Distribution of attacked plants along trails in leaf cutting ants (Hymenoptera: Formicidae): consequences in territorial strategies
6
0
0
Texto completo
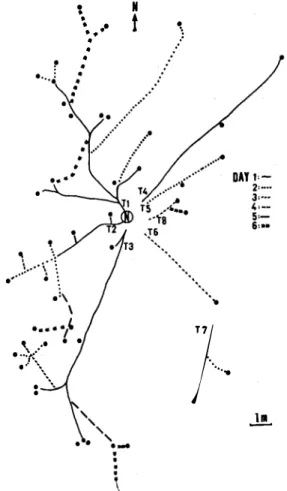
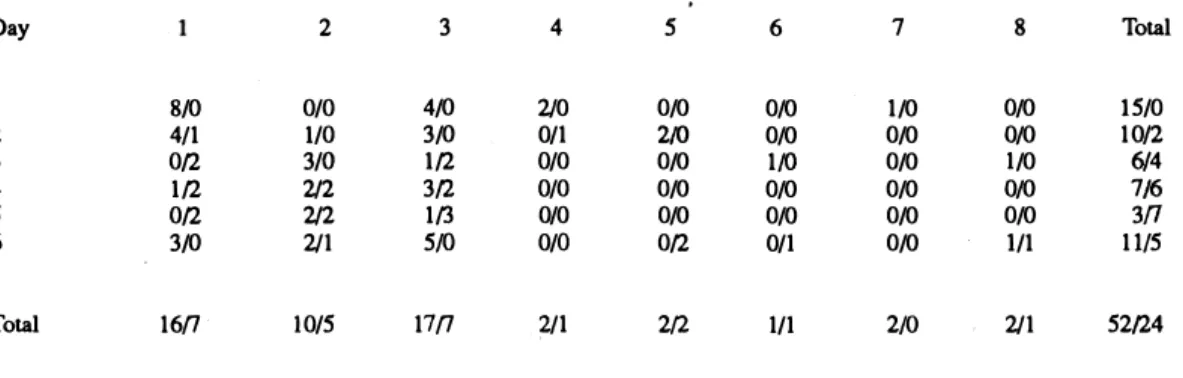
(2) 892. REVISTA DE BIOLOGIA lROPICAL. cations in the geometry of foraging territories (powler & Stiles 1980). Much has been written about why trail systems exist in leaf-cutting ants, but almost nothing about their consequences in relation to territorial defense. Even though trails ate constructed to harvest palatable patches, it seems probable that they also have an "inertial" defensive function in protecting t he potential and res tricted vegetation patches from possible competitors. It is known that leaf-cutting ants do frequently defend their territory, especially against other Atta species (Vilela & Howse 1986). Spatial distribution between colonies is an effective measure to reduce the aggression between sorne ant species ( HOlldobler & Lumsden 1980). Intraspecific attacks in Atta include attacks on founding queens from other nests, on smaller colonies, and battles between columns of workers from different nests (powler 1977, Jutsum 1979, and personal field observation). This study presents preliminary evidence that supports the hypothesis that rotation in harvest activity and trail results in a general and unpredictable covering of the area around the nest. This characteristic could be selectively advantag e o u s to leaf-cutting ants as an "inertial" defense strategy. T h e resear c h was conducted on Barro Colorado Island (BCI), Panamá, during the late rainy season, October 1989. BCI, in Gatun Lake, is covered with tropical deciduous and semi-evergreen seasonal forest, estimated to be from 75 to 200 years old, and Atta spp. are common. We selected one adult Atta cephalotes (L.) nest (5 m aprox. o f ratio) near the Shannon-Snyder Molino trail, and for seven consecutive days (except for one rainy day without ant activity) revisited it each moming. Each day we mapped: (a) number, distance and orientation (angle respect to the north) of the attacke d/abandoned trees, and ( b) number, length, and orientation of new/abandoned trails. Ang ular data were analyzed w i t h angular statistics (Batschelet 1981). Attacked plants per trail show a clustering in their angular disposition with regard to the main trail orientations (pig. 1 and 2). The distribution of trail orientations can not be distinguished from random (Rayleigh test, P> .55, Bastchelet 1981, Fig. 2). A random distribution of the location of the abandoned trees (with 1, 2, 3 and 4 consecutive days of attack), also cannot be. rejected (Rayleigh test, P= .25, Bastchelet 1981) (pig. 3). Data of attacked and abandoned trees is presented in Appendh: 1, and its detall per day and trail in Table 1. 11. t. \ .• • •. •. t. .)'. •. \. ..... ....i". ,}7.. .. ... .•. •. ". . ...... ,......... .. . ... • •. •. ..!!.. Fig. 1. Map of one aduh Atta cephaJotes nest (N) with the physica1 trails (T) (Jines) and attacked (circ1es) during seven sampling days. The different lines utilized represent \he day oí the " inauguration'i.. trees. trail. We found two kinds of indirect evidence that could support the hypothesis of territorial defense: (a) the distribution of trails angularly covering a large (pig. 1 and 2), and (b) the angular orientation of abandoned could not be distinguished from'random distribution (Table 1 and Fig. 3). Even though we sampled only seven days, the probabiliy t of ant présence was great in a wide foraging area. A visible consequence of the rotation in the attack of trees and in the of trails is an unexpected presence of ants within the nest area, and a general t covering of the foraging terriory (pig. 1 and 3).. area. use. trees.
(3) FARlI & SIERRA: Leaf-QlltÍng ant strategies. 893. TABLE 1 Newly attacked (left) and Mwly abandoMd (right) trees per day and per trail. Too. 1 2 3 4. S. 6 Total. 4. S. 6. 7. 0/0 2/0 0/0 0/0 0/0 Ofl. 0/0 0/0. S/O. 210 0/1 O/O O/O O/O O/O. 0/0 0/0 0/1. 1/0 0/0 0/0 0/0 0/0 0/0. 17n. .2/1. 2/2. 1/1. 2/0. 2. 3. Ofl lfl Ofl 3/0. O/O l/O 3/0 2!2 2!2 211. 4/0 3/0. 16n. lO/S. Day 8/0. 4/1. lfl. 3fl 1(3. 1/0. 8. Total. 0/0. IS/0. OJO OJO. 0/0 1 /1. 10/2 614 7/6 3n 11 /S. 211. S 2/24. 1/0. Number of attacked trees/number of abandoned trees. More Iban 46% of Ihe total harvested trees were abandoned.. Since ant colonies are sessile it is an obvious advantage to dominate a wide foraging area, and to posses a maximize harvest panem that also minimizes competition (Traniello 1989). There are many demonstrations of aggression between ant colonies (Fowler 1977, Jutsum 1979, Vilela & Howse 1986). Therefore, we suspect that the defense of the foraging area. �.. /�r. plays an importa n t role in the nests development and maintenance in ecological and evolutionary time. The foraging effort is centered on trails ( Sheperd 1982) and the majority of these are conspicuous even though they are inactive during long periods (Gamboa 1975). For the nest studied, the attacked trees are highly angularly associated with the main trails (Fig 2). •. 11. ,, "'� ....... -_ .......... .,. .. ... Fig. 2. Angular distribution (S) of attacked trees per trail m. Attacked trees associated with each trail have different symbols, show a c1ustering in their angular disposition with regard to the main trail direction. Considering the mean v ectors (TI, T 2, etc. ) as a trails direction. a regular (random) distribution cannot be rejected (Rayleigh test P> .SS, Bastchelet 1981)... Fig. 3. Angular distribuuon of trees abandoned before they were defoliated during the study periodo Different symbols represent trees wilh different numbers of continuous days being attacked. A regular (random) distribution can nOl be rejected (Rayleigh test P= .2S, Batschelet 1981)..
(4) 894. REVISTA DE BIOLOGIA TROPICAL. TABLE2 A,.,lÚIJr distributioll of trails UtSOfM leafcuttUtg afÚS IlUI8 Autor. Ant spec:ie. Luao et al. (1973). Gamboa (1975). Olerrett (1968). Atta colombica. Acromyrmex versicolor. Atta cephalotes. S. 4. 4. meantrail. 7S.3. 76.8. 147.2. r. 0.40. 0.18. 0.34. 62.8. 73.8,. 6S.6. number of trails. Raylegh leIt (l'IIldam distribulion ). P=0.47S. 0,86< P <0.89. O.SS< P <O.S8. Data were obWned from nest maps of me papen cited. In a1l me cases regular disuibution (randommes) can nOl be rejected (Rayleigh test, Batscbe1et 1982).. Since both plants and colonies are sessile, the permanent trails would be acting as a "physical memory" showing the spatial location of patches that will be palatable in some period of the year. The temporal variations in their use would be following the qualitative variations of the resources. In this way we could explain the permanence of trails that apparently are not being used, bul would be in some perlod of the year. This pattem of apparen t "spatial stasis" and "temporal dynamism" associated with a pennanent search for new plants, would have important con se quences related to the defense of the foraging territory. The leafcutters'activity depends not only on a number of ptll"ameters of the "food resources" (quality," quantity, c hemical defenses), bul also on either the presence or the absence of alien foragers within the discovered patches (HOlldobler & Lumsden 1980). The resources previously occupied by competitors are clearly less attractive than the non visited ones (HOlldobler & Lumsden 1980). Therefore, the unpredictable presence of ants covering a large foraging area is obviously advantageous. On the other hand, our data about the angular rotation of attacked trees durlng the sampling period do not show a systematic movement. There are no studies about the eXÍSlence of a systematic rotation in the use of ttails and in the foraging cycles of leaf-cutting ants. and a few antecedents of rotation inant foraging columns (but see Franks & Fletcher. 1983).Since the leafcutler foraging area is seen as a nest centered sector system (HOlldobler & Lumsden 1980), activity on each trail could possibly be independent of the others over a short time perlod. Anl trails have more than one function (Traniello 1989)', and their existence is strong physical and chemical signs of an active colony. Generally, the disposition of trails covers the greatest part of the area around the leaf-cutting ant nest. This pattem can be seen in nests on other Attini species (Table 2), which could indicate a general pattem in leaf -cutting ants. In conclusion, a consequence of the partial defoliatio n of trees is lhe temporary abandonment of trails and the establishment of "new" ones, with a general and unpredictable ant presence in the foraging area. Therefore, we suggest that this rotation in the trail use may have an indirect but important role in the territorial defense.. ACKNOWLEDGENrnNTS This research was supported by a short-tenn fellowship to Alejandro G. Farji Brener from theSmithsonian Tropical Research Institute. We are grateful for the use of BCI Island facilities. The project and the manuscript benefited greatly from discussions with W. Eberhard D. Windsor, H. Fowler and J.Silva. •.
(5) FARn & SIERRA: Leaf-c:uuing ant strategies. REFERENCES. Batschelet, E. 1981. Circular Statistics in Biology. Academic Press, New York. Cherren, J.M . 19. 6 8. The foraging behavior of Alta cepluJlotes (Hymenoptera, Formicidae). 1: Foraging panems and plant species anacked in tropical rain foresL J. Anim. Ecol. 37: 387-403. Fowler, H.O. 1977. Some factors influencing colony spacing and s urvival in the grass -cutting ant Acromyrma /andoltifraticornis (Formicidae, Attini) in Paraguay. Rev. Biol. Trop: 25: 89-99. Fowler, H.O. & E.W. Stiles. 1980. Conservative resource management by leaf cutting anu? The role of Coraging, territories trails, and environmental patchiness. Sociobiology s: 25-41.. Appendix 1: Trail utilized, nest d istance, attack day, abandon day, and angular orientation of harvested trees by one adult Atta cepahlotes nest during six activity days.. Trail. Halldobler, B. and O. Lumsden. 1980. Territorial strategies in anU. Science 210: 732-739.. 1-1. 1-2a 1-2B. 1-2c 1-3 2. . Rockwood, L.L. 1977. Foraging panems and plant selectiOll in Costa Rica leaf-cutting anU. J.N.Y. Ent. Soc.89(4):222-233.. 21 15 24. 3 3-a. 3-b. Sheperd. J. 1982. Trunk trails and searching strategy of a leaf-cuning ant, Atta colombica. Behav. Ecol. Sociobiol. 11:77-84. Tranie11o, J.F.A. 1989. Foraging strategies oC ants. Ann. Rev. Entomol. 34:191-210. Vilela, E.F. & P.E. Howse. 1986. Territoriality in Leafcutting anu, Atta spp . 11I C.S. LoCgren & A.R.K. Vander Meer (eds.) Fire Anu and Leaf-Cutting Anu: Biology and Man agemenL Westview Studies in Insect Biology, LondOll and Boulder.. 36 37 42 47 42 17 18 19 2 11 28 24. Hubbell, S.P. & D.F. Wiemer. 1983. Host plant selection by an attine ant. 111 p. Jaison (ed.) Social Inscets in the tropics, v ol 2: 13S-1S4. Univ. of París Press, París, France;. Lugo, A .E., E.O. Famwonh, d. Pool, P. Jerez & O, Klaufman. 1973. The impact oC the leaf cutter ant Atta colombica on the energy flow oC a tropical wet Corest. ecoIogy S4: 1292-1301.. 6 10 15 23 23 25 26 34. Howard, J.J. 1990. Infidelity oC leaCcutting anU to host plants: resource heterogeneity or deCense induction? Oecología 82: 394-401.. Jutsum, A.R. 1979. Interspecific agression in leaf-cutting anu. Anim. Behav. 27:833-838.. Rl. R2 Thetha. TI. T2 total trees. Franks, N.R. & C.A. Fletcher. 1983. Spatial pattems in army ant foraging and rnigration: Eciloll burchelli 011 Barro Colorado Island, Panama. Behav. Ecol. Soclobiol. 12: 261-270. Oamboa, O.J. 1975. Foraging and leaf-cutting of the desert gardening ant Acromyrma versicolor versicolor (P.) (Hymenoptera, Forrnicidae). Oecología 20: 100-110.. 89S. 4. 29 29 32 4 27 40 40 30 32 27 32 45 45 45 50 83 63 83 83 50 25 40. 8 12 20 27 28 27 30 35 38 38 50 53 47 29 19 20 4 12 30 26 23 17 40 50 51 57 4 27 41 42 46 41 41 42 46 46 46 51 86 66 86 87 52 25 42. 340 10 15 347 348 332 332 332 335 328 355 346 14 300 295 290 270 260 244. 250 240 250 220 210 211 220 205 200 210 209 220 215 215 216 192 191 193 184 180 185 182 185 183 45 40. 1 1. 2. 2. 1 2 1 1 1 2 2 6 6 4 6. 5. 5 3 4. 6. 6 6 1 1. 8. 9 10 11 12 13. 1. 1 3 5 2 3 3 4 4 5 6 6 1 1 1 2 3 4 4 4 1 2 2 5 6 6. 1 2 3 4 5 6 7. 3 6 4 4 5 5 -. -. 4 4 5 5 5 3 3 -. -. -. -. 2. 14 15 16 17 18 19 20 21 22 23 24. 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45.
(6) 896. 5 6. 7 8. REVISTA DE BIOLOGIA TROPICAL. 9 17 24 16 10 7 5. 9 19 24 16 l3 7 5. 58 58 134 160 150 70 80. 2. 2 3 1 4 3 6. 6 6 6 -. -. 6 -. 46 47 48 49 50 51 52. R1 is the direct tree distance to the ant nest, and R2 the real distance in the field following the trail. Thetha is the angle respect to me north for each tree. TI is the frrst attack day, T2 the abandon day, and ( ) with ant harvest activity at the day 7. -.
(7)
Figure
Documento similar
A January effect is present in nominal terms, but it is negative in real and dollar adjusted returns.. The January effect present in nominal terms cannot be explained by
Here we have seen an absence of leptin in AF, gastric content, stomach, and umbilical cord at days 18 and 19 of gestation, followed by a sudden appearance at day 20 and a
The volume of material in a nest was related to the male’s contribution to nest building and, in agreement with our hypothesis, in a multiple regression analysis, male tail length
The distribution of berberine and sanguinarine was analyzed in roots, stems and leaves of mature Argemone mexicana plants, along with that of tran- scripts corresponding to
26 A dose of galunisertib 300 mg/day administered as 150 mg two times per day for 14 days on/14 days off treatment has been established in a phase II study as an appropriate
They split into two different nests if the number of living agents is equal to the maximum and the amount of food in the nest is larger than a given value.. A new nest is created in
n oteS on food - plant . — Bartsch (2003) mentions the larvae as being hidden amongst low plants in rocky habitats during the day, at ground level, often taken in vicinity of
Experimental infections in susceptible BALB/c mice treated with 2.5 mg/kg body weight/day once every other day for a total of 15 days showed that indotecan cleared more than 80% of
