Fortalecimiento de la infraestructura de investigación y desarrollo de biotecnología con un enfoque en Una Salud: Interfaz Animal-Hombre-Ecosistema. Conservación y uso de la biodiversidad microbiana marina: mejorar la contribución de los océanos al desarrollo económico nacional. Ser un buen ejemplo para mí y para muchos otros estudiantes, por su famosa frase: que aunque la situación sea difícil hay que recordar “si la maestra Reyna pudo, por qué yo no”.
Ramón Cepeda Palacios por su ayuda con los análisis estadísticos, por su tiempo, dedicación, consejos. Fuiste el investigador más original que he conocido xD. A CIBNOR por permitirme el espacio y facilidad de transporte, y a Posgrad por la gran calidad que nos ofrece.
INTRODUCCIÓN
Los beneficios del uso de macroalgas como complemento de la dieta animal están determinados en parte por sus valores nutricionales (Evans & Critchley, 2014) y en particular por la presencia de elementos/compuestos antioxidantes e inmunoestimulantes como polisacáridos sulfatados, carotenoides, polifenoles. , vitaminas y minerales (Sachindra et al., 2007; Devi et al., 2008; García-Casal et al., 2009; Michalak et al., 2011; Hwang et al., 2015). A pesar de conocer los beneficios de este tipo de aditivos, hasta la fecha existen pocos estudios que evalúen parámetros antioxidantes y de respuesta inmune en cabras (Fike et al., 2001; Saker et al., 2001). Por lo tanto, este trabajo examinó los efectos de la suplementación con Sargassum spp.
ANTECEDENTES
- Importancia socioeconómica de los caprinos
- Sistema digestivo en rumiantes
- Sistema antioxidante en vertebrados
- Enzimas antioxidantes
- Superóxido dismutasa
- Catalasa
- Sistema inmune en vertebrados
- Respuesta inmune innata
- Respuesta inmune adaptativa
- Suplementación animal
- Macroalgas marinas como suplemento
En particular, estos tres tipos de SOD se expresan en dosis altas en el riñón (Gongora et al., 2008). Las células dendríticas (CD) juegan un papel fundamental en la regulación de la respuesta inmune. Se trata de proteínas que se forman en la respuesta inmune para combinarse específicamente y con gran fuerza con los antígenos.
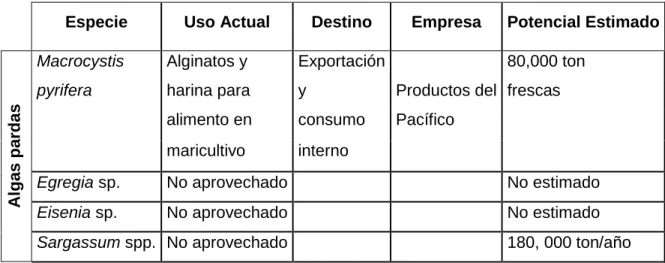
Además de ser un recurso natural renovable con amplia distribución a lo largo de la costa del Pacífico (en la costa de Baja California se reconocen cuatro especies de macroalgas pardas de interés económico, Cuadro 1) (Ortiz et al., 2006). Estos compuestos juegan un papel importante contra diversas enfermedades y procesos de envejecimiento, estimulando y fortaleciendo la respuesta inmune y antioxidante de los seres que los consumen (Cardona et al., 2013).

JUSTIFICACIÓN
La suplementación animal con algas marinas se ha informado durante siglos, y la especulación sobre su uso como fuente potencial de alimento no es nueva (Evans & Critchley, 2013). En ovinos suplementados con Sargazo se determinó la digestibilidad in vivo, la degradabilidad de la materia seca, así como el pH ruminal y los ácidos grasos volátiles (Marin et al., 2009). 2003) alimentaron con algas marrones a ovejas y luego midieron la degradación de la materia seca y la digestibilidad de la materia orgánica. También se ha utilizado otra macroalga marina parda como es Ascophyllum nodosum (Tasco, por su nombre comercial). nodosum para medir la temperatura rectal y las tasas de respiración en climas de alta temperatura.
2001) midió la capacidad reductora de corderos alimentados con pienso tratado con A. 2001) midió la respuesta de monocitos de toros alimentados con pienso tratado con Tasco.
HIPOTESIS
OBJETIVOS
Objetivo general
Objetivos específicos
MATERIALES Y MÉTODOS
- Diseño Experimental
- Homogenización de las muestras y cuantificación de proteína
- Respuesta antioxidante
- Actividad superóxido dismutasa
- Actividad catalasa
- Respuesta inmune
- Mieloperoxidasa
- Antiproteasa
- Lisozima
- Inmunoglobulina A y G (IgA-IgG)
- Análisis estadístico
Para determinar la concentración de proteína de los homogeneizados, se utilizó el concentrado de reactivo de colorante para ensayo de proteínas Bio-Rad en una microplaca, de acuerdo con las instrucciones del fabricante. Finalmente, la microplaca se leyó en un lector de placas (Bio-Rad, 3550-UV) a una longitud de onda de 595 nm. Se realizaron diluciones de las muestras, se tomaron 160 µL de cada una de las diluciones por triplicado y se dejaron reposar por 15 min con 40 µL de reactivo de Bradford, la microplaca se leyó en el lector de microplacas a 595 nm.
Los blancos se incubaron a 37°C durante 20 minutos y se midió la absorbancia a 405 nm en un lector de microplacas. Las muestras se trataron de la misma manera que el blanco 1 y se incubaron bajo los mismos parámetros. Este método se basa en la reacción de catalasa con metanol en presencia de una concentración óptima de H2O2.
Se colocaron 25 µl de tampón de ensayo diluido, 25 µl de metanol y 5 µl de peróxido de hidrógeno diluido (por triplicado para cada una de las diluciones) y 50 µl de las diluciones en una microplaca ELISA. Se agregaron 25 µL de hidróxido de potasio 10 M y 50 µL de Purpald, la placa se cubrió de la luz y se puso en agitación durante 10 min T.A. Luego se agregaron 50 µL de ácido sulfúrico 4 M y la placa se leyó a 450 nm en un lector de microplacas.
La actividad antiproteasa total se midió mediante la capacidad de las muestras para inhibir la actividad de tripsina. StatSoft, Inc. 2011) Para los análisis estadísticos se utilizó STATISTICA (Data Analysis Software System), versión 10. Se realizó un análisis de varianza unidireccional (ANOVA) para determinar el efecto de las diferentes dietas sobre los parámetros de respuesta inmune y antioxidantes.

RESULTADOS
Cuantificación de proteína
Respuesta antioxidante
- Superóxido dismutasa
- Catalasa
Determinaciones de la actividad superóxido dismutasa de las diferentes dietas suministradas a cabras en los 12 tejidos. Por otro lado, en la actividad catalasa (Cuadro IV) sólo 3 de los tejidos se vieron afectados significativamente (omaso, bazo y moco intestinal), lo que sugiere que la suplementación de Sargassum spp.

Respuesta inmune
- Mieloperoxidasa
- Antiproteasa
- Lisozima
- Inmunoglobulina A y G
Determinaciones de la actividad catalasa de las diferentes dietas suministradas a cabras en los 12 tejidos. Se representa la actividad antiproteasa (Figura 6) en el moco intestinal, expresada como porcentaje de inhibición de tripsina. El tratamiento 2 fue significativamente diferente del control y de los tratamientos 3, 5 y 6 al tener un menor porcentaje de inhibición que dicho grupo.
El tratamiento 5 tuvo diferencias significativas con respecto al resto de los tratamientos incluido el control. En la cinética de la actividad antiproteasa (Figura 7) se observa una disminución de dicha actividad en los tratamientos y 6 con respecto al control en la muestra 2, mientras que a partir de la tercera muestra se observa un ajuste entre los tratamientos 2, 5 y 6 con respecto a el control, a partir de ahí se mantuvo un incremento significativo durante el resto del muestreo. El grupo de tratamiento que mostró diferencias significativas respecto al control y el resto de grupos fue el número 4.
En la cinética de la lisozima (Figura 9), el comportamiento de dicha enzima fue errático para la mayoría de los grupos, los grupos presentaron altibajos durante 6 semanas. Hubo correlación entre el grupo Ss2.5 y Ss2.5+S3; en la tercera semana tuvieron un pico de actividad (16 y 15 U/mL, respectivamente) por encima del resto de los grupos. Los grupos Ss5 y Ss5+S3 fueron los que presentaron menor índice de actividad lisozima, incluso bajo el control.
En la Figura 10, correspondiente a IgA, los tratamientos 2, 3, 4 y 6 difirieron significativamente del control, mientras que en el quinto tratamiento no existieron. El cuarto tratamiento presentó los mayores valores de absorción, generando diferencias significativas con el resto de los grupos. En la Figura 11 se muestran las absorbancias para IgG, los grupos 2, 4 y 6 fueron los que mostraron diferencias significativas con respecto al control, incluyendo los grupos 2 y 6 fueron los tratamientos con mayores diferencias entre grupos.

DISCUSIÓN
- Suplementación con Sargassum spp. y reto con sebo rancio
- Cuantificación de proteína
- Actividad antioxidante
- Superóxido dismutasa
- Catalasa
- Respuesta inmune
- Lisozima
- Mieloperoxidasa
- Antiproteasa
- Inmunoglobulina A y G
Del mismo modo, Côrtes et al. 2012) encontraron que cuando se suplementaba con infusiones de aceite de linio no oxidado, este aceite no tenía ningún efecto sobre la actividad de SOD en plasma. En la mayoría de los tejidos, la concentración de proteínas fue mayor en los animales suplementados con Sargassum spp. Informan que no hubo diferencias estadísticamente significativas (p<0,05) en la determinación de SOD en relación con el control.
Contrario a lo mencionado, de los 12 tejidos analizados, 6 (abomaso, yeyuno, colon, músculo renal y mucosa intestinal) de ellos presentaron índices superiores en la determinación de SOD con respecto al control, estos tejidos pertenecían principalmente al grupo 4. . a excepción del abomaso y los músculos. Informan que los niveles plasmáticos de transcripción de SOD aumentaron. 2004) observaron un aumento en la actividad de SOD en llamas alimentadas con Ascophyllum nodosum y expuestas a estrés térmico. Esto es importante ya que se ha visto que el estrés oxidativo puede estar asociado con un aumento en la incidencia de trastornos de salud como la fiebre de la leche y el desplazamiento del abomaso (Miller et al., 1993).
Esto es importante ya que uno de los aspectos más importantes en la producción animal es la calidad de la carne (Surai, 2002). Sgorlon et al., (2006), tras determinar glutatión peroxidasa en ovejas suplementadas con vitamina E, licopeno y polifenoles, informaron que no hubo cambios en la actividad de la enzima en cuestión. Informan que no hubo diferencias estadísticamente significativas (p<0,05) en la determinación de catalasa y glutatión peroxidasa con relación al control.
En la cinética sérica hubo correlación entre los grupos Ss2.5 y Ss2.5+S3; en la tercera semana tuvieron un pico de actividad (16 y 15 U/mL, respectivamente) por encima del resto de los grupos. En la cinética de la mieloperoxidasa (Figura 9), se observa estabilidad en la actividad de esta enzima durante las primeras 3 semanas. Son proteínas que se forman en la respuesta inmune para combinarse específicamente y con gran fuerza con los antígenos (Parham, 2005).
CONCLUSIONES
RECOMENDACIONES
LITERATURA CITADA
Mammary gene expression and antioxidant enzyme activity and concentration of the mammalian lignan enterolactone in milk and plasma of dairy cows fed flax lignan and injected with linseed oil in the abomasum. Effect of hot season and feeding on oxidative status and metabolic profile in mid-lactation dairy goats. Hepatic and cardiac oxidative stress and other metabolic changes in broilers with ascites syndrome.
Use of subtherapeutic antibiotics in livestock (as supplements or feed additives) with the induction of resistance in human bacterial pathogens, the extent and complexity of the problem is becoming increasingly clear. Oxidation of biological systems: oxidative stress phenomena, antioxidants, redox reactions and method for their quantification. Effect of macroalgae enriched with microelements on egg quality parameters and mineral content of eggs, eggshell, blood, feathers and manure.
Effect of vitamin E supplementation on growth, nutrient utilization, mineral balance and immune status of goats exposed to arsenic. Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea antarctica. Effect of dietary selenium and vitamin E content on lipid peroxidation in breast muscle tissue of broiler chickens during storage.
Physiological Aspects of Ruminant Digestion and Metabolism, Proceedings of the Seventh International Symposium on Ruminant Physiology, Academic press Inc. Effects of feeding dietary antioxidant in oxidized fat diets on lactation and antioxidant status of the cow. Effect of seaweed supplementation on the oxidative status of plasma in dairy cows during the periparturient period.