ESCUELA TÉCNICA SUPERIOR DE INGENIERÍAS
AGRARIAS
Departamento de Producción Vegetal y Recursos Forestales
CLIMATE AND FOREST GROWTH IN MEDITERRANEAN
ENVIRONMENTS:P
INUS PINASTER ANDP
INUS SYLVESTRIS INS
PAINPresentada por
Stella Marys Bogino
para optar por el título
de Doctora
Dirigida por
Dr. Felipe Bravo Oviedo
Dra. María José Fernández Nieto
UNIVERSIDAD DE VALLADOLID
ESCUELA TÉCNICA SUPERIOR DE INGENIERÍAS
AGRARIAS
Departamento de Producción Vegetal y Recursos Forestales
CLIMATE AND FOREST GROWTH IN MEDITERRANEAN
ENVIRONMENTS:P
INUS PINASTER ANDP
INUS SYLVESTRIS INSPAIN
Stella Marys Bogino
Ingeniera Agrónoma
Especialista en Docencia Universitaria
Magíster en Ciencias Agropecuarias
Tesis doctoral
Directores
Dr. Felipe Bravo Oviedo
Dra. María José Fernández Nieto
CERTIFICAN:
Que la Ingeniera Agrónoma (M.Sc.) Stella Marys Bogino ha realizado bajo nuestra dirección el trabajo que, para optar al Grado de Doctor, presenta con el título “Climate and forest growth in Mediterranean environments: Pinus pinaster and Pinus sylvestris in Spain”.
Por lo que mediante este Trabajo de tesis, se solicita el reconocimiento del Doctorado europeo de la doctoranda, quien realizó una estancia de cuatro meses en la
Georg-August Universität, Göttingen, Alemania, bajo la supervisión del Dr. Christoph
Klein (Profesor titular de dicha Universidad).
Y para que así conste a los efectos lo firmamos en Palencia, a 30 de Junio de 2008.
Vº.bº Vº.bº
ABSTRACT
Climate change will affect Mediterranean forests by modifying driving factors related to different processes and functions. Relationship between tree growth and climate is crucial to develop forestry strategies to mitigate climate change impact. Dendroclimatological techniques were applied in sampling sites placed in central Spain to analyse the association between Pinus pinaster and Pinus sylvestris and
climatic variables in Mediterranean environments. The objectives of this thesis were: a) To analyse the relationship between P. sylvestris and P. pinaster tree-ring width and
climatic variables (precipitation and temperature). b) To explore possible shifts in the association between climate variables and tree growth over time. c) To investigate the effects of drought and pulse and interpulse water events on radial growth of P. sylvestris. d) To explore the age effect on climate-growth relationship of P. sylvestris.
e) To estimate the potentiality of Intra-annual density fluctuation (IADFs) in P. pinaster as dendroclimatological proxy. f) To estimate the climatic variables that drive
the carbon thirteen variability (δ13C) in tree ring of P. sylvestris and P. pinaster. g) To
analyse the variability of δ13C related to water use efficiency in both species. Results showed that radial growth of P. pinaster and P. sylvestris was positively correlated
with rainfall during and prior to the growing season. Mean temperature effect varied according to site altitude. P. pinaster has a changing association between radial growth
and climate variables, initiated in the 1980s. A climatic response according to the age was also found in P. sylvestris: young stands have higher variability than old stands.
Correlation between successive years was also detected in old stands. There was a significant association between radial growth of P. sylvestris and drought index (DRI).
The periodicity related to pulse and interpulse events was two or two and a half years suggesting the strong impact of the interannual precipitation variation on radial growth. IADFs in P. pinaster showed: an increase in IADF frequency from the 40
decade to the present; the mean frequency of IADF was higher in younger than in older trees; a negative correlation between radial growth and IADFs and the IADFs may be predicted using a logistic model and monthly rainfall and temperature as independent variables. The values of δ13C in P. pinaster and P. sylvestris were affected by moisture,
rainfall, maximum temperature and solar radiation. P. pinaster and P. sylvestris
RESUMEN
El cambio climático afectará los bosques mediterráneos pues altera los factores que controlan sus diferentes funciones y los procesos que tienen lugar en ellos. La relación existente entre el crecimiento de los árboles y el clima es esencial para aplicar y desarrollar estrategias forestales cuyo objeto sea mitigar el impacto del cambio climático. En este contexto, se aplicaron distintas técnicas dendroclimatológicas en varios sitios de muestreo emplazados en el centro de España, para analizar la relación entre Pinus pinaster y Pinus sylvestris y las variables climáticas en los ambientes
Mediterráneos. Los objetivos de esta tesis fueron: a) Analizar la relación entre el ancho de los anillos de crecimiento y las variables climáticas (precipitación y temperatura). b) Explorar posibles cambios en la asociación entre las variables climáticas y el crecimiento de los árboles a través del tiempo. c) Investigar los efectos de la sequía y de los pulsos e interpulsos de agua sobre el crecimiento radial de P. sylvestris d)
Explorar el efecto de la edad de los árboles en la relación clima-crecimiento de P. sylvestris. e) Estimar la potencialidad de las variaciones interanuales de la densidad
(FIAD) en P. pinaster como herramienta dendroclimatológica. f) Estimar las variables
climáticas que controlan la variabilidad del carbono trece (δ13C) en los anillos de crecimiento de P. sylvestris y P. pinaster. g) Analizar la variabilidad del δ13C en
relación con el uso del agua, en las dos especies. Los resultados demostraron que el crecimiento radial de P. pinaster y P. sylvestris se correlaciona positivamente con la
precipitación, tanto antes como durante la estación de crecimiento. El efecto de la temperatura media varió de acuerdo con la altitud del sitio de muestreo. La relación entre el crecimiento radial de P. pinaster y las variables climáticas mostró una
variación a través del tiempo que se inició en la década del ochenta. P. sylvestris
manifestó una respuesta climática en función de la edad: los árboles jóvenes tienen más variabilidad de su crecimiento radial explicada por el clima, que los árboles viejos. En los árboles viejos se detectó correlación entre los anillos de crecimiento de años sucesivos. Hay una correlación significativa entre el crecimiento radial de P. sylvestris
y el índice de sequía relativa. La periodicidad relacionada con los pulsos e interpulsos de agua fue de dos y dos años y medio, lo que sugiere un fuerte impacto de la variación interanual de la precipitación sobre el crecimiento radial. Las variaciones intraanuales de la densidad mostraron: un aumento de la frecuencia de FIAD en P. pinaster desde la
década de los cuarenta hasta la actualidad; la frecuencia media de FIAD fue más alta en los árboles jóvenes que en los viejos y también se detectó una correlación negativa entre el crecimiento radial y las FIAD. Las FIAD podrían predecirse usando un modelo logístico que tiene a la precipitación y la temperatura mensual como variables independientes. La variabilidad del isótopo carbono trece (δ13C) en P. pinaster y P. sylvestris está afectada por la humedad, la precipitación, la temperatura máxima y la
radiación solar. En los ambientes Mediterráneos, P. pinaster y P. sylvestris son
AGRADECIMIENTOS
A mi director Felipe Bravo Oviedo y Co-Directora María José Fernández Nieto, por su apoyo permanente.
A las Universidades de Valladolid, España, y Georg-August Universität, Göttingen,
Alemania, en las que desarrollé este programa de doctorado.
Al Programa Alβan, Programa Europeo de Becas de Alto Nivel para América Latina, beca # E05D049920AR; al Programa de Becas Erasmus de la Universidad de Valladolid y a los
proyectos de la Junta de Castilla y León, España, Código VA096A05, y del Ministerio de Educación y Ciencia, España, Códigos AGL2004-07094-C02-02 y AGL2007-65795-C02-01 por la ayuda económica.
A la Agencia Estatal de Meteorología, España, por haberme facilitado los datos climáticos. A la Universidad Nacional de San Luis, Argentina, y a la Escuela Remedios Escalada de San Martín, Argentina, por la licencia otorgada durante estos tres años para la realización del presente doctorado.
Un especial agradecimiento a Vicente Rozas, Esteban Jobbágy y Mariano Morales por los inestimables comentarios realizados a esta tesis. A María Laura Cangiano por las correcciones del texto en Inglés, a Iñaki Etxebeste Larrañaga y Reinhard Langel por la inestimable ayuda en los capítulos de densidad e isótopos, respectivamente, y a Cristóbal Ordóñez por la ayuda en el trabajo de campo.
A Wilson Lara, Iñaki Etxebeste Larrañaga y Javier Castaño, por la edición final de la tesis. A Celia Redondo por su ayuda incondicional con la documentación, siempre que la he necesitado.
A aquellas personas que me han hecho sentir la vida un poco más fácil durante estos tres años lejos de mi querido país: en España: Encarna Rodríguez García y familia, Antonio Sanz, María Gómez, Antonio Urchaga, Pilar Valbuena, Claudia Escudero, Jorge Martín García y familia, Iñaki Etxebeste Larrañaga, Leticia Botella, Yésica Palavecini, Celia Herrero de Aza y familia, Patricia Recio, Lucía de Soto Suárez, Darío Martín Benito, José Reque y familia, Wilson Lara, Javier Castaño, Amelework Kassa, Luis Fernando Osorio, Gonzalo Álvarez Baz, Laura Benito y Charo Litardo y familia. En Argentina: Elena Riccio, María Victoria Vega, Mirta Gómez, Zunilda Furlán, Hernando Casagrande, Liliana Gabutti, Edgardo Guerrero, Graciela Ardú, Emilio Lotto, Ana María Bogino, Andrés Bogino, Daniel Bogino, Anselmo Avila, María Pachoud, Patricia Pagnone, Lautaro Barbaglia, Agostina Serrano, Rosita Giraudo, Zoraide Fernández Montero, Salvador Micali, Ana María Careaga, Claudia Dustchazky, Olga Callovi, Iván Niveyro, Patricia Vidal, María Eugenia Moreno, Maximiliano Dellacassa y Nedda Rimondi. En Alemania: Hardy Schilling, Frank Bräuer, Michael Barthel, Lena Hollmann, Frank Stein, Anja Sölter, Nicola Köberl and Benjamin Jung.
A mis alumnos de la Escuela y de la Universidad en Argentina, por todo lo que me han enseñado durante estos 20 años. Al estudiante Juan Politano quien dio su vida luego de salvar a mucha gente en la explosión de la Universidad de Río Cuarto el 5 de diciembre de 2007, mi dolor y mi gratitud.
ACKNOWLEDGEMENTS
Thanks to my director Felipe Bravo Oviedo and Co-Director María José Fernández Nieto for their permanent support.
To the Universidad de Valladolid (Spain) and Georg-August Universität, Göttingen
(Germany) where I did this PhD Program.
To the Programme Alβan, the European Union Programme of High Level Scholarships for Latin America, scholarship # E05D049920AR; the Erasmus Scholarship of the Universidad de Valladolid and the research projects from the Regional Castilla y León government (Spain),
Project Code VA096A05 and the Ministerio de Ciencia y Educación (Spain), Project Code
AGL2004-07094-C02-02 and AGL2007-65795-C02-01 for economical support.
To the Agencia Estatal de Meteorología (National Meteorological Agency, Spain) for
providing meteorological data.
To the Universidad Nacional de San Luis (Argentina) and the Escuela Remedios Escalada de San Martín (Argentina) for the extended leave they gave me along these three years.
Special thanks to Vicente Rozas, Esteban Jobbágy and Mariano Morales for the very useful comments made to this work. Thanks are also extended to María Laura Cangiano for improving the English version, to Iñaki Etxebeste Larrañaga and Reinhard Langel for helping me with the density and isotope chapters, respectively, and to Cristobal Ordoñez for field assistance.
Thank are also extended to Wilson Lara, Iñaki Etxebeste Larrañaga and Javier Castaño for the final edition of the thesis.
To Celia Redondo for helping me with my documents whenever I needed it.
To those people who have supported me throughout these three years far away from my beloved land. In Spain: Encarna Rodríguez García and family, Antonio Sanz, María Gómez, Antonio Urchaga, Pilar Valbuena, Claudia Escudero, Jorge Martín García and family, Iñaki Etxebeste Larrañaga, Leticia Botella, Yesica Palavecini, Celia Herrero de Aza and family, Patricia Recio, Lucía de Soto, Darío Martín Benito, José Reque and family, Wilson Lara, Javier Castaño, Amelework Kassa, Laura Benito, Charo Litardo and family, Gonzalo Álvarez Baz and Luis Fernando Osorio. In Argentina: María Victoria Vega, Mirta Gómez, Zunilda Furlán, Hernando Casagrande, Liliana Gabutti, Edgardo Guerrero, Graciela Ardú, Emilio Lotto, Ana María Bogino, Andrés Bogino, Daniel Bogino, Anselmo Avila, María Pachoud, Patricia Pagnone, Lautaro Barbaglia, Agostina Serrano, Rosita Giraudo, Zoraide Fernández Montero, Salvador Micali, Elena Riccio, Edith Rojas, Ana María Careaga, Claudia Dustchazky, Olga Callovi, Iván Niveyro, Patricia Vidal, María Eugenia Moreno, Maximiliano Dellacassa and Nedda Rimondi. In Germany: Hardy Schilling, Frank Bräuer, Michael Barthel, Lena Hollmann, Frank Stein, Anja Sölter, Nicola Köberl and Benjamin Jung.
To my school and university students for everything you have taught me along these years. My sorrow and gratitude to the student Juan Politano, who died after saving the life of many people in the explosion at the Universidad Nacional de Río Cuarto the 5th of December 2007.
mi dulce y amada hija María Laura.
Si piensan en las cualidades que una persona perfecta debería tener, tal vez puedan comprender cómo es ella.
I think about you and I am very happy. This thesis is for you, my sweet and beloved daughter María Laura.
CONTENTS
ABSTRACT... vii
RESUMEN ...ix
AGRADECIMIENTOS ...xi
ACKNOWLEDGEMENTS ... xiii
CONTENTS...xvii
1. INTRODUCTION ...1
1.1. Climate change and Mediterranean environments ...1
1.2. Species studied...1
1.2.1. Pinus pinaster Ait...1
1.2.2. Pinus sylvestris L....2
1.3. Dendrochronological tools available for climate growth relationship analysis..3
1.3.1. Tree-ring growth...5
1.3.2. Intra-annual density fluctuations (IADFs) ...5
1.3.3. Isotopes...5
1.4. Objectives ...6
1.4.1. Tree-ring growth-climate relationship ...6
1.4.2. Tree-ring growth-atmospheric indexes relationship ...6
1.4.3. Tree-ring growth-water availability relationship ...6
1.4.4. Intra-annual density fluctuations ...7
1.4.5. Isotope study...7
2. MATERIALS AND METHODS ...9
2.1. Study sites...10
2.2. Dendrochronological methodology...10
2.2.1. Fiel and laboratory work...10
2.2.2. Climate-growth relationship ...11
2.2.3. Intra-annual density fluctuations analysis...13
2.2.4. Isotope analysis ...14
2.3. Climatic data...15
3. RESULTS ...17
3.1. Chronology description...17
3.2. Climate growth association: multiple stepwise and bootstrapped analysis ...17
3.3. Climate growth association over time: the Kalman filter technique in P. pinaster...17
3.4. NAO and SOI effect on radial growth ...18
3.5. Climate and radial growth of P. sylvestris...18
3.6. IADFs and climate results...19
3.7. Carbon isotopes and climate results...19
4. DISCUSSION ...21
5. CONCLUSIONS...27
6. CONCLUSIONES ...29
REFERENCES ...31 LIST OF ORIGINAL ARTICLES... I Growth response of Pinus pinaster Ait. to climatic variables in central Spanish forests
Bogino (2008)
xviii
SOI and NAO impacts on Pinus pinaster Ait. growth in Spanish forests... V
Resumen... V Drought and climate effect on radial growth of Pinus sylvestris L. in its southern and
western distribution threshold... VII Resumen... VII Climate and intra-annual density fluctuations in Pinus pinaster in Spanish woodlands
...IX Resumen...IX Stable carbon isotope 13C in Pinus pinaster and Pinus sylvestris tree rings: climatic
signals and water use efficiencyin Mediterranean environments...XI Resumen...XI
TABLA DE CONTENIDOS
RESUMEN ...ix
AGRADECIMIENTOS ...xi
CONTENIDOS...xvii
1. INTRODUCCIÓN ...1
1.1. El cambio climático y los ambientes Mediterráneos...1
1.2. Especies estudiadas...1
1.2.1. Pinus pinaster Ait...1
1.2.2. Pinus sylvestris L....2
1.3. Herramientas dendrocronológicas para el análisis de la relación crecimiento-clima ...3
1.3.1. El ancho de los anillos de crecimiento...5
1.3.2. Las fluctuaciones intraanuales de la densidad (FIAD)...5
1.3.3. Los isótopos...5
1.4. Objetivos...6
1.4.1. Relación entre el clima y el ancho de los anillos de crecimiento...6
1.4.2. Relación entre los índices atmosféricos y el ancho de los anillos de crecimiento ...6
1.4.3. La relación entre la disponibilidad de agua y el ancho de los anillos de crecimiento ...6
1.4.4. Fluctuaciones intraanuales de densidad (FIAD) ...7
1.4.5. Estudio de isótopos...7
2. MATERIALES Y MÉTODOS ...9
2.1. Los sitios de estudio...10
2.2. La metodología dendrocronológica...10
2.2.1. El trabajo de campo y de laboratorio ...10
2.2.2. La relación del clima con el crecimiento ...11
2.2.3. El análisis de las fluctuaciones intraanuales de densidad...13
2.2.4. El análisis de isótopos...14
2.3. Los datos climáticos...15
3. RESULTADOS...17
3.1. Descripción de las cronologías ...17
3.2. Relación clima-crecimiento: regresión múltiple y análisis bootstrapped ...17
3.3. La relación del clima con el crecimiento a través del tiempo: el filtro de Kalman en P. pinaster...17
3.4. El efecto de la NAO y del SOI sobre el crecimiento radial...18
3.5. Clima y crecimiento radial de P. sylvestris...18
3.6. FIAD y clima ...19
3.7. Los isótopos del carbono y el clima...19
4. DISCUSION ...21
6. CONCLUSIONES ...29
BIBLIOGRAFÍA ...31 LISTA DE ARTÍCULOS ORIGINALES... I La respuesta del crecimiento de Pinus pinaster Ait. a las variables climáticas en los
Bogino (2008)
2
El impacto deL SOI y de la NAO sobre el crecimiento de Pinus pinaster Ait.en los
bosques esañoles... V Resumen... V El efecto de la sequía y del clima sobre el crecimiento radial de Pinus sylvestris L. en
el límite de su distribución sudoeste. ... VII Resumen... VII El clima y las fluctuaciones intraanuales de densidad en Pinus pinaster en los bosques
españoles...IX Resumen...IX El isótopo carbono 13C en los anillos de crecimiento de Pinus pinaster y Pinus
sylvestris: señal climática y eficiencia en el uso del agua en los ambientes
1. INTRODUCTION
1.1.CLIMATE CHANGE AND MEDITERRANEAN ENVIRONMENTS
Climate change will affect Mediterranean forests by modifying driving factors related to different processes and functions. Complex interactions between climate and biotic and abiotic factors are leading to more complex forest management in the future. Mediterranean environments in the Iberian Peninsula, which are water limited ecosystems, are characterised by summer droughts and high interannual variability of precipitation and temperature. In summer months, when temperature is favourable for growth, moisture is limiting; whereas in winter, when moisture is available, low temperature limits further growth (Mooney and Dunn, 1970). In arid and semi-arid ecosystems, where water is a limiting resource, water availability experiences two phases: pulse, when the resource is available, and interpulse, when water availability is too low for plant use (Noy-Meir, 1973; Goldberg and Novoplansky, 1997).
Projections of climate change impact on the European distribution of higher plants in 2050 suggest that the Iberian Peninsula could be one of the most vulnerable areas for species lost (Bakkenes et al., 2002). Climate shifts have already taken place in the region, as exemplified by the reduction of rainy days by 50 and 30% over the southern coast of Spain and the Pyrenees in the last century, respectively (Esteban-Parra et al., 1998; Rodrigo et al., 2000). On the other hand, the mean annual temperature has increased about 1.6º C in the Iberian Peninsula over the last century (Parry, 2000; IPCC, 2007).
Global change effect on tree growth is essential to understand tree growth and productivity response to the warming environment, and the impact that this response could have on global warming and on the other hand, is crucial to understand and model the carbon sequestration process in forests and to develop forestry strategies to mitigate climate change impact.
1.2.SPECIES STUDIED
1.2.1. Pinus pinaster Ait.
The Mediterranean Maritime pine (Pinus pinaster Ait.) is a characteristic species
in Mediterranean forests, and is mainly located in the Iberian Peninsula (Blanco et al., 1997). This species shows a wide ecological range (Nicolas and Gandullo, 1967), and can survive under high or low temperatures, under regular or variable rainfall as well as under severe droughts. It is adapted to the extremely cold winters of the continental climate in central Spain and to the mild, temperate winters of the Atlantic coast in the western Iberian Peninsula [Alía et al., 1996; Blanco et al., 1997 (Figure 1)].
Bogino (2008)
2
and high temperatures in summer. The soils are sandy and rocky and very well drained. Even though the Mediterranean Maritime pine had been used widely on plantations in the Iberian Peninsula, this species grows naturally in some areas, where it is the best adapted woody species (Blanco et al., 1997). Fossil P. pinaster cones and
pollen dating from the Superior Pliocene have been found in the Iberian Peninsula, suggesting a Pre-Mediterranean origin for this species and evolution in tropical-like environments that gradually incorporated seasonal aridity (Di Castri, 1981). Authors suggest that the Iberian System is the natural origin of and centre of distribution for this species (Blanco et al., 1997).
The Mediterranean Maritime pine has rarely been used for Dendrochronology and Dendroclimatology because stands older than 100 years are very difficult to find. In addition, trees from the lower mountain level often have false rings and most trees have been damaged by resin harvest in the past (Schweingruber, 1993).
1.2.2. Pinus sylvestris L.
Scots Pine (Pinus sylvestris L) covers a very wide ecological spectrum: across
Eurasia, together with other species, it dominates forest landscapes from boreal region in northern Europe and Russia to the western Mediterranean mountains in southern Europe.
0º
200 km
40 Nº ATLANTIC
OCEAN
PORTUGAL
FRANCE
MEDITERRANEAN SEA
Figure 1. Distribution of P. pinaster (gray), and P. sylvestris (black) in Spain
(Adapted from www.mma.es).
around 8ºC. On the northern forest border, temperatures in July are about 10ºC, while in the Mediterranean region they can reach 22ºC. In Western Europe, annual precipitation is 2500 mm, but in southern Europe only 400 mm. On all natural sites, Scots pine grows up to 300 year old and it could reach 600 years on dry sites.Scots Pine is the most widely distributed species of pine in the world (Blanco et al., 1997); along with Norway spruce [Picea abies (L.) H. Karst.], it possesses the highest
dendrochronological potential in Europe (Schweingruber, 1996). Forests of
P. sylvestris in Spain are the southern and western distribution threshold worldwide of
that species and occupy drier areas than in the other parts of the world (Barbéro et al., 1998; Figure 1). These dry areas of distribution of this species which usually grows in humid environments are the first places to investigate the effects of increased aridity (Martínez Vilalta and Piñol, 2002). Besides, in assessing the impact of global warming on ecosystems, any changes in tree growth are likely to occur first in those tree stands placed at the ecological boundary of the species (Tessier et al., 1997). In Spain,
P. sylvestris stands occupy 1 210 000 ha, split approximately in 50% pure and 50%
mixed stands (DGCN, 1998; 2002).
1.3.DENDROCHRONOLOGICAL TOOLS AVAILABLE FOR CLIMATE GROWTH
RELATIONSHIP ANALYSIS
Dendrochronology is the science of dating tree rings of woody species. Due to the cambial activity, a tree ring is formed every year and the analysis of tree rings provides useful information about both the structure of dated rings and applications to environmental and historical questions (Kaennel and Schweingruber, 1995). Dendroecology is a sub field of Dendrochronology which utilizes dated tree rings to study ecological problems and the environments, for example: endogenous disturbances due to interactions between trees; exogenous disturbances as for example: climate impact, fire, insect outbreaks, etc. (Figure 2).
4
1959. The seed germinates. The tree starts its life cycle. Cuban revolution happens.
1969. For several years one stem affects tree growth. So reaction
wood is produced. Neil Armstrong walks on the
Moon.
1977. Crowns and roots of trees start to compete for light and water. Vicente Alexaindre is awarded with the
Literature Nobel Price.
1979. Some trees are cut down. As the tree number is reduced, more environmental resources are available for the
remaining trees. Snow falls on Sahara.
1985. A dramatic drought period starts. Growth decreases due to the lack of water. Nevado del Ruiz volcano erupts in Colombia,
killing an estimated 23,000 people. 1996. A fire affects the woodland.
The tree survives thanks to its thick bark. The Talibans seize power in Afghanistan.
Viking I and II arrive in Mars. 1999. Tree growth was strongly
affected by an insect outbreak. Trees start to compete again and are
very weak.
The outbreak is repeated for several years.
War is in Kosovo. 2007. Tree dies at 48 years old and turns into a
wildlife refugee. Decay starts. My daughter María Laura is 20 years old.
1961. Tree grows without competition. Light and moisture conditions are adequate.
Herbivores take no notice of this seedling. The Berlin wall is built.
1.3.1. Tree-ring growth
Tree-ring widths of conifers offer some of the best climatological reconstructions of semiarid regions (Schweingruber, 1996), as in the case of annual precipitation estimates in Mediterranean climates (Tessier et al., 1997). On the other hand, different climatic variables (mainly derived from precipitation and temperatures but also derived from atmospheric pressure balance) can be used to express climatic oscillation and to derive relationships between tree growth parameters and climatic signal (D’Arrigo and Jacoby, 1992; Jones et al., 1997).
Tree-ring growth is influenced by several simultaneous environmental factors: solar radiation, temperature, water precipitation, soil nutrient content, etc. Depending on conditions and species, one or more of these factors can become limiting for tree growth (Fritts, 1976). Climate variability often explains past tree growth (Hughes, 2002), helping in the projection of future growth responses in the context of climate change (Yeh and Wensel, 2000).
1.3.2. Intra-annual density fluctuations (IADFs)
Intra-annual density fluctuations (IADFs), which include false rings, growth bands, double rings and multiple rings, are anomalies in ring growth that are formed by latewood-like cells within the earlywood, or earlywood-like cells within the latewood (Fritts, 1976).
When IADFs are properly identified, intra-annual structures from trees growing in xeric environments may be used to develop records of growing seasons characterised by early drought and followed by moist conditions (Villalba and Veblen, 1996). Species growing under a Mediterranean climate, which is characterised by summer drought and high inter-annual variability in precipitation and temperature, commonly show special anatomical characteristics in tree rings (Schweingruber, 1993). Consequently, the importance of incorporating intra-annual features or anomalies in radial growth may be useful for ecological and climatological interpretation (Tessier et al., 1997).
1.3.3. Isotopes
The isotopic composition of carbon stored in the growth rings of trees may represent a record of variations in 13C/12C as a result of physiological responses to environmental changes (Francey and Farquhar, 1982). This ratio is expressed as δ13C, the proportional deviation of the 13C/12C ratio from the international accepted PDB carbonate standard (Craig, 1957). Since cellulose is not transferred between annual growth rings, intra and interannual seasonal events are recorded permanently in δ13C signal in tree ring (Tans et al., 1978).
Bogino (2008)
6
related to each other (Farquhar et al., 1989). The crucial interdependence between carbon and water relations of plants can be particularly well studied by the use of stable carbon isotopes. Determination of δ13C was suggested as a sensitive long-term monitoring of physiological changes (Francey and Farquhar, 1982).
1.4.OBJECTIVES
This thesis has a main objective: to explore the impact of climate on pine trees growth under Mediterranean conditions. This general objective is asses by the following specific objectives.
1.4.1. Tree-ring growth-climate relationship
The relationship between tree-ring growth and climatic variables is essential to predict the future growth trend of P. pinaster and P. sylvestris. The knowledge of the
growth response of these species to past climate variability can help us to explore how its populations, and the ecosystems they dominate, will behave in the future.
Objective 1: To analyse the relationship between P. pinaster and P. sylvestris
tree-ring width and climatic variables (precipitation and temperature), and to explore possible shifts in the association between these climate variables and tree growth over time as a result of changing environmental conditions.
1.4.2. Tree-ring growth-atmospheric indexes relationship
There are no previous studies made on the impact of the SOI and NAO indexes on conifers growing in the Iberian Peninsula. As many scientists argue about both indexes have global impact on the earth’s surface, it could be an excellent opportunity to analyse the relationship between these indexes and the growth of woody species.
Objective 2. To analyse the relation between the P. pinaster‘s tree-ring width and
the NAO and SOI atmospheric indexes in Eastern Spain.
1.4.3. Tree-ring growth-water availability relationship
In the Iberian Peninsula, P. sylvestris grows in its southern and western distribution
threshold worldwide and under water limited conditions. Therefore, dendroclimatological studies may provide valuable information about both, the association with climatic variables and the interannual water availability effect (pulse and interpulse) on radial growth.
Objective 3. To investigate the drought effect and pulse and interpulse water events on radial growth of P. sylvestris and to analyse cambial age effect on
1.4.4. Intra-annual density fluctuations
As conifers growing in dry environments tend to produce false rings, and as dendroclimatological studies concluded that P. pinaster is very susceptible to rainfall
during the growing season [especially in spring and early summer (Paper 1)], the study of IADFs in P. pinaster may be a useful tool for enhancing dendroclimatological
studies based on tree-ring width fluctuations, and for reconstructing past intra-annual climate events.
Objective 4. To estimate the potentiality of intra-annual density fluctuation (IADFs) features in P. pinaster as dendroclimatological proxy.
1.4.5. Isotope study
There is not information about stable carbon isotope in tree rings of P. pinaster and P. sylvestris growing under Mediterranean climate conditions. Stable carbon isotope
variability may provide very valuable information about the climatic variables which affect this variability and WUE as a result of globally changed conditions.
Objective 5. To estimate the climatic variables that determine the δ13C signal in
P. sylvestris and P. pinaster tree rings, and to analyse the variability in δ13C in relation
2. MATERIALS AND METHODS
Dendroclimatological tools (tree-ring width, IADFs and carbon isotopes) were applied on Pinus pinaster and Pinus sylvestris, two species that constitute pure and
mixed forests in Mediterranean ecosystems in the Iberian Peninsula. Dendroclimatology, analysing tree ring and climate association, give information about past growth-climate relationship which may provide very valuable information about future tree growth on changing climatic conditions. Linear and non-linear models were applied to estimate this association between climatic conditions and tree ring. Figure 3 summarizes the main facts of the thesis.
Mediterranean terrestrial Ecosystems
Pinus pinaster Pinus sylvestris
Linear and non-linear Models Climate
Temperature Precipitation Solar radiation
Humidity Atmospheric indexes Forest communities
Radial growth
δ13C
IADF TREE RING Mediterranean terrestrial
Ecosystems
Pinus pinaster Pinus sylvestris
Linear and non-linear Models Climate
Temperature Precipitation Solar radiation
Humidity Atmospheric indexes Forest communities
Radial growth
δ13C
IADF TREE RING
Figure 3. Main facts of the thesis that relate the parts involved in the dendroclimtological study of
P. pinaster and P. sylvestris in the Iberian Peninsula. Dark lines mean incoming variables and
Bogino (2008)
10
2.1.STUDY SITES
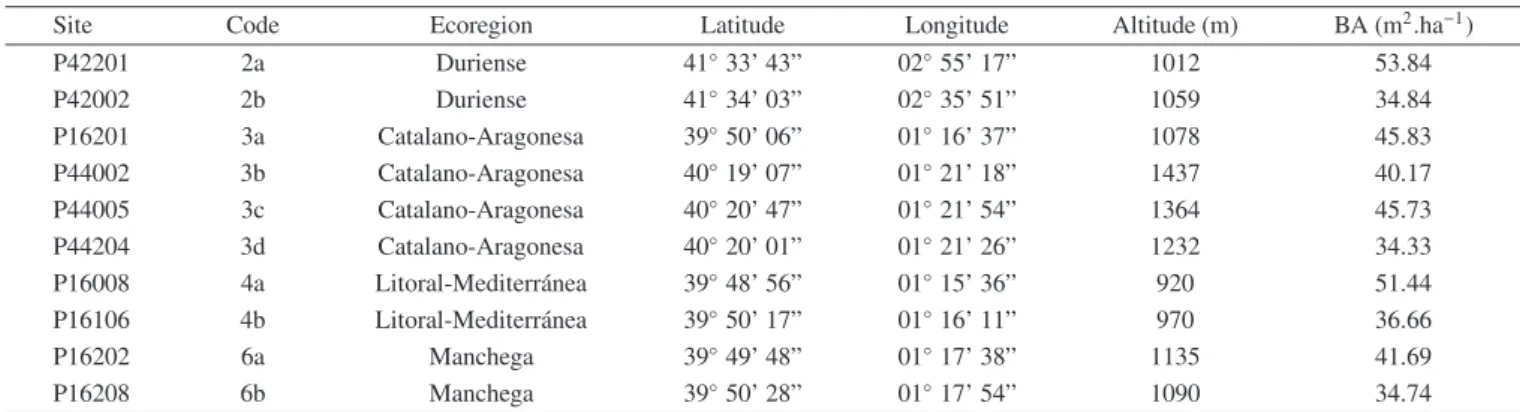
Ten and six sampling sites for P. pinaster and P. sylvestris, respectively, were
selected from different bioclimatological ecoregions where these species grow in the Iberian Peninsula. Each ecoregion has common physiographic, climatic and lithological characteristics (Elena Roselló et al., 1997; Table 2.1).
2.2.DENDROCHRONOLOGICAL METHODOLOGY
2.2.1. Fiel and laboratory work
During the summer of 2006, two cores were extracted, at 1.30 m above ground level, from fifteen dominant and co-dominant trees at each sampling site. As growth of tapped trees may be affected by resin extraction, such trees were avoided. Cores were glued on to channelled wood, dried for two weeks and polished with progressively thinner sandpaper. In order to detrmine the calendar year in which a tree ring was formed, tree rings were dated by use of a binocular microscope Nikon SMZ1000 (20X), according to standard dendrochronological techniques (Stokes and Smiley, 1968; Fritts, 1976; Cook and Kairiukstis, 1990). The cores in transverse sections were scanned at high resolution (2.000 dpi) with an Epson Expression 1640 XL scanner (0.01-mm accuracy), and rings were measured by use of the WinDENDRO programme (Regent Instrument Inc., 2002). The v6.06P COFECHA programme (Holmes, 2001; Grissino-Mayer, 2001; available at www.ltrr.arizona.edu) was applied to assess measurement and dating accuracy. COFECHA allows calculation of the Pearson’s correlation indices for the indexed tree-ring series and a master reference chronology
Table 2.1. Ecoregion classification, geographical position and mean basal area of ten P. pinaster (Pp)and six P.sylvestris (Ps) sampling sites across its natural distribution area in the Iberian Peninsula.
Site Eco-region Sp. Lat. N Long. W Alt. (m) BA (m2 ha-1)
P42201 Duriense Pp 41º 33' 43” 02º 55' 17” 1012 53.84
P42002 Duriense Pp 41º 34' 03” 02º 35' 51” 1059 34.84
P16201 Catalano-Aragonesa Pp 39º 50' 06” 01º 16' 37” 1078 45.83
P44002 Catalano Pp 40º 19' 07” 01º 21' 18” 1437 40.17
P44005 Catalano Pp 40º 20' 47” 01º 21' 54” 1364 45.73
P44204 Catalano Pp 40º 20' 01” 01º 21' 26” 1232 34.33
P16008 Litoral-Mediterranea Pp 39º 48' 56” 01º 15' 36” 920 51.44
P16106 Litoral-Mediterranea Pp 39º 50' 17” 01º 16' 11” 970 36.66
P16202 Machega Pp 39º 49' 48” 01º 17' 38” 1135 41.69
P16208 Machega Pp 39º 50' 28” 01º 17' 54” 1090 34.74
in a series of consecutive, partially overlapping segments of a length specified by the user. Dating is essential for any dendroclimatological study, and it is impossible to compare climatic variables in one specific year with tree-ring growth if the individual tree-ring series are not dated correctly.
2.2.2. Climate-growth relationship
To eliminate biological trends in tree-ring series and to minimize growth variations that are not shared by most trees (Fritts, 1976), the v6.05P ARSTAN programme (Cook and Holmes, 1984; Holmes, 2001; available at www.ltrr.arizona.edu) was used. Standardisation removes geometrical and ecological trends while preserving interannual high-frequency variations that are presumably related to climate. To obtain a master chronology at each study site, the standardized series were averaged. These temporal series or master chronologies expressed the annual variations in P. pinaster
and P. sylvestris radial growth at the population level in each sampling site.
Chronology quality was evaluated using mean sensitivity (MS), is the degree to which a tree reacts to environmental factors or the measure of the year-to-year variability (Schweingruber 1996); signal-to-noise ratio (SNR), the proportion of the variability explained by climate or other causal factors divided by the residual or unexplained variability (Fritts and Swetnam, 1989); and expressed population signal (EPS), which describes how a finite sample estimates the hypothetical infinite population (Wigley et al., 1984, Briffa, 1995). Chronology was considered confident with an EPS value higher than 0.85.
2.2.2.1. Climate-growth relationship at regional scale
The common variance between all chronologies was analysed using a Principal Component Analysis for the common growth period 1952-2005 (Sokal and Rohlf, 1995) using Infostat V.2 (Di Rienzo et al., 2002). The variance explained by the first principal component (PC1) was used as an indicator of the similarity among the chronologies. PC1 was used as a regional chronology of P. pinaster in central Spain.
To determine the climatic variables that control P. pinaster radial growth, the
average of the meteorological station data were compared with the regional chronology (PC1).
2.2.2.2. Climate-growth relationship at local scale
Bogino (2008)
12
improve the statistical significance of the correlation coefficients. In this analysis, 1000 bootstrap interactions were made.
2.2.2.3. Climate-growth relationship over time
The Kalman filter, which estimates regression models by time-varying coefficients, was applied to determine if climatic variable effect changed over time (Fritts, 1998). The output of this analysis included predicted tree-ring indexes and the confidence interval (95% level); therefore, when the interval did not include a zero value, the regression coefficient was considered as significant (Kalman filter, V5.17 PRECON programme).
2.2.2.4. Drought index effect and pulse and interpulse analysis
The drought index (DRI) of Thornthwaite (1948) was applied to detect pulse-interpulse effect on radial growth of P. sylvestris. The DRI at each sampling site was
calculated from the months that at the response function analysis showed a significant association between precipitation and radial growth (May and June at Miñon, Oña and Arauzo de Miel sites; June and July at El Espinal site and July and August at Molino Piqueras and Amogable sites). Monthly DRI was calculated by the formula,
PET
P
DRI
=
−
(2.1)where DRI is the drought index, P = monthly precipitation and PET = the potential evapotranspiration estimated from the monthly mean temperature and the geographical position of the meteorological station.
2.2.3. Intra-annual density fluctuations analysis
Once the cores were dated accurately, they were re-examined in a binocular microscope Nikon SMZ1000 (20X). Differentiation between a true tree ring and an IADF was established by the clear boundary in the true annual ring and the progressive and gradual transition in cell size and wall thickness in the IADFs (Masiokas and Villalba, 2004).
Photographs were obtained with a digital camera Leica DFC290 with a binocular microscope Leica S8APO. As the number of samples changed over time, the relative frequency was calculated with the following formula:
N
n
F
=
, (2,2)where F is the number of IADFs per year; n the number of trees that formed the IADF and N the total number of trees analysed. As the number of samples changed over time, the bias in the frequency was assessed by calculating the stabilized IADF frequency (f), according to the formula of Osborn et al. (1997), so
5 . 0
F
f
=
. (2,3)The Pearson’s correlation between stabilized IADFs was applied to detect common patterns in IADFs at different sites. ANOVA and Fisher’s test were used to detect significant differences in stabilized IADF between sites and over time. To determine the variability in IADFs over time, sampling sites of more than 100 years old were subdivided into two periods (between 1886 and 1939 and between 1940 and 2005). The Pearson’s correlation between the frequency of IADFs and the residual tree-ring series (Paper 1) was also calculated to analyse the relationship between IADFs and radial growth (Sokal and Rohlf, 1995). Infostat V.2 software (Di Rienzo et al., 2002) was used to statistical analysis.
The nonlinear logistic equation form was chosen to model the probability of occurrence of IADFs in P. pinaster rings,
(
)
10
.
1
+
− −=
ze
P
(2,4)where P is the probability of IADFs and Z = b0+b1(x1)+b2(x2)+…+bk(xk) + ε; where x1; x2... xk are the climatic variables and b0; b1; b2 ….. bk are unknown parameters of the model, ε is a normal random error N(0,1), and e is the exponential operator.
Bogino (2008)
14
was used to find the best model. The alternative fits were evaluated on the basis of Akaike information criterion (AIC), the area under the receiver operating characteristic (ROC) curve and the expected behavior - as indicated by the signs of the parameters estimates. The area under the ROC curve can be considered as an estimator of accuracy. This curve, which is widely used in health sciences but not in Dendrochronology, relies on false/true-positive/negative tests, and the sensitivity is indicated by the proportion of correctly classified events and the specificity by the proportion of correctly classified non-events (Hair et al. 1998). PROC LOGISTIC of SAS 9.1 (SAS Institute Inc., 2004) was used to fit the model.
2.2.4. Isotope analysis
The δ13C isotope was determined on whole wood (Schleser et al., 1999a; Babour et al., 2001). Cellulose was not extracted from the wood as these two components were shown to yield highly correlated signals (Borella and Leuenberger, 1998) and even higher climatic signal can be detected in the untreated material (Loader et al., 2003). Powdered material was obtained for each tree ring with a Micromot 40E instrument with a 0.5 mm thick needle. The samples were obteined after identification of the previously dated tree ring by use of a binocular microscope. Each sample was analysed individually, the material was weighted, and for each tree ring, 0.2-0.3 mg of powered material was placed into tin capsules. The stable carbon isotope was measured with an NA 2500 elemental analyser (CE Instruments, Rodano, Italy), with an isotope ratio mass spectrometer (Finningan MAT Delta plus, Bremen, Germany).
The isotopic composition (δ13C) of samples was determined with the formula,
(
)
(
)
1 312 13 12 13 13
10
/
/
)
‰
(
=
×
− PDB sampleC
C
C
C
C
δ
(2,5)where δ13C (‰) is the proportional deviation from the international Peedee belemite (PDB) carbonate standard (Craig, 1957)
As a previous study showed a changing association between climatic variables and growth of P. pinaster (Paper 1) from the 1980s onwards, and phenological changes
have been reported over the last 25 years (Peñuelas et al., 2002), the period analysed was between 1975 and 1999. The same period was considered for P. sylvestris.
climatic variables that were best correlated with δ13C (***p<0.001) were grouped to
construct simple linear regression models, as follows (Sokal and Rohlf, 1995):
ε
+
+
=
a
bx
y
, (2,6)where y is δ13C; x is the sum of climatic variables; a and bare unknown parameters
of the model and ε is a normal random error N(0,1).
Statistic analysis was carried out with the Infostat programme (Di Rienzo et al., 2002).
2.3.CLIMATIC DATA
Monthly precipitation and mean monthly temperature by the Agencia Estatal de Meteorologia, Spain, were used to detect climate-growth relationship. The
meteorological data provided belong to meteorological stations placed within 30 km of the sampling sites (Table 3). The data recorded cover a period of, at least, 50 years. The data from four meteorological stations were averaged to obtain a regional climatic record to be applied in the regional dendroclimatic analysis of P. pinaster.
Table 3. Meteorological station data from the Agencia Estatal de Meteorología, Spain
Location Lat. (N)
Long. (W)
Alt. (m)
Rain. (mm)
Temp. (ºC)
Period
El burgo de Osma 41º 35' 10" 03º 04' 02" 895 529.5 10.54 1932-2005 Cuenca 40º 04' 00" 02º 08' 17" 956 541.42 12.43 1956-2005 Pantano de la Toba 40º 13' 19" 01º 55' 33" 1154 764.68 10.22 1944-2005 Observatoriode Soria 41º 46' 00" 02º 28' 00" 1082 418.34 10.66 1944-2005 Miranda del Ebro 42º 40' 42" 02º 57' 20" 520 529.97 12.08 1936-2005 Villafría 42º 21' 22" 03º 37' 57" 890 564.67 10.15 1943-2005 Aldea del rey Niño 41º 34' 35" 04º 42' 02'' 1160 522.24 9.17 1935-2005 Yemeda 39º 45' 40'' 01º 43' 17'' 868 405.13 12.41 1950-2000 Cella 40º 27' 20'' 01º 17' 27'' 1023 370.81 12.01 1939-2006
The HOM component (Homogeneity of Meteorological Data) of the Directory Programme Library for Dendrochronology (Holmes, 1983; available at www.ltrr.arizona.edu) was used to determine the homogeneity of the climatic variables.
3. RESULTS
3.1.CHRONOLOGY DESCRIPTION
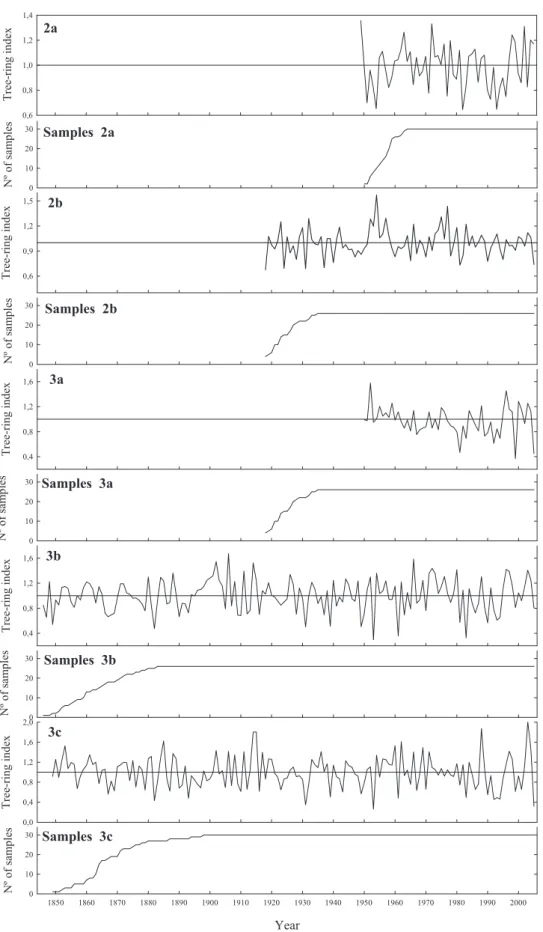
One chronology of Pinus pinaster was not included in the analysis because its time
span was only 30 years and it would have limited the period analyzed of all series. The nine and six chronologies of P. pinaster and Pinus sylvestris, respectively had
high SNR, EPS and percentage of the variance accounted for the first eigenvector; this suggests that they reflect a strong common signal, presumably related to climatic factors. The descriptive statistics showed that mean sensitivity varied from 0.1857 to 0.3179 and from 0.1556 to 0.2669, standard deviation varied from 0.1650 to 0.3108 and from 0.1858 to 0.3527, for P. pinaster and P. sylvestris, respectively. SNR varied
from 27.615 to 68.444 and from 10.676 to 24.747 and EPS values varied from 0.958 to 0.986 and from 0.914 to 0.961, for P. pinaster and P. sylvestris, respectively. All
chronologies analysed have high SNR (values always over 10) and EPS (always over 0.914), while the variance accounted by the first eigenvector is over 35 %; these facts suggest a strong common signal related to environmental climatic factors (Tables 3, I and III).
In P. pinaster, despite the diversity of tree ages and the difference in bioecological
regions, a clear common macroclimatic signal expressed by the first principal component (PC1) of the PCA was found. PC1 explained 56% of the total variance of indexed tree-ring growth among the nine chronologies. All the chronologies had a positive correlation with PC1, indicating they shared a common variance (Figure 4 and Table 4, I).
3.2.CLIMATE GROWTH ASSOCIATION: MULTIPLE STEPWISE AND BOOTSTRAPPED
ANALYSIS
Radial growth, at both regional and local scales, in P. pinaster was strongly
associated with climatic variables, especially rainfall during and prior to the growing season in all the sites analysed. Mean temperature effect changed according to the sampling site, from insignificant at the highest altitudinal sites to positively significant in winter at the lowest altitudinal position sites (Figure 6, I).
As a result of the PCA analysis, PC1 was considered indicative of the interannual high-frequency variation of tree-ring growth at regional level for the common period 1952-2005. This chronology was correlated with regional climatic variables, precipitation and temperature, and the results showed that more than 60.8% of the total variance in radial growth was explained by climate (Figure 5, I).
3.3.CLIMATE GROWTH ASSOCIATION OVER TIME: THE KALMAN FILTER TECHNIQUE
IN P. PINASTER
Bogino (2008)
18
The change in association between growth and climatic conditions took place during the 1980s. During this period, precipitation changed from insignificant to positively significant at four sampling sites. This change occurred in winter prior to the growing season or at the beginning of the growing season. Temperature also showed a changing association with growth at two sampling sites. Temperature became negatively significant on radial growth in springtime (May) at one sampling site; this change in association may be related to the intense drought and the warming that characterised the 1980s in the peninsula. Finally a positive change was found in the other sampling site in relation to winter temperature (February) (Table 5, I).
3.4.NAO AND SOI EFFECT ON RADIAL GROWTH
The total variance explained by atmospheric indexes varied from 8.95 to 37.46%. The total variance explained by the NAO and SOI indexes is higher in the sites at higher positions. In these places there is a significant negative association between the NAO index and growth during January and March or December and March, but only March is significant in the bootstrapped analysis. Only one place showed a positive association with NAO values in September prior to the growing season, but this association was not significant in the bootstrapped analysis. The association with the SOI was positive in all the analysed sites, but it was not significant, in the correlation coefficient and the in the bootstrapped response function. Only one site showed a negative association with the SOI, shown by the bootstrap coefficient during March previous to the growing season (Figure 2, II).
3.5.CLIMATE AND RADIAL GROWTH OF P. SYLVESTRIS
Tree-ring width was positively correlated with rainfall in the growing season at all sites. Mean temperature effect varied according to site. A climatic response according to age was also found: young stands had higher variability explained by climatic variables than old stands (Figure 3, III).
Because the six sites analysed showed a clear association with rainfall in the growing season, the months where this association was significant at each sampling site were selected for DRI calculation. The Pearson’s correlation coefficient between tree-ring index and DRI showed a significant association in all sampling sites (* p < 0.05; **p < 0.01;***p < 0.001) (Table 4 and Figure 4, III).
The ACF of the six chronologies performed to detect autocorrelation between successive tree-ring growth values suggested that autocorrelation coefficients in young stands (less than 100 years old) were not significant. However, this association changed in old stands (more than 100 year old), where a significant association between current year tree growth with that of one, two or three years before was detected (Figure 5, III).
showed by a periodicity of two or two and a half years, except for only one sampling site (Table 5 and Figure 6, III).
3.6.IADFS AND CLIMATE RESULTS
P. pinaster is characterised by different anatomical features. A total of 11 930 tree
rings were analysed and a total of 1 038 IADFs were detected in the trees, 8.7% of the tree ring analysed (Table 3, IV). Even though the species showed different anatomical IADFs they were rather easy to identify by correct preparation of the samples. Samples showed an increase in IADFs from the 1940s to the present, except in only one site (Table 6, IV). The absence of IADFs at this site may be attributed to the extreme thinness of the tree rings (less than 0.3mm), which made it impossible to detect IADFs (Figure 4, IV). There were IADFs at all sampling sites in 1961 and 1983, with a stabilized frequency higher than 3. A low frequency of IADFs was recorded in the 1970s at all sampling sites (Figure 3, IV).
The Pearson’s correlation between stabilized IADFs was positive and significant for all comparisons between sampling sites (*p<0.05 and ***p<0.001) (Table 4, IV).
The results of the ANOVA and Fisher’s test performed to detect significant differences between IADFs at different sampling sites (from the common growth period 1953-2005) showed that younger sampling sites have higher IADFs than older sampling sites. However, Fisher’s test does not enable complete separation of young and old stands. The higher mean IADFs also coincided with the highest site index, but as these sites are also the youngest in terms of cambial age, it was not possible to ensure that the higher IADFs are related to site index (Table 5, IV).
Significant differences were found in the last 120 years in the stabilized IADFs. The Pearson’s correlation coefficients showed a negative significant correlation (*p<0.05; *** p<0.001) between tree-ring growth and stabilised IADF at all sampling
sites. The results suggest that IADFs restrict growth at all sampling sites (Table 7, IV). The logistic function used to predict the possibility of the occurrence of IADF estimated that 18 monthly climatic variables out of 24 had a significant effect on predicting future IADFs. Drought events in May and July had a positive impact on IADF while wet periods in April and June also promoted IADF. Such pulses in precipitation (rainy months follow by dry months) are typical of Mediterranean areas. Increases in temperatures also had a generally positive impact on IADFs (Table 8, IV). The value of the area under ROC curve (0.84) shows that the accuracy of model is good enough to use it to predict occurrence of IADFs (Figure 5, IV).
3.7.CARBON ISOTOPES AND CLIMATE RESULTS
Bogino (2008)
20
A negative significant correlation between δ13C and radial growth was found for the 1975-1999 period. The Pearson’s correlation coefficient for the residual tree-ring chronologies and the mean δ13C was r: -0.83 (***p < 0.001) for P. pinaster and r:-0.41
(*p < 0.05) for P. sylvestris.
Pearson’s correlation coefficient for climatic variables and δ13C in P. pinaster
showed that moisture is a driving factor affecting the variability of δ13C between winter and summer (January to July). Rainfall also had a negative effect on δ13C between winter and spring although the effect was only statistically significant in April and May. Monthly mean maximum temperature in March had a positive significant effect on δ13C and monthly hours of solar radiation in January, April and May (Figure 4, V). Pearson’s correlation coefficient for climatic variables and δ13C of P. sylvestris
showed that moisture is a driving factor affecting δ13C variability in summer (July) and in autumn (October). Rainfall in October also had a negative effect on δ13C. Monthly mean maximum temperature in summer (June and July) had a positive significant effect on δ13C as well as hours of solar radiation in October (Figure 5, V).
For P. pinaster RH from January to July, rainfall from January to May, maximum
mean temperature from April to July and hours of solar radiation of January, April and May were selected to construct four different simple linear regression models that relate each variable and δ13C (Figure 6, V). For P. sylvestris RH in July and October
4. DISCUSSION
Pinus pinaster is a reliable species for dendrochronological studies, showing good
correlation between trees growing at the same sampling site, high signal related to total noise and accurate statistical values that mean clear response to environmental factors.
P. pinaster also shows accurate performance in studying the association between tree
growth and global change showed by a changing relationship with climatic variables over time.
A common growth pattern among all series has been detected. PCA analysis suggests a clear strong common variance among all the sampling sites (explaining over 56% of variability) and a positive correlation (**p<0.01 and ***p< 0.001) with the
PC1 axis. Although the sampling sites included trees of different ages and trees from different eco-regions, all the series can be considered to share a common variance related to causal factors.
The total variance explained by average climatic variables and regional radial growth index of P. pinaster (60%) can be considered higher than the most common
findings obtained in dendroclimatic studies worldwide. Tree rings rarely cover more than 60% of the variance registered in instrumental records, and 40 to 50% is quite a common level (Fritts, 1991; Jones et al., 1998). Richter et al. (1991) found 68% of the total variance explained by precipitation and temperature in pine species growing in the Iberian Peninsula.
Rainfall is the dominant climatic variable that has a significant association with
P. pinaster growth, as the stepwise correlation analysis indicated. Rainfall effect
constitutes the significant climatic variable in both, the regional and local analysis. Temperature showed a changing significant effect on tree growth that varied according to the sampling site. No effect was recorded at the sites placed at the highest positions and a negative effect was recorded at five sampling sites. The difference found out in this study is that temperature effect could not be simplified to a common response in all the sampling sites because local issues determine the association between this variable and radial growth.
Although P. pinaster shows a strong association with precipitation and the
temperature impact is lower than that of rainfall, its increase might raise evaporation, limit environmental moisture and, consequently, restrict growth. However, the predicted temperature trend until 2080 will probably not lead to an unsuitable environment for P. pinaster (Harrison et al., 2006). As P. pinaster is a
pre-Mediterranean species that has suffered different environmental changes [from subtropical environments to Mediterranean ones (DiCarlo, 1931)], it can also deal with very atypical severe droughts, such as that of 1994 (Peñuelas et al., 2002), and it shows a better water-use efficiency than P. sylvestris (Martínez-Vilalta and Piñol, 2002).
These facts indicate that P. pinaster might be well adapted to summer rainfall
reductions, as IPCC forecasts in the Iberian Peninsula.
Bogino (2008)
22
The climatic warming of the 1980-1995 period was characterised by intense droughts that produced severe damage in woody species (Peñuelas et al., 2001). Our findings indicate that some climatic variables (May temperature and April-May rainfall) have changed from insignificant to significant in the last 30 years (from 1980 until now). This coincides with previously-reported phenological changes in plant life (related to temperature) in the Mediterranean region. In addition, these changes started during the 1980s, which could mean a changing association with climatic factors (Peñuelas et al., 2001; 2002).
It is difficult to find a simple linear correlation between radial growth of
P. pinaster and atmospheric indexes because their global effects and their impact on
regional climatic variables are not yet completely understood.
In these results, the total variance explained by NAO and SOI indexes suggested that the signal is weak if it is compared with regional climatic variables. However, the negative correlation with NAO in winter in two sites, and the changing effect of SOI index over time in one site, offer new information about the association between atmospheric indexes and coniferous species growing in the Iberian Peninsula.
Although atmospheric indexes explain less variability than other regional climatic variables, these results emphasised that these indexes effects could be recorded on tree ring and they could have a sensible effect in growth of woody species, even if their action centres are located too far away from the analysed sites.
Pinus sylvestris is a useful species in dendrochronological studies showing
accurate statistical values that mean clear response to environmental causal factors. The six chronologies have high mean sensitivity (MS), expressed population signal (EPS), signal to noise ration (SNR) and percentage of the variance accounted for the first eigenvector, suggesting a strong common signal to related-climatic environmental factors.
The association between growth and climate shows that rainfall in the growing season has a positive significant effect on radial growth among all sites, no matter what their altitudinal location, geographical position or cambial age are. Differences in rainfall effect within the growing season were found: at sites located at the lowest altitudinal position, this effect happened in spring (May and June); in contrast, with sites placed at a higher altitudinal position, the association occurred in late spring and summer. This different association may be due to temperature. Sites located at the lowest altitudinal positions are warmer; consequently, the growing season starts early, and the spring rainfall is a driving factor that affects growth. However, sites located at the highest altitudinal position are colder, the growing season starts later and growth depends on summer rainfall. These results emphasised how important the within-season precipitation dynamic is; it can be equal to or more important than the within-seasonal or annual total for plant growth (Fay et al., 2000; Knapp et al., 2002).
The significant association between radial growth and DRI suggests that drought is a key factor affecting P. sylvestris growth in its southern and western distribution area.
ring growth is affected by previous growth. Young stands have higher variability explained by climatic variables in the response function analysis (from 53 to 69%) and are not vulnerable to previous growth; in contrast, old stands have a lower percentage of the variability explained by climatic variables (from 31 to 39%) but are vulnerable to previous growth. Since ecosystems have the capacity to store water, they clearly have a “memory” of past precipitation events. As a result of such a “memory” effect, it is not correct to understand rainfall effect as a single seasonal or annual event (Schwinning et al., 2004). This study confirms the “memory” of past events when autocorrelation analysis is performed, suggesting an association between present growth with previous growth years in old P. sylvestris stands. When growth periodicity
is analysed to determine pulse and interpulse periods in P. sylvestris growth, it is clear
that the strong DRI variability and the significant effect that this index has on tree-ring growth result in a strong variability between successive years that is reflected as periods of two or two and a half years in the time series analysis. Even though, depending on the sampling site, other cycle periods were detected, two-year periodicity is stronger than other long-period cycles.
P. pinaster showed nine different anatomical structures that confirm the tendency
of Mediterranean species, and this species in particular, to develop special anatomical structures (Schweingruber, 1993). Even though IADFs, which occur in all series at all sampling sites, may previously have limited dendroclimatological studies in
P. pinaster (Schweingruber, 1993), when the correct date is obtained, the significant
association between IADFs and radial growth or climatic variables provides a useful proxy for complementing and enhancing dendroclimatological data (Paper 1).
Latewood is formed from carbohydrates produced during photosynthesis which is the result of water stress and temperature (Kozlowski et al., 1991). IADFs of
P. pinaster growing in Tuscani Italy had a higher 13C/12C isotope ratio in latewood
than earlywood, which suggests a better water use efficiency (De Micco et al., 2007). The present results emphasised the impact of drought events on IADFs (significant effect of rainfall in the growing season) and suggest a physiological response of P. pinaster to unfavourable climatic conditions and the development of anatomical
structures that lead to better water use efficiency. Isotope analysis in the Iberian Peninsula, showed that P. pinaster makes more efficient use of water under severe
drought events than P. sylvestris and Pinus nigra Arnold (Martinez Vilalta and
Piñol, 2002).
Previous studies that relate IADFs to growth of P. sylvestris in Switzerland
determined a positive association between tree-ring growth and IADFs (Rigling et al., 2001); these results contrast with the present results in which a significant negative relationship between radial growth and IADFs was found, suggesting that the environmental variables that produce IADFs also limit growth.
The effect of climate on IADFs of P. pinaster is determined by both the significant