Saps més que ningú com d'"ingrats" poden ser els complexos de cadena. Les metodologies aplicades per a l'estudi dels complexos que formen CRM i l'estudi de la via endògena de la biosíntesi de CoQ10 han millorat significativament la qualitat analítica i el diagnòstic en pacients amb sospita de malaltia mitocondrial i/o deficiència de CoQ10.
INTRODUCCIÓ
El mitocondri i les malalties mitocondrials
- El mitocondri: estructura i funció
- La cadena respiratòria mitocondrial
- Genètica mitocondrial
- Desordres mitocondrials
Es necessita una quantitat suficient d'ADNmt per a la producció d'algunes de les subunitats que formen els complexos de la cadena respiratòria mitocondrial i, per tant, per a la producció d'energia. Aquesta àmplia gamma fa que sigui difícil trobar la causa molecular de les deficiències del sistema OXPHOS.

El CoQ 10
- Funcions del CoQ 10
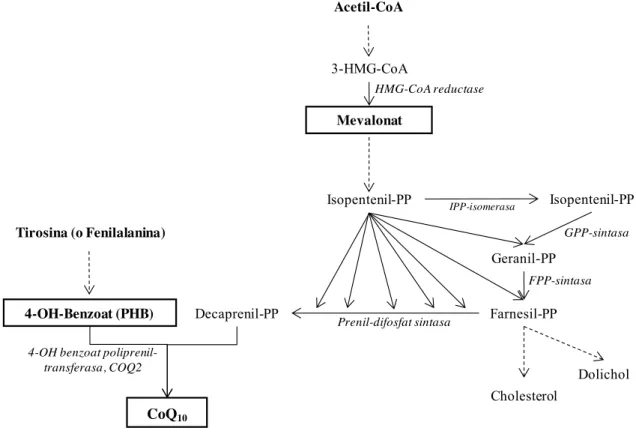
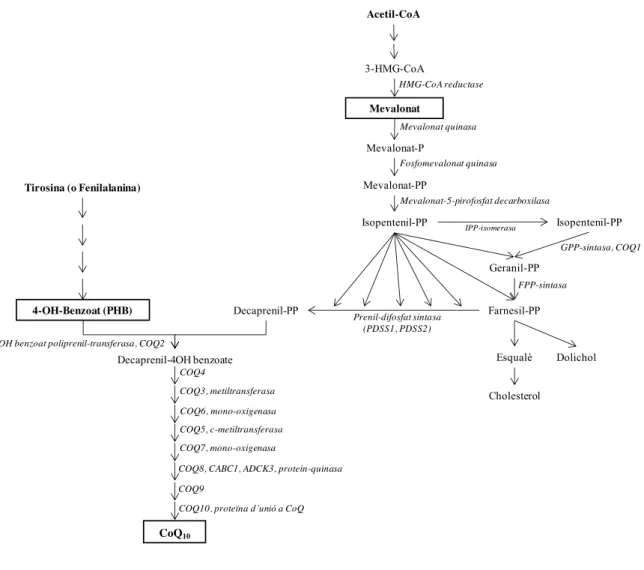
- Via de síntesis i gens implicats
- Deficiències de CoQ
El principal enzim regulador de la via del mevalonat és la HMG-CoA reductasa, que afecta principalment la síntesi de colesterol. Aquest gen codifica una de les dues subunitats de la poliprenil difosfat sintasa, el primer enzim de la via de síntesi de CoQ10.
Diagnòstic bioquímic de les alteracions del sistema OXPHOS i els defectes de CoQ 10
La majoria dels pacients amb deficiència de CoQ10 presenten anomalies en l'activitat de la cadena respiratòria mitocondrial, que s'estudien mitjançant anàlisis espectrofotomètriques. Per tant, la biòpsia muscular serà el teixit escollit per realitzar els estudis enzimàtics del sistema OXPHOS i la quantificació de CoQ10.
OBJECTIUS
Establir contactes amb altres laboratoris d'estudi d'enzims CRM per homogeneïtzar la metodologia, permetent la creació d'un punt de suport entre els diferents centres de diagnòstic. Implementar la tècnica d'electroforesi a Blue Native per a l'estudi d'enzims CRM i demostrar que és una bona eina complementària a les tècniques espectrofotomètriques.
MATERIAL I MÈTODES
OBTENCIÓ I CULTIU DE FIBROBLASTS
La biòpsia de pell es realitza en condicions estèrils, en medi de cultiu i a temperatura ambient. Sembrar els explants de manera homogènia en un matràs de 25 cm3 que conté medi MEM-GlutaMAX suplementat amb NBCS al 15% i penicil·lina-estreptomicina 125 U/ml. Deixeu el matràs en posició vertical, sense que el medi toqui els explants, a la incubadora a 37ºC i 5% CO2 durant dues hores aproximadament per assegurar-vos que estiguin ben fixats.
A continuació, col·loqueu el matràs en posició horitzontal i deixeu-lo a la incubadora fins que s'uneixi.
DETERMINACIÓ DE PROTEÏNES PEL MÈTODE DE LOWRY
Inoculeu per triplicat 5 µL de cada punt de la línia estàndard en ordre ascendent i mostra (si la mostra està massa concentrada, diluïu ¼ amb aigua MiliQ).
DETERMINACIÓ DE PROTEÏNES PEL MÈTODE DE BCA
Inocular per triplicat 10 µL de cada punt de la línia estàndard en ordre ascendent i de la mostra.
ACTIVITAT CITRAT SINTASA EN FIBROBLASTS (Mètode de Srere)
Rentar el pellet dues vegades amb solució salina (0, 9 M NaCl) i finalment tornar a suspendre amb 400 L de tampó SETH.
TEST DE VIABILITAT CEL·LULAR
Compteu les cel·les dels 4 quadrants, ignorant les cel·les que toquen la línia dels marges dret i inferior. Centrifuga la suspensió cel·lular (1 ml) de nou durant 10 minuts a 1100 rpm i torna a suspendre amb 5 ml de medi DMEM normal. Calculeu el volum de suspensió cel·lular necessari per obtenir una suspensió a una concentració de 5000 cèl·lules/100 µL.
0,1 mL = X2 mL de suspensió de cèl·lules Ajusteu el volum a 4 mL amb medi de cultiu. Mostra les cèl·lules que s'han de tenir en compte mitjançant el recompte (glòbuls blancs) i les que no s'han de comptar (glòbuls vermells). L'endemà, substituïu el medi de cultiu als pous designats per als tractaments específics per 100 µL de medi de cultiu "problemàtic" (tractat o suplementat).
Afegiu 100 µL de DMSO estèril a cada pou i torneu a suspendre els cristalls pipetant repetidament (la solució es torna porpra). Es considera que el tractament afecta la viabilitat de les cèl·lules si el percentatge aconseguit és inferior al 80%.

BIOSÍNTESI DE CoQ 10 EN FIBROBLASTS
Pesar els mg de 13C6-PHB i α-ciclodextrina necessaris segons el volum de medi d'incubació a preparar (2 mL de medi d'incubació per pou de cèl·lules). Agafeu el volum de 2H3-Meva 8,84 mM necessari segons el volum de medi d'incubació a preparar (2 mL de medi d'incubació per pou de cèl·lules). Quan el cultiu està confluent, un dia abans d'iniciar la sobrecàrrega amb substrats etiquetats, tripsinitzar-lo i inocular-lo amb medi DMEM normal en una placa de 6 pous: un matràs confluent es divideix en 4 pous.
Quan s'ha completat la sobrecàrrega, rentar les cèl·lules una vegada amb PBS estèril, tripsinitzar i centrifugar a 1100 rpm durant 10 min. Sonicar les cèl·lules dues vegades durant 5 segons cadascuna i emmagatzemar-les congelades a -80 °C fins al seu ús. En l'homogeneïtat cel·lular obtingut, determineu la concentració de proteïnes mitjançant el mètode Lowry, l'activitat de la citrat sintasa (mètode Srere) i la CoQ10 (2H3-CoQ10 i 13C6-CoQ10), que es van sintetitzar a partir dels corresponents precursors marcats.
La detecció de COQ10 es produeix en mode d'ions positius mitjançant el monitoratge de reaccions múltiples (MRM). Per comparar-lo amb l'estàndard intern (COQ9), l'àrea de cada pic es converteix en un valor de CoQ10 expressat en PMOL.
- Preparació de l’extracte de teixit
- Activitat del complex I (NADH ubiquinona oxido-reductasa)
- Activitat del complex II (Succinat ubiquinona oxido-reductasa)
- Activitat del complex III (Ubiquinol-citocrom C oxido-reductasa)
- Activitat del complex IV (Citocrom C oxidasa)
- Activitat combinada dels complexes I+III (NADH citocrom C oxido- reductasa)
- Activitat combinada dels complexes II+III (Succinat citocrom C oxido- reductasa)
- Activitat Citrat Sintasa
La disminució de l'absorbància a 340 nm es mesura quan el NADH s'oxida a NAD. L'activitat del complex III ve donada per la diferència entre l'activitat mesurada amb i sense antimicina. Es mesura l'augment de l'absorbància a una longitud d'ona de 550 nm que es produeix quan es redueix el citocrom c.
Es mesura la disminució de l'absorbància a una longitud d'ona de 550 nm que es produeix quan s'oxida el citocrom c. Prepareu una solució de citocrom C 100% reduïda: col·loqueu 600 µl de la solució inicial de citocrom C 100 µM en una cubeta semimicro i afegiu-hi uns quants grans de ditionit sòdic (la solució es tornarà rosa salmó). Es torna a blanquejar l'espectrofotòmetre amb la solució 100% oxidada i llegeix l'absorbància de la solució 100% reduïda.
Transferiu alíquotes de 5-10 µL de la solució reduïda al 100% a la solució inicial de citocrom C i torneu a llegir l'absorbància. Calcula la diferència entre l'activitat de la cubeta que conté el sobrenedant i la que només conté medi de reacció.
BLUE NATIVE ELECTROFORESIS EN GELS DE POLIACRILAMIDA (BN-PAGE) EN MOSTRES DE MÚSCUL I FIBROBLASTS
- BLUE NATIVE electroforesis
- Tinció del gel amb Blau de Comassie
- Activitats enzimàtiques en gel de Blue Native
- Western-Blot dels complexes de cadena respiratòria mitocondrial a partir d’un gel de Blue Native
Escórrer la solució de tinció (es pot reciclar) i cobrir el gel amb la solució de lleixiu Coomassie. Escorreu la solució de decoloració i hidrateu el gel amb aigua destil·lada durant almenys uns 30 minuts. El gel es pot emmagatzemar en aigua o plastificar (sistema Bio Rad).
Un cop s'executi el gel degradat, talleu-lo segons els complexos que vulgueu estudiar. Per aturar l'activitat dels complexos I, II i IV, submergiu el gel en la solució fixadora (en una campana de flux laminar) durant 15 min. Un cop fixades les bandes d'activitat, el gel es pot deixar completament submergit en aigua destil·lada fins l'endemà.
Un cop s'hagi executat el gel de gradient, separeu el gel d'apilament del gel de gradient. Talla un tros de membrana de nitrocel·lulosa de la mateixa mida que el gel.

EXTRACCIÓ D’ADN
Si el precipitat no sembla prou consistent, torneu a repetir el pas anterior, s'incuba durant 5 minuts sobre gel i es torna a centrifugar. Recollir el sobrenedant amb una pipeta estèril i transferir-lo a un tub estèril de 15 ml que conté 1 ml d'isopropanol al 100%. Descartar el sobrenedant abocant i assecar el tub amb paper absorbent, deixant-lo breument cap per avall.
EXTRACCIÓ DE ARN I OBTENCIÓ DE cADN
Afegiu 350 µl de solució d'extracció d'ARN al pellet de fibra, barregeu-lo amb una pipeta estèril i transferiu-lo a una columna QIAshredder amb tub de recollida. Afegiu el mateix volum d'etanol al 100% (350 µL) al filtrat, barregeu-lo amb una pipeta estèril i transferiu tot el contingut a la columna RNeasy mini spin amb tub de recollida. Afegiu 30-50 µl d'aigua sense RNasa directament al filtre de columna i deixeu-lo rehidratar durant 1 minut.
CERCA DE MUTACIONS EN cADN i ADN DE GENS IMPLICATS EN LA VÍA DE SÍNTESI DE CoQ 10
Un cop realitzada la PCR, assegureu-vos que s'ha amplificat correctament carregant 5 µl de producte de PCR barrejat amb 3 µl de tampó de càrrega (LB) en un gel d'agarosa al 2%. L'observació de les bandes s'aconsegueix incubant el gel durant 10-15 minuts en una solució de bromur d'etidi (intercalador de base) i exposant a llum UV amb l'equip AlphaImager.
RESULTATS I DISCUSSIÓ
Implementació d’una nova tècnica per a l’estudi espectrofotomètric de l’activitat dels enzims que formen la cadena respiratòria mitocondrial (CRM)
- Estudi espectrofotomètric de l’activitat dels enzims de CRM en teixit muscular
- Estudi espectrofotomètric de l’activitat dels enzims de CRM en fibroblasts cultivats
- Situació actual
- Estudis de complex I
- Estudis de complex II
- Estudis de complex III
- Estudi del complex IV
- Estudis de complex V
- Consideracions en els estudis enzimàtics mitjançant la tècnica del Blue Native
El complex III no es va poder investigar a causa de les dificultats en el procés de reducció de la decilubiquinona. L'estudi dels enzims de la cadena respiratòria es va realitzar de la mateixa manera que per al teixit muscular. L'activitat enzimàtica del complex I en gel es va estudiar en 3 pacients (pacients 1-3) que van mostrar poca activitat en un estudi d'activitat CRM realitzat a partir del teixit muscular.
La banda corresponent al complex II del pacient 4 (P4) era similar a la del control, mentre que estava absent o molt feble en el pacient 5 (P5) (Figura 10). El defecte de l'activitat del complex II es va confirmar en l'estudi de les activitats de BN en gel (Figura 12-A) i per a BN-WB la banda corresponent al complex II era de menor intensitat en comparació amb la mostra de control (Figura 12-B). Per tant, es va continuar l'estudi dels enzims de la cadena respiratòria mitocondrial per BN a partir de fibroblasts cultivats (figura 13), ja que no hi havia prou teixit muscular disponible.
En el pacient 10, es va observar un clar dèficit en l'activitat del complex III en l'estudi basat en la biòpsia muscular. Es van estudiar 15 pacients (pacients 19-33) amb sospita d'un defecte en l'activitat del complex V, i en 5 es va observar un canvi en l'activitat d'aquest complex. Es va observar una clara deficiència del complex V tant en les activitats en gel com en WB.
En el teixit muscular, es va observar un clar defecte en l'activitat d'aquest complex (Figura 22-A), mentre que no es va observar cap canvi en l'estudi realitzat amb fibroblasts cultivats (Figura 22-B,C).

Estudi de la via de síntesi del CoQ 10 en fibroblasts de pacients amb sospita de deficiència primària o secundària de CoQ 10
- Posta a punt de la metodologia
- Validació del mètode
- Estudi de la via de biosíntesi de CoQ 10 en pacients seleccionats
- Anàlisi d’intermediaris de la via acumulats
- Consideracions per l’estudi de la via de biosíntesi de CoQ 10 en fibroblasts
D'altra banda, s'ha especulat sobre l'existència de determinats factors genètics que poden conferir una certa susceptibilitat a desenvolupar deficiències secundàries de CoQ10 (Sacconi 2010). Els resultats obtinguts en fibroblasts van mostrar que utilitzant PHB-13C6 com a precursor de la via (Figura 25-A), la biosíntesi de CoQ10 va augmentar linealment. Biosíntesi de CoQ10 utilitzant PHB-13C6 i Meva-2H3 com a substrats per a la via a diferents concentracions i incubació. vegades.
Es va observar que la quantitat final de CoQ10 etiquetada va disminuir quan es van utilitzar concentracions de precursors Meva-2H3 superiors a 56 µM (figura 25-C). En estudiar la via biosintètica de CoQ10 mitjançant substrats marcats, es van obtenir valors dins del rang normal, demostrant que la deficiència de CoQ10 observada era secundària. En pacients afectats per NPC (pacients 6 i 7), es va observar que la biosíntesi de CoQ10 era normal quan s'utilitzava Meva-2H3 com a precursor de la via, mentre que es va produir la formació de CoQ10.
No es va observar cap millora en la concentració final de CoQ10 en l'estudi d'aquests pacients (taula 17). Cribratge mutacional de gens relacionats amb la via de síntesi de CoQ10 en pacients amb deficiència de CoQ10.