Mostraremos a dinâmica dessas estratégias ao jogar DP repetidos utilizando noções básicas de teoria dos jogos e análise qualitativa de equações diferenciais ordinárias no plano. Palavras-chave: Evolução da cooperação, Dilema do Prisioneiro infinitamente repetido, estratégias reativas, equação replicadora, equilíbrio de Nash. Mostraremos a dinâmica dessas estratégias ao jogar DP repetidos utilizando noções básicas de teoria dos jogos e análise qualitativa de equações diferenciais ordinárias no plano.
![Figura 1: Resultados dos experimentos de Nowak e Sigmund, figura extra´ıda de [20].](https://thumb-us.123doks.com/thumbv2/123dok_es/12434695.0/8.892.251.644.374.819/figura-resultados-experimentos-nowak-sigmund-figura-extra-ıda.webp)
Matriz de pagamento
Supõe-se que tal situação se repete com uma frequência suficientemente elevada e que a recompensa para cada indivíduo que escolhe uma determinada estratégia é a sua aptidão ou “aptidão”, ou seja, o número de descendentes que terá na próxima geração tardia. . Cada descendente utiliza a mesma estratégia de seu ancestral e, para simplificar, a reprodução é considerada assexuada. A teoria dos jogos evolutivos não se baseia, portanto, na racionalidade dos seus jogadores, mas considera uma população de jogadores que interagem aleatoriamente no jogo e onde cada jogador participa com uma estratégia fixa.
Equil´ıbrios de Nash
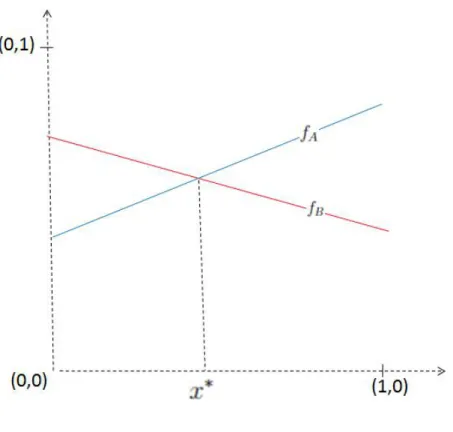
Uma estratégia que é um equilíbrio de Nash é a melhor resposta para si mesma, ou seja, Uma estratégia que é um equilíbrio de Nash estrito é a melhor resposta para si mesma, ou seja, Esta definição diz-nos que uma estratégia q∈Sn é um equilíbrio de Nash se, quando escolhida por dois jogadores, nenhum deles se arrepende de saber que o seu oponente a escolheu.
Equa¸c˜ ao do Replicador caso n = 2
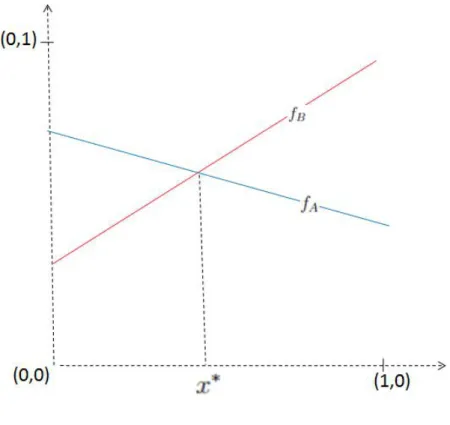
Se nenhuma das estratégias AnemB for equilíbrio de Nash, existe a solução de equilíbrio x∗ ∈(0,1). Se ambas as estratégias A e B são equilíbrios estritos de Nash, existe a solução de equilíbrio x∗ ∈(0,1). Se apenas a estratégia B for equilíbrio de Nash (estrito ou não), então não existe solução de equilíbrio x∗ ∈(0,1).
Equa¸c˜ ao do Replicador caso geral
Uma estratégia reativa [14, 12] no dilema do prisioneiro iterado é tal que a escolha de C ou D por um jogador depende unicamente da escolha do oponente na rodada anterior e é determinada por um par da forma (p, q), com p , q ∈ [0,1]. Se um jogador com estratégia S(p, q) enfrenta um adversário com estratégia ˆS(ˆp,q), seu nível de confiança cn determina o nível. O retorno em estado estacionário para a estratégia S(p, q) versus S(ˆˆ p,q), no Dilema do Prisioneiro infinitamente repetido, é dado por.
An´ alise assint´ otica
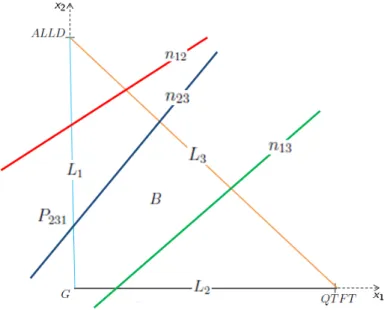
Regi˜ ao de interesse e pontos de equil´ıbrio
P231: É o ponto de equilíbrio onde as estratégias G e ALLD coexistem na ausência de QT FT. Nosso objetivo é provar a existência e relevância biológica dos pontos de equilíbrio para a equação do replicador. Veremos que variando qem (0,1] os pontos de equilíbrio podem entrar ou sair da região biológica B.
Portanto, a partir de qgreen haverá uma faixa de valores q em que existirão os pontos de equilíbrio biologicamente relevantes P123, P132 e P231. É importante notar que nenhum dos pontos de equilíbrio biologicamente relevantes que existiam no intervalo anterior (qblue, qred) foi perdido no intervalo (qred, qblack). É importante notar que para grandes valores de q temos os pontos de equilíbrio biologicamente relevantes P123 no lado L3, P132 no lado L2 e Q dentro da área B e que esta situação persiste para qualquer valor de q até a partir de qblack , isso é ∀q ∈ (qblack,1].
Aumentando ainda mais o valor de q, alcançamos o valor qblu e assim a reta n23 passa pela origem do plano (x1, x2), e assim o ponto de equilíbrio P231 não é mais biologicamente relevante. Neste ponto dos valores de q temos os pontos de equilíbrio biologicamente relevantes P123 no lado L3, o ponto de equilíbrio P132 no lado L2 e o ponto de coexistência das três estratégias Q dentro da região aoB. Observemos que, neste caso, o ponto de equilíbrio de coexistência biologicamente relevante Q "aparece" para valores "pequenos" de q no lado L1.
Também é importante notar que para grandes valores de q temos pontos de equilíbrio biologicamente relevantes P123 no lado L3, P132 no lado L2 e Q dentro da região B e que esta situação permanece ´em para qualquer valor de q de qgreen , ou seja, ∀q∈ ( qverde,1).
Alguns resultados ´ uteis
Nenhum outro ponto de equilíbrio é biologicamente relevante, mas é importante notar que neste caso os outros pontos de equilíbrio coincidem no vértice E2 da região B. d) Para grandes valores de q, ou seja, q > qverde, temos o biológico pontos de equilíbrio relevantes P123 no lado L3, P132 no lado L2 e o ponto de coexistência das três estratégias Q dentro da região B, mas o ponto de equilíbrio P231 não é mais biologicamente relevante. Pelo argumento dado acima, o resultado 3.12 segue se tomarmos o máximo da expressão (∗) como K4 e o mínimo como K3, sendo ambos positivos devido às relações entre os parâmetros T, R, T e S. Portanto, se r é pequeno o suficiente, temos:. O próximo corolário afirma que o crescimento do insumo a13(q) da matriz de pagamentos 3.3, visto como uma função de q, não é apenas positivo como mostrado no Teorema 3.4.7, mas é inicialmente “muito grande”, e para valores grandes de q “muito pequeno”, esse comportamento é o mesmo na entrada a31.
No Teorema que apresentamos a seguir, determinamos a sequência no lado L3 dos pontos P123, P233 e P133, que será de extrema importância na determinação de quando o ponto de equilíbrio de coexistência das três estratégias se torna biologicamente relevante. Vamos estudar as raízes desta equação quadrática: 3.18) Por fim, para completar esta seção, apresentaremos resultados que definem os parâmetros qgreen, qblue, qred e qblack e os relacionam com as coordenadas dos pontos Pijk, que determinam quando os pontos de equilíbrio P231 e P132 se tornam biologicamente relevantes. 0, pois o denominador é negativo neste intervalo, enquanto o numerador é sempre negativo devido a (3.10) e à Proposição 3.4.7.
Observe que o parâmetro qgreen define o valor de q a partir do qual o ponto de equilíbrio P132 se torna biologicamente relevante. Notemos que a Proposição 3.20 anterior mostra que o parâmetro qblue define o valor de q a partir do qual o ponto de equilíbrio P231 deixa de ser biologicamente relevante. A seguir mostramos um Lema que será utilizado na definição do parâmetro qred e posteriormente também.
A partir das propriedades da função µ provadas no Lema 3.4.15, os teoremas desta Proposição seguem facilmente.
Se G1 < 0, ∃qblack ∈ (qred,1], tal que ∀q ∈ (qblack,1] temos que o ponto de equilíbrio da coexistência das três estratégias torna-se biologicamente relevante. Os teoremas 3.4.11 e 3.5.1 mostram que o ponto de equilíbrio de coexistência Q ainda não é biologicamente relevante As observações acima, juntamente com o Teorema 3.4.11, mostram que o ponto de equilíbrio de coexistência Q não é biologicamente relevante.
As observações acima, juntamente com o Teorema 3.4.11, mostram que o ponto de equilíbrio de coexistência Q não é biologicamente relevante. As proposições 3.4.17 e 3.5.3 juntamente com o Teorema 3.4.11 mostram que o ponto de equilíbrio de coexistência Q ainda não é biologicamente significativo. A Proposição 3.4.17, juntamente com os Teoremas 3.5.2 e a ordem em L3 mostram que o ponto de equilíbrio de coexistência Q torna-se biologicamente significativo.
O Teorema 3.4.11 e a proposição 3.6.2 mostram que o ponto de equilíbrio de coexistência Q ainda não é biologicamente relevante. 3.4.14, o que prova que o ponto de equilíbrio P231 é biologicamente relevante, mas o ponto P132 ainda não é biologicamente relevante. O que foi dito acima, juntamente com o Teorema 3.4.11, Proposição 3.6.2 e Proposição 3.6.3, mostra que o ponto de equilíbrio de coexistência Q é biologicamente relevante.
O Teorema 3.4.11, a Proposição 3.5.1 e a Proposição 3.5.3 mostram que o ponto de equilíbrio de coexistência Q ainda não é biologicamente significativo. Na Seção 3.3, identificamos os pontos de equilíbrio biologicamente relevantes para o nosso exemplo. É importante notar que apenas para q ∈ (qblack,1] o ponto de equilíbrio de coexistência Q acaba sendo biologicamente relevante, representando a coexistência de três estratégias.

Por um lado, se considerarmos 1, 2 suficientemente pequenos, o Teorema 3.5.4 mostra que para q ∈(qblack,1] os pontos de equilíbrio P132 ∈ L2, P123 ∈L3 são biologicamente relevantes e o ponto de equilíbrio de coexistência Q torna-se biologicamente relevante. Os retratos de fase possíveis neste caso são 12, 12R e 13 e ainda não temos argumentos para excluir dois deles. Por um lado, se encontrarmos 1, 2 suficientemente pequeno, do Teorema 3.6.4 afirmamos que o os pontos de equilíbrio P123 ∈L3 e P231 ∈L1 são biologicamente relevantes e o ponto de equilíbrio de coexistênciaQ não o é.
Estamos então na mesma situação que em I.1 do exemplo 4.1, e portanto com esta informação relativa ao limite da região B, temos que o retrato de fase neste caso pertence à classe de equivalência 38R. Por um lado, se considerarmos que 1, 2 são suficientemente pequenos, segue-se do Teorema 3.6.4 que para q ∈ (qblack, qblue) os pontos de equilíbrio P123 ∈ L3 e P231 ∈ L1 são biologicamente significativos e o ponto de equilíbrio The conceito de coexistência torna-se biológicamente importante. Portanto, a dinâmica na fronteira B é dada pela Figura 4.8, que pode ser identificada com as classes de equivalência Bomze 9Re 10R, das quais podemos extrair o diagrama 10R pelo argumento de vizinhança P123 usado em I.1 do Exemplo 4.1.
Observemos também que a estratégia G com q = qgreen acaba sendo a estratégia chamada GT F T Generous Tit for Tat, ou seja, neste caso GT F T =S(1, qgreen). Por um lado, se considerarmos 1, 2 suficientemente pequenos, temos do Teorema 3.6.4 que para q ∈ (qazul, qverde) os pontos de equilíbrio P123 ∈ L3 e o ponto de coexistência Q são biologicamente relevantes, mas o ponto de equilíbrio P231 ∈ L1 é biologicamente relevante. Portanto, a dinâmica na fronteira de B é dada pela Figura (4.9), que podemos identificar inequivocamente com o diagrama Bomze 15R.
Estamos então na mesma posição que em I.1 do Exemplo 4.1, e assim com esta informação referente ao limite da região B, temos que os possíveis retratos de fase neste caso são 12, 12R e 13, e você ainda não tem argumento de que você pode descartar dois.

Falaremos de desenvolvimento parcial da cooperação quando os pontos do conjunto de trajetórias ω-limite em Γ tiveremx2 = 0. Por fim, diremos que houve desenvolvimento total da cooperação quando o conjunto de trajetórias ω-limite em Γ for o equilíbrio ponto E3. Seguindo os intervalos definidos na seção 4.1, podemos observar que para q ≤qgreen temos uma evolução de cooperação total.
Por fim, para valores “grandes” de q, ou seja, q > qzeze, não temos argumentos precisos para afirmar se houve uma evolução cooperativa, pois estamos entre três retratos de fase possíveis, onde um deles (12) propõe a ponto de equilíbrio interno Q como atrator, o outro (12R) como repulsor e o último (13) como centro. Conjecturamos que no caso G1 < 0 e q > qblack, o diagrama de Bomze associado será sempre 12 e que, portanto, haverá evolução cooperativa fraca neste caso, pois, exceto por uma órbita derivada de E3. Por fim, para valores “grandes” de q, ou seja, q > qgreen, não temos argumentos precisos para afirmar a evolução da cooperação, pois mais uma vez nos encontramos entre três possíveis retratos de fases, onde.
Suspeitamos que no caso G1 > 0 e q > qgreen o diagrama de Bomze correspondente será sempre 12R e que neste caso não haverá portanto evolução da cooperação, pois, com exceção de uma órbita que vai de Q a P123 , todas as outras órbitas convergem para o ponto E2. Neste caso, como nos casos anteriores, temos que para valores ‘pequenos’ de q, ou seja, q≤qverde, chegamos à evolução da cooperação total. Suspeitamos que no caso G1 = 0 e q > qgreen o diagrama de Bomze correspondente será sempre 13 e que neste caso haverá portanto uma evolução fraca da cooperação, como nas órbitas fechadas em torno do ponto Q fração de indivíduos de tipo G são positivos.
Evolução da cooperação entre antropóides virtuais: um modelo computacional baseado em agentes.