Ramón Molina, gracias por tu apoyo, disponibilidad y por enriquecer este trabajo con tu valiosa experiencia. Las principales APC del sistema respiratorio son las células dendríticas pulmonares (DC), las DC de los ganglios linfáticos y los macrófagos alveolares pulmonares (PAM).
INTRODUCCIÓN
Sin embargo, la mayoría de los estudios que evalúan la susceptibilidad de las CD al virus del síndrome respiratorio y reproductivo porcino (PRRSV) se han realizado in vitro con CD derivadas de monocitos (Mo-DC) o derivadas de médula ósea (BM). -DC) (Flores-Mendoza et. al., 2008;.3 La identificación y caracterización de poblaciones de APC respiratorias porcinas contribuirá a una mejor comprensión de su implicación en la inmunopatología de enfermedades respiratorias porcinas como el PRRS.
ANTECEDENTES
Células Dendríticas (DCs)
- Receptores y Poblaciones de DCs
- Generalidades de las Células Presentadoras de Antígeno en el Cerdo
- El Papel de las Células Presentadoras de Antígeno en Infecciones Virales 12
- Síndrome Reproductivo Y Respiratorio Porcino
- Etiología del PRRS
- Mecanismo de Infección del PRRSV y Respuesta Inmune
Origen, localización, desarrollo y maduración de la población celular de antígenos murinos (Satpathy et al., 2012). CD163 es un receptor necesario para la infección de una célula diana por PRRSV (Calvert et al., 2007).
HIPÓTESIS
OBJETIVOS
General
Particulares
MATERIALES Y MÉTODOS
Caracterización Fenotípica y Funcional de las APCs de Pulmón de Cerdo
- Animales
- Obtención del Pulmón y de las Células Presentadoras de Antígenos
- Anticuerpos y Citometría de Flujo
- Aislamiento de Células MHC-II + por MACS
- Aislamiento de Linfocitos de Sangre Periférica (PBLs)
- Reacción Leucocitaria Mixta (MLR)
- Ensayo de Internalización de Antígenos
Las células mononucleares se recogieron con una pipeta Pasteur y se lavaron dos veces durante 10 minutos a 1200 rpm a 15 °C. Transcurrido el tiempo de cultivo, las células no adherentes (NAC) se retiraron cuidadosamente del matraz y el matraz se lavó 2 veces con 10 ml de PBS estéril. Luego las células se incubaron con anti-MHC-II (molécula de presentación para antígenos de origen proteico, clon H42A; VMRD) conjugado con R-PE Cy7 (LNK112PECY7; AbDSerotec, Oxford, .. VMRD) conjugado con R-PE (LNK022RPE) AbDSerotec, Oxford, Reino Unido).
Además, las células se incubaron con anti-CD163 (receptor del complejo haptoglobina-hemoglobina, clon 2A100/11; AbDSerotec, Oxford, Reino Unido) conjugado con el kit de etiquetado Zenon® Alexa Fluor® 647 Mouse IgG1 (Z250008; ). Primero, las células se marcaron con anticuerpo anti-MHC-II de ratón y se incubaron a 4 °C durante 30 minutos, luego se lavaron dos veces con 2 ml de PBS/FBS al 2 %. Las células resultantes se contaron y se fijaron a una concentración de 1 x 106 células/ml en RPMI-1640 completo.
Se utilizaron células MHC-II+ enriquecidas con MACS en ensayos de funcionalidad e infección por PRRSV. Las células mononucleares de sangre periférica se lavaron tres veces en RPMI y se resuspendieron en medio RPMI-1640 completo. Luego, se lavaron tres veces con PBS frío al 0,1% de FBS y se marcaron con anti-MHC-II para analizar la capacidad de internalización del antígeno por parte de las células MHC-IIhigh.
Evaluación de la Susceptibilidad de las APCs de Pulmón de Cerdo al PRRSV
- Inoculación con el PRRSV
- Evaluación de la Replicación Viral
- Evaluación de Apoptosis y Necrosis en las Células Infectadas
Para las pruebas de endocitosis y fagocitosis, la internalización de dextrano y látex, respectivamente, se determinó en 1 x 106 células/ml. Se añadieron 0,5 µm de dextrano (marcado con isotiocianato de fluoresceína) (FD70, Sigma-Aldrich, St Louis, MO, EE. UU.) hasta una concentración final de 400 µg/ml. Se añadió látex de 0,5 μm (marcado con poliestireno modificado con carboxilato de color naranja fluorescente) (L5530, Sigma-Aldrich, St Louis, MO, EE. UU.) a una concentración de 3,7 x 108 perlas/ml.
Las células se incubaron durante 1 h a 37 °C y, para evaluar la internalización específica, se utilizaron tubos con células tratadas en las mismas condiciones pero incubadas durante 1 h a 4 °C. Se analizaron mediante citometría de flujo y para evaluar la unión no específica de partículas de dextrano o látex, los porcentajes positivos en células incubadas a 4 °C se restaron de los porcentajes obtenidos en células incubadas a 37 °C (Loving et al., 2007) . C en varios momentos y 72 h), donde se recogieron células y sobrenadantes de cada momento para evaluar la replicación viral, el fenotipo, la funcionalidad, la apoptosis y la necrosis.
Luego se realizaron diluciones seriadas diez veces de los sobrenadantes obtenidos en diferentes tiempos de las células inoculadas con PRRSV y se incluyeron como control negativo las células tratadas con simulacro y como positivo el virus NVSL-9587 con una concentración viral previamente determinada (1 x 107). control. Log10 CCID50/mL). Las células se cultivaron durante 72 horas a 37°C y la placa se revisó cada 24 horas para detectar el efecto citopático. La capacidad del PRRSV para inducir apoptosis se evaluó en L-C, LN-C y PAM utilizando el kit de detección de apoptosis anexina V-FITC (POAF, Sigma-Aldrich, St Louis, MO, EE. UU.), siguiendo las recomendaciones del proveedor. (Pujhari). et al., 2014).
Análisis Estadístico
RESULTADOS
Caracterización Morfológica, Fenotípica y Funcional de las APCs de Pulmón de
- Morfología de L-Cs, LN-Cs y PAMs
- Caracterización Fenotípica de las Células Presentadoras de Antígeno de Pulmón
- Reacción Leucocitaria Mixta por las L-Cs, LN-Cs y PAMs
- Capacidad de Internalización de Antígenos por las APCs Pulmonares
- Producción de Lisozima por las APCs
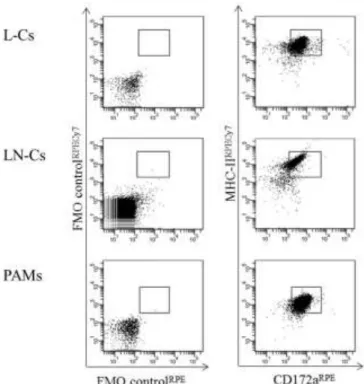
El porcentaje de células contaminantes se determinó en las células MHC-II+ enriquecidas en MACS (L-Cs, LN-Cs y PAMs). Por otro lado, el porcentaje de células MHC-II+CD3+ fue bajo en las tres poblaciones (≤ 1,8%). La evaluación de las células MHC-II+CD79a+ se realizó en células permeabilizadas y se detectó una cantidad baja de estas células en las tres poblaciones (≤ 1,7%).
Las células presentadoras de antígeno pulmonar (L-C), las células presentadoras de antígeno de los ganglios linfáticos mediastínicos (LN-C) y los macrófagos alveolares pulmonares (PAM) MHC-II+ enriquecidos con MACS se analizaron en función de su tamaño (FSC) y complejidad. (SSC)). Se analizaron las células presentadoras de antígeno pulmonar (L-C), las células presentadoras de antígeno de los ganglios linfáticos mediastínicos (LN-C) y los macrófagos alveolares pulmonares (PAM) en función de la expresión de MHC-IIhigh y CD172a+. Se realizó un análisis de la expresión del receptor en células presentadoras de antígeno pulmonar (L-C, barras negras), células presentadoras de antígeno de ganglio linfático mediastínico (LN-C, barras grises) y macrófagos alveolares pulmonares (PAM, barras blancas). .
Se incubaron células MHC-II+ enriquecidas con MACS con dextrano o látex durante una hora a 4° o 37°C. Para evaluar la capacidad de las células para producir lisozima después de la fagocitosis, se midió la expresión intracelular de lisozima en células MHC-II+ enriquecidas con MACS. Se cultivaron células presentadoras de antígeno pulmonar (L-C), células presentadoras de antígeno de ganglio linfático mediastínico (LN-C) y macrófagos alveolares pulmonares (PAM) MHC-II+ enriquecidos con MACS con látex o P.
Evaluación de la Susceptibilidad de las APCs de Pulmón de Cerdo al PRRSV 38
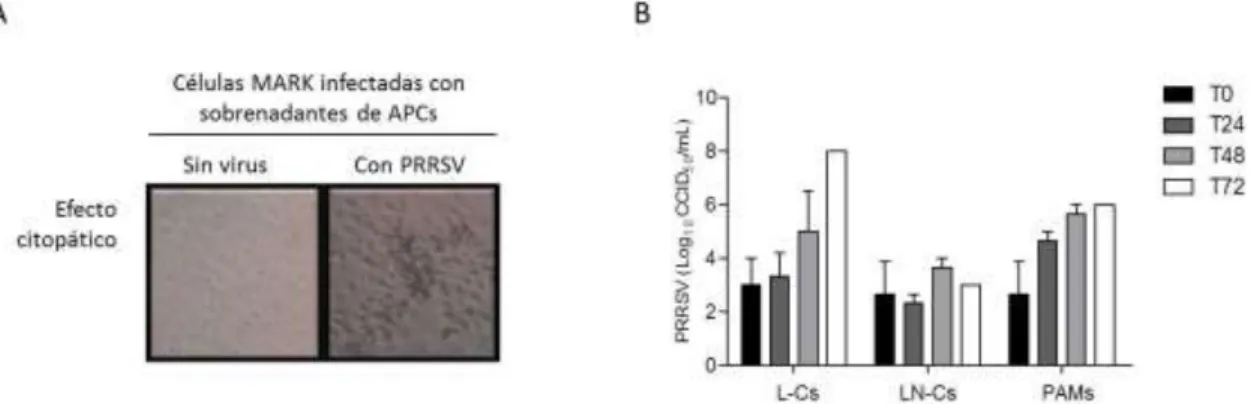
En tres poblaciones celulares, se observó un efecto citopático a partir de las 24 horas, pero solo en L-C y en PAM se observó un aumento en la cantidad de partículas virales con el tiempo (Figura 15B). Aunque no se observaron diferencias estadísticas con respecto al tiempo, estos resultados indican que PRRSV se replicó claramente en L-C pero no en LN-C; Se necesitan más pruebas para confirmar esto. Para comprender el efecto causado por la replicación de PRRSV en las APC, se evaluaron las características fenotípicas de tres poblaciones celulares después de la inoculación de PRRSV (Figura 16).
En el caso de la expresión de MHC-II y CD172a, se vio afectada en L-C y LN-C, donde se observó una ligera disminución en las células con PRRSV en comparación con las células simuladas, en PAM no hubo efecto aparente. . Aunque hubo una ligera disminución en los marcadores de presentación y activación en L-DC y LN-DC, esta diferencia no fue muy evidente entre las células simuladas y las de 0,1 MOI. Se observó que el virus disminuía la capacidad de inducir linfoproliferación en L-C y LN-C y aparentemente no tuvo efecto sobre PAM (Figura 17).
Finalmente, se evaluó la capacidad del PRRSV para inducir apoptosis o necrosis en células infectadas. En las L-C con MOI, se observó un aumento en la apoptosis y necrosis temprana y tardía a las 48 h después de la infección, y a las 72 h también se observó un aumento en la necrosis. Estos cambios se compararon con lo observado en células con simulación (Figura 18B). De manera similar, esto sucedió en los LN-C con MOI, pero solo con las células en necrosis en los tiempos 24, 48 y 72 h; Sin embargo, en las células con simulacro se detectó un elevado número de células en necrosis, posiblemente esta población fue más afectada por el cultivo ex vivo que las L-C y las PAM.
DISCUSIÓN
La expresión de CD163 en las L-C sugiere que estas células podrían ser macrófagos; sin embargo, existe evidencia de que existen CD porcinas que expresan CD163, como las generadas in vitro con la citocina hematopoyética tirosina quinasa similar a Fms (Flt3L) (Guzylack-Piriou et al., 2010). Mientras que en el estudio de Marquet, las células se obtuvieron de linfa aferente, donde solo se encontraron CD migratorias (Marquet et al., 2011). CD206 (manosa) es un receptor similar a lectina dependiente de calcio (tipo C; CTLR), su funcionalidad está asociada con endocitosis y presentación cruzada (Burgdorf et al., 2006; Wollenberg et al., 2002).
Esta información es consistente con lo informado en CD11c+ L-C por Loving et al. 2007), los autores observaron que las células inducían una mayor proliferación de linfocitos T que las CD derivadas de monocitos o los macrófagos derivados de monocitos. Esto era de esperar ya que los macrófagos son las células más especializadas en la fagocitosis (Gordon et al., 1974). En el presente estudio, se evaluó preliminarmente la infectividad del PRRSV en L-C y se utilizaron PAM como control positivo, ya que se sabe que son la principal célula diana del PRRSV (Duan et al., 1997a,b).
Además, la replicación podría probarse en las L-C; Sin embargo, estos resultados contradicen lo descrito previamente por Loving et al. 2007), ya que no detectaron replicación viral en estas células. También se observó que la capacidad de inducción de linfoproliferación de las L-C se ve afectada y esto puede estar relacionado con la expresión reducida de las moléculas de presentación del antígeno MHC-II, descritas previamente en las Mo-DC (Park et al., 2008). 49 en comparación con aquellos que solo se burlaron, esto también coincide con lo reportado anteriormente (Wang et al., 2007).
CONCLUSIÓN
Duan, X., Nauwynck, H.J., Pensaert, M.B., 1997b, Virus quantification and identification of cellular targets in lung and lymphoid tissues of pigs at different time intervals after inoculation with porcine reproductive and respiratory syndrome virus (PRRSV). Guilliams, M., Lambrecht, B.N., Hammad, H., 2013, Division of labor between lung dendritic cells and macrophages in defense against pulmonary infections. Haverson, K., Singha, S., Stokes, C.R., Bailey, M., 2000, Professional and nonprofessional antigen-presenting cells in the porcine small intestine.
Jingwen, Z., Danting, H., Xin, G., Xinna, G., Hanchun, Y., 2011, The increase of CD4+CD25+ T cells in the peripheral blood of pigs persistently infected with porcine reproductive and respiratory syndrome virus is infected. Labarque, G., Van Gucht, S., Nauwynck, H., Van Reeth, K., Pensaert, M., 2003, Apoptosis in the lungs of pigs infected with porcine reproductive and respiratory syndrome virus and associations with the production of apoptogenic cytokines. Neumann, E.J., Kliebenstein, J.B., Johnson, C.D., Mabry, J.W., Bush, E.J., Seitzinger, A.H., Green, A.L., Zimmerman, J.J., 2005, Assessing the economic impact of porcine reproductive and respiratory syndrome on swine production United States.
Suradhat, S., Thanawongnuwech, R., 2003, Upregulation of interleukin-10 gene expression in the leukocytes of pigs infected with porcine reproductive and respiratory syndrome virus. In the first three experiments, the dead cells were excluded with Fixable Viability Dye eFluor®. Lysozyme production was evaluated in L-Cs, LN-Cs and BAL-Cs stimulated with 222.
Next, to characterize the phenotype of the APC cell populations in the FSC/SSC gate 256. Next, we assessed the dextran or latex uptake capabilities of MHC-II+ enriched L-Cs, LN-284.
