejas de pelo de raza criolla a las cuales se les restringió la lactancia a periodos de 30 minutos al día, se obtuvo como resultado que las hembras del grupo experimental Gc promediaron una CC de 3,5 puntos, en el peso de las hembras los grupos experimentales Gc y G2 promediaron 37 kg y 33 kg respectivamente, en el periodo de días abiertos el grupo experimental G2 promedio 78 días y el grupo experimental G1 promedio 96 días. Esto demostró que la implementación de un sistema de amamantamiento restringido podría disminuir los días abiertos con relación a la primera ovulación post parto, sin alterar significativamente el peso de las hembras y de sus crías, donde posiblemente sería un sistema apto para la implementación del manejo post parto en las granjas ovinas..
(7) INTRODUCCIÓN. Actualmente en Colombia, de forma similar a lo que ocurre en otras regiones con sistemas de producción desarrollados en climas tropicales, la ovino-cultura no contribuye significativamente a la economía nacional, pero desempeña un importante papel social, en los recursos económicos y alimenticios de los campesinos de las zonas marginales. La producción ovina nacional se caracteriza por estar sustentada, en su gran mayoría, por sistemas de explotación extensivos con escaso nivel tecnológico (Castellano Mendez & C, 2010). En la actualidad, la información al respecto de las características productivas y reproductivas de los rebaños nacionales es escasa, y consecuentemente, los sistemas de producción ovina carecen de estrategias de manejo adecuadas que ayuden a sortear los principales problemas productivos que las aquejan, ya que implementando nuevas alternativas y tecnologías se podría generar un nuevo mercado a nivel internacional debido a la facilidad y rusticidad de los ovinos para los terrenos de estas regiones.. El conocimiento de los indicadores reproductivos es requisito esencial para el sostenimiento eficiente de los sistemas de producción animal. Los índices reproductivos afectan directamente la productividad de las explotaciones y esta relación se hace más evidente en determinados periodos del ciclo productivo (Stellfug JN & JW, 2001). La fase que sigue al parto es un periodo crítico para la.
(8) productividad de los hatos ovinos, es durante este período que las hembras cesan su actividad reproductiva y de esta manera la continuidad de la multiplicación del rebaño se detiene.. El estado nutricional de las hembras en el periodo conocido como periparto y el estímulo generado por el acto de amamantar, han sido señalados como los dos factores que en mayor medida tienen influencia en la duración del anestro posparto (RD, 1990). Buscando la implementación de nuevas alternativas y el mejoramiento de los procesos productivos, se hace necesario sean estudiados los beneficios que produciría la implementación del amamantamiento restringido en las razas ovinas del trópico de pelo corto, evitándose de ésta manera la presencia de anestro prolongados en hembras de cría que produzcan un retraso en la recuperación posparto de las hembras, situación que puede mejorar mediante la implementación del amamantamiento restringido propuesto. Con el propósito también de evaluar condición corporal y rendimientos en la ganancias de peso. La implementación del amamantamiento restringido consistió en limitar la periodo e intensidad del amamantamiento lo cual es una alternativa para atenuar los efectos negativos del anestro de las ovejas cría de pelo corto, convirtiéndose en opción viable de bajo costo para mejorar el desempeño reproductivo de las ovejas, contribuyendo así, al aumento de la eficiencia productiva de los rebaños del país..
(9) 1. OBJETIVO GENERAL. Evaluar el efecto de la implementación de un sistema de amamantamiento restringido sobre el balance energético y desempeño reproductivo posparto de ovejas de pelo.. 1.1 Objetivos específicos. . Evaluar el efecto del sistema de amamantamiento restringido sobre el intervalo parto- primer celo, el intervalo parto- ovulación y los días abiertos (intervalo parto- preñez) de ovejas de pelo.. . Evaluar el efecto del sistema de amamantamiento restringido el peso de las crías al destete, la ganancia diaria de peso y la condición corporal de las ovejas de pelo..
(10) 2. MARCO TEORICO. 2.1 Regulación del desarrollo folicular y ovulación de la hembra ovina.. Durante los periodos de actividad reproductiva, las ovejas manifiestan ciclos sexuales con una duración media de 17 días, de los cuales aproximadamente 3036 horas corresponden al periodo de receptividad sexual.. Una vez restaurada la actividad cíclica después de un periodo de anestro, ya sea la etapa pre-púber, el anestro postparto, el hipotálamo mediante su secreción de GnRH tiene una capacidad de acción, tanto de generar pulsos frecuentes (horarios) de LH en la hipófisis, como de activar la descarga básica de LH, la cual es responsable de la ovulación (López Sebastián A. S., 1993).. El primer ciclo ovárico después de un periodo de anestro no va precedido de síntomas de celo, y tanto su duración como la secreción de progesterona por parte del cuerpo Iúteo, son inferiores a lo normal. Suelen ser necesarias una o dos de estas ovulaciones antes de que aparezca la primera receptividad sexual por lo tanto, en el comienzo de la fertilidad. Aunque ha sido ampliamente puesta en evidencia la regulación de la dinámica del desarrollo y crecimiento de los folículos.
(11) por parte de las gonadotropinas, el papel de cada una de ellas (FSH y LH), durante todo el periodo de la foliculogénesis aún no es del todo comprensible.. En las primeras observaciones de las descargas de FSH y LH, a lo largo del ciclo estral de la oveja, se puede apreciar como las concentraciones plasmáticas de FSH, tienen un brusco, incremento comenzando unas horas después del inicio del celo (alrededor de cuatro horas), (Bazer, 1989) seguido de un incremento posterior, aproximadamente 24 horas después, y que coincide con el momento de la ovulación. Este segundo incremento fue considerado el responsable del reclutamiento y selección de los folículos que han de ovular en el próximo ciclo (Cahill, 1980).. En el caso de la LH sólo se observa una descarga que coincide con la primera descarga de FSH, la cual es responsable de la ovulación, apareciendo ésta 20-26 horas después del comienzo de la liberación de LH (Bartlewski, 2000). Sin embargo, la secreción pulsátil de LH, varía a lo largo del ciclo, en términos de variaciones de la frecuencia de esos pulsos, cuando son más altas las concentraciones de progesterona, los pulsos de LH aparecen con una frecuencia menor de tres horas. Sin embargo, a partir de la luteolisis, el día 15 del ciclo, los pulsos de LH se incrementan hasta alcanzar una frecuencia horaria, siendo este.
(12) incremento de los pulsos el responsable de la maduración final del folículo y formación del folículo preovulatorio.. La FSH, por el contrario, no presenta una secreción pulsátil, observándose tan sólo una disminución de su secreción durante la fase folicular, previamente a la aparición del celo a pesar de que desde hace años, gracias a la enorme sensibilidad y exactitud de las técnicas de análisis radio-inmunológicas, son bien conocidos los niveles plasmáticos de ambas gonadotropinas, el papel concreto de la FSH y LH en el control de la formación del folículo preovulatorio no está bien determinado (López Sebastián A. S., 1993). Se señala que la FSH parece ser la responsable del crecimiento del folículo, y durante su fase de reducción de las concentraciones plasmáticas en la fase folicular se produce la selección del folículo o folículos destinados a ovular. Por otro lado, la actividad esteroidogénica del folículo es responsabilidad de la LH, la cual controla la secreción de andrógenos por parte de las células de la teca folícular, que a su vez son transformados a estradiol, gracias a la actividad aromatasa inducida por la FSH, en las células de la granulosa (Acosta, 1983).. La FSH es la responsable de la selección del folículo preovulatorio, mientras que la LH tendría un efecto posterior en la maduración final del mismo. La FSH como.
(13) hormona clave en la recuperación y selección de folículos (López Sebastián A. S., 1993).. Crecimiento y selección folicular. La foliculogénesis se puede definir como un proceso de crecimiento y desarrollo de los folículos, desde el "pool" de folículos primordiales, hasta el estadio de preantral y antral, culminando con la atresia y rara vez con la ovulación. A partir del día 70 de la gestación en la oveja, el ovario del feto mantiene ya folículos en fase de crecimiento, estando presentes todos los folículos primordiales en el momento del nacimiento. De estos folículos primordiales, más del 99,9% nunca llega a ovular, pasando de la fase de crecimiento a la atresia (Jolly, 1994).. Los folículos preovulatorios están presentes a lo largo de todas las fases del ciclo estral y durante los periodos de anestro y son siempre equivalentes en número a la tasa de ovulación correspondiente a cada raza (Cahill, 1980). Las fases de desarrollo folicular incluyen la recuperación del folículo primordial e incorporación a la fase de crecimiento, acontecimiento que no es gonadotropo-dependiente, interviniendo factores intraováricos de crecimiento (algunos péptidos intraováricos identificados como tales: IGFs, EGF, TGF, FGFs, etc). Pasan después a un periodo a través del cual los folículos alcanzan un diámetro de 2mm, cuando son plenamente dependientes de las gonadotropinas, con una máxima concentración.
(14) de receptores para la FSH en la granulosa y para la LH en las células tecales (López Sebastián A. S., 1993).. En la fase de selección, durante la cual algunos folículos pasan al crecimiento y otros a la atresia, se establece por los más desarrollados una fase de dominancia, en la cual mantiene un sistema de defensa frente a la atresia. Los folículos comienzan a formar el antrum a partir de los 0,2mm de diámetro, siendo muy rápido el crecimiento final, ya que pasan en unos 5 días de 0,5 a 2mm, y es a partir de este diámetro cuando pueden ser seleccionados para ovular a partir del momento de la luteolisis, alcanzando el tamaño preovulatorio, a partir de 6mm, en 3-4 días (McNeilly, 1984).. 2.2 Función de las gonadotropinas.. En las diferentes fases del desarrollo folicular las gonadotropinas juegan un papel fundamental, aunque no por igual en todas las fases, incluso algunas de ellas aparentan ser gonadotropo-independientes. La total retirada de gonadotropinas de.
(15) la circulación sanguínea mediante la hipofisectomía, resulta en un fallo de la formación de folículos antrales (Dufour, 1979).. Durante la fase folicular del ciclo estral en la oveja, cuando se produce la selección del folículo que va a ovular, coinciden tres aspectos de notable interés: . Se ha producido la luteolisis.. . Esta lleva consigo el incremento de la pulsatilidad de Ia LH.. . Hay una disminución de la secreción de FSH.. El mecanismo de inhibición del crecimiento de los folículos subordinados que ejerce el dominante, parece que se lleva a cabo por dos vías. En el líquido folicular del folículo dominante, se ha encontrado un factor no esteroide, una glicoproteína (inhibina) sintetizada por las células de la granulosa. Esta glicoproteína capaz de inhibir el desarrollo folicular se pensaba desde hace tiempo que debía actuar a través de canales sistémicos, no intraováricos. Se ha demostrado que actúa directamente sobre la hipófisis, inhibiendo la secreción de FSH, sin que se hayan demostrado relaciones intraováricas entre folículos (Acosta, 1983).. El segundo mecanismo de inhibición del folículo dominante, basado en su secreción de estradiol. Esta inhibición, la realiza sin embargo vía hipotálamo.
(16) mediante un feed-back negativo que ejerce. Al desaparecer el folículo dominante, se produce una nueva descarga de FSH, con la formación de un nuevo folículo dominante. Este mayor acumulo de estradiol en este folículo, le permite además mantenerle libre de atresia por la acción que tuviera su propia inhibina (Karsch, 1980). A pesar de la depresión de FSH durante la fase folicular, el folículo durante este periodo sigue siendo plenamente dependiente de la acción de esta gonadotropina.. Ovulación y cuerpo lúteo. La fase folicular, coincidiendo con el momento del celo, el folículo preovulatorio llega a su máximo tamaño y presenta una enorme capacidad de síntesis de hormonas esteroides. El estradiol, el estrógeno más activo secretado por la granulosa del folículo, alcanza al igual que la androstenodiona su concentración máxima aproximadamente 24 horas antes del comienzo del celo. Coincidiendo con el incremento de los niveles de LH, cuyos pulsos aumentan su frecuencia a partir de la luteolisis, y que por medio de un feed-back positivo con el hipotálamo provocan la descarga preovulatoria de LH. Esta descarga aparece pocas horas después del comienzo del celo, siendo la responsable de la ovulación, que aparece entre veinte y veinticuatro horas después, junto con la formación del cuerpo Iúteo. El incremento en la frecuencia de pulsos de LH a partir de la luteolisis y la descarga preovulatoria de LH poco después del comienzo del celo producen cambios en la estructura del folículo que inducen la ovulación y posterior formación del cuerpo Iúteo. Estos cambios.
(17) producen una pérdida de la organización compacta de las células de la granulosa, con desaparición de la lámina basal y gran producción de mucopolisacáridos (Hay, 1977).. El cuerpo Iúteo se forma fundamentalmente a partir de las células de la granulosa y de la teca del folículo que se ha roto en el momento de la ovulación, dando lugar a las futuras células luteales grandes y pequeñas, respectivamente, que se encuentran en el mismo número que las foliculares pero aumentan su tamaño (O'Shea, 1979). La LH, principal hormona luteotrófica en los rumiantes (Niswender, 1985), provoca la síntesis de progesterona por parte de las células luteales, principalmente las pequeñas, con mayor número de receptores a la LH. Las concentraciones de progesterona comienzan a ser detectables en sangre periférica a partir de los tres días del celo. Esta hormona es la responsable de la gestación, relajación del Útero y reducción de la excitabilidad del endometrio. Otras hormonas producidas por el cuerpo Iúteo, y de gran interés, son la relaxina y la oxitocina.. Su perfil de concentraciones plasmáticas es similar al de la progesterona, con concentraciones máximas los días 14 Y del ciclo (Flint, 1983)..
(18) 2.3 Restablecimiento de la actividad ovárica después del parto.. Uno de los parámetros que de forma más directa incide sobre los rendimientos reproductivos, es el intervalo entre el parto y la primera ovulación fértil. La duración de este intervalo condiciona de manera directa el número de corderos por oveja y año, y finalmente delimita los márgenes de producción de una determinada explotación. Este intervalo, como cualquier otro en términos de eficacia. reproductiva,. es. totalmente. dependiente. de. múltiples. factores,. principalmente la interacción entre la época del año en que se produce el parto y la condición corporal en ese momento, junto con la variabilidad individual, nutricional y de manejo. La interacción entre la lactación y la época del año es decisiva en la determinación del periodo parto-primera ovulación fértil (Restall, 1977).. La condición corporal en el momento del parto juega un papel de gran importancia en el restablecimiento de la actividad reproductiva, ya que el estado nutritivo de estas hembras puede variar el intervalo a la primera ovulación desde 1 a 6 meses (López Sebastián A. S., 1993)..
(19) Durante la lactación los niveles de prolactina están notablemente aumentados en la oveja, debido al estímulo que provoca la succión en su secreción. En las oveja se ha llegado a demostrar que la prolactina no es la responsable de la inhibición de la ovulación durante la época de luz creciente o durante la lactación, situaciones en las cuales los niveles circulantes son los más altos (Brunet, 1991). Sin embargo, resulta de gran interés la observación de como los niveles de esta hormona. evolucionan. en. combinación. con. estos. dos. factores.. Los. glucocorticoides, concretamente el cortisol, hormona a la también se le ha atribuido un cierto papel en los mecanismos inhibitorios de la ovulación, sobre todo por su acción sobre la pulsatibilidad de la LH (Dufour, 1979), se produce igualmente un pico en la concentración plasmática similar al de prolactina, después del parto está además condicionada por el estado uterino, principalmente por su capacidad de regresión, siendo en muchos casos la limitante en el intento de fecundaciones tempranas después del parto aunque el ovario haya recuperado la capacidad ovulatorio (Sebastian, 1988).. Efecto del amamantamiento sobre el anestro posparto. Desde el punto de vista de la biología reproductiva, la principal causa del anestro observado en el posparto de las hembras mamíferas, no está relacionado a fallas en la actividad ovárica, una vez que la administración exógena de hormonas gonadotróficas.
(20) puede desencadenar el proceso ovulatorio, pero sí a una inhibición en la secreción pulsátil de la hormona liberadora de gonadotrofinas (GnRH) por parte del hipotálamo y consecuentemente de la hormona encargada del desarrollo y la maduración final del folículo preovulatório, esto es, de la hormona luteinizante (LH) por parte de la hipófisis anterior (Zalesky, 1990). La inhibición de la secreción de cantidades adecuadas de LH, en primer lugar, está dada por el efecto negativo que ejerce el estradiol de origen placentário sobre eje hipotalamo-hipófisis, sin embargo éste efecto cesa algunos días después del parto (Brooks, 1986), en segunda instancia, el estímulo generado por el efecto del amamantamiento de la(s) cría(s) sobre las hembras, induce la secreción de cantidades crecientes de opioides hipotalámicos, específicamente del péptido β-endorfina (Boland, 1990).. Los péptidos opioides acentúan el efecto negativo del estradiol sobre la secreción pulsátil de GnRH, y de esa manera, un estímulo continuo de amamantamiento impide la restitución de la actividad del eje hipotálamo-hipófisis, prolongando así el anestro en el periodo posparto (Acosta, 1983). En contraste, una reducción en la concentración. de. opioides. puede. ser. observada. si. el. estímulo. del. amamantamiento es retirado (Williams, 1994), y de esta manera la actividad cíclica se restituye. En ovinos, este efecto puede ser observado si se realiza un destete precoz de las crías o si el amamantamiento se restringe a 2 sesiones de 30 min.
(21) por día (Morales, 2004). También ha sido propuesta una interacción entre los efectos del amamantamiento y el desbalance energético que sufren las hembras en el posparto sobre la duración de la inactividad cíclica reproductiva. Los nutrientes consumidos en esta etapa, deben ser divididos para cumplir con las tareas de mantenimiento, producción de leche y desarrollo de las crías (Loudon, 1987). La insulina, la hormona de crecimiento y el factor de crecimiento similar a insulina (IGF-1) han sido señaladas como las sustancias responsables de priorizar la movilización de las reservas nutricionales, y todas ellas son sensibles de presentar variaciones de acuerdo al estado nutricional (Zaragoza & Angosto, 2006, p. 98), Conceptos alternos han surgido al respecto del efecto que presenta el amamantamiento sobre el desempeño reproductivo. Ha sido planteado que existe un componente de identificación materna y de selectividad hacia las propias crías, que regularía la respuesta de las hembras hacía el amamantamiento. En estas condiciones, es posible que solo las crías pertenecientes a determinada hembra, puedan inducir el efecto inhibitorio sobre la secreción pulsátil de GnRH (Williams, 1994)..
(22) 2.4 Metabolismo energético de ovejas en el periparto. El estado metabólico de un rumiante puede modificar la repartición de los nutrientes y su uso en diferentes vías catabólicas y anabólicas, a través de mecanismos como la homehorresis o la priorización de la distribución de la energía para satisfacer las demandas del organismo (Bauman, 1980). Un período de balance energético negativo alrededor del parto es considerado como la causa primaria del desarrollo de trastornos metabólicos y pérdidas productivas y reproductivas asociados a ellos (Robert J. Van Saun, 2000).. En rumiantes, ocurre una mínima absorción de glucosa originaria del intestino; aproximadamente 90% de la síntesis de glucosa ocurre en el hígado a través de la gluconeogénesis. Los principales precursores gluconeogénicos son el propionato y aminoácidos; otros precursores son el lactato absorbido en el rumen y, el glicerol liberado durante la lipólisis. La glucosa es el principal precursor de glicerol, la cual aumenta la disponibilidad de glicerol y favorece la lipogénesis. La estimulación de la movilización de triacilgliceroles en animales que presentan un balance energético negativo ocurre como efecto de compensación por la carencia de glicerol debido a las bajas concentraciones plasmáticas de glucosa (Boden, 1999)..
(23) El control del metabolismo de los lípidos durante el período periparto, es esencial para mantener la homeostasis corporal; fisiológicamente, los cambios hormonales, la liberación de catecolaminas y el aumento de la actividad del sistema nervioso simpático debido al estrés alrededor del parto, resulta en un incremento en la movilización de ácidos grasos no esterificados (NEFA) desde el tejido adiposo (Kaneko, 1989) (Herdt, 2000). La concentración plasmática de NEFA fue tres veces mayor en el primer mes posparto en ovejas alimentadas en sistema de confinamiento que en las ovejas en pastoreo, mientras que la concentración urea fue menor en las ovejas confinadas; los autores indican que esto se debió a la diferencia en el suministro alimenticio, que es inferior en animales confinados que a pastoreo al aire libre (Álvarez-Rodríguez, 2009).. Durante la lipólisis, los NEFA son transportados a otros tejidos unidos a la albúmina plasmática, mientras que el glicerol liberado se disuelve libremente en el plasma. En el hígado NEFA puede ser (Castellano Mendez & C, 2010) β-oxidado a acetil-CoA (Stellfug JN & JW, 2001), esterificado a triacilgliceroles los cuales son exportados como lipoproteínas de muy baja densidad (colesterol - VLDL) (Zurek, 1995), almacenado en el hígado en forme de triacilglicerol o destinado a la cetogénesis (SOUZA, 1999). Los principales productos de la cetogénesis son el acetoacetato, β-hidroxibutirato (βHB) y acetona formados a partir de la.
(24) condensación de dos moléculas de acetil-CoA (Ospina, 2010). En ovejas preñadas, un sistema de alimentación restringido llevó a una significante reducción en el peso vivo comparado a la alimentación control (media de 65 y 78 kg, respectivamente), pero se observó un cambio mínimo o ningún cambio en las concentraciones plasmáticas de glucosa, proteínas totales, triacilgliceridos, colesterol y lactato (Tanaka, 2008).. El tejido muscular también está involucrado en los mecanismos homeostáticos durante períodos críticos; los músculos utilizan lípidos y cuerpos cetónicos como fuente energética, además de la movilización de aminoácidos para soportar la gluconeogénesis hepática (Palmquist, 1994). Cuando las reservas lipídicas se consumen ocurre una sobre demanda de proteínas corporales para sostener la escasez de energía. El intenso uso de aminoácidos musculares genera un agotamiento. de. importantes. proteínas. estructurales. y. enzimáticas. en. consecuencia del déficit energético (Waltner, 1994).. La desnutrición en las ovejas también ha tenido consecuencias en los fetos, por lo que la glucemia y las concentraciones de lactato fueron más bajos que en los controles (no desnutridos) (Oliver et al., 2005). En ovejas gestantes, la alimentación restringida conllevó a una reducción del peso corporal en comparación con el control de la alimentación (peso vivo de 65 y 78 kg,.
(25) respectivamente), sin embargo, no presentó diferencias en las concentraciones plasmáticas de glucosa, proteínas totales, TG, colesterol y lactato o en la actividad de la lactato deshidrogenasa.. A su vez, Cuando se produce una balance energético negativo el cual suele presentarse en el período periparto, puede haber un exceso en la producción de cuerpos cetónicos (hipercetonemia) produciendo trastornos metabólicos como la cetosis. Se observó que la hipercetonemia disminuye la producción de glucosa hepática resultando en una reducción significativa de la glucemia de ovejas no preñadas y no lactantes, en el preparto y durante la lactancia (Schlumbohm, 2004).. 2.5 Condición corporal. La medición se realiza mediante una palpación a nivel lumbar, que permite estimar la cantidad de grasa bajo la piel, indicando los niveles de reservas energéticas que posee el animal. Está relacionado en forma positiva con el estado de engrasamiento (de la Isla Herrera, Aké López, Ayala Burgos, & González-Bulnes, 2010). La evaluación de la condición corporal (CC), permite realizar correcciones en el manejo y correcciones necesarias para incrementar la eficiencia productiva y reproductiva de los animales según su estado fisiológico la técnica se basa en el.
(26) principio, que el lomo es la última parte en que se acumula la grasa subcutánea y la primera en perderla. La cc en el ovino se evalúa palpando las apófisis espinosas y transversas de las vértebras lumbares con los dedos. La dureza de los huesos, el grosor de los músculos y el espesor de grasa deben evaluarse tocando el área del lomo, arriba y hacia atrás de la última costilla. (Mueller, 2015).. 2.6 Destete de corderos. Durante los primeros meses de vida los corderos se alimenta, fundamentalmente, de la leche de la madre, que aporta hasta un 75% de sus requerimientos energéticos, a medida que avanza la lactancia, disminuye el aporte energético de la leche producida por la hembra. Se debe tener en cuenta que las ovejas producen distintas cantidades de leche, circunstancia que depende, entre otros factores, de las exigencias del cordero. La producción láctea a partir de los 2 ó 3 meses posteriores al parto tiende a estabilizarse y luego declina; paralelamente el cordero inicia a consumir proporciones crecientes de pasto (Weary D. M., 2008). Respuesta de estrés al destete artificial la remoción abrupta y completa de los corderos antes de la edad del destete natural es una situación estresante para la oveja y para el cordero, provocada por diferentes estresores: . Emocional, generado por la ruptura del vínculo madre-cría..
(27) . Cambios en el ambiente físico y social, ya que al menos uno de ambos es alojado en un nuevo ambiente, muchas veces con otros animales con los que no tenía contacto previo.. . Cese de la succión, lo que implica el cese de los cambios neurofisiológicos inducidos por la misma.. . Cambio nutricional, generado por la pérdida de la leche y su sustitución completa por otros alimentos.. La respuesta del animal es consecuencia de todos estos estresores en conjunto, pero la importancia relativa de cada uno ha sido escasamente estudiada. Luego de. la. separación, la. oveja. y el cordero. aumentan. la frecuencia. de. comportamientos relacionados a la búsqueda del otro. Se observa un aumento de la frecuencia de vocalizaciones (Orgeur P. B., 1999) y del tiempo que pasan parados y caminando, además de que frecuentemente costean (cuando los animales se mueven repetidas veces a 1 a 2 m de manera paralela al alambrado modificando la dirección de ida y vuelta en forma continua), comportamiento que en los corderos se asocia exclusivamente a la pérdida de la madre (Damián, 2013)..
(28) El aumento en la cantidad de vocalizaciones y el despliegue del comportamiento costear son los cambios de comportamiento más indicativos de la respuesta de estrés al destete. Generalmente estos cambios se producen rápidamente, alcanzan su máxima frecuencia en pocas horas, y se manifiestan durante 3 a 4 días. El rápido cese de estos comportamientos podría estar relacionado a su falta de efectividad, ya que generan un importante costo sin lograr respuestas positivas (Weary D. M., 1995).. Además, al aumentar los comportamientos relacionados con la búsqueda de la cría o de la madre, las ovejas y corderos reducen los comportamientos de alimentación y descanso: los animales pasan menos tiempo echados, pastando y rumiando luego del destete (Damián, 2013). En corderos recién destetados alimentados en base a pasturas o con sustitutos lácteos, se ha observado que algunos corderos redirigen el comportamiento de succión intentando mamar de otros corderos (Pérez-León, 2006). La pérdida de la madre como consecuencia del destete es un factor central en la respuesta de estrés de los corderos.. El destete también provoca cambios fisiológicos agudos indicadores de estrés en las ovejas y corderos, como un aumento rápido en la concentración sérica de cortisol, (Pérez-León, 2006) que es uno de los principales indicadores endocrinos de la respuesta de estrés en ovinos. (Möstl, 2002) El aumento de la secreción de.
(29) cortisol está asociado con la modulación de la respuesta inmune del animal (Griffin, 1989), ya que en los rumiantes se ha reportado que el destete artificial afecta negativamente la función inmune (Ungerfeld, 2011). La cantidad de anticuerpos sintetizados luego de un desafío antigénico es menor en los corderos durante el período post-destete que en otros momentos (Napolitano, 1995). Luego del destete en las ovejas se observa un aumento de los leucocitos (Orgeur P. M., 1998), y una disminución de la concentración de proteínas totales y globulinas (Freitas-de-Melo, 2013).. Dado que el destete es un manejo estresante y que afecta el sistema inmune, puede aumentar la susceptibilidad del animal a enfermedades (Watson, 1991).. El destete también afecta negativamente el crecimiento de los corderos, el que puede ser más marcado en los corderos destetados antes de completar el desarrollo del rumen si no son criados artificialmente (Caneque, 2001). Este retraso en el crecimiento también podría ser explicado como consecuencia del peor estatus inmunológico y el aumento de la susceptibilidad a parasitosis gastrointestinales, además de los cambios de comportamiento indicadores de estrés (Damián, 2013) (Orgeur P. B., 1999). En síntesis, los cambios de comportamiento y las respuestas fisiológicas tienen consecuencias negativas tanto.
(30) a nivel productivo como en el bienestar de los animales, por lo que es importante desarrollar alternativas para disminuirlas..
(31) 3. METODOLOGÍA. El presente estudio se llevó acabo en el Centro Académico Agroindustrial El Ciruelo de la Universidad Cooperativa de Colombia, el cual se encuentra ubicado en la vereda Bejaranas bajo, en el municipio de San Gil Departamento de Santander; con una altura de 1114 m.s.n.m., una temperatura media de 24º C, una máxima de 32º C y una mínima de 16º C, con una extensión de 19.5 hectáreas. El tipo de investigación realizada fue de tipo experimental, donde se utilizaron 23 hembras de raza criolla entre 2 a 5 partos, las cuales fueron sincronizadas después de una previa selección con respecto a condiciones sanitarias y en una buena CC, se realizó adecuación de dos potreros para la implementación del amamantamiento restringido. En estas adecuaciones fueron introducidas las ovejas de los grupos experimentales, conformado por ocho ovejas cada uno con sus respectivas crías y un macho reproductor en cada grupo y un grupo control con 7 ovejas y un macho reproductor.. Los ovinos fueron manejados en un sistema de pastoreo rotacional en praderas permanentes de Cynodon plectostachium y Brachiaria sp con libre acceso a agua; además, se les suministró 30 g/animal/día de suplemento mineral ganasal® al 8%, los corderos fueron suplementados con Pithecellobium dulce..
(32) 3.1 Grupos experimentales. Las 23 hembras seleccionadas fueron asignadas de forma aleatoria, a uno de los siguientes grupos, según el sistema de manejo del amamantamiento restringido hasta el momento del destete (90 días). 3.1.1 Control (GC): Amamantamiento sin restricción (ad libitum); las hembras permanecieron junto a los corderos 24h/d, desde el día del nacimiento, hasta el destete. Este grupo consta de 7 ovejas y 11 corderos. 3.1.2 Grupo 1 (G1): Amamantamiento restringido a partir del tercer día de nacimiento de la cría. Este grupo constó de 8 ovejas y 10 corderos. Las ovejas ingresan al corral con las crías 30 minutos en horas de la mañana y 30 minutos en horas de la tarde para permitir la lactancia de las crías. 3.1.3 Grupo 2 (G2): Amamantamiento restringido, a partir de los 15 días de nacimiento de la cría. Este grupo consta de 8 ovejas y 10 crías. Las ovejas ingresan al corral con las crías 30 minutos en horas de la mañana y 30 minutos en horas de la tarde para permitir la lactancia de las crías..
(33) 3.2 Evaluación de la condición corporal y pesaje de las hembras. Fue evaluada la condición corporal en los mismos días experimentales de la obtención de las muestras de sangre, en una escala de 1,0 a 5,0 con intervalos de 0,25 puntos, donde 1,0 representa un animal muy delgado y 5,0 un animal obeso.. 3.3 Evaluación del peso al destete de los corderos.. Los corderos serán pesados al nacer y al destete en la semana 12. El pesaje se realizará con una báscula calibrada previamente a cada pesaje.. 3.4 Evaluación del balance energético negativo.. Se obtuvo, de cada hembra, una muestra de sangre mediante la venopunción de la yugular utilizando tubos vacutainer® sin anticoagulante y otro con heparina. Las muestras para el análisis de metabolitos indicadores de balance energético se obtuvo en las horas de la mañana, una vez por semana desde la 1ª semana posparto hasta la 5ª semana posparto, y quincenalmente a partir de la 6ª hasta las 12ª semana de lactancia, cuando se lleva a cabo el destete. Se determinarán las concentraciones de beta hidroxibutirato (βHB), glucosa, colesterol total, urea, proteína total y albúmina mediante kits enzimáticos Randox®. Las muestras serán.
(34) analizadas en espectrofotómetro semiautomático (Metrolab 1600 plus ®, Tucamán, Argentina). En el laboratorio, se centrifugaron las muestras a 3.000 rpm durante 20 minutos para la obtención de suero y plasma, los cuales serán congelados a -20 ºC hasta su posterior análisis. En total se obtuvieron 18 muestras por cada hembra las cuales son 9 muestras de suero sanguíneo y 9 muestras con concentraciones plasmáticas para un total de 414 muestras en total con todas las hembras de la investigación.. 3.5 Evaluación del intervalo parto-primer celo. La detección del primer celo posparto se evaluó mediante observación directa, para lo cual fueron utilizados tres machos enteros con chaleco marcador, los cuales permanecieron con las hembras todo el día y fueron remplazados cada quince días durante todo el periodo de muestreo para evitar el agotamiento. Para la detectar la presencia de los celos fueron las hembras fueron observadas dos veces/día (mañana y tarde). Las ovejas marcadas fueron consideradas hembras que entraron en el periodo de celo. Con el objetivo de determinar si el celo detectado en las ovejas fue seguido de ovulación, serán determinadas las concentraciones de progesterona en el suero sanguíneo. Para este propósito se colectaron muestras de sangre a través de la venopunción de la vena yugular cada 3 días desde el día del celo hasta el día 15 posterior a este (total de 6 muestras/hembras). Una vez en el laboratorio, las muestras de sangre fueron.
(35) centrifugadas a 3.000 rpm durante 20 minutos para la obtención del suero sanguíneo, que fue envasado en crioviales de 1,5 ml e inmediatamente después las muestras de suero se congelaron a -20 ºC hasta su posterior análisis. La concentración de progesterona en las muestras será determinada a través de la técnica de radioinmunoanálisis. Posteriormente se realizó el diagnostico de gestación (45 días post servicio), mediante ultrasonografía transrectal. La presencia de contenido uterino correspondiente a una vesícula gestacional, un feto y las membranas placentarias serán tomadas como indicadores de preñez..
(36) 4. RESULTADOS Y DISCUSIÓN. 4.1 Evaluación de la condición corporal y el pesaje de los animales.. El peso vivo presenta la ventaja de ser fácil de medir, pero al existir grandes variaciones en el tamaño de los animales existentes en un rebaño, no refleja de manera adecuada de la masa corporal de un animal, se pueden ver afectados los datos obtenidos por el peso del contenido digestivo y en el caso de las ovejas gestantes por el peso de los fetos y de los líquidos fetales. La condición corporal resulta ser un método de fácil aplicación que no necesita de ningún equipo para su determinación y además no se encuentra afectado por los estados fisiológicos del animal, la condición corporal lo único que se requiere un grado de entrenamiento.. 4.1.1 Condición Corporal. La condición corporal representa un importante indicador de las reservas corporales de energía disponibles en las ovejas, y tiene importantes efectos sobre la función reproductiva. En el transcurso de las 12 semanas se evaluó la CC semanalmente de los grupos experimentales GC, G1, G2, en donde. registraron al momento del parto, se. encontraban en una condición corporal de 3,40 ± 0,1 puntos, donde en los primeros 15 a 20 días post parto es normal que pierdan de 0.5 puntos debido a los.
(37) cambios metabólicos y movilización de energía para suplir el desgaste que conlleva la producción de leche lo cual con buena nutrición y suplementación se recuperara a los 40 días post parto (Figura 1).. Figura 1. Evaluación de la condición corporal de ovejas de pelo en el transcurso de las 12 semanas.. Promedio de CC 3,50. CC. 3,25. 3,00 G1 G2. 2,75. Gc 2,50. Semana Promedio de Condición Corporal de las hembras evaluadas en las 12 semanas donde ± ANOVA (P<0.05). G1 = grupo 1 amamantamiento restringido 3 días post parto (n=8) G2 = grupo 1 amamantamiento restringido 15 días post parto (n=8) GC = grupo control amamantamiento sin restricción(n=7).. El estudio demostró que en las primeras dos semanas se presenta una pérdida considerable de peso, en la cual es notorio el G1 presentando la pérdida más significativa de los 3 grupos durante el periodo del peripato presentando una perdida promedio de 0.25 puntos comparada con G2 y GC que solo perdieron en.
(38) promedio 0.5 punto, los resultados obtenidos por GC fueron los más favorables. En el transcurso de las doce semanas se muestra una estabilidad de la perdida de condición corporal; ya en la semana 5 se manifiesta una estabilidad hasta la semana 9 cuando los grupos experimentales G1 y G2 empiezan a recuperar la condición corporal hasta la semana 11 en la cual estabilizan nuevamente. El grupo Gc después de que se estabilizo en la quinta semana hasta la semana 12 no evidencio una ganancia para la recuperación de la CC, debido a la constante secreción de leche por parte de los corderos. Una baja de la condición corporal durante el posparto, resulta en una disminución en la fertilidad. Se ha demostrado que en las primeras semanas después del parto, el gasto o pérdida de energía para producción de leche, mantenimiento y funciones corporales. Para llenar este déficit, la oveja se ve obligada a tomar reservas corporales. Las ovejas con el mayor balance energético negativo a menudo experimentan un gran número de días abiertos. Folículos preantrales son reclutados durante periodos de severas deficiencias de energía y producen óvulos de mala calidad, así como, una disfunción del cuerpo lúteo. Así es que en la primera ovulación mientras no se restaure el balance energético serán liberados óvulos de mala calidad o degenerados y además la formación de un cuerpo lúteo con pocos niveles de progesterona. La actividad folicular ovárica aumenta conforme el animal entra en un balance energético positivo. Las hormonas que parecen ser influenciadas por un status energético positivo son: LH, FSH, Insulina, BST, etc. (Bazer, 1989).
(39) 4.1.2 Evaluación de peso.. Una forma práctica de evaluar la cantidad de energía almacenada en forma de grasa y músculo por la hembra en un momento dado que se debe realizar en horas de la mañana o con un previo ayuno para evitar obtener datos erróneos. La medición del nivel de reservas por CC resulta más útil que el registro del peso corporal, no sólo por cuestiones prácticas sino porque la información que brinda puede prescindir del biotipo, estado reproductivo y llenado del tracto digestivo de los animales. (Jorge Hugo Veneciano, 2014). La comparación. de los grupos experimentales en referencia al pesaje. (kilogramos) de las hembras evaluadas durante la investigación, se observó que los grupos Gc, G1, G2, iniciaron con un promedio de peso de 36 ±1 kilogramo reflejando que los tres grupos se comportan de una manera muy similar hasta la novena semana donde el grupo Gc obtiene una mayor ganancia de peso. Al peso que registraron al momento del parto y en el transcurso de las 12 semanas, en general se presentó una variación de 2 kg (Figura2)..
(40) Figura 2. Evaluación de peso (kilogramos) de los tres grupos experimentales en ovejas de pelo en un periodo de 12 semanas.. Promedio de peso 39. Kilogramos. 37 35 33 31. G1. 29. G2. 27. Gc. 25. Semanas Promedio de peso hembras evaluadas en las 12 semanas donde ± ANOVA (P<0.05). G1 = grupo 1 amamantamiento restringido 3 días post parto (n=8) G2 = grupo 1 amamantamiento restringido 15 días post parto (n=8) GC = grupo control amamantamiento sin restricción (n=7).. Los tres grupos experimentales se comportaron muy paralelamente en el transcurso de las 12 semanas, exceptuando el grupo Gc el cual tuvo una pérdida de peso hasta la quinta semana debido al periodo del periparto, a partir de esta semana las ovejas encontraron el balance energético estabilizando su peso hasta la semana del destete de los corderos, las crías disminuyeron los periodos de lactancia al presentar el cambio de alimentación de la leche por forraje verde. Los grupos G1 y G2 tuvieron pérdidas de pesos similares y un comportamiento parecido en el transcurso de las 12 semanas, debido a la interrupción del pastoreo normal dos veces al día por 30 minutos durante los periodos de lactancia lo cual alteró el proceso fisiológico normal de rumia generando periodos de ayuno más.
(41) prolongados y periodos de stress por el manejo que fueron sometidos las ovejas, no se presentaron perdidas de pesos significativamente en ninguno de los tres grupos.. 4.2 Peso promedio de las crías.. El destete es el proceso de separar las crías de la madre en un momento determinado, terminando de esta manera con el suministro de la leche materna esta práctica es de manejo rutinario en la mayoría de producciones; El peso al destete es un registro de gran importancia determinando la ganancia de kilos que obtuvo la cría desde el momento de nacer hasta que va hacer separado de la madre para pasar a una nueva etapa en la producción donde evalúa la habilidad materna, calidad del alimento y conversión nutricional. (Chieza, 2010) Los corderos de estudio fueron suplementados con una mezcla de harinas diseñada para pequeños rumiantes que contiene 13.78% de proteína, 5.17% de grasa, 7.50% de fibra, 10.74 de cenizas y 9.29% humedad, se les suministro 30g/animal/día de sal mineralizada al 8%, Pithecellobium dulce (gallinero) dos raciones al día, con libre acceso de agua.. Los grupos experimentales evaluados evidenciaron la ganancia de peso (kilogramos) de los corderos que se registraron al final del estudio. Realizando una.
(42) comparación de los tres grupos experimentales de corderos donde se puede evaluar el peso al momento del destete,. en donde el grupo GC registro un. promedio de peso de 13,1 kg. El grupo G2 con un promedio de ganancia de peso de 9,88 kg y el grupo G1 con un promedio peso de 7,2 kg. (Figura 3). Figura 3. Evaluacion del peso (kilogramos) al destete de la crias de ovejas de pelo en el periodo de 12 semanas.. Peso promedio de crias 14. 13,1. 12 9,88. Kilogramos. 10 8. 7,2 G1. 6. G2 Gc. 4 2 0 G1. G2. Gc. Grupos experimentales Peso promedio de las crías donde ± ANOVA (P<0.05) G1 = grupo 1 amamantamiento restringido 3 días post parto (n=8) G2 = grupo 1 amamantamiento restringido 15 días post parto (n=8) GC = grupo control amamantamiento sin restricción (n=7).. En el estudio se reflejó que los corderos del grupo Gc registraron las mayores ganancias de peso con 13,1 kg en promedio, el amamantamiento continúo y la habilidad que adquieren al estar con las hembras lactantes desarrollan un mayor aprendizaje para desarrollarse más fácilmente en el rebaño. En relación el grupo.
(43) G1 que promedio un peso de 7,2 kg donde registro el peso más bajo de los tres grupos estudiados evidenciando una diferencia entre los dos grupos de aproximadamente 6 kg. El amantamiento restringido que se utilizó en este grupo presentó cambio brusco en la dieta de los corderos los cuales fueron sometidos a un desarrollo prematuro del rumen y a una adaptación metabólica, modificando la digestión de un mono-gástrico del lactante a la fermentación y digestión de un rumiante adulto. Se debe tener en cuenta que pasa de una dieta líquida a una sólida incentivada por el hombre con el tipo de ración que se le suministra (Chieza, 2010).. Como en estudios similares los grupos control siempre mostraron un mejor comportamiento a diferencia de los grupos restringe el amantamiento (Gladis Morales-Terán1, 2004, p. 168).. 4.3 Evaluación de intervalo parto – primer celo.. De los múltiples factores condicionantes en los sistemas de producción ovina, la reproducción ocupa un lugar fundamental, ya que la rentabilidad económica de las explotaciones está condicionada en gran medida por el rendimiento reproductivo de las hembras adultas. Rendimiento reproductivo que se mide en número de corderos producidos por oveja a lo largo de su vida y que viene determinado por.
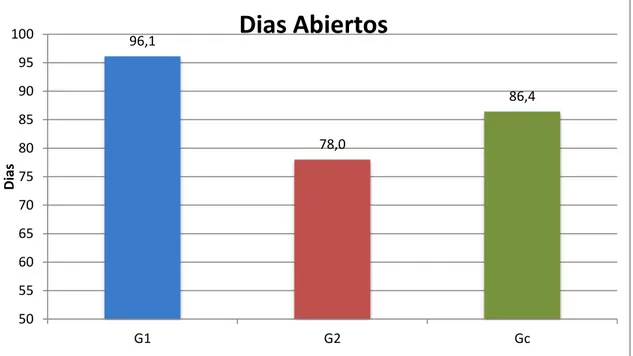
(44) parámetros como la edad al primer parto, el intervalo entre partos, la prolicifidad y la duración de la vida útil (Mueller, 2015). Realizando una comparación de los 3 grupos experimentales se evaluó el periodo de días abiertos. El grupo Gc promedio 86,4 días, el grupo G1 promedio 96,1 días y el grupo G2 promedio 78 días abiertos. (Figura 4) Figura 4. Periodo de días abierto de las ovejas de pelo de los tres grupos experimentales. 100. 96,1. Dias Abiertos. 95 90. 86,4. 85 78,0. Dias. 80 75 70 65 60 55 50 G1. G2. Gc. Días abiertos de las ovejas, donde ± ANOVA (P<0.05) G1 = grupo 1 amamantamiento restringido 3 días post parto(n=8) G2 = grupo 1 amamantamiento restringido 15 días post parto (n=8) GC = grupo control amamantamiento sin restricción (n=7).. El grupo G2 registro el menor tiempo en promedio con 78 días para la reactivación ovárica post-parto por la restricción del amantamiento a partir de los 15 días postparto disminuyendo la secreción de la leche, reduciendo el perdido del periparto, iniciando su reactivación hormonal más rápido que los otros dos grupos experimentales. El grupo G1 en relación al grupo G2 fue el que obtuvo un.
(45) promedio mayor con 96,1 días periodo para iniciar primera ovulación post-parto evidenciando una diferencia de 18 días entre los dos grupos estudiados debido a una caída en la condición corporal y la pérdida de peso (Fig. 1 y 2), del grupo G1 se vio afectado su primera ovulación post-parto. En estudios similares el amamantamiento ejerció un efecto inhibitorio en el restablecimiento de la actividad ovárica postparto en ovejas Pelibuey (Gladis Morales-Terán1, 2004). En hembras con nutrición balanceada se ha comprobado, la formación de ondas foliculares con presencia de folículos dominantes de diámetro semejante al de un folículo ovulatorio, a partir de la segunda o tercera semana posparto (Jolly, 1994), pero en las animales con nutrición deficiente, aunque también se desarrollan ondas foliculares durante el posparto temprano, se ha observado una disminución del diámetro y de la vida media del folículo dominante y un menor número de folículos. Al presentarse una disminución de peso y de cc, el anestro posparto es de mayor duración que en las que no los disminuyen (Angel, 1996). Aumentando el número de celos silenciosos y de celos con tiempos más cortos..
(46) 5. CONCLUSION. La implementación de un sistema de amamantamiento restringido posiblemente podría disminuir los días abiertos con relación a la primera ovulación post parto, sin alterar significativamente el peso de las hembras y de sus crías, donde posiblemente sería un sistema apto para la implementación del manejo post parto en las granjas ovinas..
(47) 6. RECOMENDACIÓN. Dar un alimento balanceado de mejor calidad con el cual se van a suplementar los corderos para no disminuir el rendimiento de los pesos al momento del destete, administrando también en épocas de verano a todos los animales.. Mejorar las instalaciones de corrales para el manejo de la producción ovina y el manejo que se tiene en los potreros.. Implementar un plan sanitario para el hato ovino disminuyendo la carga parasitaria en los corderos.. Tratar el agua que se suministra en la producción..
(48) 7. BIBLIOGRAFÍA. Acosta, B. T. (1983). Nursing enhances the negative effect of estrogen on LH release in the cow. Journal of animal science, 1530-1536. Álvarez-Rodríguez, J. S. (2009). The effect of the spring management system on blood metabolites and luteal function of ewes on Mediterranean mountain areas. Small Ruminant Research,, 18-26. Arroyo, J., Magaña-Sevilla, H., & Camacho-Escobar, M. (2009). REGULACIÓN NEUROENDOCRINA DEL ANESTRO POSPARTO EN LA OVEJA. Tropical and Subtropical Agroecosystems. Bartlewski, P. M. (2000). THE RELATIONSHIPS BETWEN OVAïUAN ANTRAL FOLLICLE DYNGMICS, LUTEGL, FUNCTION AND ENDOCRINE VARIABLES IN EWES. Doctoral dissertation, College of Veterinary Medicine, University of Saskatchewan, Saskatoon. Bauman, D. E. (1980). Partitioning of nutrients during pregnancy and lactation: a review of mechanisms involving homeostasis and homeorhesis. Journal of dairy science, 1514-1529. Bazer, F. W. (1989). Establishment of pregnancy in sheep and pigs. Reproduction, Fertility and Development, 237-242. Boden, G. &. (1999). Effects of fatty acids and ketone bodies on basal insulin secretion in type 2 diabetes. Diabetes, 577-583. Boland, M. P. (1990). The use of ultrasound to monitor ovarian function in farm animals. Agbiotech News and Information, 841-844. Brooks, A. N. (1986). Opioid modulation of LH secretion in the ewe. Journal of reproduction and fertility, 693-708. Brunet, A. G. (1991). Effect of season on plasma concentrations of prolactin and cortisol in pregnant, non-pregnant and lactating ewes. Animal Reproduction Science. Cahill, L. P. (1980). Influences of season, cycle and breed on follicular growth rates in sheep. Journal of reproduction and fertility, 321-328..
(49) Caneque, V. V. (2001). Effect of weaning age and slaughter weight on carcass and meat quality of Talaverana breed lambs raised at pasture. Animal Science, 85-95. Chieza, D. (2010). El destete como herramienta para un mejor negocio ganadero. instituto de promocion de la carne vacuna argentina. Damián, J. P. (2013). Behavioural response of grazing lambs to changes associated with feeding and separation from their mothers at weaning. Research in veterinary science, 913-918. de la Isla Herrera, G., Aké López, J. R., Ayala Burgos, A., & González-Bulnes. (2010). Effect of body condition and season of the year on estrous cycle, estrous, follicular development and ovulation rate in Pelibuey ewes under tropical conditions. Veterinaria México, 167-175. Departamento de Producción Animal Estación Experimental Bariloche Instituto Nacional de Tecnología Agropecuaria. (2015). Actualización en Producción Ovina 2015. IX Curso de Actualización en Producción Ovina (pág. 12). San Carlos de Bariloche: ISBN 978-987-521-651-8. Dufour, J. C. (1979). Short-and long-term effects of hypophysectomy and unilateral ovariectomy on ovarian follicular populations in sheep. Journal of reproduction and fertility, 301-309. Flint, A. P. (1983). Evidence for a systemic role for ovarian oxytocin in luteal regression in sheep. Journal of reproduction and fertility, 215-231. Freitas-de-Melo, A. B. (2013). Progesterone administration reduces the behavioural and physiological responses of ewes to abrupt weaning of lambs. animal, 1367-1373. Gladis Morales-Terán1, A. P.-M.-S.-d.-R.-S. (2004). CONTINOUS OR RESTRICTED SUCKLING AND ITS RELATIONSHIP TO THE LENGHT OF POSTPARTUM ANESTROUS IN PELIBUEY EWES. Agrociencia 38: , 165171. Griffin, J. F. (1989). Stress and immunity: a unifying concept. Veterinary immunology and immunopathology, 263-312. Hay, M. F. (1977). Changes in the Graafian follicle population during the follicular phase of the oestrous cycle. In Proceedings..
(50) Herdt, T. H. (2000). Ruminant adaptation to negative energy balance: Influences on the etiology of ketosis and fatty liver. Veterinary Clinics of North America: Food Animal Practice, 215-230. Jolly, P. D. (1994). Physiological effects of undernutrition on postpartum anoestrus in cows. Journal of reproduction and fertility, 477-492. Jorge Hugo Veneciano, C. A. (2014). CRÍA Y RECRÍA DE BOVINOS. Sitio Argentino de Producción Animal. Juan Gonzalo Castellano Mendez, J. C., & C, C. L. (2010). Prospectiva de investigación y desarrollo tecnologico para la cadena productiva carnica ovina-caprina en colombia . ISBN. 978-958-8536-21-7. Kaneko, J. J. (1989). Carbohydrate metabolism and its diseases. Clinical biochemistry of domestic animals, 44-80. Karsch, F. J. (1980). Feedback basis of seasonal breeding: test of an hypothesis. Journal of reproduction and fertility, 521-535. López Sebastián, A. S. (1993). Aspectos característicos de la fisiología de la oveja. Revista Científica. López Sebastián, A. S. (1993). Aspectos característicos de la fisiología de la oveja. Revista Científica. Loudon, A. (1987). Nutritional effects on puberty and lactational infertility in mammals. Proceedings of the Nutrition Society, 203-216. Manazza, J. ( 2006). MANEJO DE CARNEROS Y OVEJAS EN SERVICIO A CAMPO. EEA INTA Balcarce *Grupo Sanidad Animal, 1. McNeilly, A. S. (1984). . Changes in FSH and the pulsatile secretion of LH during the delay in oestrus induced by treatment of ewes with bovine follicular fluid. Journal of reproduction and fertility, 165-172. Morales, G., Pro, A., Figueroa, B., Sánches, C., & Gallego, J. (marzo-abril, 2004). amamantamiento continuo o restringido y su relacíon con la duracion del anestro postparto en ovejas pelibuey. agrociencia, vol. 38 num. 2, 167-168. Möstl, E. &. (2002). Hormones as indicators of stress. Domestic animal endocrinology, 67-74..
(51) Napolitano, F. M. (1995). Influence of artificial rearing on behavioral and immune response of lambs. Applied Animal Behaviour Science. Niswender, G. D. (1985). Regulation of luteal function in domestic ruminants: new concepts. Recent Prog Horm, 51-101. Olivera Angel, M., Castillo, J., Ruiz, T. E., & Jiménez, C. (1996). Días abiertos en vacas brahman y su relación con peso y condición corporal. Revista Colombiana de Ciencias Pecuarias, 66-67. Orgeur, P. B. (1999). Psychobiological consequences of two different weaning methods in sheep. Reproduction Nutrition Development, 231-244. Orgeur, P. M. (1998). Artificial weaning in sheep: consequences on behavioural, hormonal and immuno-pathological indicators of welfare. Applied Animal Behaviour Science, 87-103. O'Shea, J. D. (1979). The small luteal cell of the sheep. Journal of anatom. Ospina, P. A. (2010). Evaluation of nonesterified fatty acids and β-hydroxybutyrate in transition dairy cattle in the northeastern United States: Critical thresholds for prediction of clinical diseases. Journal of Dairy Science, 546-554. Palmquist, D. L. (1994). The role of dietary fats in efficiency of ruminants. The Journal of nutrition, 1377-1382. Pérez-León, I. O. (2006). Reducing mother-young separation distress by inducing ewes into oestrus at the day of weaning. Animal Welfare, 383-389. RD, R. (1990). Nutrition and postpartum rebreeding in cattle. Journal of Animal Science, 853-862. Restall, B. J. (1977). The influence of season of lambing and lactation on reproductive activity and plasma LH concentrations in Merino ewes. Journal of reproduction and fertility. Robert J. Van Saun, D. P. (2000). Pregnancy toxemia in a flock of sheep. Journal of the American Veterinary Medical Association, 1536-1539. Roblero, M. &. (2014). Somatotropina bovina recombinante (rbST) y su efecto en la inducción de la ovulación en ovejas pelibuey amamantando. Schlumbohm, C. &. (2004). Hyperketonemia impairs glucose metabolism in pregnant and nonpregnant ewes. . Journal of dairy science, 350-358..
(52) Sebastian, A. L. (1988). . Effects of lactation status, progestogen and ram exposure on response to cloprostenol in ewes during the anestrous season. Theriogenology, 279-289. SOUZA, P. D. (1999). Efeito da amamentacao sobre o desempenho reprodutivo pos-parto em ovelhas da raca Santa Ines. Ciencia veterinaria tropical, 115. Stellfug JN, H. P.-R., & JW, W. (2001). Reproductive performance of ewe lambs from ewes from different selection practices with or without induced estrus. Animal Reproduction Science, 185-193. Tanaka, Y. M. (2008). Plasma metabolite concentrations and hepatic enzyme activities in pregnant Romney ewes with restricted feeding. Research in veterinary science, 17-21. Ungerfeld, R. H. (2011). Behavioral and physiological changes in early-weaned multiparous and primiparous beef cows. animal,, 1270-1275. Waltner, S. S. (1994). Relationships of body condition score to production variables in high producing Holstein dairy cattle. Journal of Dairy Science, 3410-3419. Watson, D. L. (1991). Effect of weaning on antibody responses and nematode parasitism in Merino lambs. Research in veterinary science, 128-132. Weary, D. M. (1995). Signalling need: costly signals and animal welfare assessment. Applied Animal Behaviour Science, 159-169. Weary, D. M. (2008). Understanding weaning distress. Applied Animal Behaviour Science, 22-41. Williams, G. L. (1994). Sensory and behavioural control of gonadotrophin secretion during suckling-mediated anovulation in cows. Journal of reproduction and fertility, 463-475. Zalesky, D. D. (1990). Suckling inhibits release of luteinizing hormone-releasing hormone from the bovine median eminence following ovariectomy. Journal of animal science, 444-448. Zaragoza, F. M., & Angosto, M. C. (2006). ENFERMEDADES METABÓLICAS. Madrid: INSTITUTO DE ESPAÑA; REAL ACADEMIA NACIONAL DE FARMACIA..
(53) Zurek, E. F. (1995). Metabolic status and interval to first ovulation in postpartum dairy cows. Journal of Dairy Science, 1909-1920..
(54)
Figure



Outline
Documento similar
Entre nosotros anda un escritor de cosas de filología, paisano de Costa, que no deja de tener ingenio y garbo; pero cuyas obras tienen de todo menos de ciencia, y aun
o Si dispone en su establecimiento de alguna silla de ruedas Jazz S50 o 708D cuyo nº de serie figura en el anexo 1 de esta nota informativa, consulte la nota de aviso de la
The 'On-boarding of users to Substance, Product, Organisation and Referentials (SPOR) data services' document must be considered the reference guidance, as this document includes the
In medicinal products containing more than one manufactured item (e.g., contraceptive having different strengths and fixed dose combination as part of the same medicinal
Products Management Services (PMS) - Implementation of International Organization for Standardization (ISO) standards for the identification of medicinal products (IDMP) in
Products Management Services (PMS) - Implementation of International Organization for Standardization (ISO) standards for the identification of medicinal products (IDMP) in
This section provides guidance with examples on encoding medicinal product packaging information, together with the relationship between Pack Size, Package Item (container)
Package Item (Container) Type : Vial (100000073563) Quantity Operator: equal to (100000000049) Package Item (Container) Quantity : 1 Material : Glass type I (200000003204)
