José Armando Carrillo Fasio por su orientación académica en el logro de los objetivos de investigación. Cambios en la fisiología de las plantas y la interacción planta-ambiente en función de la escala de tiempo de exposición al estrés hídrico.
INTRODUCCIÓN
Estrés Hídrico
- Efectos del Estrés Hídrico
- Mecanismos de Defensa al Estrés Hídrico
- Ajuste osmótico
- Regulación osmótica
- Osmolitos
- Pinitol
- Prolina
- Glicina betaína
- Inductores de Estrés Hídrico
Esta acumulación es un indicador de adaptación al estrés hídrico, por lo que se utiliza como criterio de selección de especies con excepcional tolerancia o resistencia (Arteaga et al., 2020). En tomate, se demostró la regulación de la actividad enzimática del metabolismo de la prolina en respuesta a condiciones variables de estrés hídrico (Fujita et al., 2003).

Bioestimulante
Plantas Halófitas
- Procesos Adaptativos en Plantas Halófitas
Esto se debe a desequilibrios iónicos, deterioro del intercambio de gases, pérdida de la homeostasis del agua y cambios en las vías metabólicas (Rahman et al., 2021). La mayoría de las especies de esta familia tienen características xeromorfas que les permiten evolucionar en ambientes extremos y cambiantes; incluso favorece su crecimiento reproductivo, que incluye la floración tardía y la vitalidad de las flores, el polen y las semillas; mencionado anteriormente como resultado de un proceso adaptativo que les permitió optimizar los mecanismos de alimentación durante el crecimiento vegetativo y los cambios hormonales (Yuang et al., 2019). Para ello, están dotadas de barreras apoplásticas en las células de la epidermis y endodermis de las raíces, que dificultan el flujo masivo de iones durante el proceso de transpiración.
Todavía se están haciendo esfuerzos para comprender los mecanismos que lo regulan. ultrafiltración del sistema radicular, ya que representan una oportunidad para dirigir programas de mejoramiento genético para reducir el impacto de la salinización del suelo en la agricultura. Esta capacidad está presente principalmente en las células parenquimatosas, que utilizan un mecanismo para reducir la actividad de los canales de vacuola tras la exposición a ambientes salinos. La mayor retención de K+ está más asociada a una actividad de inactivación de radicales libres mediante mecanismos de la vía antioxidante no enzimática, lo que permite evitar daños por acumulación de secuestro de sodio en las vacuolas de las células parenquimatosas.
Otras plantas halófilas que comparten este mecanismo son Salsola drummondii, Achras sapota y Sarcocornia fruticosa (Zeng et al., 2018; Rahman et al., 2021). d) Otros mecanismos individuales o asociados son la adaptación osmótica, cambios en las vías fotosintéticas, redistribución general de sales, simbiosis microbiana, entre otros mecanismos en descubrimiento y evolución (Sairam et al., 2006; Ma et al., 2020; Rahman et al. al., 2021).
Sesuvium portulacastrum
Portulaceae Prolina Fagales Casuarinaceae Prolina Lamiales Acanthaceae Glicina betaína Malpighiales Rhizophoraceae Prolina, sacarosa Myrtales Combretaceae Mannitol rehegua. Ko ka’avo ojapo yvoty rósa guive púrpura peve ha sa’ieterei morotĩ año pukukue, hi’a oñesirkunsida (cápsula) oguerekóva semilla morotĩ michĩ ha hũva, peteĩteĩ yva ojapo 50 semilla peteĩ cápsula-pe (Lonard ha Judd, 1997; Lokhande et al., 2009; ( Luettge ha ambuekuéra, 1989). Umi variación tesape, temperatura ha salinidad rehegua omoheñói germinación vai, upévare pe método de propagación ha e estolon rupive (Martínez et al., 1992).
Se ha destacado su importancia en desalinización, fitorremediación y protección ambiental a través del conocimiento de los mecanismos del comportamiento de las halófitas así como de la señalización metabólica para el manejo del estrés. Además de la producción de prolina, una mayor salinidad también provoca una mayor acumulación de glicina betaína (Lokhande et al., 2010). La glucosa-6-fosfato produce mionositol, que actúa como precursor del pinitol (Vernon et al., 1992).
La aplicación exógena de mioinositol de origen sintético vía hojas en Capsicum annuum sometidas a sequía, mostró su potencial como inductor de tolerancia a dicho estrés (Yildizli et al., 2017).

El Cultivo de Tomate
Las condiciones de prueba experimental fueron la comparación de la acumulación de indicadores de daño por estrés (físico, químico y bioquímico) en plantas control absoluto no expuestas a sequía (riego diario) versus plantas con 7 días sin riego (control) sin y con tratamiento. dosis µM de mioinositol). Sinaloa lideró la producción de las entidades federativas con el 60% de la producción nacional, de la cual las variaciones en el volumen de cosecha están relacionadas con la estacionalidad de la siembra. Lo anterior muestra que el sistema de producción de tomate en Sinaloa representa una importante fuente de empleo y por lo tanto influye en la economía en los diferentes niveles de la cadena.
Este es el caso de la reducción de la disponibilidad de agua que presiona sobre alternativas sustentables de producción de alimentos; A este estado de escasez de agua se suman problemas de gestión de recursos y baja eficiencia en los sistemas de riego convencionales (IICA, 2015). La crisis del agua es un problema latente en la agricultura, que requiere innovación en los sistemas hídricos y la gestión de su uso eficiente, y por lo tanto este recurso se ha convertido en una limitante importante en la producción sostenible de alimentos (Rijsberman, 2006). Esta condición de la especie fue sugerida por Yang y Lu (2005) como una ventaja ya que la aplicación exógena de glicina betaína podría usarse para evaluar su posible utilidad como inductor de tolerancia al estrés.
Existen informes controvertidos sobre la función inductora de tolerancia de la glicina betaína, pero en tomate tuvo un efecto para inducir tolerancia a condiciones de estrés hídrico (salinidad y sequía), como se informó (Makela et al., 1998ab; Heuer, 2003).
HIPÓTESIS
OBJETIVOS
Objetivo General
Objetivos Específicos
MATERIALES Y MÉTODOS
- Localización del Áreas de Colecta de Sesuvium portulacatrum L
- Preparación de Extracto de S. portulacastrum
- Eficiencia de Extracción
- Prueba de Fitotoxicidad del extracto de S. portulacastrum L
- Identificación de Componentes por Barrido Cromatográfico GC/MS
- Cuatificación del Osmolito Pinitol en Extracto
- Material Vegetal
- Inducción de Estrés Hídrico con Polietilenglicol
- Aplicación Foliar de Extracto de S. portulacastrum
- Diseño del Experimento
- Determinación de Contenido Relativo de Agua (CRA)
- Determinación del Contenido de Materia Seca
- Determinación de Clorofila
- Determinación de Prolina Libre y Ácido Abscísico
- Análisis de Datos
Al recibirlo, se eliminó la arena sobrante de la cosecha, se lavó con jabón y se enjuagó abundantemente con agua potable. La cuantificación de los indicadores de eficiencia de extracción se realizó sobre muestras filtradas con papel filtro Whatman No. La absorbancia se midió a 620 nm en un espectrofotómetro UV-visible (Cary 60, Agilent); para la cuantificación se realizó una curva de calibración de 0 a 80 µg simultáneamente con un estándar de glucosa (Sigma.
El ensayo se realizó por cromatografía de gases con el método de derivatización a acetatos de alditol de Albersheim et al. 1967), que consistió en la hidrólisis de la muestra con 500 µL de ácido trifluoroacético (2 N que contenía 50 µg de mio-inositol como estándar interno, ya que dicho ciclitol no fue detectado en el extracto) durante 1 h a 121°C, luego eliminar el TFA por evaporación con metanol; La derivatización se inició con una reacción de reducción con 150 µL de NaBH4 (20 mg mL-1 en NH4OH 1N) y una posterior acetilación con 200 µL de anhídrido acético y 20 µL de 1-metilimidazol como catalizador. La integración del área se realizó utilizando el software MS Workstation versión 6.5 (SP1) y el cálculo de la concentración se realizó a partir de una curva estándar de pinitol. Para ello y con base en la revisión de la literatura, se consideró como inductor del estrés hídrico un valor de potencial osmótico (Ψs) de -0,7 MPa, el cual se logró con 244 g de PEG 6000 por L-1 H2O según la ecuación de Michel. y Kaufmann (1973).
La cuantificación se realizó a partir de 200 mg de material fresco extraído en frío con 2 ml de etanol:agua (40:60 v/v).
RESULTADOS
- Cinética de Extracción (Sólidos Disueltos y Azúcares Totales)
- Prueba de Fitotoxicidad del Extracto de S. portulacastrum
- Identificación de Componentes del Extracto de S. portulacastrum
- Efecto del Extracto de S. portulacastrum en el Estrés Hídrico
- Apariencia Relativa de Hidratación
- Contenido Relativo de Agua (CRA)
- Contenido de Clorofila (a y b)
- Prolina
- Ácido Abscísico (ABA)
El extracto etanólico de Guasaba mostró una concentración de pinitol de 1.097 mg ml-1, correspondiente a 5,5 mg g-1 (peso fresco). De acuerdo con el valor de CRA, el contenido de materia seca aumentó, confirmando un estado de humedad reducida en los tejidos, afectado principalmente por la dosis de 0,45 µmol y el día 3 de inducción post-estrés. Los principales efectos de la aplicación de dosis foliares de pinitol en función del valor de materia seca reiteran que la dosis baja de 0,15 µmol fue la que devolvió el efecto de deshidratación del PEG a los valores más cercanos al control no inducido (Figura 13). ). .
El valor medio general para la dosis de 0,15 µmol fue del 13%, lo que se consideró la mejor protección contra el estrés hídrico basado en la retención de humedad del tejido. Los efectos de deshidratación del tratamiento con 0,30 y 0,45 µmol de pinitol pueden considerarse como un efecto combinado de la reacción de privación de agua y descartarse como efectos negativos porque se utilizaron las dosis del extracto S. Este osmolito fue el indicador fisiológico más importante en este estudio y a diferencia de los otros indicadores. , el grado de su acumulación respondió claramente a la inducción de estrés hídrico y a la inducción de dispersión por 0,15 µmol de pinitol, en asociación con las variables CRA y materia seca.
Por otro lado, el bajo contenido de prolina en plantas sin inducción de estrés coincide con los datos reportados por (Claussen, 2005), quienes reportaron valores de prolina de 1.5 µmol g-1 (peso fresco) en tomate Counter. El valor de CRA de la peor condición de estrés en cebada fue de 60%, lo que resultó en la acumulación de 24.4 µmol g-1 de peso fresco en comparación con el contenido basal de 0.1 µmol g-1 de peso fresco del control no estresado; mientras que la administración exógena de dl-ciclopentano-1,2,3-triol resultó en una reducción de la gravedad de la deshidratación con PCR leve. Se evaluó el contenido de ácido abscísico en hojas de plantas de tomate con estrés hídrico inducido y tratadas con diferentes dosis del extracto S. Las medias para días que no tienen una letra común son significativamente diferentes p reportaron una concentración de ácido abscísico en plantas no estresadas. tomates de unos 2500 picomoles g- 1 y un aumento correspondiente debido a la influencia del estrés salino y.

CONCLUSIONES
RECOMENDACIONES
Ghnaya T, Nouairi I, Slama I, Messedi D, Grignon C, Abdelly C, Ghorbel MH (2005) Effects of cadmium on growth and mineral nutrition of two halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum. Ghnaya T, Slama I, Messendi D, Grignon C, Ghorbel MH, Abdelly C (2007) Effect of Cd2+ on K+, Ca+ and N uptake in two halophytes Sesuvium portulacastrum and Mesembrynathemum crystallinum: implications for growth. Lokhande VH, Nikam TD, Penna S (2010b) Differential osmotic adaptation to iso-osmotic salt and PEG stress in vitro in the halophyte Sesuvium portulacastrum L.
Messedi D, Labidi N, Grignon C, Abdelly C (2004) Limits imposed by salt on the growth of sesuvium halophyte portulacastrum. Papageorgiou GC, Murata N (1995) Extremely strong stabilizing effects of glycine betaine on the structure and function of the oxygen-evolving photosystem II complex. Rehem BC, Bertolde FZ, De Almeida AF (2012) Regulation of gene expression in response to abiotic stress in plants.
Slama I, Messedi D, Ghnaya T, Savouŕe A, Abdelly C (2006) Effects of water deficit on growth and proline metabolism in Sesuvium portulacastrum.
Enrollamiento foliar y reducción de crecimiento de frutos, causado por el desarrollo
Estructura molecular de los principales osmolitos con capacidad de osmoprotección
Bases fisiológicas y moleculares de la tolerancia al estrés hídrico
Ilustración de la respuesta a estrés y bioestimulantes en plantas
Sesuvium portulacastrum o verdolaga de playa
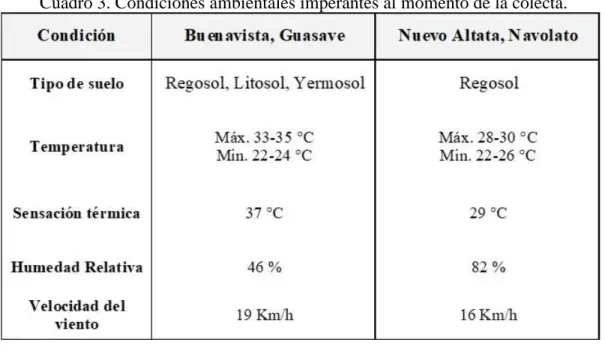
Localización geográfica de los puntos de colecta
Arreglo y distribución de macetas con plántulas de tomate tratadas con diferentes
Corrida cromatográfica (LC MS-MS) y curva de regresión para la cuantificación de
Cinética de extracción etanolática de sólidos a partir de biomasa de S
Hojas de tomate expuestas a diferentes concentraciones de extracto de S
Contenido relativo de agua (% CRA) en hojas de plantas de tomate Tisey DRD
Porcentaje de materia seca de hojas de plantas de tomate con estrés hídrico
Diferencias en apariencia de hojas de tomate post-exposición a estrés hídrico con
Contenido de clorofila a (A) y clorofila b (B) en hojas de plantas de tomate con
Contenido de prolina libre en hojas de plantas de tomate con estrés hídrico inducido
Contenido de ácido abscísico en hojas de plantas de tomate con estrés hídrico