El propósito de esta tesis es profundizar en los efectos no deseados derivados de la inserción y/o expresión de transgenes en plantas, utilizando como modelo los granos de arroz. De los resultados obtenidos en la tesis podemos concluir que el impacto del transgén sobre la planta huésped puede ser muy diferente, desde impedir su viabilidad o fertilidad hasta prácticamente cero.
INTRODUCCIÓN 1
Construcción de plantas de arroz transgénico para la síntesis de derivados de BP100 114 IV.2.1 Diseño y construcción de PAM derivados de BP100 para las mismas. Subclonación de genes de interés derbp100, 128 IV.2.5 Generación de plantas de arroz utilizando MG S-derbp100.
INTRODUCCIÓN
La planta de arroz como modelo de estudio
La aplicación de estas tecnologías ha servido para caracterizar los genes relacionados con las características agronómicas de la especie Oryza sativa. Sin duda, los estudios genómicos de las especies de Oryza sativa han evolucionado enormemente en la última década.

Transformación genética
El desencadenante de la infección por Agrobacterium son los compuestos fenólicos (como la acetosiringona) liberados por las células vegetales heridas, así como las moléculas de azúcar liberadas por las células vegetales en respuesta a una infección patógena, que provocan la expresión coordinada de genes de virulencia. (para) presente en el plásmido Agrobacterium Ti (Stachel et al., 1986). Hoy en día existen otras tecnologías, como PIG (pistola de entrada de partículas) o ACCELTM (aceleración por descarga eléctrica) (Shewry et al., 2008).
Aplicaciones de la transgénesis
Los ARNip (de 21 a 24 nucleótidos) se forman a partir de moléculas de ARN bicatenario (ARNds) mediante hidrólisis por la enzima DICER (Bernstein et al., 2001). Esta modificación produjo la variedad Golden Rice2, con mayores niveles de β-caroteno (Paine et al., 2005).

Análisis de plantas MG para la seguridad de alimentos y piensos; y del medio ambiente
Estos nucleótidos unidos a su vez llevan un protector sensible a la luz para que el ciclo pueda repetirse (Pease et al., 1994). Alternativamente, las técnicas de resonancia magnética nuclear (RMN) permiten la detección cuantitativa de metabolitos de forma no destructiva y pueden usarse para el análisis de un gran número de muestras (Cellini et al., 2004).

Péptidos antimicrobianos
Así, pueden presentar selectividad reducida, cierta actividad citotóxica y/o hemolítica y alta sensibilidad a las proteasas (Lui et al., 2010). La expresión de CecA en arroz confiere resistencia al hongo Magnaporthe grisea (Coca et al., 2006).

OBJETIVOS
Evaluación de diferencias transcripcionales (mediante hibridación de microarrays) y fenotípicas (rasgos agronómicos) entre líneas que expresan constitutivamente transgenes que codifican derivados de BP100 y la línea convencional más cercana (Senia). Comparación de los efectos no deseados de transgenes que codifican diferentes proteínas o péptidos antimicrobianos en plantas, especialmente derivados de AFP y BP100.
MATERIAL y MÉTODOS
A. Métodos de manipulación de ácidos nucleicos
A.1 Amplificaciones de fragmentos de ADN mediante la Reacción en Cadena de la Polimerasa
La enzima utilizada en este sistema es una ADN polimerasa Taq modificada que está inactivada y requiere altas temperaturas para su activación. a) La hibridación T óptima dependerá de la Tm de los cebadores. La formación de producto y la detección de fluorescencia se producen simultáneamente, mientras que siempre se tiene acceso a la cinética de la reacción de amplificación. El número de copias (C) de ARN o ADN (según el ensayo) de un gen determinado en una muestra de prueba se calculó a partir de la ecuación de primer grado [1] introduciendo el valor Ct obtenido de la reacción qPCR.
Para la comparación entre muestras, el valor de C fue relativo al número de copias de un gen endógeno en la muestra.

A.2 Purificación de productos de PCR
Centrifugar durante 1 minuto a 13.000 rpm para eliminar completamente cualquier rastro de etanol en la columna.
A.3 Electroforesis en gel de agarosa (AGE)
A.4 Extracción de fragmentos de ADN de gel de agarosa
A.5 Purificación fenólica y precipitación con etanol de soluciones de ADN
Seque el sedimento invirtiendo el tubo con la tapa abierta para eliminar completamente los restos de etanol.
A.6 Subclonajes
Un factor importante para la ligadura óptima entre el inserto y el vector es su relación molar en la mezcla de reacción (fórmula [4]). Se recomienda una relación molar de inserto:vector de 10:1 para fragmentos de tamaños similares; por el contrario, se utilizará una relación molar de inserto:vector de 2:1 para fragmentos de tamaños significativamente diferentes. Otro factor importante es la concentración de ADN total en la mezcla, que debe estar entre 10 y 20 ng/μL, y el volumen de la mezcla, que no debe exceder los 25 μL.
En cuanto al volumen de la mezcla, es importante que no supere los 25 µL, ya que esto puede reducir la eficiencia del proceso de transformación posterior.
A.7 Minipreparaciones de ADN plasmídico
No desactivar la enzima puede reducir la eficiencia (o incluso inhibir) el proceso de transformación. Añadir 300 l de tampón A3 (que contiene clorhidrato de guanidina) para neutralizar la solución y permitir que el ADN se adhiera bien dentro de la columna. Se recomienda lavar el ADN con 500 µL de tampón AW (que contiene clorhidrato de guanidina) mediante centrifugación durante 1 minuto a 13.000 rpm.
Para eliminar contaminantes como sales, metabolitos u otros componentes celulares solubles, lave la columna con 600 l de tampón A4 (que contiene etanol).
A.8 Secuenciación de fragmentos de ADN
Resuspender el sedimento en 30 l de reactivo de supresión de plantilla (Applied Biosystems, Foster City, CA, EE. UU., Foster City, CA, EE. UU.) y rehidratar durante 20 minutos a temperatura ambiente. Transfiera los 30 µl a microtubos ABI 310 (Applied Biosystems, Foster City, CA, EE. UU.) y ejecute las reacciones de secuenciación en el analizador genético ABI PRISM TM 310.

A.9 Transformación genética de bacterias
Ambos métodos tienen como objetivo desestabilizar la membrana bacteriana para permitir la transferencia de ADN al interior de la célula. Añadir una alícuota de la reacción de ligadura o 100 ng de plásmido a una alícuota de células competentes y mezclar suavemente. Incubar las células en hielo durante 30 minutos para permitir que el ADN se adhiera a la pared celular.

B. Construcción de plantas de arroz MG
B.1 Construcción de vectores para expresión en planta Oligonucleótidos
La enzima utilizada para la síntesis fue la polimerasa Expand High Fidelity de Roche (Roche Diagnostics, Barcelona, El vector pGEM®T-Easy se obtuvo de la empresa comercial Promega (Promega Corporation, Madison, EE. UU.). Este vector fue modificado con el gen de la proteína verde fluorescente, gfp, y sus elementos reguladores para la selección de callos de arroz transformados positivamente.
La selección de clones pCAMBIAgfp::derbp100 se realizó mediante PCR (Taq ADN polimerasa, Invitrogen, Life Technologies, Nueva York, EE. UU.) con los cebadores pUCGF_for y seqUBI_rev usando como ADN molde una suspensión de una ufc aislada seleccionada de placas LB-Hagar.

B.2 Transformación de plantas de arroz
Aspirar la suspensión bacteriana con una pipeta y dejar escurrir el callo sobre papel de filtro esterilizado. Transfiera los callos a un medio selectivo N6 que contenga cefotaxima (C) y timentina (T) (para eliminar Agrobacterium) e higromicina (H), para aquellas transformaciones con selección positiva para resistencia a herbicidas. Antes de transferir el callo al medio selectivo, retire los restos de Agrobacterium en papel de filtro estéril.
El callo resistente a la higromicina comienza a crecer y desarrollar nódulos brillantes, mientras que el callo inferior se vuelve necrótico.

B.3 Obtención de líneas homocigotas Aclimatación
Las condiciones de aclimatación de las plantas in vitro creadas durante este tema se variaron según el esquema representativo de la Figura III.10. Las semillas T1 producidas a partir de plantas T0 son, según la ley mendeliana, 50% hemicigotas (aA), 25% homocigotas (AA) y 25% no transgénicas (aa). Con el objetivo de identificar plantas homocigotas para el transgén (AA), se sembraron 24 semillas T2 de cada evento T1 en medio MS (para seleccionar las viables) y luego en N6.0 + higromicina.
El procedimiento para la obtención de líneas homocigotas de plantas con el gen de selección gfp se realizó de la misma forma que el proceso descrito, excepto la selección de semillas y plantas, que se analizaron bajo fluorescencia.

C Análisis de plantas de arroz transgénico
C.1 Cultivo de material vegetal MG
C.2 Análisis de ADN genómico
La técnica se basa en la separación de la mezcla de ADN mediante gel de agarosa (III.A.3), su transferencia a una membrana de nailon y finalmente la detección del fragmento deseado mediante hibridación específica con una sonda marcada. La síntesis de la sonda se diseñó para construirse mediante una reacción de PCR con α-32P-dATP. Este sistema se basa en marcar la sonda con la enzima fosfatasa alcalina, que degradará el sustrato dioxetano, provocando quimioluminiscencia.
Corte un trozo de membrana de nailon Hybond-N+ del tamaño del gel (6 x 9 cm para un gel pequeño).

C.3 Análisis transcriptómico de plantas de arroz MG Extracción de ARN mensajero
Dependiendo de la concentración de la solución de ARN utilizamos dos tratamientos diferentes, rutinario o estricto. La comparación de la fluorescencia emitida por las dos muestras hibridadas paralelas permitió determinar la sobre o subexpresión de los genes analizados. Los valores de fluorescencia obtenidos mediante hibridación de microarrays se procesaron para obtener el nivel de expresión de cada gen en la muestra analizada.
La determinación de la eficacia de amplificación de un gen y la obtención del valor Ct del mismo gen en la muestra problema se describe en el apartado III.A.1.
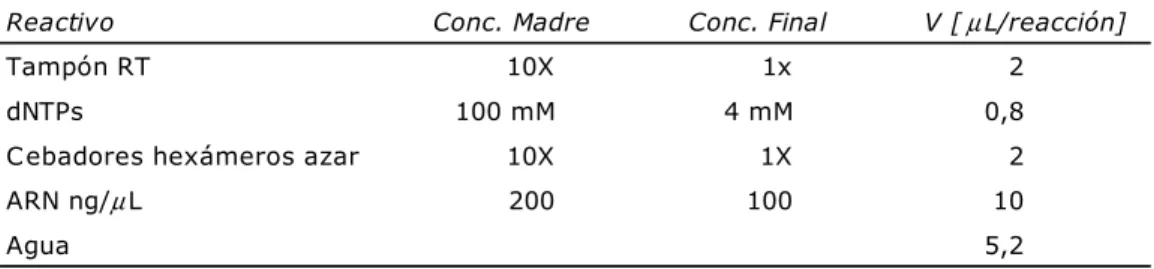
C.4 Análisis de actividad antibacteriana
Para cada extracción, y antes de comenzar a preparar el material, añadir 10 µL de la solución inhibidora de proteasa por mililitros de tampón de extracción. Diluir la solución madre a 0,1 mg/ml para la construcción de la línea estándar de acuerdo con las diluciones que se muestran en la Tabla III.24. Para preparar el complejo proteína-colorante se procedió como se indica en los puntos 2 al 4 para la formación de la línea estándar.
Para analizar el efecto de los péptidos sobre la viabilidad de las bacterias, se realizó un análisis de muerte celular.

C.5 Ensayos de resistencia a estrés Resistencia a estrés oxidativo
Resuspender la suspensión de esporas de la placa usando una pipeta estéril y pasar a través de un filtro Miracloth estéril (Calbiochem, Merck, Darmstadt, Alemania). Pasada una semana de la siembra se realiza un riego con un suplemento de nitrato (15 g/L) durante una semana más para favorecer la posterior infección de la planta con M. Cortar la segunda hoja y colocarla con el envés hacia arriba con ayuda. mediante fórceps o la mano sobre una placa de agar + medio kinetina (Sigma-Aldrich, St. Louis, MO, EE. UU.).
Después de 7 días de infección, analizar las hojas con el programa de análisis de imágenes APS Assess v2.0.

C.6 Ensayo agronómico. Evaluación de los componentes de producción
RESULTADOS
Análisis transcriptómico de plantas de arroz resistentes a plagas
- Comparación transcriptómica de Senia-afp respecto a Senia mediante hibridación por microarray
- Análisis de genes con expresión diferencial sobre-representados en Senia-afp La representación del total de genes con expresión diferencial sobre el genoma de la línea MG
- Evaluación del impacto del proceso de transformación sobre las diferencias transcriptómicas
- Evaluación de las diferencias transcriptómicas entre diferentes eventos MG de Senia-afp y Senia
- Regulación de las secuencias por el efecto del nivel de expresión del transgén El transgén de las líneas de arroz MG S-afp analizadas hasta el momento está regulado por el
- Regulación de secuencias por el efecto herida en plantas Senia y Senia-afp La presencia del transgén en una gran variedad de plantas MG, ha demostrado conferir a la
Los valores de fluorescencia obtenidos de la hibridación se analizaron conjuntamente mediante el programa Robust Multichip Average (RMA) para obtener los valores de expresión de los genes analizados. En el Apéndice 1 se describe la función de los genes correspondientes a las secuencias que mostraron expresión diferencial. Los resultados obtenidos en este estudio han revelado que la expresión diferencial del 50% de los genes se deriva de la presencia del transgén.
Se muestra que el resto de los 7 genes (40%) están regulados solo en S-afp, lo que indica que su regulación depende del nivel de expresión de la aplicación.

- Diseño y construcción de PAM derivados de BP100 para su expresión en planta Secuencias peptídicas quiméricas
- Estrategia de clonaje versátil
- Construcción de los vectores intermediarios
- Construcción de los vectores de transformación
- Generación de plantas de arroz MG S-derbp100 mediante transformación por Agrobacterium
- Análisis del efecto de los transgenes derbp100 sobre callos y plantas MG Los resultados de las transformaciones de callos con el gen de selección gfp evidencian el
- Análisis molecular de plantas MG S-derbp100
- Análisis fenotípicos
- Ensayo de componentes de rendimiento de eventos MG S-derbp100
Las secuencias del gen derbp100 (con la secuencia de aminoácidos a continuación) se presentan en las Figuras IV.6 y IV.9. La Figura IV.13 representa esquemáticamente la posición del casete de expresión derbp100 en el vector de transformación. Finalmente, los niveles de ARNm de derbp100 se cuantificaron mediante PCR en tiempo real (qPCR).
Los niveles de expresión del gen de selección y los transgenes derbp100 se normalizaron con respecto a los niveles de ARNm de los genes endógenos. Construcción de los vectores de expresión derbp100 con el gen de selección hpt II. Las secuencias de cebadores se muestran encuadradas en azul (SouthUBI_for) y verde (SouthBP_rev).

Análisis transcriptómico de plantas de arroz MG diseñadas para la síntesis de derBP100
- Comparación transcriptómica mediante análisis de hibridación en microarray Análisis y validación
Mientras que los genes regulados en S-bp217-1 en relación con Senia estaban en su mayoría reprimidos en S-bp217 (42 genes, 7 de ellos más de 5 veces; en comparación con 22 genes inducidos, 6 de ellos más de 5 veces), el La mayor proporción de los regulados en S-bp213-5 se expresa más intensamente en la planta transgénica (28 genes inducidos, 6 de ellos más de 5 veces; frente a 9 genes reprimidos, 1 de ellos más de 5 veces). Por tanto, aproximadamente la mitad de los genes regulados en S-bp213-5 en relación con Senia también muestran expresión diferencial entre S-bp217-1 y Senia. La descripción de cada uno de los genes regulados se recoge en la tabla IV.18.
Patrones de expresión de genes reprimidos en S-bp217-1 en comparación con Senia, en otras líneas S-derbp100.
