Esquema hipotético de los eventos moleculares durante el proceso de preinfección en Colletotrichum spp. En el periodo de 24 a 72 hdi se observó colonización del patógeno, se expresaron los genes CUT1, GLS1 y CHS1; De 96 a 120 hdi se observaron hifas de infección secundaria y formación de acérvulos; Los niveles de transcripción de los genes CUT1, GLS1 y CHS1 estaban elevados.
INTRODUCCIÓN GENERAL
El Capítulo VI aborda la tercera pregunta de investigación que incluye un estudio básico-experimental enfocado en determinar los niveles de expresión de genes relacionados con la patogenicidad (CUT1, CHS1 y GLS1) de C. Diversidad morfológica, patológica y genética de las especies de Colletotrichum responsables de la antracnosis en papaya. Carica papayaL).
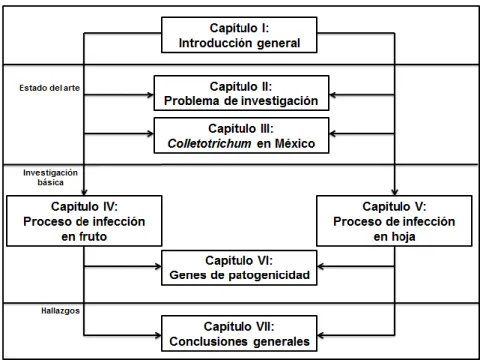
PROBLEMA DE INVESTIGACIÓN
El hongo mantiene un pH alto en el área de infección secretando amoníaco (Kramer-Haimovich et al., 2006). Durante las últimas etapas de la infección fúngica, las hifas invaden las células epidérmicas y del mesófilo, provocando la muerte de la célula huésped (Perfect et al., 1999). Por otro lado, Roberts y Snow (1984) describieron el proceso de infección de C. truncatum en su interacción con el algodón; observaron la germinación de conidios y la producción de apresorios en la cutícula cuatro horas después de la inoculación.
Las observaciones del proceso de infección revelaron penetración directa de la cutícula y colonización intramural, endofítica y necrotrófica.

PRIMER ARTÍCULO
Debido a su importancia científica y económica, la especie Colletotrichum ocupó el octavo lugar en la clasificación mundial de patógenos vegetales (Dean et al., 2012). Colletotrichum consiste en el estado asexual (anamórfico) de Glomerella (teleomorfo), perteneciente al phyllum Ascomycota, clase Hypocreomycetidae, orden Glomerellales, familia Glomerellaceae (Réblová et al. 2011). Por otro lado, Colletotrichum presenta diferentes morfotipos, como un micelio circular, de margen entero, de color gris, con masas conidiales de color naranja (Fig. 8E) (Torres-Calzada et al., 2012).
Para superar las deficiencias de la identificación tradicional, actualmente el uso combinado de la caracterización molecular junto con el diagnóstico tradicional representa una técnica apropiada para estudiar complejos en especies de Colletotrichum (Hyde et al., 2009b). La región más utilizada es el espaciador transcrito interno (ITS) del ADN ribosomal nuclear (Schoch et al., 2012). Se han reportado 82 especies de Colletotrichum en monocotiledóneas como maíz (Zea mays) (Álvarez, 1976), henequén (Agave fourcroydes) (Quijano-Ramayo et al., 2002), sorgo (Sorghum vulgare) (Álvarez, 1976) y dicotiledóneas como el plátano.
Por otro lado, Villanueva-Arce et al. fragariae en interacción con frutos de manzana. Por otro lado, Rodríguez-López et al. 2013) estudiaron el proceso de infección de C. gloeosporioides en su interacción con frutos de aguacate. Existen ejemplos de control biológico de antracnosis en manzana, papaya, aguacate, mango, entre otros (Carrillo-Fasio et al., 2005; Landero-Valenzuela et al., 2016).
Además, Patiño-Vera et al. 2005) desarrollaron un proceso de fermentación en planta piloto para la producción de levadura R. Disponible en línea: http://www.fungaldiversity.org/fdp/sfdp/FD39-1-E.pdf Hyde KD, Nilsson RH, Alias', Ariyawansa HA , Blair JE, et al.

SEGUNDO ARTÍCULO
Un aspecto importante del proceso de patogénesis es la estrategia de infección utilizada por el patógeno (PERFECT et al., 1999). Los tejidos permanecieron en las soluciones durante 3 horas y finalmente se agitaron dos veces en un procesador de tejidos centrífugo (Thermo Scientific, STP 120, GER) (CASARRUBIAS-CARRILLO et al., 2002). Las especies de Colletotrichum forman apresorios melanizados para facilitar la adhesión y penetración, mediante acción mecánica y lisis enzimática (KLEEMANN et al., 2012).
En los apresorios, se observó una mancha que carecía de melanina, lo que sugiere un poro donde se requiere que ocurra la segregación enzimática durante la penetración (RANATHUNGE et al., 2012). En el chile, la cutinasa es esencial para provocar la degradación de la cutícula (AUYONG et al., 2015). La producción de polifenoles está relacionada con la limitación del crecimiento de los hongos en el sitio de la infección (VARGAS et al., 2012).
Por otro lado, en este estudio no fue posible observar hifas debajo de la cutícula como en el estudio de Diéguez-Uribeondo et al. 2005); Sin embargo, se observaron hifas intramurales en las paredes de las células epidérmicas del huésped (Figura 11C), por lo que se puede inferir que las hifas de infección cruzaron la capa subcuticular a lo largo de la pared celular de la epidermis (RANATHUNGE et al., 2012) . Según Pring et al. 1995), las hifas intramurales crecieron en las paredes celulares de las células epidérmicas. En esta etapa, el patógeno obtiene nutrientes del contenido celular del huésped, por lo que la infección se asocia principalmente con la producción de enzimas líticas como proteasas y enzimas activas en carbohidratos que degradan los polisacáridos de las paredes celulares (O'CONNELL et al. , 2012). ).

TERCER ARTÍCULO
Resumen Colletotrichum truncatum es un hongo patógeno que causa antracnosis en una variedad de huéspedes; Se desconoce el proceso de infección de este patógeno en hojas de papaya, así como el comportamiento de la misma cepa C. El objetivo de este estudio fue describir el proceso de infección de la cepa C. Colletotrichum truncatum se mediante técnicas histológicas. como patógeno intramural subcuticular en papaya y como hemibiotrofo intracelular en guisante; Por tanto, la estrategia de infección depende del huésped.
Esta especie utiliza diferentes mecanismos de infección, comportándose como un patógeno hemibiotrófico intracelular en guisante (Pisum sativum), haba (Vicia faba) y lenteja (Lens culinaris) (O'Connell et al., 1993; Latunde-Dada y Lucas, 2007) y como patógeno intramural subcuticular en chile (Capsicum annuum), garbanzo (Cicer arietinum), maní (Arachis hipogaea) y frijol (Phaseolus vulgaris) (Pring et al., 1995; Ranathunge et al., 2012). Las primeras hifas de infección emergieron de los apresorios entre las 18 y 20 h, a diferencia de lo que sucedió en las hojas de papaya (Fig. 14A). Aunque había hifas de infección primaria en la luz celular, no se observó lisis.
La característica principal de la invasión intracelular es la formación de hifas de infección primaria; Estos pueden tener diferentes morfologías dependiendo de la especie (O'Connell et al., 2000). Primera entrada de infección (HI) que emerge de los apresorios (a). b) Sección longitudinal (paradérmica) de la hoja a 36 HDI. Hifas de infección (HI) que emergen de los apresorios (A), así como hifas primarias lobuladas (HP) delimitadas por la pared de la célula huésped (PC).

CUARTO ARTÍCULO
Estos genes están relacionados con la producción de estructuras de infección y el desarrollo del patógeno durante el proceso de infección en hojas de papaya Maradol, lo que sugiere que la síntesis de quitina y β-1-3 glucanos está relacionada con la etapa de producción de estructuras de infección. , conidios y acérvulos) mientras que la cutinasa desempeña una función secundaria probablemente para la lisis de la cutícula del huésped. Se han estudiado genes relacionados con la patogenicidad en especies de Colletotrichum; Durante el proceso de infección, el patógeno requiere estructuras especializadas como apresorios e hifas de infección para llevar a cabo la infección con éxito (O'Connell et al., 2012). En hongos patógenos es importante la regulación de la formación de polímeros de la pared celular, así como el cambio estructural durante la morfogénesis relacionado con el proceso de infección; La β-1-3 glucano-sintasa (GLS1) es necesaria para la rigidez.
La expresión de este gen coincide con las etapas de germinación y formación de apresorio (etapa de penetración) durante el proceso de infección de C. 2015) reportaron la expresión del gen de la cutinasa en la interacción C. Según Rojo-Báez et al. 2016), durante el estado necrotrófico del proceso de infección de C. La expresión de GLS1 está relacionada con la patogenicidad en Colletotrichum graminicola, ya que fue sobreexpresada durante su interacción con el maíz a los 12 y 72 hdi, lo que coincide con el período temprano de infección donde La síntesis de β-1-3 glucanos son indispensables en la rigidez de la pared celular del apresorio, la esporulación asexual, la adhesión y diferenciación del apresorio funcional, así como para el rápido crecimiento de las hifas de infección secundaria durante el estado necrotrófico (Oliveira-García et al. al., 2013).
En este estudio, la expresión de GLS1 durante las últimas horas de la infección (120 hdi) (Fig. 16B) sugiere que el patógeno requiere la producción de β-1-3 glucanos durante el estado necrotrófico para la producción de estructuras de infección como las hifas. de infección y conidios. Aunque la expresión del gen quitina sintasa durante las últimas horas de la infección (120 hdi) no fue estadísticamente significativa, se observó un marcado aumento en los niveles de transcripción (Fig. 17B), que probablemente esté relacionado con la necesidad de garantizar la estabilidad en la célula. Pared de hifas, acervulas y conidias de infección secundaria. En este estudio se analizaron los niveles de expresión temporal relativa de varios genes de Colletotrichum truncatum relacionados con la producción de estructuras de infección y el desarrollo del patógeno durante su interacción con las hojas de papaya Maradol.

CONCLUSIONES GENERALES
Integrando los resultados de los objetivos propuestos podemos conocer el proceso de infección de C. Las etapas de adhesión, germinación y penetración se realizaron durante las primeras horas de la inoculación (2 a 24 hdi). En esta etapa se observó la germinación de conidios a través de tubos germinales, así como la producción de apresorios melanizados; Además, se sobreexpresaron los genes de la β-1-3 glucano sintasa y de la quitina sintasa, que codifican la formación de importantes polímeros de la pared celular como los β-1-3 glucanos y la quitina, que podrían haber permitido al patógeno formar tubos germinales y producir una pared celular funcional para la apresoria.
En esta etapa también comenzó la expresión del gen de la cutinasa; donde el patógeno logró penetrar exitosamente la cutícula de papaya con actividad enzimática. En el periodo de 24 a 72 idh; Se observa colonización con el patógeno, en esta etapa la expresión de los genes CUT1, GLS1 y CHS1 indica la cooperación de las enzimas cutinasa, β-1-3 glucano sintasa y quitina sintasa, para que el patógeno pueda desarrollarse y dispersarse a través de las hifas. infección. Finalmente, se observaron hifas de infección secundaria y formación de acérvulos entre 96 y 120 hdi; En esta etapa, la colonización y reproducción de C.
En el caso de los genes GLS1 y CHS1, forman estructuras de infección como hifas, acérvulos y conidios de infección durante el estado necrotrófico del proceso de infección. Por otro lado, se determinó el nivel de expresión génica de tres genes relacionados con la patogenicidad en esta investigación; Sin embargo, una limitación del estudio fue que no se realizó ni transformación del vector ni silenciamiento génico para demostrar definitivamente la participación de los genes en el proceso de infección de C.








