Casa Abierta al Tiempo
Universidad Autónoma Metropolitana
UNIDAD IZTAPALAPA
UNIVERSIDAD AUTONOMA METROPOLITANA
UNIDAD IZTAPALAPADIVISION DE CIENCIAS BIOLOGICAS Y DE LA SALUD DEPARTAMENTO DE CIENCIAS DE LA SALUD
,y Título del trabajo:
1
“CUANTIFICACION DEL DAÑO AL DNA CAUSADO POR /ESTRÉS OXIDATIVO Y EL POSIBLE EFECTO
PROTECTOR DE B c u
/ Tesis que presenta la alumna: O h i a Huchín Espinosa de
los
MonterosMatrícula: 923307 13
Para la Obtención del Grado de: Licenciada en Biología Experimental.
Asesora: M. En Biol. Exp. Mina Konigsberg Fainstein.
Agosto/ 02.
ANTECEDENTES:
1 . I ENVEJECIMIENTO Y ESTRÉS OXIDATIVO
El envejecimiento es un fenómeno que se caracteriza por el deterioro de las funciones
celulares. Sin embargo, las bases moleculares del envejecimiento aún no están
totalmente comprendidas (Harman, 1956).
En la actualidad no existe una teoría única que pueda explicar el fenómeno. Sin
embargo, una de las hipótesis más aceptada, es la que relaciona la mitocondria y el
estrés oxidativo con el envejecimiento (Lenaz, 1998). El concepto de que la
mitocondria está íntimamente involucrada en el envejecimiento deriva de la teoría de
Harman (Harman, 1956), que propone que el daño producido por los radicales libres o
especies reactivas de oxígeno (ROS), inducen los cambios que producen el
envejecimiento. Todos los seres vivos están expuestos a ROS exógenos entre los que
se encuentran los rayos UV, la nicotina del tabaco, la contaminación y ROS
endógenos como subproductos de la cadena respiratoria y otros proceso metabólicos.
Puesto que se calcula que entre el 1 y el 4 YO de oxígeno que utiliza la cadena
respiratoria mitocondrial, es reducido en forma incompleta hacia ROS (Dimitry, 1997);
se ha observado que específicamente en el complejo Ill (Ubiquinol citocromo c
reductasa) la salida de electrones produce al radical superóxido ( 0 2 ), que a su vez
genera al peróxido de hidrógeno H 2 0 2 ,que no es propiamente un radical libre pero
que posteriormente produce al extremadamente agresivo radical hidroxilo OH
(Halliwell, 1984; Christoph, 1988). Sin embargo, para contrarrestar el efecto de estas
dismutasa, el glutatión, y las vitaminas C y E, que funcionan como protectores contra
los radicales libres de las ROS (Lenaz, 1998).
La mitocondria al ser la mayor fuente de producción de las ROS, está constantemente
expuesta a ellos y puede ser suceptible a un ataque de los mismos (Christoph, 1988) .
Existen evidencias de que las ROS pueden dañar a los componentes de la
mitocondria disminuyendo la actividad de las enzimas y el potencial de la membrana
mitocondrial. El daño por estrés oxidativo en los componentes de la mitocondria en
particular y la célula en general incluye peroxidación de lípidos, oxidación de proteínas
y mutaciones en el DNA (Halliwell, 1984).
El daño al DNA mitocondrial tiene especial interés, ya que al no tener histonas de
protección ni enzimas de reparación, es altamente suceptible al daño por estrés
oxidativo (Christoph, 1988) . La acumulación de mutaciones en el DNA mitocondrial
inducidas por una exposición continua al ataque de radicales libres, codificará errores
en las cadenas del DNA mitocondrial, que se traducirán como errores en la síntesis de
proteínas y en una cadena de respiración defectuosa (Lenaz, 1998 ; Yau-Hei, 1998),
incluso se han reportado una inducción extensiva.de fragmentaciones y deleciones
( más de 187 diferentes deleciones se presentan en una lista mostrada por Ozawa)
(Pang, 1994) después de 3 días de exponer a estrés oxidativo a una línea celular
humana transformada de fibroblastos. Así mismo se han observado una serie de
mutaciones y deleciones en el DNA mitocondrial durante el envejecimiento y
enfermedades asociadas al mismo (Lenaz, 1998 ; Yau-He¡, 1998).
En el análisis realizado en el DNA mitocondrial de personas de edad avanzada, es
común”. Esta deleción es un fragmento que tiene una longitud de 4977 pares de bases
y se localiza entre
los
8483 y 13459 pares de bases (Yau-Hei, 1998).FACTOR DE NECROSIS TUMORAL a (TNF-a)
El TNF-a es una citocina llamada “Factor de necrosis tumoral ” ; estructuralmente es
una proteína de 17 kDa producida por macrofagos y otros tipos celulares. TNF-a fue
identificado por su potencial citotóxico y se ha relacionado como un mediador letal
agudo de infecciones crónicas(Tracey, 1995).
El gen de TNF-a humano, esta localizado en el cromosoma 6 cerca del locus HLA-B
codificando la prohormona del aminoácido 233 (Wang et al., 1985; Spies et al., 1986;
Davies et al. , 1987; Muller et al., 1987), ésta puede servir para promover el precursor
de la membrana plasmatica, sin embargo el secretor de TNF-a maduro se encuentra
en la en la superficie de la membrana extracelular (Kriegler et al., 1988; Jue et al.,
1990). AI exponer a la membrana a TNF-a extracelular se encontró que había
disminución de la bioactividad como en las células expuestas a citotoxicos (Kriegler et
al., 1988 ; Jue et al., 1990 ; Perez et al., 1990).
Los estudio cristalograficos de la estructura de TNF revelaron una similitud con una
“campana” compuesta por tres monomeros asociados no covalentemente dentro un
triímero que forma un eje de tres pliegues de simetría (Jones et al., 1989). Con esta
estructura, cada monomero de 157 aminoacidos esta atrás del pliegue sobre sí mismo,
como un P-plegamiento ; las interacciones entre los tres pliegues adyacentes
estabilizan la estructura del triímero (Sprang and Eeck, 1992). El amino terminal de
cada monomero se encuentra expuesto en la superficie y el carboxilo terminal se
estructural sugieren que las mutaciones que desestabilizan la asociación trimérica del
monómero provocan la pérdida de la actividad biológica (Lin, 1992).
La biosíntesis de TNF-a esta en gran parte regulada por eventos moleculares que han
sido cuidadosamente estudiados en el monocitolmacrófago.
Clones de cDNAs que codifican para dos receptores de TNF-a, han facilitado el
aislamiento y la secuenciación del fragmento de péptido que procesa la actividad
inhibitoria de TNF-a (Gray et a1.,1990 ; Loetscher et al., 1990 ; Nophar et al., 1990). Se
ha probado que éstos péptidos llamados “Proteinas ligadoras de TNF-a” (TNF-a BPI y
TNF-a BP2) provocan degradación proteolítica en los receptores de TNF-a ; y estos
receptores a su vez, son componentes de una región intracelular del segmento
transmembranal y un dominio extracelular correspondiente a los TNF-a BPS (Howard
et al., 1991 )
Bcl-2
Bcl-2, una proteína relacionada con la apoptosis y se ha reportado que además
prolonga la viabilidad celular en presencia de estrés oxidativo (David, 1993). Esta
proteína se localiza en la membrana interna mitocondrial, membrana nuclear y retículo
endoplásmico (Juliane, 1998) y forma parte de una gran familia de moléculas
antiapoptóticas, sin embargo, el mecanismo por el cual aumenta la longevidad en
JUSTIFICACION :
Desde tiempos remotos, los hombres han tratado de encontrar alguna manera de
impedir el deterioro progresivo al que todos estamos condenados : el envejecimiento.
Hoy en día se conocen algunas enfermedades degenerativas relacionadas con el
envejecimiento, las más comunes y devastadoras han sido relacionadas con el
metabolismo mitocondrial. Ciertas enfermedades como Huntington, Parkinson y la
degeneración cerebral presentan una patología con un común denominador que afecta
a diferentes áreas específicas con atrofia celular que pueden ser debidas a un
metabolismo oxidativo reducido y una baja síntesis de ATP (Lenaz, 1998)(Yau-Hei
1 998).
Como se mencionó anteriormente, la molécula de Bcl-2 prolonga la viabilidad celular
frente a un reto de estrés oxidativo, sin embargo se desconoce si Bcl-2 tiene la
capacidad de proteger al DNA celular de daño durante dicho estrés.
Por lo tanto el objetivo de este trabajo será evaluar la capacidad de Bcl-2 para prevenir
el daño al DNA generado por estrés oxidativo, ya sea con H 2 0 2 o con TNF.
Para ello se utilizará una línea celular de fibroblastos de pulmón de ratón (L929) que
sobreexpresa a Bcl-2; se le someterá a distintos tratamientos de estrés oxidativo y se
evaluará el daño y protección sobre el DNA en las células silvestres y en las que
sobreexpresan Bcl-2.
Este estudio contribuirá a entender el mecanismo de acción de Bcl-2, así como al
fenómeno del envejecimiento celular y su relación con el fenómeno de apoptosis y
OBJETIVO GENERAL :
Evaluar el daño generado por estrés oxidativo sobre las células L929 que
sobreexpresa Bcl-2 y las que no
lo
hacen.OBJETIVOS PARTICULARES :
1 .I Encontrar las condiciones óptimas para evaluar el daño en el DNA de las células
tratadas con H202 (concentración y tiempo de exposición).
1.2 Encontrar las condiciones óptimas para evaluar el daño en el DNA de las células
L929 tratadas con TNF (concentración y tiempo de exposición).
1.3 Someter a las células P6 (silvestres) y C.2 (que sobreexpresan Bcl-2) a tratamiento
con la concentración y tiempo de H202 y TNF establecidos previamente.
METODOLOGIA :
Mantenimiento del cultivo celular.
La línea celular que sobreexpresa Bcl-2 (C.2) se mantuvo en medio mínimo esencial
(MEM) suplementado con 10% de suero fetal bovino y 2pg/mI de puromicina, mientras
que la línea celular que no sobreexpresa Bcl-2 (P6) se mantuvo en MEM
suplementado con 10% de suero de ternera. El medio se reemplazó cada 3 días. Las
células se crecieron a 37"C, en botellas de plástico desechables (Nunc, Kamstrup,
Denmark) en una atmósfera de 95% de aire y 5% de COZ.
Establecimiento de las condiciones óptimas.
Nuestro grupo de investigación realizó trabajos previos para evaluar el efecto protector
de la sobreexpresión de Bcl-2 en tratamientos crónicos de 1 a 6 días con
concentraciones crecientes desde O hasta 10 mM estableciendo que la concentración
óptima era de 0.05 mM por 3 días. Para tratamientos agudos la concentración óptima
fue de 2 mM por 4 horas. Sin embargo al tratar de utilizar estas concentraciones a
dichos tiempos, fue imposible recuperar las células al momento de despegarlas.
Posiblemente esto se deba al daño que ya presentaban las células. De manera que se
decidió utilizar las mismas concentraciones (0.2mM y 0.05mM) de H202 a tiempos de
10, 20 y 30 minutos, y se les llamó tratarnientos de alta y baja concentración.
Tratamiento con TNF : Para el TNF se realizaron experimentos a los 30 minutos,
1,3,6,9,12 y 16 horas. Con una concentración de 5qg/ml. Dicha concentración se
estableció previamente en el laboratorio del Dr. Zentella del IFC, UNAM como la
Tratamiento comparativo de las diferentes líneas celulares
Para el tratamiento de baja concentración se adicionó 0.05mM de H 2 0 2 a las células
P6, C.2 y P R I durante 30 minutos; El tratamiento de alta concentración se realizó
adicionando 2 mM de H202 a las células P6, C.2 y PRI durante 30 minutos. Para el
tratamiento con TNF, se agregó 5qg/ml de la citocina a las 3 y 6 horas.
Cuantificación del daño al DNA
Se utilizo la técnica del ensayo cometa o electroforésis celular alcalina, para
observar el rompimiento en las cadenas sencillas de DNA y se cuantificó en un
analizador de imágenes marca Sinoptics (Collins, 1997).
Ensayo Cometa :
Preparación de las muestras
1. Se sembraron las células de tal manera que el día del ensayo se tuvieron 100,000
células /cm2
2. Se tripsinizaron y se transfirieron a tubos Ependorff
3. Se centrifugaron a 4,000 rpm por 4 min
4. Se tiro
el
sobrenadante y el botón se resuspendió en 40pl de PBS5. Se tomaron
lop1
del homogenado celular por preparación y se depositaron en otrotubo Ependorff conteniendo 75111 por preparación de agarosa de bajo punto de
fusión (LMP) al 0.5% tibia
Preparación de los minigeles
1. Se depositó una gota de 1 2 0 ~ ~ 1 de agarosa regular al 1 Ohsobre un cubreobjetos y se
sin burbujas. La agarosa debe estar tibia y el cubreobjetos sobre una parrilla
caliente para que no gelifique antes de tiempo.
2. Se puso el portaobjetos en hielo hasta que gelifico
3. Con mucho cuidado se deslizó el cubreobjetos y sobre el lado que no se usó se
agrego una gota de 80pl de la mezcla del homogenado celular que se encuentra en
agarosa LMP. Se cubrió con el portaobjetos cuidando de no hacer burbujas.
4. Se puso en hielo hasta que gelificó
5. Con mucho cuidado se deslizó el cubreobjetos y se deshecho.
6. En un nuevo cubreobjetos se agrego una gota de 85pl de agarosa LMP 0.5% y se
tapó con el portaobjetos que contiene las 2 capas anteriores.
7. Se puso en hielo hasta que gelificó
8. Se quitó el cubreobjetos y se depositó el portaobjetos con las 3 capas de agarosa
en la solución de lisis fría.
9. Se dejaron las muestras en esta solución por 24h (pueden estar hasta 7 días) en
refrigeración.
Electroforesis
1. Se sacaron los geles de la solución de lisis y se enjuagaron con el amortiguador de
corrida
2. Se acomodaron en la cámara de electroforesis, se cubrieron con la solución de
corrida y se dejaron estabilizar por 20 minutos
3. Pasado el tiempo de equilibrio se corrieron los geles por 20 minutos a un voltaje de
RESULTADOS :
I. Tratamientos de alta y baja concentración con peróxido de hidrógeno
En la gráfica 1 .I ,se muestran las diferencias de migración del cometa entre las células
P6 y C.2 que han sido tratadas con H202 a baja concentración ( 0.05 mM ) a diferentes
tiempos. Aquí se pueden ver los distintos grados de daño temprano al DNA que
provoca el H202 en las condiciones establecidas; cabe notar que en el tiempo cero
(células control), también se nota daño, y esto es debido a que se están cuantificando
eventos únicos; a los 10 minutos, en las células P6 se observa una menos migración
con respecto a las células C.2 de 10 minutos, en éstas últimas, se observan
migraciones desde 1 hasta 60 micómetros, en cambio, en las P6
solo
migraciones de 1hasta 40 micrómetros. A los 20 minutos si se puede observar una pequeña protección
en las C.2, ya que su migracón va desde 1 hasta 40 micrómetros, y en las P6
migraciones desde 1 hasta 60 micrómetros, y a los 30 minutos, en las C.2, se observan
daños desde 1 hasta 80 micrómetros, pero en las P6, se observa daño desde 1 hasta
40 y de 40 hasta 1 OO.
En la gráfica 1.2 se muestran las diferencias de migración del cometa entre las células
P6 y C.2 que han sido tratadas con H202 a alta concentración ( 0.2 mM ) a diferentes
tiempos. En este caso se observa de manera muy evidente como aumenta el daño, es
decir la migración del cometa, con respecto al tiempo. En especial en las células P6.
La figura 1.3 es una comparación de los tratamientos anteriores en las dos líneas
en ambos tratamientos hay una diferencia significhtiva que no existe a los 10 y 20
minutos.
2. Tratamiento con TNF
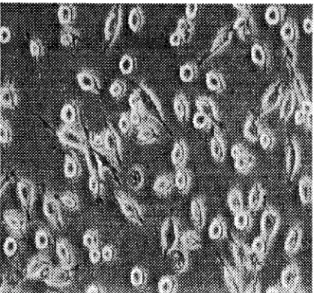
Las células P6 se trataron con Snglml de TNF durante los tiempos antes mencionados
y como se observa en la figura 2.1 existe un daño al DNA que va aumentando
conforme aumenta el tiempo. Si se comparan estos resultados por los mostrados en
las fotografías (figura 2.2), se observa que aún y cuando las células no mostraban
daño aparente, ya presentaban daño temprano en el DNA.
3.
Tratamiento comparativo de las diferentes líneas celularesPara evaluar el efecto protector de Bcl-2 de una manera más evidente, se realizó un
experimento comparativo y simultáneo entre las célluas P6 y c.2, junto con otra línea
celular (PRI) a la cual se le insertó el plásmido de pBabePuro sin Bcl-2, pero que se
sabe que debido a esa transfección, son resistentes a TNF.
En la figura 3.1 se observan los resultados de la migración de los cometas a los
diferentes tratamientos con peróxido y TNF. Es importante notar que el
comportamiento de las células P6 fue como lo que se esperaba, mientras que en las
células c.2 si se encontró un efecto protector. Sin embargo en las células P R I se
1
P6 expuestas a 0.05mM [H202]
migración del cometa (um)
c.2 e n presencia de 0.05mM [H202]
cn
O
C
al
>
al al
U
z
c )
o
1
Migracióndel cometa (urn)
P6 expuestas a 0.2mM [H202]
40
20
tJ 10
longitud del cometa
tiempos, O, 10 20, y 30 m i n
longitud del cometa
0-20, 2 1-40, 4 1-60,
61-80, 81-100
c.2 expuestas a 0.2mm [H202]
40
/ I
35
1
I U
migración del cometa (urn)
P6 y c.2 expuestas a 0.05mM [H202]
70
60 E
4 50
40
-
Sc
9)
-
30
.O
.- 20
10 0
O
O 1 O' 20' 30'
tiempo
P6 y c.2 expuestas a 0.2mM [H202]
80
,
IO 1 O' 20' 30'
tiempo
azul = P6, rojo= c.2
P6 expuestas a Bnglml de TNF
30
16
6 tiempode exposici6n ( h )
longitud del cometa (um)
P6 expuestas a 5nglml de TNF
" ~ . . .. - . . ." .. ~. . . ~ ~~ ~ ~~
O 0.5 1 3 6 9
tiempo de exposición (h)
Células L929 control
1
oox
Células L929 expuestas a
Células L929 expuestas a
5
ng/mL de TNF por 6h
Figura 2.2b
Células L929 expuestas
a
migración
del cometa
(Urn)
140
120
I
O0
80
60
40
20
O
control
H202
TNF TNF
3h
6h
Q
p6
c.
2
o
PRI
J
DISCUSION Y CONCLUSION:
La molécula de Bcl-2 es una proteína que se ha observado que prolonga la viabilidad
celular frente a un reto de estrés oxidativo, sin embargo se desconoce si Bcl-2 tiene la
capacidad de proteger al DNA celular de daño durante dicho estrés. Es por ello que en
este trabajo se evaluó el daño temprano producido por el peróxido de hidrógeno sobre
las células silvestres y las que sobreexpresan Bcl-2. Con respecto a los resultados con
los tratamientos de peróxido se encontró que las células c.2 (que sobreexpresan Bcl-2)
protegen al DNA del daño con H 2 0 2 . Los resultados fueron significativamente
diferentes en las células tratadas por media hora, y más evidentes en las de alta
concentración que en las de baja.
El factor de necrosis tumoral alfa (TNF) es una citocina producida por macrófagos
activados que al unirse a sus receptores de membrana p55 y p75 induce muerte por
apoptosis en diferentes células tumorales o inmortalizadas. En estudios previos del
laboratorio se observó que las células L929 (P6) empiezan a morir 12 h después de
haber sido expuestas al TNF, pero durante este periodo, el número de células no
aumenta como en las células control. Pensando que este bloqueo del ciclo celular
pudiera deberse a daño sobre el genoma, es por ello que en este trabajo también se
evaluó el daño que produce esta citocina a lo largo del tiempo, para después evaluar
la protección que confiere Bcl-2. Los resultados indican que se confirma el efecto
protector en las células c.2 tanto para los tratamientos con peróxido como con TNF.
daño a las 6 horas,
lo
cual se podría explicar por un efecto reparador en dichascélulas. Que también se observa en las c.2 pero no de manera tan evidente.
Sin embargo, hasta el momento no se conoce el mecanismo por el cual se genera
REFERENCIAS :
Christoph R., Jeen-Woo P., y Bruce N. A.(1988) ; Normal oxidative damage
to mitochondrial and nuclear DNA ; Proc." Natl. Acad. Sci. USA ; 85 : 6465-
6467.
Collins, A.R., Dobson, J.P. Dusinska, M. Kennedy, G . , Stetina, R.(1997) ; The
comet assay : what can it really tell us ; Mufat. Res ; 375 : 183-193.
David M . , Zoltan N , , Xiao-Ming, Curt L., and Stanley J.(1993) ; Bcl-2 functions
in an antioxidant pathway to prevent apoptosis ; Cell Press ; 75 :241-251.
Dimitry B.Z ; Boris F . K ; Alevtina E.K ; Michail Y.V. and Ljubava D.Z.
(1 997) ;Mitochondria revisted. Alternative functions of mitochondria ;
Bioscence Reports ; 1 7(6) :507-519
Harman, D . ( I 981 ); The aging proces ; Prdc Natl. Acad. Sci. USA ; 78 :7124-
71 28.
Halliwel B.(1984) ; Oxigen toxicity, oxigen radicals, transition metals and
dieses ; Biochem. J ; 21 9 : 1-1 4
Juliane M . , Shigemi M.(1998) ; Bcl-2 family proteins and mitochondria ; Bioch
et Bioph ; 1366 : 127-1 37.
Lenaz G. (1 998) ; Role of mitochondria in oxidative stress and agegin ; Bioch
et Bioph ; 1 336 :53-67.
Pang C.Y, Lee H.C, Yang J.H, We¡ Y.H,(1994) ; Human skin mitochondrial
DNA deletions associated with light exposure ; Arch Biochem Biophys ;
Wang, A.M., (1985) Science ; 228, 149-154.
Yau-He¡ W. (1998), Oxidative stres and mitochondrial DNA mutations in
OBJETIVOS Y METAS ALCANZADAS :
o Se evaluó el daño generado por estrés oxidativo sobre las células L929 que
sobreexpresan Bcl-2 y las que no lo expresan.
o Se encontraron las condiciones óptimas para evaluar el daño en el DNA de las
células tratadas con H 2 0 2 (concentración y tiempo de exposición).
o Se encontraron las condiciones óptimas para evaluar el daño en el DNA de las
células L929 tratadas con TNF (concentración y tiempo de exposición).
o Se sometieron a las células P6 (silvestres) y C.2 (que sobreexpresan Bcl-2) a
tratamiento con la concentración y tiempo de H202 y TNF establecidos previamente.
CRITERIOS DE EVALUACION
El presente proyecto de Servicio Social se culminó en un 100%. Se alcanzaron
todos los objetivos y metas planteadas. Inclusive se presentarán los resultados
obtenidos en el XXlll Congreso de la Sociedad Mexicana de Bioquímica a
realizarse del 19 al 24 de Noviembre del 2000 en Acapulco, Gro. (Se anexa
copia del resumen).
Sociedad Mexicana de
Bioquímica,
A.
C.
FUNDADA EN 1957 INSTITUTO DE FlSlOLOGlA CELULAR U.N.A.M. APARTADO POSTAL 70-600
C.P. 04510 MEXICO. D.F. TEL. FAX 56-22-56-11
TEL. 56-22-5643
MESA DIRECTIVA 1999-2001
’RESIDENTE
IR. EDMUNDO CHAVEZ COSSlO
/ICEPRESIDENTE
IR. HELIODORO CELE SANDOVAL
SECRETARIO TESORERO
IR. FEDERICO MARTINEZ MONTES
SUB-SECRETARIO TESORERO
3RA. GLORIA SOBERON CHAVEZ
México,
D.F.
a 10 de octubre del 2000OLIVIA HUCHIN
P r e s e n t e .
Estimado autor:
Tenemos el gusto de comunicarle que su resumen “Daño temprano al ADN inducido por el factor de necrrosis tumoral alfa en celulas L9L29” de Huchín
O.,
VenturaJ.L., Gómez E,O., Konigsberg M., Zentella A. fue aceptado para ser presentado en forma de poster con el número 442 en tamaño lm X l m durante el XXIII Congreso Nacional de la Sociedad Mexicana de Bioquímica que se celebrará del 19 al 24 de noviembre de 2000.
Atentamente,
DAÑO TEMPRANO AL ADN INDUCIDO POR EL FACTOR DE NECROSIS TUMORAL ALFA EN CELULAS L929
Huchín, O.n; Ventura, J.L.§; Gómez, E.O. §; Konigsberg, M.n; Zentella. A.
nDepartamento de Ciencias de la Salud, CBS, UAM- Iztapalapa. §Departamento de Biología Celular, Instituto de Fisiología Celular, UNAM Apartado Postal 70-243, Mexico
D.F.04510; FAX 56-22-56-1 1 , correo electrónico [email protected].
El factor de necrosis tumoral alfa (TNF) es una citocina producida por macrófagos activados que al unirse a sus receptores de membrana p55 y p75 induce muerte por