Organización de la región genómica que contiene el gen que codifica la serina/treonina quinasa PrkA……. Superposición del modelo PrkA de L.monocytogenes (azul) y la estructura de PknB (PDB: 4EQM) S. Interacciones de ATP (A) y quercetina (B) con residuos de aminoácidos presentes en el sitio de unión de ATP de PrkA… …….
INTRODUCCIÓN
17 enfermedades crónico-degenerativas y los estudios en este campo se han centrado en la búsqueda de compuestos inhibidores (Taunton et al., 2014). Los inhibidores de cinasas se clasifican en cuatro tipos, según su mecanismo de inhibición (Wu et al., 2015). Los tipos I y II son inhibidores que compiten por el sitio de unión de ATP y mientras que los tipos III y IV son inhibidores alostéricos no competitivos (Wu et al., 2015).
ANTECEDENTES
- Riesgos que Representan las Biopelículas Listeria monocytogenes para la Salud
- Formación de Biopelículas de L. monocytogenes y el Papel de las Sustancias
- Algunos Factores de Virulencia de L. monocytogenes como las Internalinas y la
- La Señalización de la PrkA es Clave en la Regulación de la Formación de las
- Potencial de Quercetina para Inhibir PrkA y la Formación de Biopelículas y
19 la instalación involucrada y las agencias federales que respondieron al brote (Kate et al., 2015). Este factor de virulencia está asociado principalmente al estilo de vida intracelular (García-del Portillo et al., 2011). El gen de serina/treonina, PrkA, está organizado en un operón (Figura 5) que incluye otros genes como lmo1821 y otros genes implicados en vías de información (modificación y metabolismo de ADN, ARN y proteínas; lmo1819, lmo1822, fmt y priA) y metabolismo intermedio (lmo1818 y lmo1825) (Lima et al., 2011).

HIPÓTESIS
OBJETIVOS
Objetivo General
Objetivos Específicos
MATERIALES Y MÉTODOS
- Etapa 1: Caracterización de la Actividad Antibacteriana y Anti-biopelícula de
- Cepas Bacterianas y Condiciones de Cultivo
- Determinación de la Concentración Mínima Inhibitoria (CMI) y Bactericida
- Efecto de la Quercetina sobre Biopelículas Maduras de L. monocytogenes
- Determinación de la Concentración Mínima Inhibitoria de Biopelículas de
- Efecto de la Quercetina Sobre la Permeabilidad de la Membrana y las
- Diseño Experimental y Análisis Estadístico
- Etapa 2: Efecto de la Quercetina Sobre la Organización Espacio-temporal deBiopelículas
- Cinética de Formación de Biopelículas de L. monocytogenes en Presencia de
- Extracción y Cuantificación de las Sustancias Poliméricas Extracelulares
- Microscopía Láser Confocal de Barrido
- Cinética de Crecimiento de L. monocytogenes en Estado Planctónico Presencia de
- Diseño Experimental y Análisis Estadístico
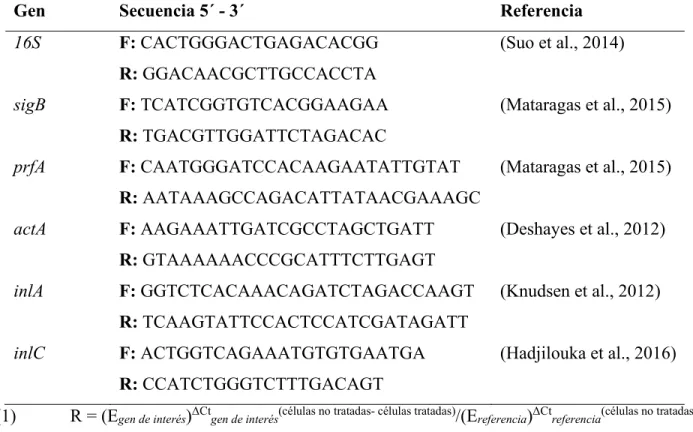
- Etapa 3: Expresión de prkA, sigB, prfA, inlA, inlC y actA en L. monocytogenes
- Diseño Experimental y Análisis Estadístico
- Etapa 4: Efecto Inhibitorio de la Quercetina contra la PrkA
- Construcción del Modelo de la Estructura de la PrkA
- Sobreexpresión y Purificación del Dominio Cinasa de PrkA
- Ensayo de Actividad Enzimática
- Análisis de Acoplamiento Molecular
- Diseño Experimental y Análisis Estadístico
Para determinar el efecto de la quercetina sobre la adhesión inicial y la formación de biopelículas, se retiraron tres discos de acero inoxidable del biorreactor después de 2 y 24 h de incubación, respectivamente. Para los ensayos de inhibición de la formación de biopelículas, los factores fueron la concentración de quercetina y el tiempo de incubación (2 y 24 h). Las proteínas totales se cuantificaron mediante reacción con ácido bicinconínico (BCA; Thermo Scientific™).
Para garantizar que el efecto de la quercetina sobre la adhesión y la producción de SPE durante la formación de biopelículas no se debiera a la pérdida de viabilidad celular, se realizaron cinéticas de crecimiento planctónico en presencia de quercetina a 0,2 mM. A partir de la pendiente de esta curva, el valor E se calculó usando la fórmula E = 2-1/pendiente. La sobreexpresión de PrkA se indujo mediante la adición de IPTG 1 mM (isopropil-β-D-1-tiogalactopiranósido).
Una vez confirmada la actividad de la proteína sobreexpresada, se evaluó el efecto inhibidor de la quercetina sobre ella. Por otro lado, se realizó un análisis dosis-respuesta para calcular la concentración de quercetina que inhibe el 50% de la actividad enzimática (CI50). El modelo PrkA obtenido como se describe en la Sección 5.4.1 se usó para realizar un análisis de acoplamiento molecular entre la quercetina y el sitio de unión de ATP de PrkA usando el complemento AutoDock Vina del programa Chimera (Trott y Olson, 2010).
La estructura tridimensional de la quercetina y el ATP se importó de la base de datos PubChem (ID: 5280343 y 5957, respectivamente).
RESULTADOS Y DISCUSIONES
Etapa 1: Caracterización de la Actividad Antibacteriana y Anti-biopelícula de
- Efecto de la Quercetina sobre L. monocytogenes en Estado Planctónico y
- Efecto de la Quercetina en la Permeabilidad de la Membrana de L. monocytogenes y
- Conclusiones
Estos resultados confirman el papel de la quercetina como agente que previene la formación de biopelículas de L. Además, existe evidencia del papel protector de las biopelículas maduras en el efecto antibacteriano de diversos compuestos (Pan et al., 2006), sin embargo, En el estudio actual no fue posible observar esto debido a la baja solubilidad de la quercetina a > 26,4 mM en el medio de cultivo. La Figura 9 muestra el efecto de la concentración de quercetina y el tiempo de exposición sobre la absorción del cristal violeta, siendo este efecto dosis dependiente (p ≤ 0,05).
Estos resultados muestran que la quercetina fue capaz de reducir la permeabilidad de la membrana de L. Si bien el presente estudio observó el efecto opuesto sobre la permeabilidad, estas diferencias pueden deberse a la composición de los liposomas utilizados y la composición lipídica de la membrana de L. L. La energía libre de la superficie refleja el carácter hidrófobo/hidrófilo de las bacterias (Shen et al. , 2017), por lo que los datos obtenidos indican que la presencia de quercetina aumenta la hidrofobicidad de L.
El valor negativo del potencial zeta refleja la carga negativa neta de la superficie bacteriana, que se atribuye a los grupos fosfato de los ácidos teicoicos de la pared celular y a los fosfolípidos de la membrana (Bisbiroulas et al., 2011). El valor negativo de ΔGadh observado entre las bacterias tratadas con quercetina y acero inoxidable no está de acuerdo con la inhibición de la adhesión observada experimentalmente. Esto podría deberse a la posible internalización de la quercetina, donde podría actuar en otros sitios objetivo e inhibir la producción de adhesinas que mejoran la unión a la superficie.
Por tanto, es necesario profundizar en este posible mecanismo evaluando el efecto de la quercetina sobre la producción de sustancias poliméricas extracelulares por parte de L.

Etapa 2: Efecto de la Quercetina Sobre la Densidad Celular, Morfología y
- Cinéticas de Formación de Biopelículas de L. monocytogenes en Presencia de
- Conclusiones
Estos resultados indican que la quercetina interfiere con la formación de biopelículas de L: monocytogenes, probablemente debido a la inhibición de la síntesis de SPE que ayudan a retener las células hijas una vez que se completa la división celular. Otro aspecto importante de la formación de biopelículas es la colonización de la superficie durante su desarrollo. El análisis cuantitativo de las imágenes (Figura 14) nos muestra que este compuesto afectó la dinámica de formación de biopelículas de L.
Los resultados confirman que este flavonoide actúa en las primeras etapas de formación de biopelículas y cambia su organización espacio-temporal. Mientras que los flavonoides como la miricetina, la hesperetina y la chalcona floretina inhibieron la formación de biopelículas de S. El proceso de formación de biopelículas es complejo y requiere la coordinación de la replicación de la superficie y la producción de SPE para colonizar la superficie y crear biopelículas maduras.
71 Según los resultados, la quercetina afecta la producción de proteínas extracelulares, reduce la adhesión y la formación de microcolonias, lo que afecta la capacidad de colonizar superficies abióticas y formar biopelículas maduras. A ambos polisacáridos se les atribuye un papel protector más que estructural en la formación de biopelículas (Brauge et al., 2016; Köseoğlu et al., 2015). En nuestro estudio, se observó que la quercetina inhibía la producción de proteínas extracelulares y afectaba la adhesión inicial y la formación de biopelículas de L.
Por tanto, el efecto inhibidor sobre la formación de biopelículas se debe a la inhibición de la acumulación de proteínas en la matriz extracelular, lo que perjudica la adhesión, la formación de microcolonias y, por tanto, el establecimiento de biopelículas maduras.

Etapa 3: Expresión de Genes inducidos por la Actividad de PrkA Relacionados
- Conclusiones
Esto sugiere que la quercetina es capaz de modular la expresión de factores de virulencia de L. Estos resultados pueden estar relacionados con la reducción en la producción de proteínas extracelulares en las biopelículas de L. En el presente trabajo se observó que con la concentración de 0,8 mM la expresión de las internalinas inlA e inlC fue 14 y 23 veces menor en comparación a células no tratadas. Por otro lado, InlC se cuantificó en mayor abundancia en biopelículas en comparación con las células planctónicas (Lourenço et al., 2013), lo que sugiere que diferentes internalinas tienen diferentes funciones durante el desarrollo de biopelículas de L.
Por tanto, la supresión observada en los tratamientos con quercetina puede estar relacionada con la inhibición de L. Asimismo, a concentraciones de 0,2 y 0,8 mM, la expresión del gen actA fue 20 y 29 veces menor, respectivamente, en comparación con las células no tratadas. fue tratado. Por tanto, es probable que el efecto sobre el establecimiento de microcolonias y maduración de biopelículas formadas en presencia de quercetina esté relacionado con la supresión del gen actA. 76 Otros autores han abordado el estudio de los flavonoides sobre la virulencia de L. monocytogenes, pero se ven limitados al no profundizar en el posible modo de acción.
2016) reportaron que la morina inhibió la actividad hemolítica de este patógeno entre un 54 y 70%; sin embargo, no se evaluó su efecto sobre la expresión génica. Esto podría indicar que, en el caso específico de la actividad hemolítica, la floretina podría actuar a nivel postraduccional contra LLO (Wang et al., 2017). En el presente trabajo observamos que la quercetina suprime la expresión de los genes inlA, inlC y actA, por lo que es probable que la inhibición de la actividad hemolítica también se deba a la supresión del gen hyl, ya que todos ellos se encuentran en el mismo gen. . isla de patogenicidad (LPI1) bajo el control de prfA, también suprimida por el efecto de la quercetina.
Los resultados muestran que la quercetina suprime la expresión de los genes de estrés (sigB) y virulencia (prfA, actA, inlA e inlC) asociados con la formación de biopelículas de L.

Etapa 4: Efecto Inhibitorio de la Quercetina Sobre PrkA de L. monocytogenes
- Modelo Tridimensional del Dominio Catalítico de la PrkA de L. monocytogenes
- Sobreexpresión y Purificación de la PrkA Mediante Cromatografía de Afinidad
- Efecto Inhibitorio de la Quercetina Sobre la Actividad Cinasa de la PrkA y Análisis
- Conclusiones
78 de las regiones antes mencionadas, se puede suponer que el pliegue del modelo construido de PrkA es estable. La Figura 19 muestra la superposición del modelo PrkA con la estructura PknB de S. Se observó que PrkA conserva los dominios y residuos de aminoácidos que participan en la orientación de la molécula de ATP y en la transferencia del grupo γ-fosfato.
En el modelo diseñado de PrkA estaban presentes residuos implicados en la estabilización de grupos fosfato y la catálisis. Lima et al. informaron por primera vez sobre la sobreexpresión y purificación del dominio catalítico de PrkA. Una vez obtenida la proteína pura y activa, se evaluó el efecto inhibidor de la quercetina sobre la actividad serina/treonina quinasa de PrkA.
En la Figura 25.A podemos ver que ambos ligandos están ubicados en la cavidad formada entre los lóbulos N y C de PrkA. La Figura 25.B muestra la disposición de los ligandos dentro de la cavidad y observamos que los anillos A y C de la quercetina se superponen con el heterociclo de adenina de la molécula de ATP. Por otro lado, en nuestro estudio observamos que la quercetina inhibía la actividad enzimática de PrkA de L.
Por lo tanto, la actividad anti-biopelícula de la quercetina puede deberse a la inhibición de la señalización mediada por ParkA.

CONCLUSIONES GENERALES
RECOMENDACIONES
Positive correlation between inlA and inlB gene expression of Listeria monocytogenes and its adhesion strength to the glass surface. Effect of nutrient limitation on Listeria monocytogenes biofilm structure grown in a microfluidic system. Differential internal A levels in Listeria monocytogenes biofilms grown on different surfaces and nutrient conditions.
Differential gene expression profiling of Listeria monocytogenes in Cacciatore and Felino salami to reveal potential stress resistance biomarkers. The in vitro antibiofilm activity of selected culinary herbs and medicinal plants against Listeria monocytogenes. A screen for kinase inhibitors identifies antimicrobial imidazopyridineaminofurazans as specific inhibitors of Listeria monocytogenes PASTA kinase PrkA.
Case series of non-invasive Listeria monocytogenes febrile gastroenteritis involving ready-to-eat meat. Morin inhibits biofilm production and reduces virulence of Listeria monocytogenes - An in vitro and in vivo approach. Comparison of protein patterns of biofilm- or planktonically grown Listeria monocytogenes by proteomic analysis.
The significance of SigN for Listeria monocytogenes static and continuous flow biofilm formation and disinfection resistance.