Este trabajo aborda la biogénesis de lactonas sesquiterpénicas tipo guayanolida a partir de germacranólidos, la transformación de germacranólidos mediante la reacción de Cope y la transformación de humuleno en un sesquiterpeno tricíclico. También se encontró que el anillo de lactona desestabiliza las emanólidos pero cambia muy poco los estados de transición del reordenamiento de Cope.
Introducción
Por ello, no es de extrañar que exista mucho interés por el estudio de los productos naturales en México. La química de los productos naturales ha cambiado significativamente desde la segunda mitad del siglo XX.
Antecedentes Generales
El estudio de los SEP es muy importante en química computacional, porque en esta superficie existen posiciones de átomos de moléculas que minimizan la energía, que no es más que geometría de equilibrio. En general, no se puede visualizar el SEP completo, ya que esta superficie se encuentra en el espacio dimensional N+1, donde N son los grados de libertad de los núcleos de la molécula.

Objetivos
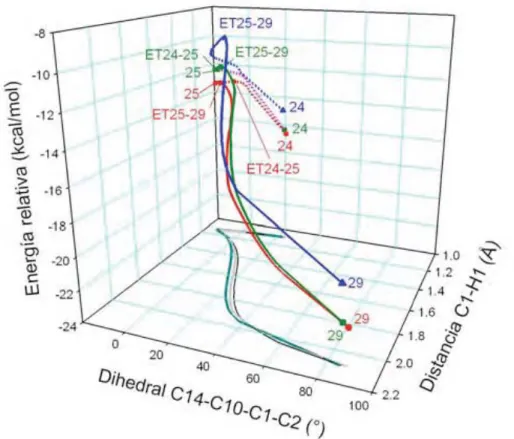
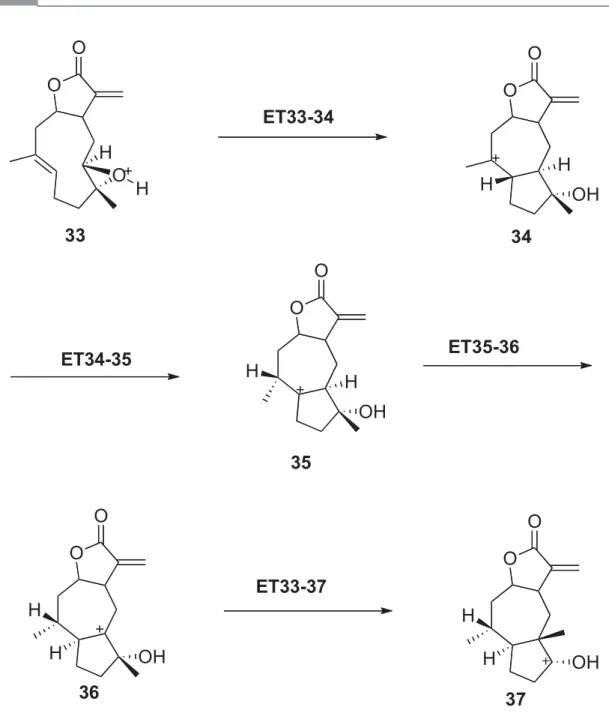
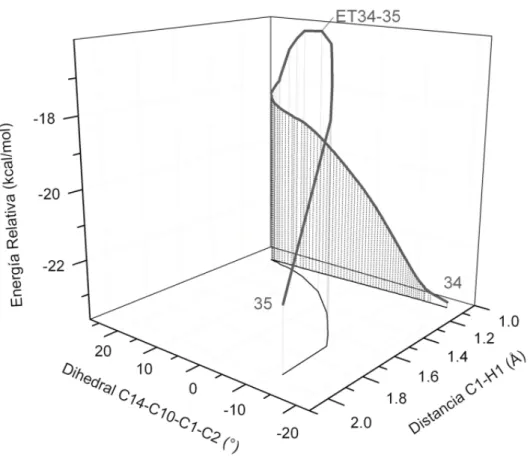
Estudio del Mecanismo de Reacción de la Biogénesis Terminal de la 8-Epiconfertina
Proyección en el plano xy de la coordenada de reacción para los funcionales mPW1B95 (Gris claro), B3LYP (Gris oscuro) y M05-2x (negro). Una de las pequeñas diferencias es el hecho de que la ET para la migración del segundo hidrógeno (ET35-36) no es tan simétrica como en el caso del epímero S.

Estudio conformacional de la 6-epi- desacetil-laurenobiolida
Esto significa que la velocidad de reacción debe estar entre los valores de 100 y 104 s-1, o lo que es lo mismo, la magnitud de la energía de activación de la interconversión debe mantenerse en el orden de 4 a 25 kcal/mol. 121. Para calcular G‡ y H‡ se realizó un análisis de la forma de las líneas de los espectros de RMN a las diferentes temperaturas con el programa gNMR.125 Solo se tomaron en cuenta temperaturas cercanas a la temperatura de coalescencia, porque en esta área donde se reducen los errores en el cálculo de k. Con la molécula 21 podemos estimar la energía resultante de la tensión anular en el segmento en cuestión.
En el espectro a -20 °C en tolueno-d8 se encuentra una situación similar, con la diferencia de que en este caso se producen desplazamientos diamagnéticos y paramagnéticos debido al carácter aromático del disolvente y cómo. Por ejemplo, C1, uno de los átomos de carbono que rota en el equilibrio conformacional y por tanto ve cambiado su entorno químico en cada uno de los confórmeros. En comparación, la barrera conformacional en el ciclohexano es del orden de 10 kcal/mol.71 La barrera de energía cuando se intercambia 17 con el intermedio 22 es de 18,39 kcal/mol, lo que hace que la energía sea mayor.
En el caso de G, la diferencia entre el experimental y el calculado también es muy pequeña, aunque en este caso es mucho más importante porque el valor de G es muy pequeño.
Estudio de la transposición de Cope en germacranólidas con geometrías Z,E y E,E
El otro posible producto de reordenamiento de Cope de 1 es el epímero C5 de 5(5', figura 6.3), que tendría un producto estereoquímico de un Cope normal (ET en forma de silla). En esta propuesta se afirma que la melampolida se isomeriza a la correspondiente germacranolida y es esta la que realiza el reordenamiento de Cope. Luego se propuso otro mecanismo de reacción en el que primero se produce la hemiacetalización y luego se produce el reordenamiento de Cope.
Los resultados en la Tabla 6.1 indican que la barrera energética de la hemiacetalización optimizó la ET y las estructuras intermedias de la transformación 1 a 2 (rojo) y 6 a 2 (verde) a través de un mecanismo en el que el reordenamiento de Cope es posterior a la hemiacetalización. El elemanolida 11 producto de la reordenación de Cope 9 y 10 parece tener una energía mucho más cercana a la melampolide cuando el anillo de lactona está abierto, ya que la diferencia de energía entre 1 y 5 es 6,8 kcal/mol, mientras que entre 9 y 11 es . sólo 2,3 kcal/mol. De lo anterior se concluye que el efecto del anillo de lactona de 5 miembros sobre el reordenamiento de Cope de los germacradienos es el mismo.
Así que una vez más llegamos a la conclusión de que el anillo de lactona de 5 miembros desestabiliza el elemanolida y no el Cope ET.

Estudio del Mecanismo de Reacción de la Biogénesis Terminal del Sesquiterpeno
En 1982 se aisló el ácido terrecíclico (6) del hongo Aspergillus terreus174, unos años antes se aisló de la misma fuente natural el quadrone (9, Esquema 7.3), que tiene una estructura muy similar al 6.175 Ambos compuestos, además, aunque Tienen una actividad antitumoral moderada, tienen una estructura tricíclica muy interesante y su origen biogenético no fue fácil de dilucidar. Varios grupos de investigación han trabajado para dilucidar la biogénesis de 6. Para ello, han incorporado acetato y mevalonato marcados isotópicamente en cultivos de Aspergillus terreus.176-179 El esquema 7.3a muestra que al incorporar [1-13C]-acetato los átomos de carbono que tienen su señal aumentada en el espectro de RMN y 11, mientras que el acetato de [1,2-13C2] contiene todas las señales. El mecanismo propuesto por Coates et al. (esquema 7.5) supone un carbocatión cercano en geometría al -cariofileno y, por lo tanto, se eligió 1f como confórmero inicial.
El mecanismo propuesto en el Esquema 7.5 muestra que en el intermedio 16 el hidrógeno y el metilo unidos a C2 y C3, respectivamente, tienen la misma dirección. Luego se propuso que el primer paso sería la rotación del enlace C2-C3 para producir el intermedio 1f' (Figura 7.3), en el que el hidrógeno y el metilo miran en la misma dirección. En el ET que conecta los intermedios 1f' y 16 (ET1f'-16), se puede observar cómo la distancia C2-C6 disminuye al mismo tiempo que se transpone C1 (Figura 7.3), por lo que se puede hipotetizar que la formación de los El enlace C2-C6 y la transposición del enlace C1 ocurren en conjunto.
Si comparamos ambos mecanismos desde un punto de vista energético (Figura 7.4), podemos concluir que el mecanismo propuesto por Cane et al.

Conclusión General
D.; Canada, J.; Jimenez-Barbero, J.; Cuevas, G., Theoretical study of the inversion and topomerization processes of substituted cyclohexanes: Importance of the 3D energy hypersurface. Tori, K.; Horibe, I.; Tamura, Y.; Kuriyama, K.; Tada, H.; Takeda, K., Re-investigation of the conformation of laurenobiolide, a ten-membered sesquiterpene lactone by variable-temperature carbon-13 NMR spectroscopy. Catalyzed cyclizations of germacranolides - Molecular structure of the cyclization products of gallicin and 8-alpha-hydroxygallicin (Shonachalin-a).
Tori, K.; Horibe, I.; Tamura, Y.; Tada, H., Simultaneous application of the nuclear Overhauser effect and an NMR shift reagent. Preparation of cyclodeca-cis,trans-diene derivatives and their Cope rearrangements.Journal of the Chemical Society, Perkin Transactions 1: Organic and Bio-Organic Chemistry. J.; Piecuch, P., Balancing dynamic and nondynamic correlation for diradical and aromatic transition states: A pairwise renormalized study of the facing rearrangement of 1,5-hexadiene.
Journal of the Chemical Society, Perkin Transactions 1: Organic and Bioorganic Chemistry Takeda, K.; Horibe, I; Minato, H., Dealing with rearrangement of some germacrane-type furan sesquiterpenes.
Apéndices
Terminal Biogenesis of 8-Epiconfertin
In this article we approach the study of the ring closure that the germacranolides (2) suffer to produce guaianolides (6, graph 1). The ability of the hybrid density functional theory (DFT) methods: B3LYP,41, B97-2,42B1B9543, and mPWB9544functionals and the MP245 method to describe the potential energy surface associated with these compounds (PES) was also evaluated. Even if many changes occur in the molecular structure of the compounds involved, the activation energy is low (Table 2).
In the case under study, compound 9 (Scheme 2) arises through the loss of one of the. The structural difference between the two groups lies in the stereochemistry of the methyl group at position C10. The absolute stereochemistry of the stereogenic center supporting the methyl group in the ambrosianolides is Swhile in helanolides is R (graph 3).
A higher barrier is for the migration of the [1,2]-methyl group with the formation of the carbonyl group, the last step in the biogenesis of pseudoguanolide.
Sesquiterpenes: The Case of 8-Epiconfertin
Introduction
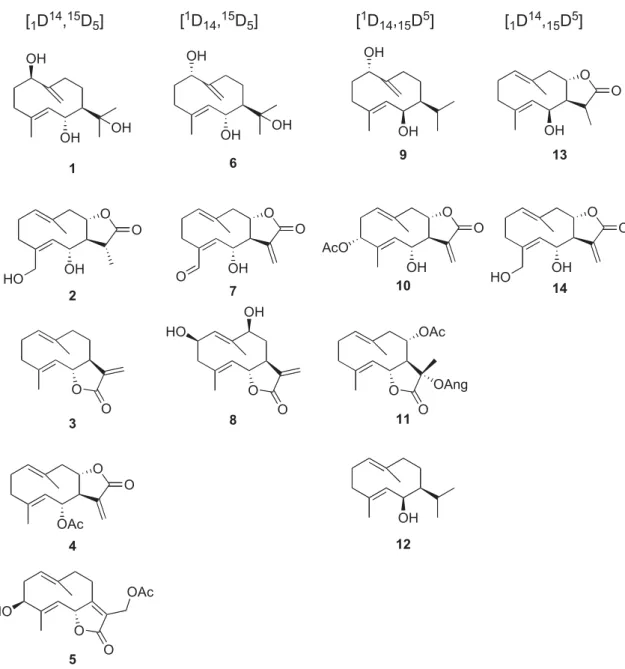
DQGWHUSHQRLGVLQVXSHULRURUJDQLVPVZKLOHGHR[\'[\OXORVHSKRVSKDWHLVXVHGE\OHVVHU RUJDQLVPV>@2QWKHRWKHUKDQGDOOWKHVHVTXLWHVHRVKVD HLURULJLQLQHLWKHUIDUQHVROVSHFLILFDOO\WKHIDUQHV\OS\URSKRVSKDWH)3316FKHPHRU QHUROLG\OS\URSKRVSKDWH>@. 7KHILUVWELRJHQHWLFK\SRWKHVLVWRH[SODLQWKHRULJLQRIWHUSHQHVLV5X]LFND¶VUXOHRI. LVRSUHQH>@$FFRUGLQJWRWKLVUXOHWKUHHPROHFXOHVRIDFHW\OFRHQ]\PH$DUHVXFFHVVLYHO\. FRQGHQVHGWRSURGXFHK\GUR[\PHWK\OJOXWDU\OFRHQ]\PH$WKURXJKWKHDFWLRQRI+0*&R$. V\QWKHWDVHWKDWWKURXJKUHGXFWLRQJHQHUDWHVPHYDORQLFDFLG$GHFDUER[\ODWLRQHOLPLQDWLRQ UHDFWLRQJHQHUDWHVLVRSHQWHQHVHVPHYDORQLFDFLG$GHFDUER[\ODWLRQHOLPLQDWLRQ UHDFWLRQJHQHUDWHVLVRSHQWHQWHVPHYDORQLFDFLG$GHFDUER LPHWK\ODOO\O. S\URSKRVSKDWHWKURXJKWKHSDUWLFLSDWLRQRILVRSHQWHQ\OS\URSKRVSKDWHLVRPHUDVH>@7KHODWWHU WZRFRPSRXQGVDOORZWKHFRQVWUXFWLRSHQWHQ\OS\URSKRVSKDWHLVRPHUDVH>@7KHODWWHU WZRFRPSRXQGVDOORZWKHFRQVWUXFWLRSHQWHQVRQVRQVLVRQVLVRQVRQVRQVLVRVLVR DOOHG KHPLWHUSHQHVGLUHFWO\GHULYHIURPDQ\RIWKHWZRFRPSRXQGVZKLOHVHVTXLWHUSHQHVKDYHWKHLU RULJLQLQWKHFRQGHQVDWLRQRIWKUHHPOHLHQVDVDF OORZVWKH ELRV\QWKHVLVRIIDUQHV\OS\URSKRVSKDWH16FKHPHRURILWVLVRPHUQHUROLG\O. 1RZDGD\VVHYHUDOIXQGDPHQWDODVSHFWVRIWKHPHYDORQDWHDQGQRQPHYDORQDWHSDWKZD\VVXFK DVWKHHQ]\PHVWKDWFDWDO\]HWKHLUWWUDQVGHVRQVRQVRQVRQVR UWKHPDUHZHOO NQRZQ>@7KHWHUPLQDOELRJHQHVLVRIWKHGLIIHUHQWQDWXUDOSURGXFWVUHVSRQVLEOHIRUWKHJUHDW VWUXFWXUDOGLYHUVLW\RIFWRQVGDH\RIFWRQVGDHV\ HUHDUHVWLOOPDQ\XQDQVZHUHG TXHVWLRQVRQWKHPHFKDQLVPVUHVSRQVLEOHIRUJHQHUDWLQJVXFKDYDULHW\RIPROHFXOHV>. amp;KHPLFDOVLPXODWLRQRIELRJHQHWLFSURFHVVVHVLVEDVHGRQWKHIDFWWKDWWKHUHDFWLRQVWKDWWDNH SODFHin vivoIROORZWKHVDPHJHQHUDOSULQVIQVQVRWLVit@ KLV FRQFHSWRQHFDQGHPRQVWUDWHWKDWLQPDQ\FDVHVDFRPSOH[HQ]\PDWLFV\VWHPLVQRWUHDOO\. Q]\PHVVHHPIXQGDPHQWDOIRUWKHVHOHFWLRQRIWKHVWDUWLQJFRQIRUPHUDQGH[SODLQWKHKLJK FKHPLFDO\LHOGVRIWKHVHWUDQVIRUPDWLRQVQWHRHRHVRH FSURFHVV FDQEHVLPXODWHGin vitro.>@. FRQILJXUDWLRQ&(&(66FKHPHWKHKHOLDQJROLGHV7&(&=WKHPHODPSROLGHV 8&=&(DQGWKH==JHUPDFUDQROLGHV97KLVZDVGHPRQVWUDWKLVRGVRQVWUDWKLVRGD XUSRVVLEOHFRQILJXUDWLRQV>@&RVWXQyOLGH>@10DQGWDPDXOLSLQ>@11 DUHH[DPSOHVRIE,EJHUPDFUDGLHQHIUXWHVFLQ>@12DQG6FKNXULROLQ>@12DQG6FKNXULDXOLSLQ QRELOLQ>@14DQGHXSDIRUPRQLQ >@15DUHH[DPSOHVRI&(&=.
Reaction Mechanism
GLVWDQFHLVWKHVDPH7KHWUDQVLWLRQVWDWHIRUWKHHOHFWURSKLOLFFORVXUHDVVRFLDWHGZLWKHDFK FRQIRUPHU34DQG44UHVSHFWLYHO\VKRZVWKHGHFUHDVHRIWKHtransDQQXODUGLVWDQFHDQGWKH LQ FUHDVHRIWKH&2+GLVWDQFHWKDWHQGVLQWKHFLVIXVLRQSURGXFW35ZLWKD&&GLVWDQFHRI cDQGWKHtransIXVLRQSURGXFW45ZLWKD&&GLVWDQFHRIc7KLVLVWKHILUVW HOHPHQWDOVWHSLQWKHPHFKDQ LVPZLWKSURGXFW45EHLQJPRUHVWDEOHWKDQ35E\DGLIIHUHQFHRI NFDOPRO7KHWUDQVLWLRQVWDWHVIRUWKHtransDQQXODUFORVXUHGLIIHUEDUHO\E\NFDOPRO 7KHORZHUVWDELOLW\RI35LQUHODWLRQWR45FD QEHDVVRFLDWHGWRWKHIDFWWKDW35SUHVHQWVDcis IXVLRQZKLFKLPSOLHVDSDUWLDOHFOLSVLQJRIWKH&&&&IJ +&&+IJ DQG&&&&IJ VHJPHQWV,QDGGLWLRQWKHPHWK\OJURXSMRLQHGWR &DQDWRP WKDWKDVORVWLWVFKDUJHDGRSWVDQDUUDQJHPHQWFORVHWRPHWK\OHQH&7KLVDUUDQJHPHQWZRXOG EHH[SHFWHGWREHPRUHHQHUJHWLFWKDQWKDWRI45ZKHUHWKHPHWK\OLVRULHQWHGLQDQ RSSRVLQJ GLUHFWLRQUHVSHFWWRWKH&PHWK\OHQHDQGWKHVHJPHQWVDVVRFLDWHGWRWKHIXVLRQPDLQWDLQDQanti GLVSRVLWLRQ2QWKHQH[WVWDJHWKHUHDFWLRQPHFKDQLVPVDUHGLIIHUHQWDQGZLO OQRWEHVLPLODU. SHUSHQGLFXODUWRWKHSODQHRIWKHUHIHUUHGDWRPV7KXVIRU35WKH+&&&DQJOHLVRI DQGRIIRU455HFHQWO\LWZDVGHVFULEHGWKDWWKHXQGHUWDNLQJRIFRQIRUPDWLRQDODQDO\VLV UHTXLUHVFRQVLGHULQ JWKHFRQIRUPDWLRQDOSURFHVVDVDVHTXHQFHRIHOHPHQWDU\FRQIRUPDWLRQDO VWHSV>@+HQFHLWLVQHFHVVDU\WRHVWDEOLVKWKHZD\LQZKLFKWKHVWDWLRQDU\VWDWHVRI PLQLPXPHQHUJ\DUHLQWHUFRQQ HFWHGWKURXJKWKH767KLVLVIHDVLEOHIRUFDWLRQ35ZKLFKLV WUDQVIRUPHGLQWRFRQIRUPHU37WKURXJK7636)LJXUH,QWKLV76WKHDQJOH&&&&. UHDFKHVDYDOXHRI7KHVWDELOL]DWLRQRIFDWLRQ36LVGXHWRK\SHUFRQMXJDWLRQ>@E\WKH SDUWLFLSDWLRQRIWZRV&&ERQGV&&DQG&&DQGDV&+ERQGRIWKHPHWK\OJURXS>@$W P3:*GSOHYHOWKHGL VWDQFH&&LVcWKH&&RIcDQG&+LV cODUJHUWKDQ&+GLVWDQFHRIWKHRWKHUK\GURJHQDWRPVRIWKHPHWK\OJURXS7KH1%2 DQDO\VLV>@RIWKHVHV&+oSLQWHUDFWLRQVHVWDEOLVKHVYDOXHVRIDQGNF DOPRO UHVSHFWLYHO\IRUHDFKLQWHUDFWLRQ.
VWDWLRQDU\VWDWHVWKH\GRUHFRJQL]HWKHIDFWWKDWWKHILUVWVHJPHQWRIWKHUHDFWLRQFRUUHVSRQGVWR WKHFRQIRUPDWLRQDOFKDQJHWHWHWHWHVHVHVHVHVHVHQVHVHVHQVHVHVHVHVHVHVHVHVHVHVHVHVHVHVHVHVHQ HFWURQLFUHTXLUHPHQWVDUH VDWLVILHGDQGWKHVHFRQGVHJPHQWFRUUHVSRQGVWRWKHWUDQVIHURIDK\GURJHQDWRP(YLGHQWO\D WUDMHFRQGVHJPHQWFRUUHVSRQGVWRWKHWUDQVIHURIDK\GURJHQDWRP(YLGHQWO\D WUDMHFRQGVHFUHD WWKHVDPHWLPHDVWKHFRQIRUPDWLRQDOFKDQJH VKRXOGEHGLVFDUGHGEHFDXVHLWUHTXLUHVDUHDFWLRQWUDMHFWRU\WKDWLVFRPSOHWHO\GLIIHPHWHFHKVHV QJHVLQWKHGLVWDQFHRIWKH&+ERQGZRXOGKDSSHQDWWKHVDPHWLPHDV WKHFKDQJHVLQWKH&&&&DQJOHZKLFKLVQRWREVHUYHGKHUH7KWKLVKLVFDFDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDVDFDVDFDVDFDVDFDVDVDVDVDFDVDFDVDVDFDFDFDFDFDVDFDFDFDFDFDFDFDFD. HOHWLRQHQHUJLHVREWDLQHGE\WKH1%2DQDO\VLVE\WKHLQWHUDFWLRQRIV&+ZLWK WKHHPSW\RUELWDORIWKHFDUEFDWLRQWKDWJRHVIURPWRNFHQWDOPHQVRNFQLQWDOP HULSODQDUWR&2QWKHRWKHUKDQGWKHVWDELOL]DWLRQRI43LVQRWSURGXFHGIRU WKLVUHDVRQVLQFHWKHLQWHUDFWLRQV&&ERQGZLWKWKHHPSW\RUELWDORIWKHFDUERFDWLRQ.
Conclusions
Biochemical 6\PSRVLD 1R $FDGHPLF 3UHVV1970 0DF0LOODQ -Recent Aspects on the Chemistry and Biosynthesis of Gibberellins, QRec. 2 it is also known that the configuration of the transring cyclization depends on the conformation of the precursor. However, this freedom is limited due to the endocyclic double bonds and the lactone ring fused to the cyclodecadiene.
The stabilization must thus be due to interaction of the double bond with the neighboring groups directly linked to it, i.e. in this segment of the molecule the energy difference must be due to the ring stress (see above). In this way, the energy differences between 12/13 and 14/15 must be due to the balance between the ring stress and the trans-annular interaction.
The energy difference could therefore be due to the trans-annular interaction of the double bonds.